Introduction
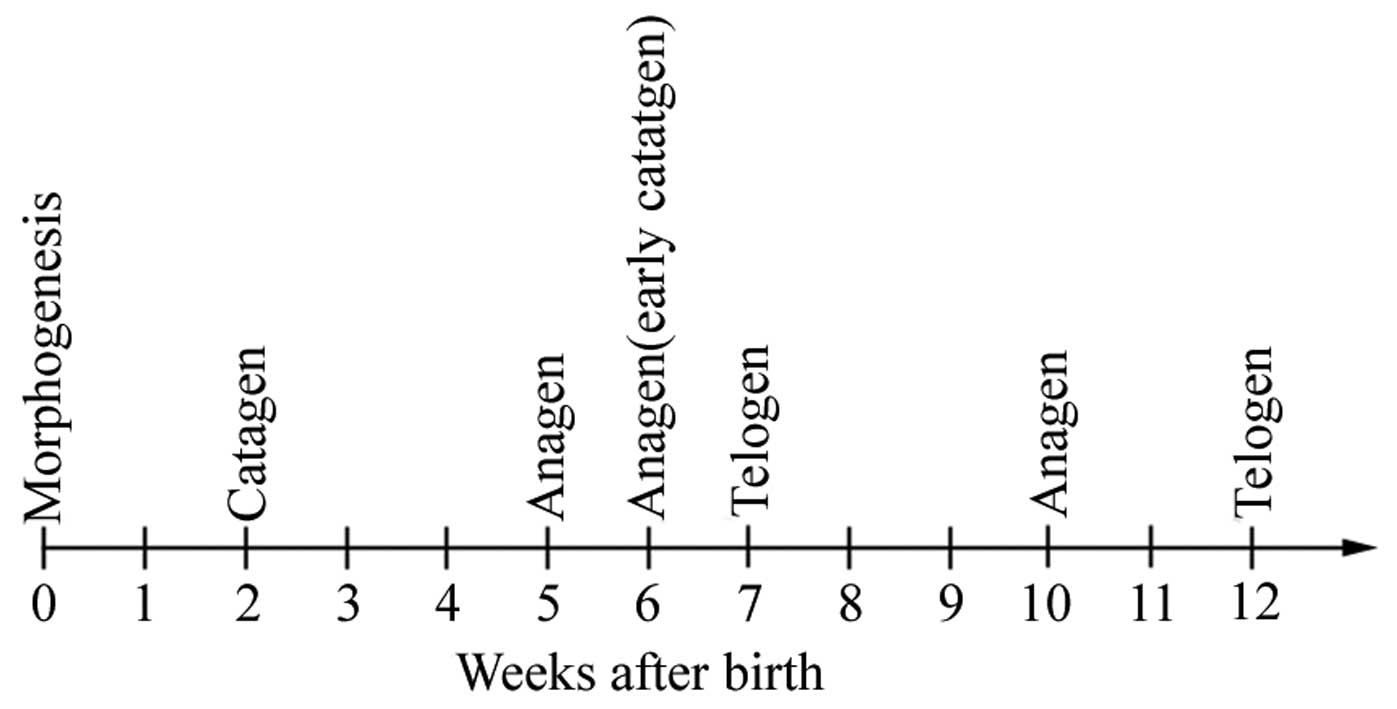
The hair cycle is an intrinsic and cyclic system of
regenerating tissue (1), which is
composed of the anagen (phases of rapid growth; 1–3 weeks), catagen
(phases of apoptosis-driven regression; ~2 days) and telogen
(phases of relative quiescence; ~2 weeks) phases (Fig. 1). The growth of hair is considered to
be a result of the cyclic remodeling of the follicle (2). There are numerous biological factors
that regulate or synchronize with the hair cycle (3), such as hormones, growth factors,
enzymes and transcriptional factors. Enzymes include ornithine
decarboxylase, aromatase and nexin 1. The hair cycle is considered
to be a process of tissue regeneration associated with
extracellular matrix (ECM) degradation and remodeling (4).
Matrix metalloproteinases (MMPs) are a family of
zinc-dependent endopeptidases that can be produced by a number of
different types of cells in the skin, including fibroblasts,
keratinocytes, macrophages, endothelial cells, mast cells and
eosinophils (5). MMPs consist of at
least 19 cloned membranes, including the type IV collagenases (such
as MMP-2 and MMP-9), metalloelastase, interstitial collagenase,
stromelysins, matrilysins and membrane-type MMPs. In addition,
increasing evidence demonstrates that MMPs can affect the signaling
pathways associated with various factors, such as insulin-like
growth factor-binding protein-3, tumor necrosis factor-α (TNF-α),
fibroblast growth factor receptor 1 and angiogenic factors
(6–10). MMP-2 and MMP-9 are important members
of the MMP superfamily, since they can degrade gelatin, collagen
IV, V, VII and X, fibronectin, and elastin (9–13).
The expression levels of MMP-2 and MMP-9 are
reported to be regulated by numerous factors. For instance, MMP-2
is regulated by transforming growth factor (TGF)-β, interleukin
(IL)-1β and interferon (IFN) in certain fibroblasts (14–17), and
Wnt in T cells (18). Similarly,
MMP-9 is induced by several regulators, including epidermal growth
factor, TGF-α, TGF-β, IL-1β and IFN-α (16,19–23). The
activities of MMP-2 and MMP-9 are specifically inhibited by tissue
inhibitor of matrix metalloproteinases (TIMP)-2 and TIMP-1,
respectively (24), which may be
vital in the regulation of connective tissue metabolism and
remodeling in numerous different organ systems, including hair
follicle.
