Introduction
Mesenchymal stem cell (MSC) research is a key topic
in current stem cell research. Multiple previous studies have
demonstrated that MSCs have therapeutic effects on acute kidney
injury (AKI), immunoglobulin A nephropathy and other renal diseases
(1–3). AKI is recognized as a common disease
which is expensive to manage, prolongs hospitalization and is
associated with increased mortality. The mortality rate for AKI has
been reported at 24% in hospital patients, and increases with the
severity of AKI (4). Clinically,
novel and more effective strategies for the treatment of AKI are
expected. MSCs have been applied and provided satisfactory
therapeutic effects for AKI in animals and humans (2,3,5); there are multiple mechanisms involved
in the therapeutic actions of MSC in AKI treatment (1).
The hypothesis that MSCs may transform to renal
tubular epithelial cells remains controversial. The discrepancy in
results is likely to be associated with the types of stem cell,
disease models and labeling and detection methods used (1,5,6). The first MSCs, and the most widely
used, are bone marrow-derived MSCs (BMSCs) (6). However, other sources of MSCs have also
emerged in clinical and research settings of renal regenerative
therapy and chronic kidney disease, including adult and fetal
tissue and umbilical cord blood (7,8).
However, previous studies on MSCs sourced from human early embryos
are limited. Compared with MSCs derived from adult tissues, a
number of features of embryonic stem cells differ, including
biological activities, such as cytokine expression and cell
adhesion molecules (9,10), differentiation potential, in
vivo migration and proliferation of embryo stem cells are
markedly increased compared with adult stem cells, and the
immunogenicity of embryonic stem cells is reduced (11–15).
Embryo-derived MSCs can be frozen in the laboratory and amplified
immediately to meet the requirement for treatment. However,
alternative sources of embryonic stem cells would be beneficial for
research and therapeutic purposes (16). Previous studies on MSCs sourced from
human early embryos are limited.
The current study hypothesized that MSCs derived
from human early embryos have an improved capability to
differentiate into tubular cells. The aim of the present study was
therefore to determine whether human embryonic MSCs (hMSCs) are
able to transform into renal tubular cells in the kidneys of
newborn mice.
Materials and methods
Cell culture and labeling with
PKH26
hMSCs were obtained from human embryos aged 4–7
weeks old, provided by Dr Minjuan Wu (Research Center of
Developmental Biology and Department of Histology and Embryology,
Second Military Medical University, Shanghai, China). The human
embryos were obtained from voluntary terminations of pregnancy with
RU486 anti-progesterone compound (17). The Committee on Ethics of Biomedicine
Research (Second Military Medical University, Shanghai, China)
reviewed and approved all human research protocols, and all donors
provided written informed consent. The hMSCs were grown as
described previously (8) and stored
at the Department of Histology and Embryology of the Second
Military Medical University (Shanghai, China). The hMSCs were
maintained in Dulbecco's minimal essential medium (DMEM;
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% fetal calf serum at 37°C in an atmosphere of
5% carbon dioxide.
As a type of lipophilic dye and emission of red
fluorescence, PKH26 can be combined with cell membranes
irreversibly and conduct fluorescence labeling for numerous types
of cells. hMSCs at passage 4 were labeled with the red fluorescent
dye PKH26 (Sigma-Aldrich, St. Louis, MO, USA) according to the
manufacturer's protocol. Briefly, the 80–90% confluence hMSCs were
trypsinized by 0.25% Trypsin solutions (Invitrogen; Thermo Fisher
Scientific, Inc.), washed using serum-free DMEM and resuspended in
1 ml of Diluent C from the PKH26 Red Fluorescent Cell Linker kit
(cat. no. PKH26-GL; Sigma-Aldrich). The cell suspension was mixed
with an equal volume of the labeling solution (containing 4 nM
PKH26; final concentration, 4 nM PKH26) and incubated at 25°C for 5
min. The staining reaction was stopped by the addition of 2 ml
fetal bovine serum, cells were washed 3 times with DMEM and
observed using epifluorescence microscopy.
In vitro counting
PKH26-positive hMSCs were observed by fluorescence
microscopy at five different time points following addition of the
PKH-26 label (24 h, 1 week, 2 weeks, 3 weeks and 4 weeks). A total
of 5 fields of view (magnification, ×400) were selected for every
time point. Red fluorescent cells were counted in each field of
view, and the labeling rate was calculated as: The number of
PKH26-positive cells/total number of cells.
Proliferation
Cell growth curves were drawn to compare the
proliferation between hMSCs and PKH26-labeled hMSCs. The cells were
grown on 24-well plates at a density of 100 cells/cm2.
Every 24 h, the cell number in 4 randomly-selected wells was
counted, and mean values were calculated using a hemocytometer
counting chamber. Cell growth curves of hMSCs and PKH26-labeled
hMSCs from days 1–7 were then calculated from these values, as a
function of incubation time.
Fluorescence activated cell sorting
(FACS) analysis
Surface markers of the hMSCs were analyzed by FACS.
The following monoclonal antibodies were used: Fluorescein
isothiocyanate (FITC)-conjugated anti-CD90 (cat. no. 328108),
anti-CD34 (cat. no. 343604) and anti-CD45 (cat. no. 368508) and
phycoerythrin-conjugated anti-CD29 (cat. no. 303004) (Biolegend).
The analysis was performed by a FACSCalibur cytometer (BD
Biosciences, Franklin Lakes, NJ, USA). hMSCs were stained with
antibody (1:100) and incubated at 4°C for 30 min. At least 10 cell
samples were acquired for each analysis.
Apoptosis was assessed by FITC-annexin V and
propidium iodide staining (Annexin V-FITC Apoptosis Detection kit,
Bipec Biopharma, Cambridge, MA, USA) according to the
manufacturer's instructions. Cells were analyzed by FACS at 594-nm
excitation (green fluorescence) for annexin-stained cells, and
excitation (red) for cells stained with propidium iodide.
Chondrogenic and adipogenic
differentiation
The hMSCs and PKH26-hMSCs were seeded onto 6-well
plates, and differentiation of these cells into chondrocytes and
adipocytes was induced at 40–50% confluence. To induce chondrocyte
differentiation, cells were cultured with chondrogenic
differentiation medium comprising 1.5×10−4 mg/ml
ascorbic acid and 1 ng/ml human recombinant transforming growth
factor-β (Sigma-Aldrich). Immunohistochemistry was performed to
examine the presence of types II collagen. Cell-seeded constructs
were rinsed with 1X phosphate-buffered saline, fixed in 4% formalin
for 24 h and embedded in paraffin. Slides were incubated with 10%
normal goat serum to block non-specific sites and mouse monoclonal
collagen type II (1:100; 1 mg/ml; cat. no. ab3092; Abcam,
Cambridge, UK) primary antibodies were applied for 1 h at 22°C.
