Introduction
The survival rate of extremely premature infants
(born at <28-weeks of gestation) has been greatly enhanced due
to multiple factors, such as technological advances, the use of
antenatal steroids, surfactant therapy and improvements in
antenatal and postnatal care (1).
However, problems remain regarding the prognosis of nervous system
injuries and neurological disabilities, including cerebral palsy
and other development disorders in premature infants (2,3). Since
the underlying mechanisms of brain injury and repair in premature
infants are unclear, there has been a lack of progress in the
development of clinical treatments for these disorders.
Iron has an important role in vivo due to its
association with hemoglobin and oxygen-carrying function. In the
brain, iron also acts as an important component of enzyme systems;
it is essential for a number of enzymes involved in
neurotransmitter synthesis, including tryptophan hydroxylase
(serotonin), tyrosine hydroxylase (norepinephrine) and dopamine
(4,5). Previous studies have suggested that
iron may contribute to nervous system development in children; an
iron deficiency during infancy has been shown to affect the
development of the nervous system, leading to poor intelligence, a
lack of concentration, learning difficulties and various other
symptoms (4,6). Conversely, other studies have reported
an increased brain iron content in various adult demyelinating
diseases, including Alzheimer's disease, Parkinson's disease,
Huntington's chorea (7–9) and obstructive cerebral hemorrhage
(10,11). Notably, iron chelation therapy,
including deferoxamine, is able to markedly alleviate these
diseases (12). In addition, it has
been hypothesized that iron increases oxidative damage and
participates in associated pathways, which may be a potential
mechanism (13).
The present study hypothesized that iron deficiency
or anemia may protect against the incidence of hypoxic-ischemic
brain injury (HIBI) in neonatal or premature infants, provided that
the anemia is not severe enough to cause serious complications.
Conversely, during the recovery stage of HIBI, the attenuation of
anemia may be beneficial, in order to promote the recovery of nerve
injury and nervous system development in the future.
The present study aimed to investigate the effects
of anemia in an immature rat model of HIBI. In addition, the
therapeutic potential of iron treatment on the rats, and its
underlying mechanism, were analyzed. The results of the present
study may provide a foundation for future studies and identify
novel therapeutic targets for brain injury function rehabilitation
in premature children.
Materials and methods
Experimental design
The present study was conducted at the Experimental
Animal Center of Beijing Friendship Hospital, Capital Medical
University (Beijing, China), during the period of July 2012 to
February 2013. All experimental procedures were approved by the
Animal Study Ethics Committee of Beijing Friendship Hospital, and
were conducted in accordance with the institutional criteria for
the care and use of laboratory animals in research.
Experimental materials
A total of 90 healthy 2-day-old Sprague Dawley (SD)
rats (weight, 6–8 g) of both genders were purchased from Beijing
Vital River Laboratory Animal Technology Co., Ltd. (Beijing,
China). The rats were maintained under a 12-h light/dark cycle at
20–26°C and 40–70% humidity, with ad libitum access to food
and water. The wastes of the animal experiments were disposed of
according to the Experimental Animal Guidelines of the Health
Ministry of the People's Republic of China.
Grouping, modeling, and
interventions
Three-day-old SD rats were randomly divided into the
following six groups: i) The normal control (NC) group (n=21); ii)
the hypoxic-ischemic (HI) group (n=21); iii) the anemia group
(n=21); iv) the HI + anemia group (n=21); v) the early iron (35
mg/kg once daily; Niferex; Schwarz Pharma, Inc., Zhuhai, China)
treatment group (n=3); and vi) the late iron treatment group (n=3).