There are a few studies investigating the
relationship between MMP-2, MMP-9, TIMP-1 and the hair cycle
(25). MMP-2, MMP-9 and TIMP-1 have
been suggested to be associated with the hair cycle in vitro
and in vivo (26). In
addition, the expression profiles of MMP-2, MMP-9, TIMP-1 and
TIMP-2 in the hair cycle remain unclear. Therefore, the present
study aimed to investigate the expression levels of MMP-2, MMP-9,
TIMP-1 and TIMP-2 throughout the hair cycle in mice.
Materials and methods
Materials
Rabbit polyclonal antibodies against MMP-2 (cat. no.
ab37150), MMP-9 (cat. no. ab38898), TIMP-1 (cat. no. ab61224) and
TIMP-2 (cat. no. ab180630) were purchased from Abcam (Cambridge,
MA, USA) and were all diluted to a concentration of 1:1,000. The
ELISA kits were purchased from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA; cat. nos. KHC3081, KHC3061, KHC1491 and
EHTIMP2).
Animals
Female C57BL/6 mice (n=5 at each week of age) aged
between 3 and 12 weeks (weight range, 9–23 g) were purchased from
the Laboratory Animal Center of Guangzhou University of Chinese
Medicine (Guangzhou, China). All animal experiments were performed
in strict accordance with the recommendations in the Guide for the
Care and Use of Laboratory Animals (2014). The mice had access to
food and water ad libitum, and were housed in a controlled
environment (temperature, 23±2°C; humidity, 55±10%; 12 h light/dark
cycle). To collect skin biopsies at each stage of the hair cycle, 6
mice at each week of age were used in this study. To ameliorate
animal welfare, the animals were used at a minimum number during
the study. As the mice reached their desired age, they were
sacrificed by CO2 gas inhalation. To obtain specific
hair cycle stage specimens, the dorsal skin of each mouse was
excised from just below the line connecting the scapulas. Each skin
specimen was separated from the subcutaneous tissue and was sampled
by punch biopsy for the examinations. Ethical approval was obtained
from the Ethics Committtee of Guangzhou University of Chinese
Medicine (approval no. 20141406-63).
Enzyme activity measurement
MMP-2 and MMP-9 activities were measured by 10%
SDS-PAGE gelatin zymography as previously described (24). Briefly, without boiling or reduction,
the samples (12 µl sample, 4 µl loading buffer) were fractionated
in an SDS-polyacrylamide gel containing 0.1% gelatin by
electrophoresis at 100 V for 90 min at 4°C. Next, the gels were
incubated in a digestion buffer (Shanghai Ann Inquiry Trading Co.,
Ltd., Shanghai, China) with bivalent metal ions to recover the
activities of MMP-2 and MMP-9. Subsequent to rinsing in distilled
water, the gels were stained with Coomassie brilliant blue (Beijing
Seitz Biotechnology Co., Ltd., Beijing, China), and then destained.
Gelatinolytic activity appeared as clear bands of digested gelatin
against a dark blue background of stained gelatin. The intensity of
the bands was measured using with Alpha Imager HP and AlphaView
software (version 1.0; ProteinSimple, San Jose, CA, USA), and was
proportional to the activities of MMP-2 and MMP-9.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA in the skin samples was isolated with
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The
RNA was quantified by measuring the absorption at 260 nm using
NanoDrop 2000 (Thermo Fisher Scientific, Inc.), and the purity of
the RNA was assayed by the ratio of absorbance at 260 nm and 280
nm. A fraction was electrophoresed with 1% agarose to verify its
integrity. The total RNA of each sample was then reverse
transcribed into cDNA with SuperScript III Reverse Transcriptase
(Thermo Fisher Scientific, Inc.). PCR was performed to measure the
mRNA levels of MMP-2, MMP-9, TIMP-1 and TIMP-2, using the primers
(synthesized by Invitrogen; Thermo Fisher Scientific, Inc.)
presented in Table I. In a 15 µl
system containing 7.5 µl 2X qPCR master mix, 2 µl primers and 5.5
µl ddH2O, GAPDH was used as a housekeeping gene and mRNA
expression was analyzed using an RT-PCR system (AB17500; Applied
Biosystems, Foster City, CA, USA) and the following thermal cycling
conditions: 40 cycles at 50°C for 2 min, 95°C for 2 min, 95°C for
15 sec and 60°C for 32 sec.
 | Table 1.Primer pair sequences used in reverse
transcription-polymerase chain reaction. |
Table 1.
Primer pair sequences used in reverse
transcription-polymerase chain reaction.
| Gene | Forward primer | Reverse primer |
|---|
| GAPDH |
5′-GGCCTCCAAGGAGTAAGAAA-3′ |
5′-GCCCCTCCTGTTATTATGG-3′ |
| MMP-2 |
5′-ATCGCAGACTCCTGGAATG-3′ |
5′-CCAGCCAGTCTGATTTGATG-3′ |
| MMP-9 |
5′-TCCAGTACCAAGACAAAGC-3′ |
5′-GAGCCCTAGTTCAAGGGCAC-3′ |
| TIMP-1 |
5′-AGTCCCAGAACCGCAGTGAA-3′ |
5′-AGAGTACGCCAGGGAACCAA-3′ |
| TIMP-2 |
5′-GGTAGCCTGTGAATGTTCCT-3′ |
5′-ACGAAAATGCCCTCAGAAG-3′ |
Enzyme-linked immunosorbent assay
(ELISA)
The expression levels of MMP-2, MMP-9, TIMP-1 and
TIMP-2 in the skin were detected using ELISA kits, according to the
manufacturer's instructions. Briefly, 100 µl prepared standard or
sample (1:40 dilution) was added to each well and incubated for 2 h
at room temperature. The plates were then washed with buffer four
times. Following washing, 100 µl biotinylated labeled primary
antibody was added to the wells, and the plates were incubated for
1 h at room temperature. After washing four times with buffer, the
plates were incubated with peroxidase-conjugated secondary antibody
for 45 min. The plates were washed three times, and 100 µl
Streptavidin-HRP Working Solution was added to each well (except
the chromogen blanks) to induce the color reaction at room
temperature for 30 min. The assay was stopped at 20–25 min. The
optical density at 450 nm was determined using an automated
microplate reader (ELx808) and Gen5 Data Analysis Software (version
2.0) (both purchased from BioTek, Shoreline, WA, USA).