Secondary antibody (anti-mouse immunoglobulin G biotin conjugate;
1:200; 2.1 mg/ml; cat. no. B7151; Sigma-Aldrich) was added for 1 h
followed by incubation with ABC reagent (Vectastain PK-400; Vector
Laboratories, Inc., Peterborough, UK) for 45 min. To induce
adipocyte differentiation, cells were cultured with adipogenic
differentiation medium comprising 1×10−8 mol/l
dexamethasone and 1×10−10 mol/l insulin (Sigma-Aldrich)
(8). Two weeks after
differentiation, adipocytes were identified by the existence of
lipid vesicles, using staining with Oil Red O (Sigma-Aldrich).
In vivo experiments
This study conformed to the Guide for the Care and
Use of Laboratory Animals (18).
Pregnant Kunming mice were purchased from the Shanghai Laboratory
Experimental Animal Center of the Second Military Medical
University (Shanghai, China). Ethical approval was provided by the
Ethics Committee of Biomedicine Research (Second Military Medical
University). All mice were allowed free access to standard
laboratory chow and tap water, housed in a room with constant
temperature (22°C) and a 12/12-h light/dark cycle.
Kunming mice (n=6; weight, 2.01 ± 0.18 g) were
narcotized by ether for 2 min, 2 days post-birth for 2 min. A
microsyringe was used to slowly inject 20 µl cell suspension in
phosphate-buffered saline containing 4×103 PKH26-hMSCs
into the left kidney at a constant rate. The uninjected right
kidney served as a control.
Renal morphology
Within 4 weeks of PKH26-hMSC transplantation, frozen
sections were extracted from kidney tissues weekly. Successive
frozen sections (thickness, 5 µm) were made following Optimal
Cutting Temperature (OCT) compound embedding. Subsequent to drying
for 30 min at room temperature, the sections were fixed with cold
acetone for 10 min. An Olympus IX70 fluorescence microscope
(Olympus Corporation, Tokyo, Japan) and a laser scanning confocal
microscope (Leica Microsystems, Inc., Buffalo Grove, IL, USA) were
used to observe and analyze the sections.
The localization of PKH26-labeled hMSCs in the
kidneys was observed 2 weeks after the injection of cells. Samples
were frozen immediately in liquid nitrogen, embedded in OCT
compound, sliced to 5 µm sections, fixed in acetone for 10 min, and
incubated for 30 min at 22°C with FITC-labeled wheat germ
agglutinin (WGA; Vector Laboratories, Inc.). Nuclei were stained
with 4,6-diamidino-2-phenylindole dihydrochloride (Sigma-Aldrich).
PKH26-positive cells were counted in 10 frozen renal sections per
mouse (n=3 mice). The number of stem cells in kidney tubules was
also counted.
Statistical analysis
Stata version 14.0 (StataCorp LP, College Station,
TX, USA) was used to perform all statistical analyses. Data are
presented as the mean ± standard error of the mean, and P<0.05
was considered to represent a statistically significant difference.
For comparison between unpaired groups, Student's t-test was
employed.
Results
Morphological observation and
apoptosis of hMSCs
In primary culture, cell adherence occurred within
12 h after plating and culture medium was changed after 24 h.
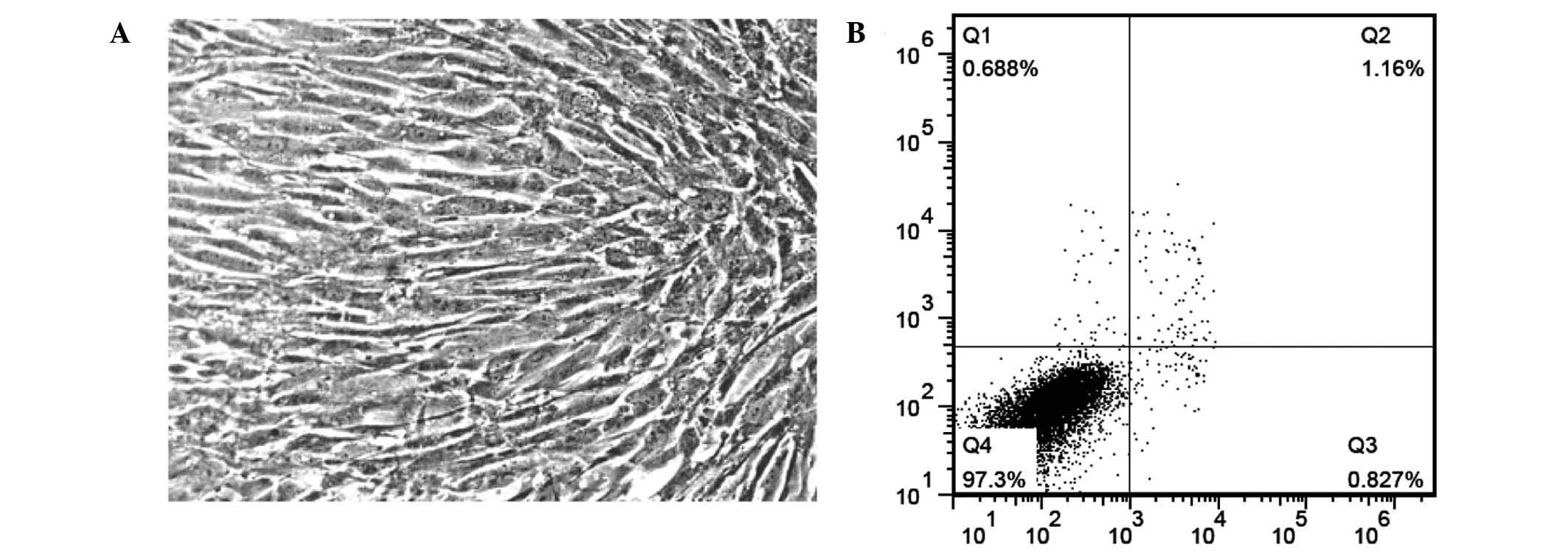
Cellular morphology was different to the primary culture. All the
stem cells were fibroblast-like at passages 4–5 (Fig. 1A). Fewer than 1% of cells at passage
5 were undergoing apoptosis when analyzed by flow cytometry
(Fig. 1B).