The NC group underwent normal feeding. Rats in the HI group
underwent bilateral common carotid artery ligation following the
intraperitoneal injection of 10% chloral hydrate (0.5 ml/kg; Meilun
Biology Technology Co., Dailan, China). The skin incisions were
sutured, and the rats were allowed to recover for 1 h, prior to
placement in a sealed water bath containing a hypoxic gas mixture
(92% nitrogen, 8% oxygen) at 37°C for 90 min. In order to establish
the anemia group, a 1-ml syringe needle was inserted at the
strongest beating points of the heart at depth ~0.5 cm, and was
used to take 0.4–0.5 ml blood, and was pressed to stop the
bleeding. Rats in the HI + anemia group were intraperitoneally
injected with 10% chloral hydrate (0.5 ml/kg) and underwent the
same surgical procedure and hypoxic exposure as the HI group, after
which they received the same treatment as the anemia group. In the
early and late iron treatment groups, rats commenced treatment with
iron (35 mg/kg body weight) on days 1 and 7, respectively,
following HI + anemia modeling. Treatments lasted until the end of
the 28-day experimental period.
Tissue specimen preparation
Tissues were collected from 4- and 6-day-old rats
for hematoxylin and eosin (H&E) staining, from 28-day-old rats
for electron microscopic examination, and 4-day-old rats for iron
content and reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) measurements. Rats were anesthetized by the
intraperitoneal injection of 10% chloral hydrate anesthesia (0.5
ml/kg), and then sacrificed by decapitation. Blood tests were
conducted immediately, following the collection of blood in an
anticoagulation tube. Five rats with hemoglobin levels >100 g/l,
tested using Horiba ABX Pentra DF-120 Analyzer (Horiba Medical,
Montpellier, France), were excluded from the anemia and HI + anemia
groups.
Rat brain tissues were harvested under strictly
aseptic conditions. The leptomeningeal vascular surface was
completely stripped, removed and preserved in neutral formalin.
Subsequently, the tissues were sliced into 14-µm sections for
H&E staining, frozen and stored in a refrigerator at −80°C for
iron content and RT-qPCR measurements. They were placed in 2.5%
formaldehyde amyl fixative (CellChip Biotechnology Co., Ltd.,
Beijing, China) for further processing by electron microscopy.
H&E staining
Brain tissues were placed in 30% sucrose overnight
at −4°C. The tissue sections were stained with 10% hematoxylin dye
for 3 min, differentiated with 0.5% ethanol hydrochloride for
several seconds and recovered with lithium carbonate saturated
solution, prior to washing with distilled water. Subsequently, the
tissue sections were stained with 0.5% eosin for 1 min, dehydrated
with gradient ethanol after washing, immersed in dimethyl benzene
and mounted with neutral gum.
Detection of iron content in the
brain
The brain tissues of rats in the NC, HI, anemia and
HI + anemia groups were weighed and washed with phosphate-buffered
saline. The tissue (0.5×0.5 cm) was then mixed with homogenate
buffer (Thermo Fisher Scientific, Inc., Waltham, MA, USA) to a
volume ratio of 1:5, and homogenized at 400 × g for 10 sec, 6
times. Then, the homogenates was centrifuged at 1,600 × g for 15
min at 4°C, and the supernatant was separated. Brain homogenates
were treated with HNO3-H2O2
solution and boiled until transformed into a colorless transparent
liquid. The iron content in the brain tissue was quantified using
atomic absorption spectrophotometry (14).
Immunofluorescence staining of
oligodendrocytes
Brain tissue was immersed in fixative (4%
paraformaldehyde), dehydrated and embedded with optimal cutting
temperature compound. The sections of rat brain tissue were
preserved at −4°C. Then, sections were incubated with polyclonal
primary rabbit anti-carbonic anhydrase II (CAII) antibody (1:200;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at 4°C overnight
followed by incubation with the DyLight 594-labeled goat secondary
antibody (1:500; ZSGB-Bio, Beijing, China) at room temperature for
30 min. Cell nuclei were stained with Hoechst 33342 (ZSGB-Bio). The
survival of oligodendrocytes around the ventricles was observed
visually using an Olympus BX51 fluorescence microscope (Olympus
Corporation, Tokyo, Japan).