Immunohistochemical analysis
The collected samples for immunohistochemical
examination were promptly fixed in 10% formalin solution. Following
dehydration and embedding in paraffin, skin sections of ~3 µm in
size were prepared, deparaffinized and incubated with 3%
H2O2 at room temperature for 5–10 min in
order to eliminate the endogenous peroxidase activity. For
histopathological examination, the sections were stained with
hematoxylin and eosin (Beijing Seitz Biotechnology Co., Ltd.). For
immunohistochemical examination, the sections were processed with
antigen repair and blocking, according to a previous study
(27,28). Briefly, slides in 0.01 M sodium
citrate buffer (Sigma-Aldrich, St. Louis, MO, USA), pH 6.0, were
incubated at 99–100°C for 20 min. Slides were then removed from
heat and kept in cool buffer at room temperature for 20 min and
rinsed in Tris-buffered saline with 0.1% Tween-20 (Aviva Systems
Biology Corporation, San Diego, CA, USA) at room temperature for 1
min. Following incubation overnight at 4°C with primary antibodies
against MMP-2, MMP-9, TIMP-1 and TIMP-2 separately, the samples
were exposed to horseradish peroxidase-conjugated anti-rabbit IgG
secondary antibody (1:5,000 dilution; cat. no. W4011; Promega
Corporation, Madison, WI, USA). Subsequently, DAB (Cell Signaling
Technology, Inc., Danvers, MA, USA) was used to visualize the
expression of proteins stained with hematoxylin and eosin.
Results
Fluctuation of MMP-2 and MMP-9
activities throughout the hair cycle
To investigate the correlation between MMP-2, MMP-9
and the hair cycle, the activities of MMP-2 and MMP-9 were
determined using samples of dorsal skin, including the hair
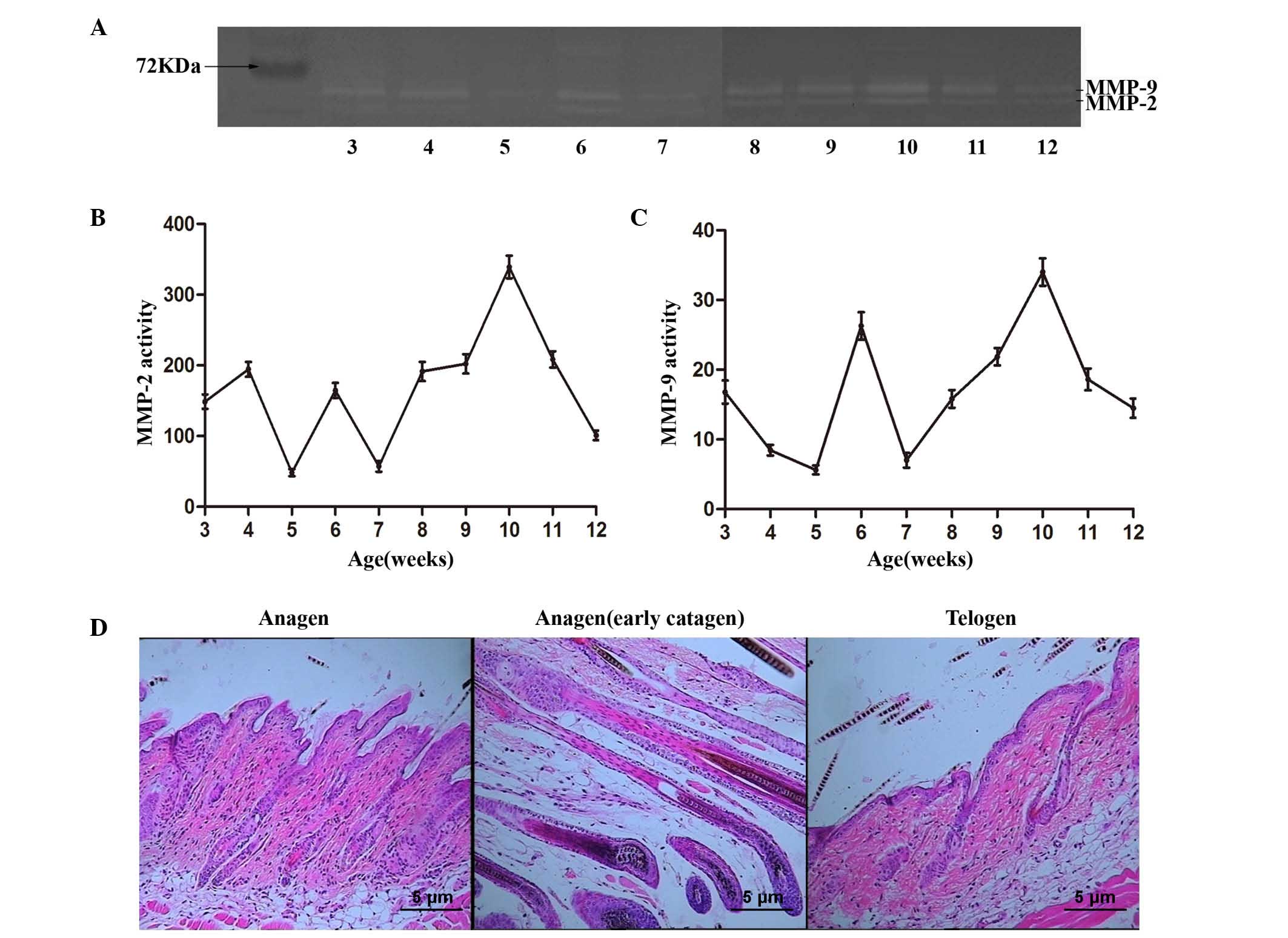
follicle. As presented in Fig. 2A,
the enzyme activities were present at each phase of the hair cycle.