In vitro observation of hMSCs
following labeling with PKH26
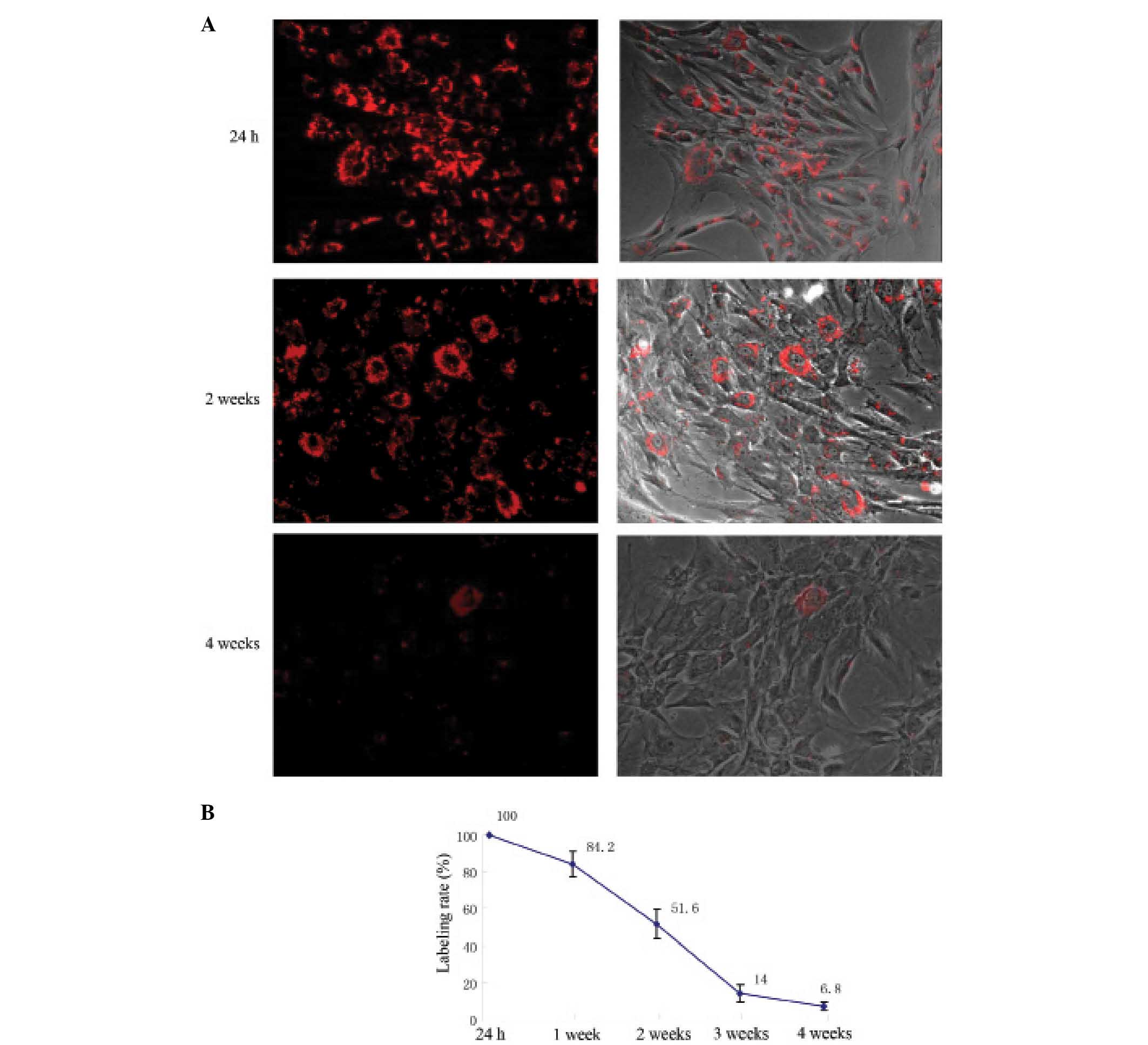
PKH26 was evenly distributed on the hMSC cell
membrane 24 h after PKH26 labeling, presenting as red fluorescence
under microscopy (Fig. 2A). The
PKH26 labeling rate was 98.6 ± 1.83% (Fig. 2B). With increased cell culture time,
fluorescence uniformity on the cell membranes decreased, the
intensity of labeling progressively weakened, and the labeling rate
continuously declined (Fig. 2). Two
weeks after PKH26 labeling, the red fluorescence was apparent in
half of the hMSCs. Four weeks after labeling, the red fluorescence
remained in ~7% of hMSCs (Fig.
2).
PKH26 labeling has no effect on hMSC
proliferation
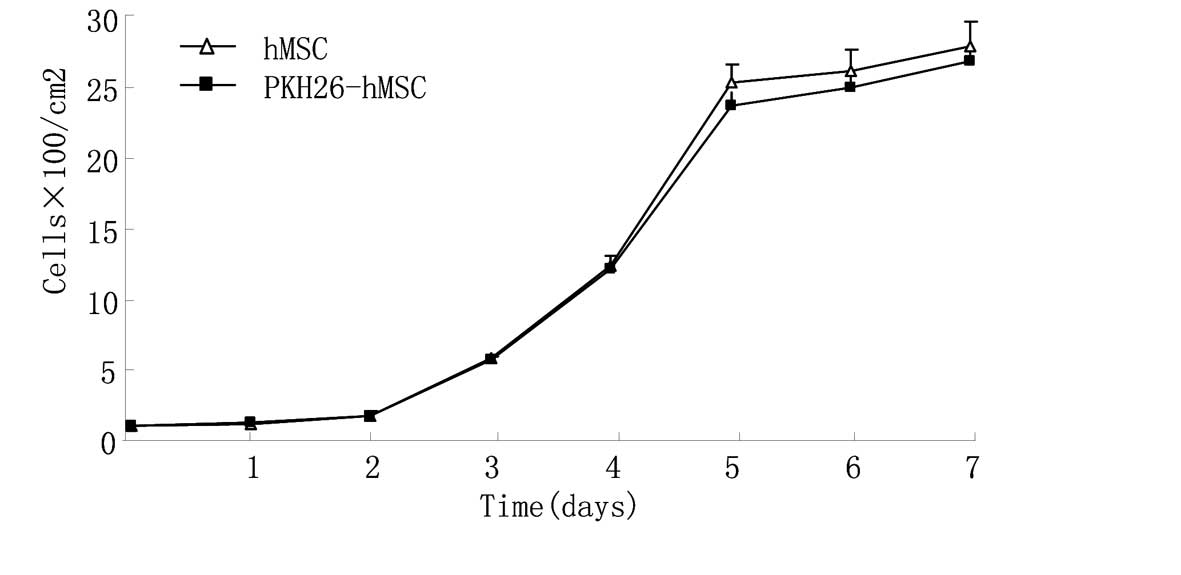
No statistically significant difference was observed
between the growth curves of hMSCs and PKH26-hMSCs (P<0.05),
which indicated that PKH26 label had no marked effect on hMSC
proliferation (Fig. 3).
PKH26 labeling does not affect the
expression of stem cell markers on hMSCs
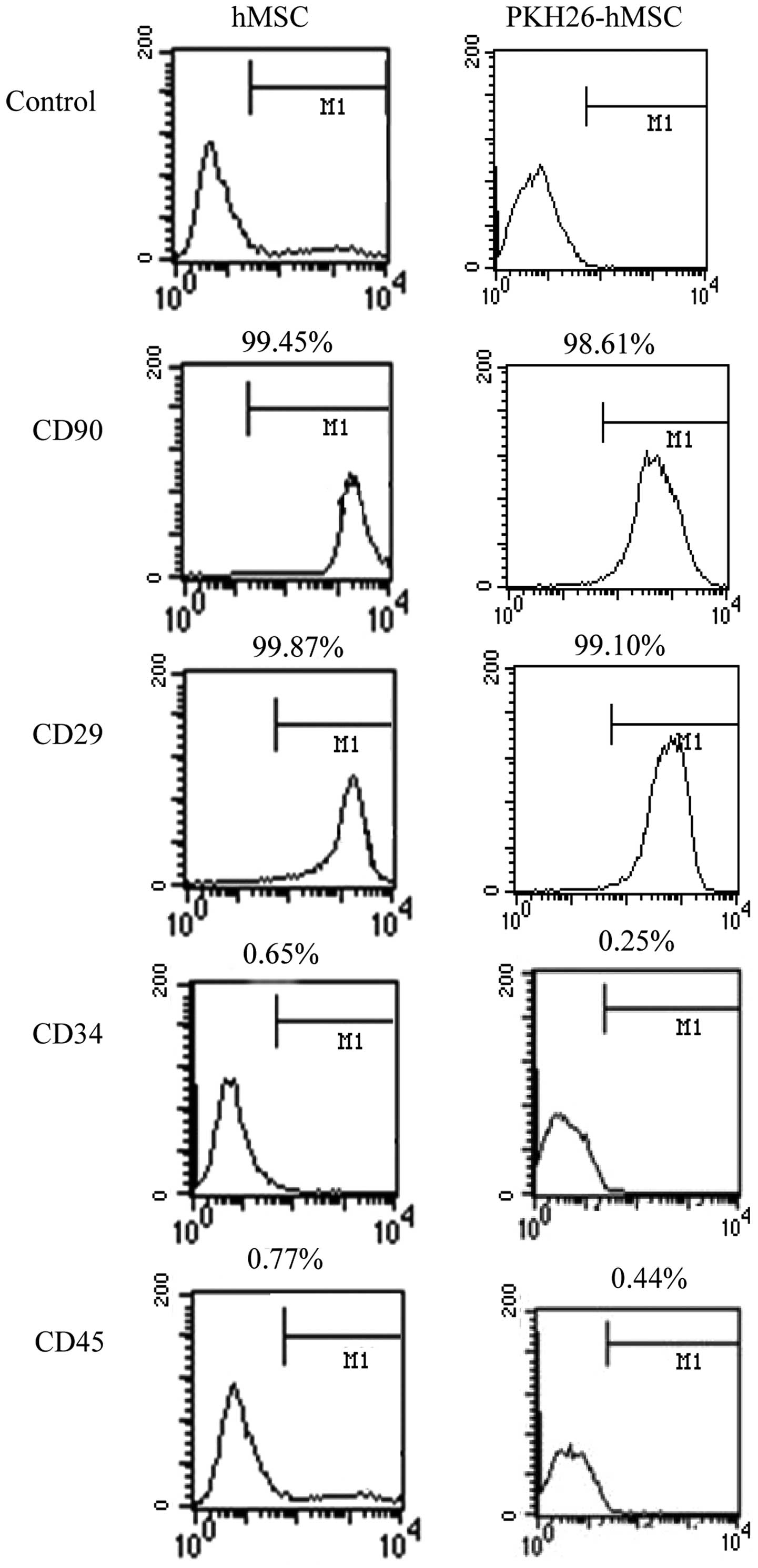
Flow cytometric analysis revealed that hMSCs and
PKH26-hMSCs were positive for the mesenchymal markers CD29 and
CD90, but negative for the markers of hematopoietic lineages CD34
and CD45 (Fig. 4).
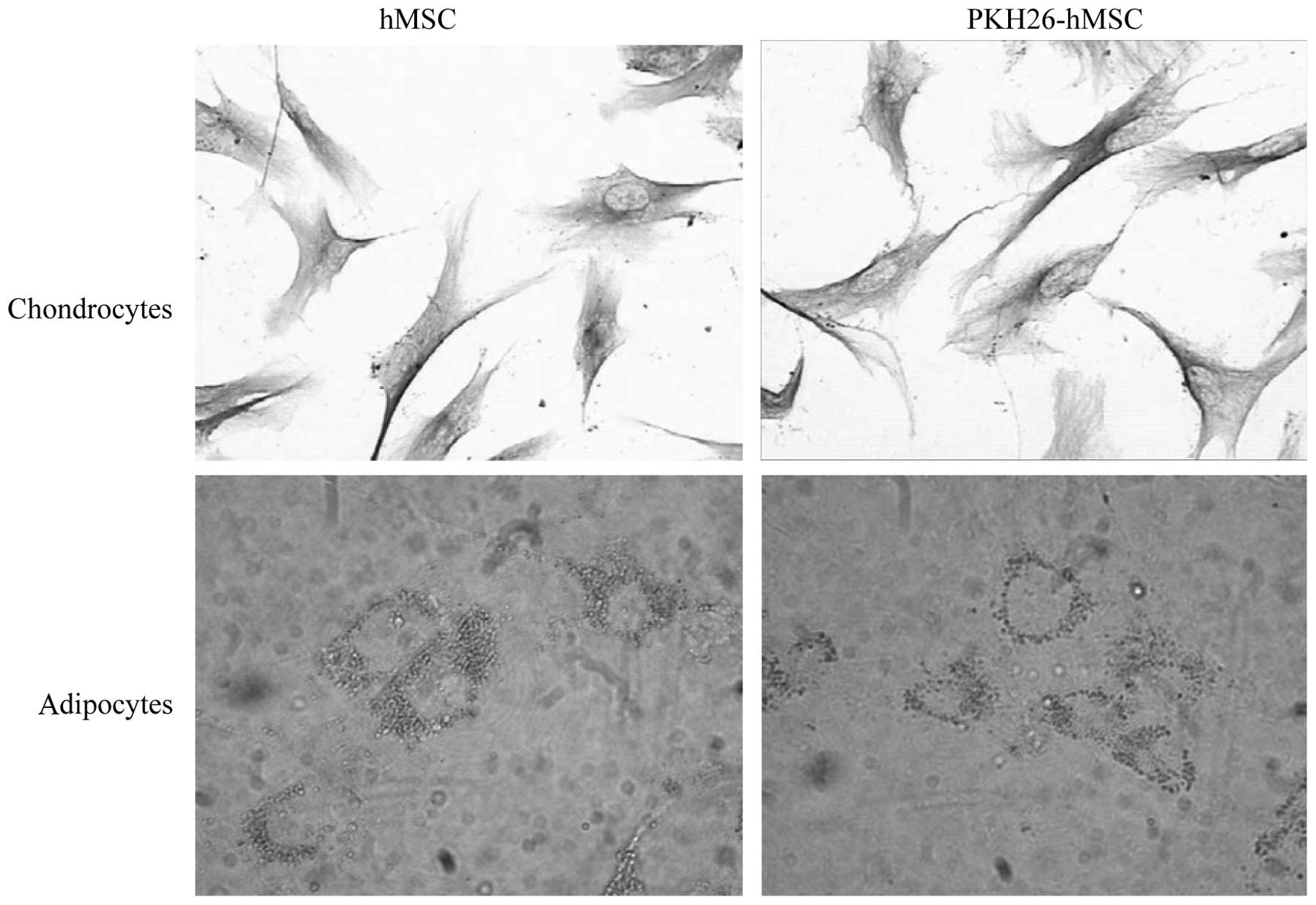
PKH26 labeling does not affect the
differentiation of hMSCs
Following adipocyte induction, the intracellular
lipid droplets continuously enlarged and fused. Subsequent to
chondroblast induction, numerous cells had irregular and polygonal
morphology, the cell volume increased and contained many particles.