The activity of MMP-2 fluctuated in the hair cycle; the activity
peaked at 6 and 10 weeks during the anagen phase, decreased during
the catagen phase, and reached its lowest level of activity at 7
and 12 weeks during the telogen phase of the hair cycle (Fig. 2B). Similarly, the activity of MMP-9
fluctuated, with a peak at 6 and 10 weeks (Fig. 2C). Typical histopathological images
of each stage during the hair cycle are presented in Fig. 2D. Cells in the cytoplasm during the
anagen phase were light or mild alkalophilic by eosin coloration,
and were eosinophilic with aging or degeneration.
Fluctuation of MMP-2, MMP-9, TIMP-1
and TIMP-2 mRNA and protein expression levels throughout the hair
cycle
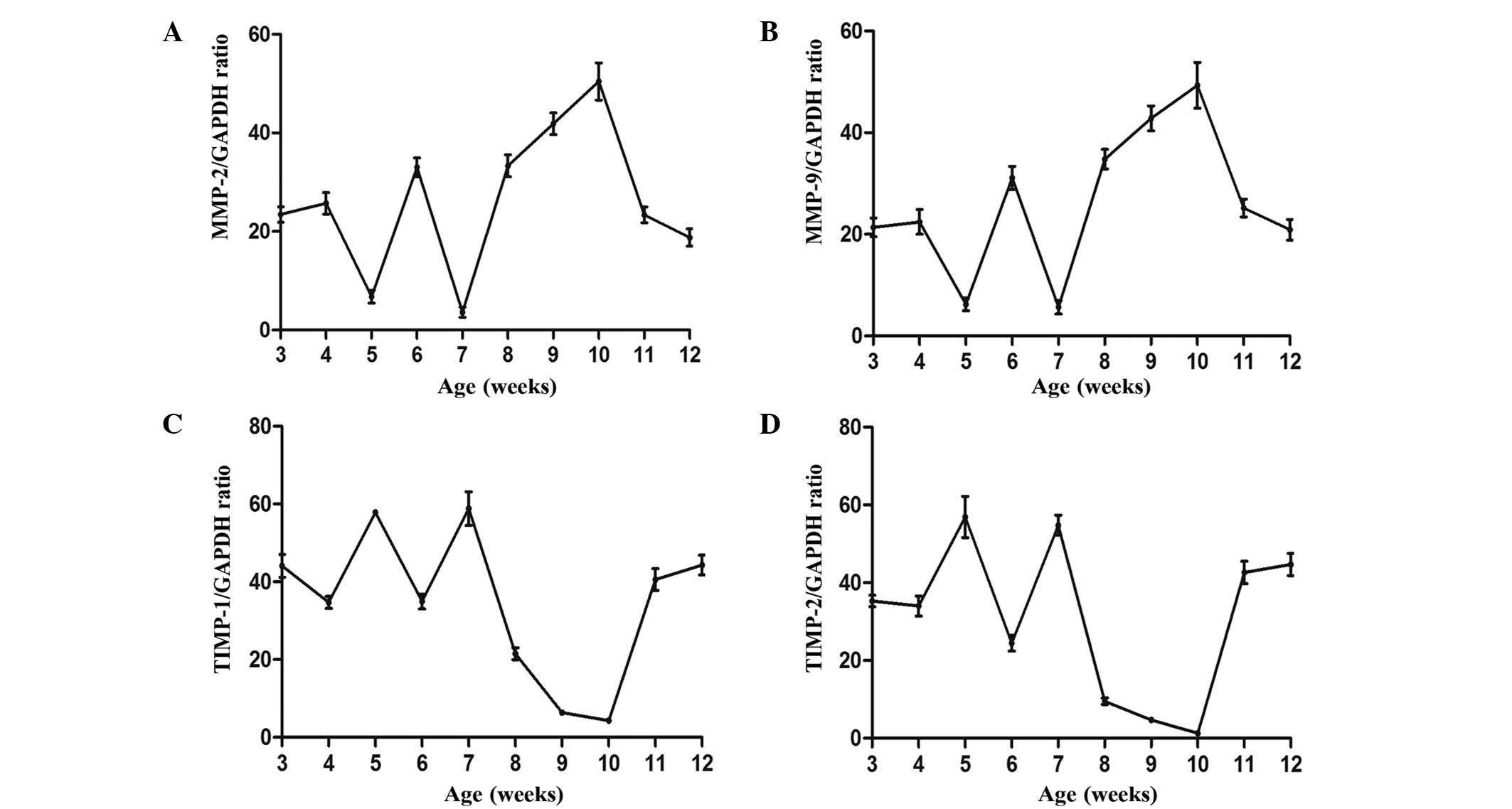
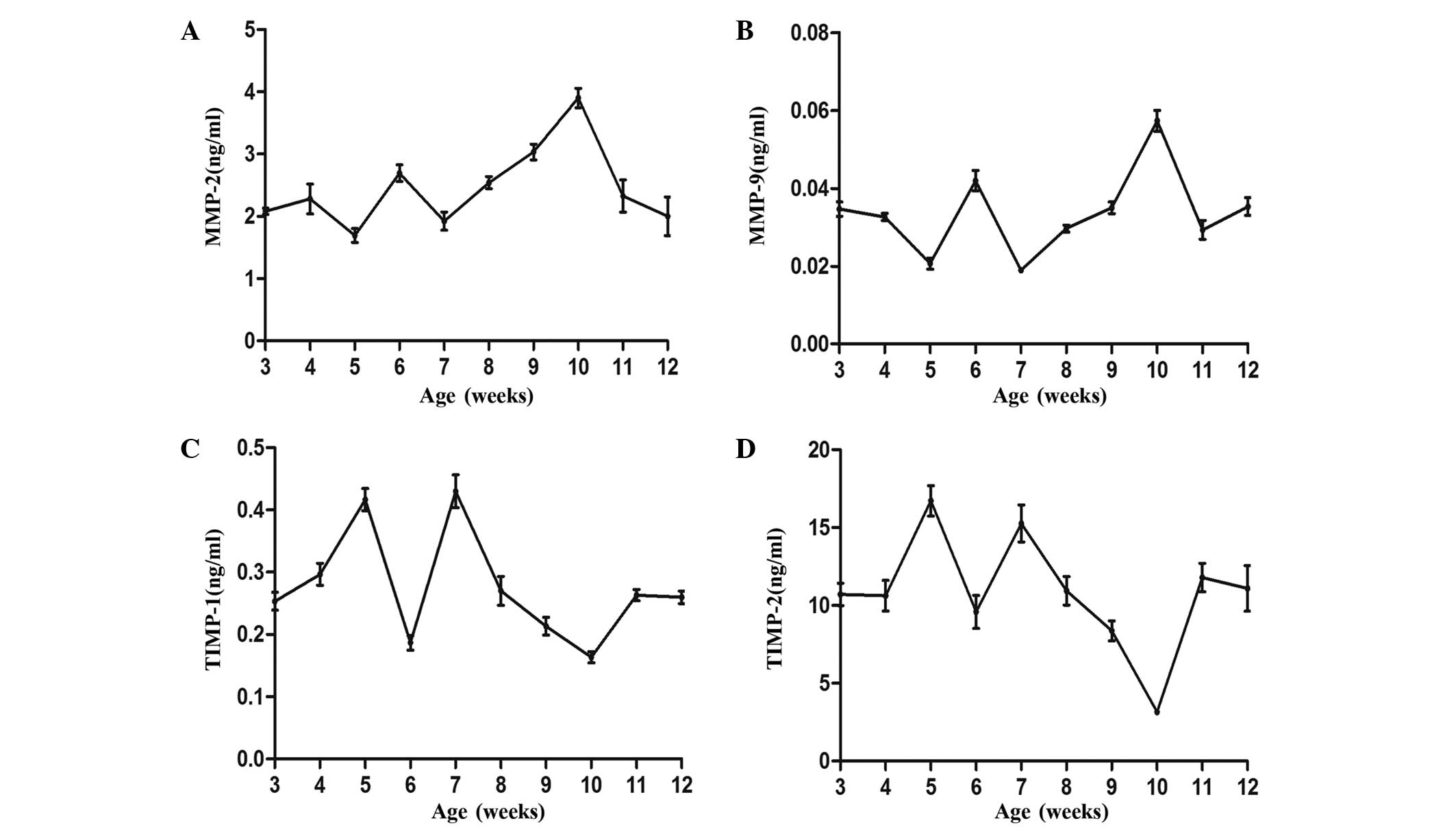
To determine whether the expression of the enzymes
of MMP-2 and MMP-9 fluctuated, RT-PCR and ELISA were performed in
order to determine the mRNA and protein expression levels,
respectively, of each protein. As shown in Figs. 3 and 4, the mRNA and protein expression levels of
the enzymes were similar and detectable throughout the hair cycle.
The expression of each protein fluctuated, with a peak at 5 and 10
weeks, and a decrease at 7 and 12 weeks, as is expected in the hair
cycle (29).
In addition, the expression levels of TIMP-1 and
TIMP-2 were determined for comparison against their targets, MMP-9
and MMP-2, respectively. Figs. 3 and
4 show the fluctuations in the mRNA
and protein expression levels of the TIMPs. mRNA and protein
expression levels were the lowest at 6 and 10 weeks, which
corresponded to the anagen phase, and were then increased during
the catagen phase, reaching peak levels at 7 and 12 weeks during
the telogen phase. These results indicate that the expression
levels of TIMP are negatively correlated with the MMP expression
levels.
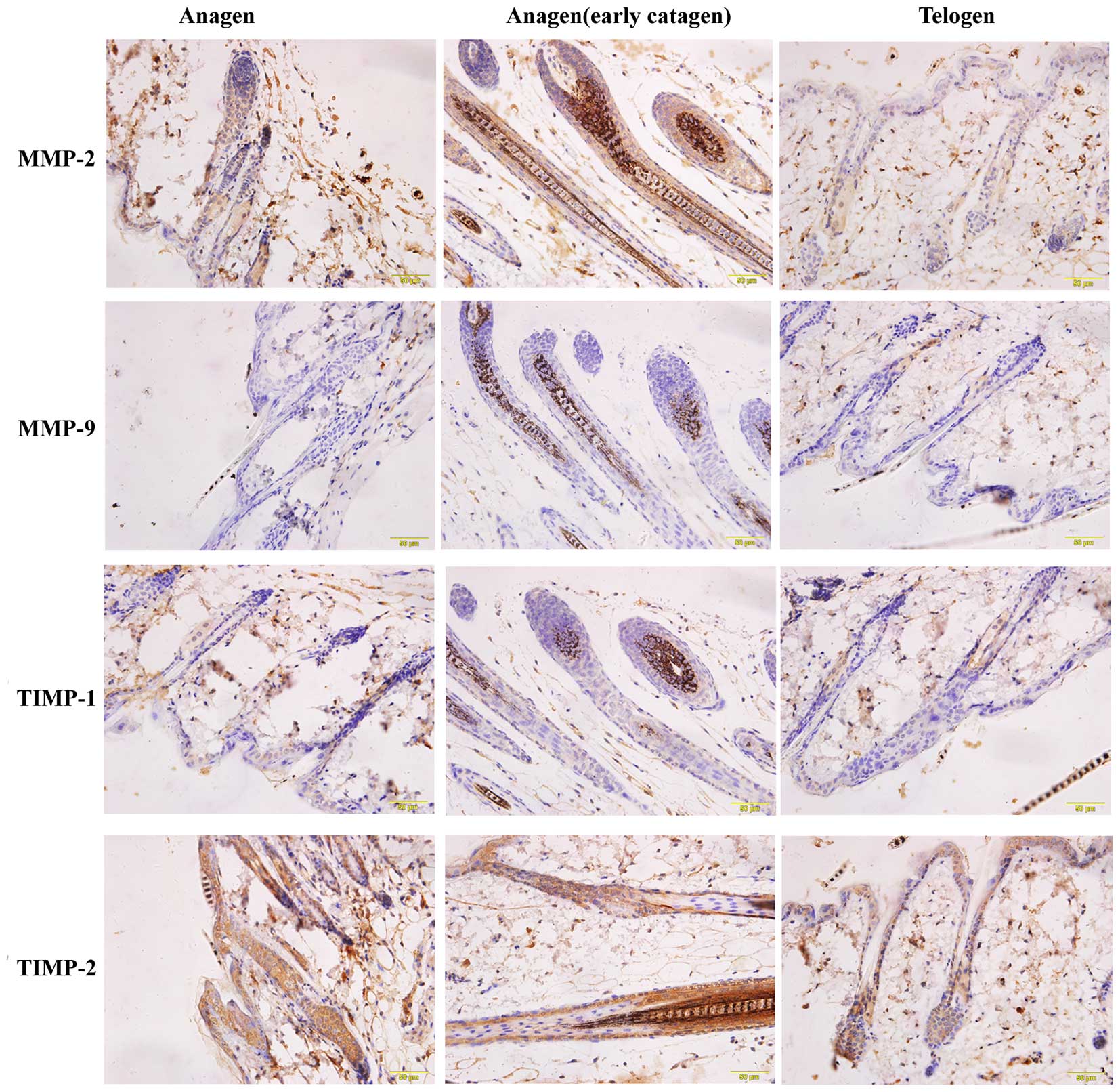
Localization of MMP-2, MMP-9, TIMP-1
and TIMP-2 throughout the hair cycle
To identify the expression sites of MMP-2, MMP-9,
TIMP-1 and TIMP-2 at each stage of the hair cycle,
immunohistochemical analysis was performed (Fig. 5). Positive staining for MMP-2 and
TIMP-2 was almost observed throughout the entire hair follicle at
each stage, although the expression was most intense during the
anagen (early catagen) phase. Positive staining was observed for
MMP-9 and TIMP-1 locally in the hair follicle, such as in the
sebaceous gland during the anagen and catagen phases, and in the
inner root sheath during the catagen phase.
Discussion
The mechanism underlying the hair cycle has been
widely studied, whereas the factors involved have yet to be
elucidated. MMPs and TIMPs are important regulators, participating
in proteolytic remodeling of ECM in various physiological pathways,
including tissue development, tissue repair and angiogenesis
(30). In the present study, the
expression levels of MMP-2 and MMP-9, typical ECM-degrading enzymes
and their inhibitors, TIMP-2 and TIMP-1, throughout the hair cycle
were investigated.
Bacterial infections of the hair follicle is
considered as primarily being caused by Staphylococcus
aureus. ECM is secreted by dermal papilla cells and involves a
cyclic process composed of degradation and renewal, which is
regulated by numerous factors, including growth factors, cytokines,
hormones, adhesion molecules and associated enzymes (13–16). It
has been reported that Wnt, TGF-β/bone morphogenic protein,
hepatocyte growth factor, and epidermal growth factor signaling
pathways serve as key regulators of hair follicle development
(31). As a downstream cytokine
regulated by the aforementioned signaling pathways, MMP-9 is an
important collagenase involved in ECM remodeling (32). Previous studies suggested that MMP-9
serves an important role in the hair cycle (25). In the present study, the expression
and activity of MMP-9 fluctuated, demonstrating peak activity
during the catagen phase and minimum activity during the telogen
phase in mice. This suggests that MMP-9 contributes towards the
hair cycle, which was also demonstrated in a previous study using
MMP-9−/− mice (26).
It has been hypothesized that other MMPs, such as
MMP-2, may compensate for the effects of MMP-9 on hair canal
formation in MMP-9−/− mice, including the changing of
the width of the hair canal and the dynamics of its formation
(26). In the current study, it was
demonstrated that the activity of MMP-2 fluctuated throughout the
hair cycle, similarly to MMP-9. However, the activity of MMP-2 was
much stronger compared with MMP-9, which is consistent with the
findings of a previous in vitro study (33). Using immunohistochemical analysis, it
was observed that in addition to the localization of MMP-9, MMP-2
was visualized at other localizations of the hair follicle at the
same stage. These results support the hypothesis that MMP-2
contributes to the hair cycle in vivo and may compensate for
the effects of MMP-9 on hair canal formation in MMP-9−/−
mice.
Divano et al (27) reported that the expression levels of
collagen fluctuate with regards to the hair cycle. The results
demonstrate that collagen expression levels increased suddenly and
significantly during the first 84 days of the telogen phase,
whereas it decreased in the last week of the phase (34). The results regarding the activities
of MMP-2 and MMP-9 in the present study may explain these data.