The cells contained numerous type II collagen positive brown
particles in the cytoplasm, identified by immunohistochemical
staining. hMSCs and PKH26-hMSCs were differentially induced into
chondrocytes and adipocytes, as demonstrated by Oil Red O staining
and type II collagen immunostaining, respectively (n=3; 30 cells
per repeat; Fig. 5).
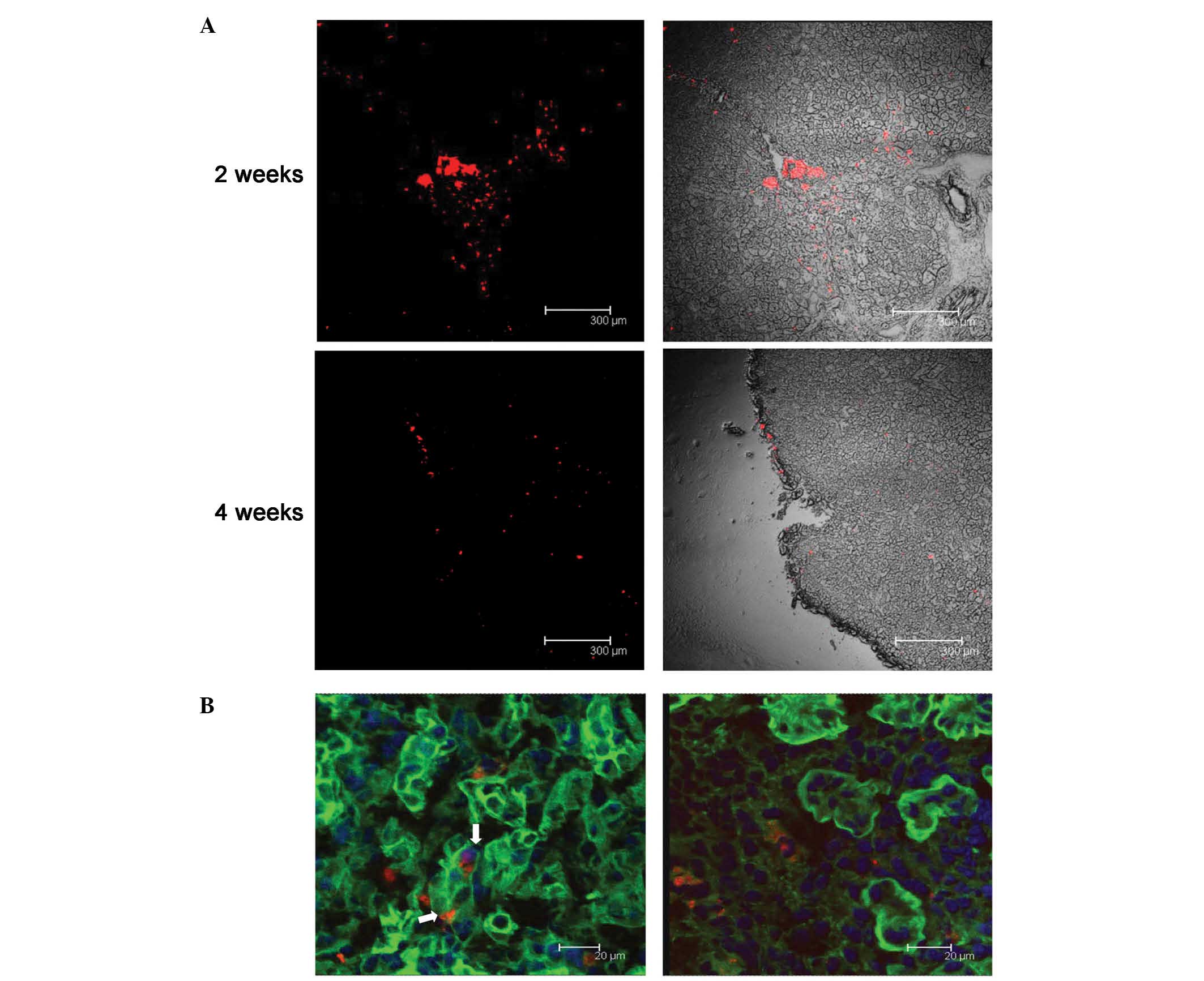
hMSCs may differentiate into renal
tubular epithelium
The PKH26-labeled fluorescent cells were observed to
assemble around the injection site 24 h after the injection.
However, two weeks later, the cells had migrated along the renal
tubule, and the red fluorescent cells were apparent in most tissue
sections. Four weeks following the injection, fluorescence remained
dispersed throughout the kidneys, although the intensity of the
fluorescence had weakened (Fig. 6A).
FITC-labeled WGA was used to visualize the glycoprotein and sialic
acid of cell membranes, and was used to label the renal tubules in
the present study. Using confocal microscopy, co-localization of
FITC-WGA and PKH26-hMSCs were observed, demonstrating these cells
in the kidney tubules. Two weeks after the cell injection, 10±2.1%
of PKH26-labeled hMSCs were demonstrated to be localized to the
renal tubules, as determined by laser scanning confocal microscopy
(Fig. 6B).