Following ECM degradation and remodeling, MMP-2 and MMP-9 may
affect the hair cycle by releasing growth factors from the ECM, and
by stimulating cell migration (35)
or activating growth factors, such as TGF-β (36). This strongly suggests that MMP-2 may
serve an important role in the hair growth cycle.
In the current study, TIMP-1 and TIMP-2 expression
levels were demonstrated to fluctuate throughout the hair growth
cycle. As expected, the expression levels of TIMP-1 and TIMP-2 were
negatively correlated to those of MMP-9 and MMP-2 respectively,
implying a negative correlation between MMPs and TIMPs. Similar to
previous immunostaining and histological studies, strong
correlations were observed between MMPs and TIMPs (37). However, the mRNA and protein
expression levels of TIMP-1 and TIMP-2 were higher when compared
with those of MMP-9 and MMP-2, respectively. In addition to
inhibition of MMPs, TIMPs are demonstrated to be multifunctional;
for instance, TIMP-2 is required for the activation of MMP-2, and
TIMP-1 has been shown to act as a growth factor (38). This suggests that the balance between
the expression levels of MMP-9 and TIMP-1, and of MMP-2 and TIMP-2
may functionally contribute towards the hair growth cycle.
In conclusion, the present study demonstrated that
the expression levels and activities of MMP-2 and MMP-9 fluctuate
throughout the hair growth cycle, and may be involved in its
regulation. However, the mechanism underlying the regulation of the
hair growth cycle by the enzymes requires further
investigation.
Acknowledgements
The present study was supported by the Natural
Science Foundation of China (grant nos. 31170949 and 81471900), the
Key Clinical Specialty Discipline Construction Program, the
Specialized Research Fund for the Doctoral Program of Higher
Education (grant no. 20124433110012), the Natural Science
Foundation of the Guangdong (grant no. S2012010009454) and the Dean
Fund (grant no. 2013Z013).
References
|
1
|
Werb Z and Chin JR: Extracellular matrix
remodeling during morphogenesis. Ann NY Acad Sci. 857:110–118.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stenn KS and Paus R: Controls of hair
follicle cycling. Physiol Rev. 81:449–494. 2001.PubMed/NCBI
|
|
3
|
Morisaki N, Ohuchi A and Moriwaki S: The
role of neprilysin in regulating the hair cycle. PLoS One.
8:e559472013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu P, Takai K, Weaver VM and Werb Z:
Extracellular matrix degradation and remodeling in development and
disease. Cold Spring Harb Perspecti Biol. 3:a0050582011.
|
|
5
|
Kähäii VM and Saariallio-Kere U: Matrix
metalloproteinases in skin. Exp Dermatol. 6:199–213. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fowlkes JL, Enghild JJ, Suzuki K and
Nagase H: Matrix metalloproteinases degrade insulin-like growth
factor-binding proein-3 in dermal fibroblast cultures. J Biol Chem.
269:25742–25746. 1994.PubMed/NCBI
|
|
7
|
Björnberg F, Lantz M, Olsson I and
Gullberg U: Mechanisms involved in the processing of the p55 and
the p75 tumor necrosis factor (TNF) receptors to soluble receptor
forms. Lymphokine Cytokine Res. 13:203–211. 1994.PubMed/NCBI
|
|
8
|
Levi E, Fridman R, Miao HQ, Ma YS, Yayon A
and Vlo-davsky I: Matrix metalloproteinase 2 releases active
soluble ectodomain of fibroblast growth factor receptor 1. Proc
Natl Acad Sci USA. 93:7069–7074. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Patterson BC and Sang QA:
Angiostatin-converting en-zyme activities of human matrilysin
(MMP-7) and gelatinase B/type IV collagenase (MMP-9). J Biol Chem.
272:28823–28825. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brooks PC, Silletti S, von Schalscha TL,
Friedlander M and Cheresh DA: Disruption of angiogenesis by PEX, a
noncata-lytic metalloproteinase fragment with integrin binding
activity. Cell. 92:391–400. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Woessner JF Jr: Matrix metalloproteinases
and their inhibitors in connective tissue remodeling. FASEB J.
5:2145–2154. 1991.PubMed/NCBI
|
|
12
|
Falkenburg JH, Harrington MA, de Paus RA,
Walsh WK, Daub R, Landegent JE and Broxmeyer HE: Differential
transcriptional and posttranscriptional regulation of gene
expression of the colony-stimulating factors by interleukin-1 and
fetal bovine serum in murine fibroblasts. Blood. 78:658–665.
1991.PubMed/NCBI
|
|
13
|
Brown PD, Levy AT, Margulies I, Liotta LA
and Stetler-Stevenson WG: Independent expression and cellular
processing of Mr 72,000 type IV collagenase and interstitial
collagenase in human tumorigenic cell lines. Cancer Res.
50:6184–6191. 1990.PubMed/NCBI
|
|
14
|
Hujanen ES, Väisänen A, Zheng A,
Tryggvason K and Turpeenniemi Hujanen T: Modulation of M(r) 72,000
and M(r) 92,000 type-IV collagenase (gelatinase A and B) gene
expression by interferons a and gamma in human melanoma. Int J
Cancer. 58:582–586. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Singh RK, Gutman M, Reich R and Bar-Eli M:
Ultraviolet B irradiation promotes tumorigenic and metastatic
properties in primary cutaneous melanoma via induction of
interleukin 8. Cancer Res. 55:3669–3674. 1995.PubMed/NCBI
|
|
16
|
Wu B, Crampton SP and Hughes CC: Wnt
signaling induces matrix metalloproteinase expression and regulates
T cell transmigration. Immunity. 26:227–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Okada Y, Tsuchiya H, Shimizu H, Tomita K,
Nakanishi I, Sato H, Seiki M, Yamashita K and Hayakawa T: Induction
and stimulation of 92-kDa gelatinase/type IV collagenase production
in osteosarcoma and fibrosarcoma cell lines by tumor necrosis
factor alpha. Biochem Biophys Res Commun. 171:610–617. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Welch DR, Fabra A and Nakajima M:
Transforming growth factorb stimulates mammary adenocarcinoma cell
invasion and metastatic potential. Proc Natl Acad Sci USA.