Discussion
MSCs are currently used to research organ
development and disease treatment, and these are predominantly
derived from adult bone marrow. It has previously been suggested
that MSCs have multiple roles, which have been investigated in the
treatment of numerous diseases (19); however, the key limitation of using
MSCs in such treatments is the low cell quantity available
(20). The number of MSCs in adult
bone marrow is extremely low, ranging between 0.01 and 0.00001% of
human bone marrow mononuclear cells, according to different donors
(21,22). Furthermore, the quantity and
differentiation of MSCs in bone marrow reduce with increasing age
(23). Compared with adult MSCs,
fetal MSC have a high proliferative capability: In identical in
vitro culture conditions, the population doubling time is
32.3±2.5 h in fetal MSCs and 116.6±22.4 h in adult MSCs (24). In culture, the number of fetal MSCs
can be increased several million fold without the loss of phenotype
such as CD19, CD44, Cd166, Cd103, SH3 and SH4 (11). Adult MSCs express a moderate amount
of human leukocyte antigen (HLA)I and HLAII, while fetal MSCs
express extremely low or no HLAI and do not express HLAII, meaning
that the immunogenicity of fetal MSCs is much weaker (14). Adult stem cell treatment may cause
hyperplasia of the lymphatic system, which is not induced by fetal
MSCs. Fetal MSCs and BMSCs can inhibit adult peripheral blood
lymphocyte (PBL) proliferation induced by numerous mitogensm such
as concanavalin A (ConA), phytohaemagglutinin (PHA),
Staphylococcus aureus (SpA) and poke weed mitogen (PWM)
(11,15). Therefore, fetal MSC treatment has
some unique advantages, such as an improved proliferative capacity,
phenotype stability and low immunogenicity.
PKH26, a lipophilic emitter of red fluorescence, may
be irreversibly bound to the cell membrane to enable fluorescent
labeling of multiple cell types (25). Importantly, under appropriate
labeling conditions, cell activity is not affected: PKH26 labeling
does not affect cell proliferation and the expression of adhesion
molecules (26,27). PKH26 is therefore an ideal tool to
study cell migration and interactions in vivo and in
vitro that is frequently used in cellular tracking studies
(25). Shao-Fang et al
(28) studied the phenotype,
differentiation, proliferation, cell cycle and apoptosis of human
umbilical mesenchymal stromal cells following PKH26 labeling,
reporting that no effect on the above indexes was observed. In the
present study, it was also demonstrated that PKH26 had no
significant effect on cell morphology and proliferation. FACS
results demonstrated that neither hMSCs nor PKH26-labeled hMSCs
expressed hematopoietic lineage markers but did express mesenchymal
cell markers. hMSCs and PKH26-labeled hMSCs were also able to
differentiate into chondrocytes and adipocytes upon induction.
Together, these analyses suggested that PKH26 labeling did not
alter the MSC characteristics in hMSCs, indicating that PKH26
labeling is an effective way to label live hMSCs.
The limitation of PKH26 labeling is the gradual
attenuation of fluorescence, occurring due to absence of sufficient
fluorescent molecules as the cells divide (25,27,29). Ude
et al (27) used PKH26 to
label MSCs and adipose-derived stem cells, revealing that the red
fluorescence remained when the cells were subcultured in
vitro to passage 6 at the 49th day, but that the fluorescence
intensity markedly weakened. In the present study, fluorescence
intensity and the number of PKH26-labeled hMSCs were reduced with
time; four weeks after labeling, only 7% of labeled cells were
observed. Based on the present results, the renal localization of
PKH26-hMSC was also observed at 2 weeks after the injection.
However, recent studies have demonstrated that BMSCs
from mice and humans may cross lineage boundaries and form
functional components of other tissues, expressing tissue-specific
proteins in organs such as the heart, liver, brain, skeletal muscle
and vascular endothelium (30–32).
Numerous experiments have also reported that stem cells have a
therapeutic effect when treating glomerular and tubular diseases
(1,6). In previous studies aiming to treat
tubular damage with stem cells, AKI has been the most widely used
disease model; however, the exact therapeutic mechanism of stem
cells in AKI remains under intensive investigation. For instance, a
previous study demonstrated that stem cells exert their therapeutic
effects in AKI through a paracrine/endocrine mechanism, and that
MSCs produce a variety of cytokines and growth factors (33,34).
Other studies revealed that microvesicles derived from human MSCs
may protect the kidneys from toxic injury through horizontal
transfer of mRNA, RNA-dependent apoptosis resistance and in
vitro proliferation (35,36).
Controversy remains, however, regarding whether MSCs protect the
kidney by transforming to renal tubular epithelium (1,5,6). Several previous studies have also
demonstrated the presence of Y chromosome staining of tubular
epithelial cells in injured kidney transplants from female donors
into male recipients, suggesting that stem cells (SCs) derived from
the male bone marrow migrate and differentiate into the epithelial
cells (37,38). Morigi et al (5) also demonstrated that the renoprotective
effects of BM-derived SC in acute renal failure are not caused by
the hematopoietic stem cells but the MSCs. Broekema et al
(39) demonstrated that the tubular
engraftment of BM-derived cells only occurs dependent on the
severity of renal damage following ischemia-reperfusion injury and
that, when transplanted, these cells acquire an epithelial
phenotype. However, additional previous studies revealed that there
was very little or no tubular incorporation, despite exogenous MSCs
having a protective effect on kidney injury (40–42). The
reason for such discrepancies between these studies is unclear, but
may be associated with the degree of severity of the model and the
protocols used, the types of stem cells and the labeling
methods.
Morigi et al (43) used human cord blood MSC in a mouse
model of acute renal failure, revealing that the quantity of stem
cells reaching the kidney was very low, and the number of stem
cells located in kidney tubules was lower still; most stem cells
were located in the renal interstitium. In this previous study,
there were 2±0.4×105 stem cells located in the kidney,
5±5% of which were located in tubules. A previous study from our
group (3) was also in concordance
with this result: When VEGF-labeled hMSCs and hMSCs were used to
treat AKI, their localizations in the kidney were 1.63±0.68 and
1.58±0.77 hMSCs/section, respectively. In this previous study, most
stem cells were located in the renal interstitium, and only a small
quantity of stem cells was located in the renal tubules. In the
present study, hMSCs were injected into the kidneys of newborn
mice, revealing that 10±2.1% of labeled stem cells were localized
to tubules, and that the proportion of MSCs within the renal
tubular epithelium was higher than that in previous studies
(3,43). The causes for the discrepancy between
this and previous studies may include the following: Firstly,
nephron development was not completed within 2 weeks of the birth
of mice; secondly, immunogenicity of stem cells from embryo was
reduced such that the transplanted stem cells were able to
participate in the development of the renal tubule during this
period (44,45). In our previous study, it was
demonstrated that the number of stem cells recruited in the kidney
through intravenous injection was very limited (3). A large number of MSCs were observed in
other organs besides the kidney, such as in the liver, lungs and
spleen, between 2 h and 4 days of hMSC infusion. In the current
study, numerous competent cells were implanted through a renal
local injection approach.
Yokoo et al (46) reported that MSC cells differentiate
into kidney structures subsequent to their injection into the rat
intermediate mesoderm at the nephrogenic site of the embryo.
However, after 6 days of injection of MSC in the developing
metanephros, it was revealed that MSC were not implanting into the
renal tubules. The authors therefore concluded that MSCs required
nephrogenic signals to participate in metanephros development. In
the present study, however, MSCs were revealed to be implanted in
tubules 2 weeks after the injection, which indicated MSC may
differentiate into tubular epithelium during kidney development.
The difference in present and previous outcomes may be attributed
to different observation times following the injection, different
animal models and types of stem cell.
A previous study by Herzog et al (47) reports cell fusion as a mechanism of
BM-derived cell epithelial differentiation, and Fang et al
(48) demonstrated that among
BM-derived cells integrated into the renal epithelium (~10% of the
cells), a number revealed signs of fusion to the renal tubular
epithelium. The present study did not exclude the fusion of hMSCs
and renal tubular epithelium of mice as a potential mechanism, but
multiple previous in vitro and in vivo studies
suggest that MSCs may express specific molecules and aquaporins of
tubular epithelium (48,49).
In conclusion, it was demonstrated that: i) PKH-26
label had no effect on stem cell proliferation and differentiation
ability; ii) hMSCs may participate in kidney development through
differentiation into renal tubular epithelium when the approach of
renal local injection into the kidney of newborn mice was adopted.
In the present study, it was therefore concluded that hMSCs may be
differentiated into renal tubule epithelium during kidney
development.
Acknowledgements
The present work is supported by the National
Natural Science Foundation of China (grant no. 81200490), the
Medical Science and Technology Development Foundation of Jiangsu
Province Department of Health (grant no. 201213).
References
|
1
|
Morigi M and Benigni A: Mesenchymal stem
cells and kidney repair. Nephrol Dial Transplant. 28:788–793.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Imai E and Iwatani H: The continuing story
of renal repair with stem cells. J Am Soc Nephrol. 18:2423–2424.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yuan L, Wu MJ, Sun HY, Xiong J, Zhang Y,
Liu CY, Fu LL, Liu DM, Liu HQ and Mei CL: VEGF-modified human
embryonic mesenchymal stem cell implantation enhances protection
against cisplatin-induced acute kidney injury. Am J Physiol Renal
Physiol. 300:F207–F218. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Selby NM, Crowley L, Fluck RJ, Mcintyre
CW, Monaghan J, Lawson N and Kolhe NV: Use of electronic results
reporting to diagnose and monitor AKI in hospitalized patients.
Clin J Am Soc Nephrol. 7:533–540. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Morigi M, Imberti B, Zoja C, Corna D,
Tomasoni S, Abbate M, Rottoli D, Angioletti S, Benigni A, Perico N,
et al: Mesenchymal stem cells are renotropic, helping to repair the
kidney and improve function in acute renal failure. J Am Soc
Nephrol. 15:1794–1804. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bussolati B, Hauser PV, Carvalhosa R and
Camussi G: Contribution of stem cells to kidney repair. Curr Stem
Cell Res Ther. 4:2–8. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chhabra P and Brayman KL: The use of stem
cells in kidney disease. Curr Opin Organ Transplant. 14:72–78.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yokote S, Yamanaka S and Yokoo T: De novo
kidney regeneration with stem cells. J Biomed Biotechnol.
2012:4535192012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hwang JH, Shim SS, Seok OS, Lee HY, Woo
SK, Kim BH, Song HR, Lee JK and Park YK: Comparison of cytokine
expression in mesenchymal stem cells from human placenta, cord
blood, and bone marrow. J Korean Med Sci. 24:547–554. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao H, Heazlewood SY, Williams B, Cardozo
D, Nigro J, Oteiza A and Nilsson SK: The role of CD44 in fetal and
adult hematopoietic stem cell regulation. Haematologica. 101:26–37.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gotherstrom C, Ringden O, Westgren M,
Tammik C and Le Blanc K: Immunomodulatory effects of human foetal
liver-derived mesenchymal stem cells. Bone Marrow Transplant.
32:265–272. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li O, Tormin A, Sundberg B, Hyllner J, Le
Blanc K and Scheding S: Human embryonic stem cell-derived
mesenchymal stroma cells (hES-MSCs) engraft in vivo and support
hematopoiesis without suppressing immune function: Implications for
off-the shelf ES-MSC therapies. PLoS One. 8:e553192013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
O'Donoghue K and Fisk NM: Fetal stem
cells. Best Pract Res Clin Obstet Gynaecol. 18:853–875. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Le Blanc K: Immunomodulatory effects of
fetal and adult mesenchymal stem cells. Cytotherapy. 5:485–489.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Le Blanc K, Tammik L, Sundberg B,
Haynesworth SE and Ringden O: Mesenchymal stem cells inhibit and
stimulate mixed lymphocyte cultures and mitogenic responses
independently of the major histocompatibility complex. Scand J
Immunol. 57:11–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Emanueli C, Lako M, Stojkovic M and
Madeddu P: In search of the best candidate for regeneration of
ischemic tissues: Are embryonic/fetal stem cells more advantageous
than adult counterparts? Thromb Haemost. 94:738–749.
2005.PubMed/NCBI
|
|
17
|
Wu M, Yang L, Liu S, Li H, Hui N, Wang F
and Liu H: Differentiation potential of human embryonic mesenchymal
stem cells for skin-related tissue. Br J Dermatol. 155:282–291.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Institute of Laboratory Animal Resources
(US). Committee on Care, Use of Laboratory Animals, and National
Institutes of Health (US). Division of Research Resources: Guide
for the care and use of laboratory animals (8th). National
Academies Press. (Washington, DC). 2011.
|
|
19
|
Schimke MM, Marozin S and Lepperdinger G:
Patient-specific age: The other side of the coin in advanced
mesenchymal stem cell therapy. Front Physiol. 6:3622005.
|
|
20
|
Sterneckert JL, Reinhardt P and Scholer
HR: Investigating human disease using stem cell models. Nat Rev
Genet. 15:625–639. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Allers C, Lasala GP and Minguell JJ:
Presence of osteoclast precursor cells during ex vivo expansion of
bone marrow-derived mesenchymal stem cells for autologous use in
cell therapy. Cytotherapy. 16:454–459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Semon JA, Maness C, Zhang X, Sharkey SA,
Beuttler MM, Shah FS, Pandey AC, Gimble JM, Zhang S, Scruggs BA, et
al: Comparison of human adult stem cells from adipose tissue and
bone marrow in the treatment of experimental autoimmune
encephalomyelitis. Stem Cell Res Ther. 5:22014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Barry FP and Murphy JM: Mesenchymal stem
cells: Clinical applications and biological characterization. Int J
Biochem Cell Biol. 36:568–584. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang ZY, Teoh SH, Chong MS, Schantz JT,
Fisk NM, Choolani MA and Chan J: Superior osteogenic capacity for
bone tissue engineering of fetal compared with perinatal and adult
mesenchymal stem cells. Stem Cells. 27:126–137. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kawaguchi K, Katsuyama Y, Kikkawa S, Setsu
T and Terashima T: PKH26 is an excellent retrograde and anterograde
fluorescent tracer characterized by a small injection site and
strong fluorescence emission. Arch Histol Cytol. 73:65–72. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gallagher SJ, Shank JA, Bochner BS and
Wagner EM: Methods to track leukocyte and erythrocyte transit
through the bronchial vasculature in sheep. J Immunol Methods.