87:7678–7682. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Samuel SK, Hurta RA, Kondaiah P, Khalil N,
Turley EA, Wright JA and Greenberg AH: Autocrine induction of tumor
protease production and invasion by a metallothionein-regulated
TGF-beta 1 (Ser223, 225). EMBO J. 11:1599–1605. 1992.PubMed/NCBI
|
|
20
|
Lyons RM, Gentry LE, Purchio A and Moses
HL: Mechanism of activation of latent recombinant transforming
growth factor beta 1 by plasmin. J Cell Biol. 110:1361–1367. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kondapaka SB, Fridman R and Reddy KB:
Epidermal growth factor and amphiregulin up-regulate matrix
metalloprotein-ase-9 (MMP-9) in human breast cancer cells. Int J
Cancer. 70:722–726. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nagase H: Activation mechanisms of matrix
metalloproteinases. Biol Chem. 378:151–160. 1997.PubMed/NCBI
|
|
23
|
Hou C, Miao Y, Wang J, Wang X, Chen CY and
Hu ZQ: Collagenase IV plays an important role in regulating hair
cycle by inducing VEGF, IGF-1, and TGF-β expression. Drug Des Devel
Ther. 9:5373–5383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sharov AA, Schroeder M, Sharova TY,
Mardaryev AN, Peters EM, Tobin DJ and Botchkarev VA: Matrix
metalloproteinase-9 is involved in regulation of the hair canal
formation. J Invest Dermatol. 131:257–260. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Larson B, Banks P, Sherman H and
Rothenberg M: Automation of cell-based drug absorption assays in
96-well format using permeable support systems. J Lab Autom.
17:222–232. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen Y, Wei X, Guo C, Jin H, Han Z, Han Y,
Qiao T, Wu K and Fan D: Runx3 suppresses gastric cancer metastasis
through inactivation of MMP9 by upregulation of TIMP-1. Int J
Cancer. 129:1586–1598. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nyhlin N, el-Salhy M, Sandstrom O and Suhr
O: Evaluation of immunohistochemical staining of human duodenal
endocrine cells after microwave antigen retrieval. Histochem J.
29:177–181. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Warford A, Akbar H and Riberio D: Antigen
retrieval, blocking, detection and visualisation systems in
immunohistochemistry: A review and practical evaluation of tyramide
and rolling circle amplification systems. Methods. 70:28–33. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tasseff R, Bheda-Malge A, DiColandrea T,
Bascom CC, Isfort RJ and Gelinas R: Mouse hair cycle expression
dynamics modeled as coupled mesenchymal and epithelial oscillators.
PLoS Comput Biol. 10:e10039142014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Patruno A, Pesce M, Marrone A, Speranza L,
Grilli A, De Lutiis MA, Felaco M and Reale M: Activity of matrix
metallo proteinases (MMPs) and the tissue inhibitor of MMP (TIMP)-1
in electromagnetic field-exposed THP-1 cells. J Cell Physiol.
227:2767–2774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schmidt-Ullrich R and Paus R: Molecular
principles of hair follicle induction and morphogenesis. Bioessays.
27:247–261. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gordon GM, Ledee DR, Feuer WJ and Fini ME:
Cytokines and signaling pathways regulating matrix
metalloproteinase-9 (MMP-9) expression in corneal epithelial cells.
J Cell Physiol. 221:402–411. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jarrousse F, Boisnic S, Branchet MC,
Beranger JY, Godeau G, Breton L, Bernard BA and Mahé YF:
Identication of clustered cells in human hair follicle responsible
for MMP-9 gelatinolytic activity: Consequences for the regulation
of hair growth. Int J Dermatol. 40:385–392. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Divano C, Cardo P, Cipriani C and
Pellerano S: Soluble and insoluble collagen and elastin in the rat
hair cycle. Arch Dermatol Res. 266:135–141. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yamazaki M, Tsuboi R, Lee YR, Ishidoh K,
Mitsui S and Ogawa H: Hair cycle-dependent expression of hepatocyte
growth factor (HGF) activator, other proteinases and proteinase
inhibitors correlates with the expression of HGF in rat hair
follicles. J Investig Dermatol Symp Proc. 4:312–315. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang M, Zhao D, Spinetti G, Zhang J, Jiang
LQ, Pintus G, Monticone R and Lakatta EG: Matrix metalloproteinase
2 activation of transforming growth factor-beta1 (TGF-beta1) and
TGF-beta1-type II receptor signaling within the aged arterial wall.
Arterioscler Thromb Vasc Biol. 26:1503–1509. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sugita H, Osaka S, Toriyama M, Osaka E,
Yoshida Y, Ryu J, Sano M, Sugitani M and Nemoto N: Correlation
between the histological grade of chondrosarcoma and the expression
of MMPs, ADAMTSs and TIMPs. Anticancer research. 24:4079–4084.
2004.PubMed/NCBI
|
|
38
|
Yang HK, Jeong KC, Kim YK and Jung ST:
Role of matrix metalloproteinase (MMP) 2 and MMP-9 in soft tissue
sarcoma. Clin Orthop Surg. 6:443–454. 2014. View Article : Google Scholar : PubMed/NCBI
|