271:89–97. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ude CC, Shamsul BS, Ng MH, Chen HC,
Norhamdan MY, Aminuddin BS and Ruszymah BH: Bone marrow and adipose
stem cells can be tracked with PKH26 until post staining passage 6
in in vitro and in vivo. Tissue Cell. 44:156–163. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shao-Fang Z, Hong-Tian Z, Zhi-Nian Z and
Yuan-Li H: PKH26 as a fluorescent label for live human umbilical
mesenchymal stem cells. In Vitro Cell Dev Biol Anim. 47:516–520.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Haas SJ, Bauer P, Rolfs A and Wree A:
Immunocytochemical characterization of in vitro PKH26-labelled and
intracerebrally transplanted neonatal cells. Acta Histochem.
102:273–280. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fox IJ, Daley GQ, Goldman SA, Huard J,
Kamp TJ and Trucco M: Stem cell therapy. Use of differentiated
pluripotent stem cells as replacement therapy for treating disease.
Science. 345:12473912014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pan Q, Qin X, Ma S, Wang H, Cheng K, Song
X, Gao H, Wang Q, Tao R, Wang Y, et al: Myocardial protective
effect of extracellular superoxide dismutase gene modified bone
marrow mesenchymal stromal cells on infarcted mice hearts.
Theranostics. 4:475–486. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rashid ST and Vallier L: Induced
pluripotent stem cells-alchemist's tale or clinical reality? Expert
Rev Mol Med. 12:252010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Togel F, Hu Z, Weiss K, Isaac J, Lange C
and Westenfelder C: Administered mesenchymal stem cells protect
against ischemic acute renal failure through
differentiation-independent mechanisms. Am J Physiol Renal Physiol.
289:F31–F42. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Semedo P, Palasio CG, Oliveira CD, Feitoza
CQ, Gonçalves GM, Cenedeze MA, Wang PM, Teixeira VP, Reis MA,
Pacheco-Silva A and Câmara NO: Early modulation of inflammation by
mesenchymal stem cell after acute kidney injury. Int
Immunopharmacol. 9:677–682. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gatti S, Bruno S, Deregibus MC, Sordi A,
Cantaluppi V, Tetta C and Camussi G: Microvesicles derived from
human adult mesenchymal stem cells protect against
ischaemia-reperfusion-induced acute and chronic kidney injury.
Nephrol Dial Transplant. 26:1474–1483. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bruno S, Grange C, Deregibus MC, Calogero
RA, Saviozzi S, Collino F, Morando L, Busca A, Falda M, Bussolati
B, et al: Mesenchymal stem cell-derived microvesicles protect
against acute tubular injury. J Am Soc Nephrol. 20:1053–1067. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Poulsom R, Forbes SJ, Hodivala-Dilke K,
Ryan E, Wyles S, Navaratnarasah S, Jeffery R, Hunt T, Alison M,
Cook T, et al: Bone marrow contributes to renal parenchymal
turnover and regeneration. J Pathol. 195:229–235. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gupta S, Verfaillie C, Chmielewski D, Kim
Y and Rosenberg ME: A role for extrarenal cells in the regeneration
following acute renal failure. Kidney Int. 62:1285–1290. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Broekema M, Harmsen MC, Koerts JA,
Petersen AH, van Luyn MJ, Navis G and Popa ER: Determinants of
tubular bone marrow-derived cell engraftment after renal
ischemia/reperfusion in rats. Kidney Int. 68:2572–2581. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lin F, Moran A and Igarashi P: Intrarenal
cells, not bone marrow-derived cells, are the major source for
regeneration in postischemic kidney. J Clin Invest. 115:1756–1764.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Duffield JS, Park KM, Hsiao LL, Kelley VR,
Scadden DT, Ichimura T and Bonventre JV: Restoration of tubular
epithelial cells during repair of the postischemic kidney occurs
independently of bone marrow-derived stem cells. J Clin Invest.
115:1743–1755. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Duffield JS and Bonventre JV: Kidney
tubular epithelium is restored without replacement with bone
marrow-derived cells during repair after ischemic injury. Kidney
Int. 68:1956–1961. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Morigi M, Rota C, Montemurro T,
Montelatici E, Lo Cicero V, Imberti B, Abbate M, Zoja C, Cassis P,
Longaretti L, et al: Life-sparing effect of human cord
blood-mesenchymal stem cells in experimental acute kidney injury.
Stem Cells. 28:513–522. 2010.PubMed/NCBI
|
|
44
|
Sorokin L and Ekblom P: Development of
tubular and glomerular cells of the kidney. Kidney Int. 41:657–664.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Narbaitz R, Vandorpe D and Levine DZ:
Differentiation of renal intercalated cells in fetal and postnatal
rats. Anat Embryol (Berl). 183:353–361. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yokoo T, Ohashi T, Shen JS, Sakurai K,
Miyazaki Y, Utsunomiya Y, Takahashi M, Terada Y, Eto Y, Kawamura T,
et al: Human mesenchymal stem cells in rodent whole-embryo culture
are reprogrammed to contribute to kidney tissues. Proc Natl Acad
Sci USA. 102:3296–3300. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Herzog EL and Krause DS: Engraftment of
marrow-derived epithelial cells: The role of fusion. Proc Am Thorac
Soc. 3:691–695. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fang TC, Alison MR, Cook HT, Jeffery R,
Wright NA and Poulsom R: Proliferation of bone marrow-derived cells
contributes to regeneration after folic acid-induced acute tubular
injury. J Am Soc Nephrol. 16:1723–1732. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yadav N, Rao S, Bhowmik DM and
Mukhopadhyay A: Bone marrow cells contribute to tubular epithelium
regeneration following acute kidney injury induced by mercuric
chloride. Indian J Med Res. 136:211–220. 2012.PubMed/NCBI
|