Quantification of mRNA expression
levels of iron regulatory protein 2 (IRP2) and transferrin receptor
(TFR) in rat brains
Total RNA was extracted from the brain tissue of the
SD rats using TRIzol reagent (Abcam, Beijing, China) and treated
with DNase I (cat. no. EN0521; Fermentas; Thermo Fisher Scientific,
Inc.). The RNA product was then examined by agarose gel
electrophoresis. Reverse transcription was performed to transcribe
the total RNA into cDNA, using a Thermo First cDNA Synthesis kit
(cat. no. 33-20102; SinoGene Scientific Co., Ltd., Beijing, China).
The reaction system consisted of 10 µl DNase, 1 µl random primer, 4
µl 5X reaction buffer, 2 µl reverse transcriptase enzyme, and
diethylpyrocarbonate water to a total volume of 20 µl. qPCR
reactions were carried out using 2X SG Green qPCR mix (with ROX;
cat. no. 22-10102, SinoGene Scientific Co., Ltd.) with a
StepOnePlus Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The primer sequences were
designed using Primer Premier 5 software (Premier Biosoft
International, Palo Alto, CA, USA) and were as follows: IRP2
forward, TTGTCCATGTTTAAAGCACTGA and reverse, ACACTGAATCTGGAGCGTCTA
(product length, 82 bp); TFR forward, TGGACTGCAGGAGACTAT and
reverse, GTACCCAGGACGACTTTAT (product length, 116 bp); β-actin
forward, CGTTGACATCCGTAAAGACC and reverse, CTAGGAGCCAGAGCAGTAATC
(product length 116 bp). β-actin was used as a reference gene. qPCR
reactions were performed under the following conditions: 10 min at
95°C, followed by 40 cycles of 15 sec at 95°C, 15 sec at 53°C and
35 sec at 72°C. An additional step was used (95°C for 15 sec, 60°C
for 30 sec and 95°C for 15 sec) for dissociation curve analysis.
Data were analyzed using the 2−∆∆Ct method (15).
Electron microscopy
Three 28-day-old newborn rats were selected from
each group and were sacrificed by decapitation, prior to extraction
of the periventricular tissues. The tissues were cut into
1-mm3 samples and immersed in a solution containing 2.0%
paraformaldehyde and 2.5% glutaraldehyde, followed by fixing with
1% osmium tetroxide for 2 h at 4°C. Subsequently, the tissue
sections were washed three times with 0.2 M sodium cacodylate
buffer for 10 min, and then three times with double distilled water
for 10 min, followed by dehydration using alcohol gradients. The
samples were further embedded in epoxy resin (Zhongke Chemical Co.,
Ltd., Tianjin, China), cut into 100 nm slices, stained with uranyl
acetate and lead citrate, and observed under an H-7650 Transmission
Electron Microscope (Hitachi, Ltd., Tokyo, Japan). The brain tissue
ultrastructure was observed under visual observation at ×20,000
magnification, and the number of myelin sheaths was counted under
×5,000 magnification; five fields of view were randomly selected
for each slice.
Statistical analysis
The results from each group were compared using
multivariate analysis of variance. Comparisons between two groups
were performed using the independent samples t-test. Correlation
analysis between two variables was conducted using linear
regression analysis. Statistical analyses were conducted using SPSS
software, version 13.0 (SPSS, Inc., Chicago, IL, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Establishment of a rat model of
anemia
A rat model of anemia was established using immature
newborn rats. The mean hemoglobin content of each group is
presented in Table I. The hemoglobin
contents differed significantly among the groups (P<0.001); the
levels in the anemia and HI + anemia groups were lower than those
in the NC and HI groups, and met the standards of the anemia model,
since they were ≤100 g/l.
 | Table I.Hemoglobin content of each group. |
Table I.
Hemoglobin content of each group.
| Group | Hemoglobin (g/l) |
|---|
| NC group |
141.75±6.54 |
| Anemia group |
79.25±9.49a |
| HI group |
150.12±12.19 |
| HI + anemia
group |
84.88±8.34a |
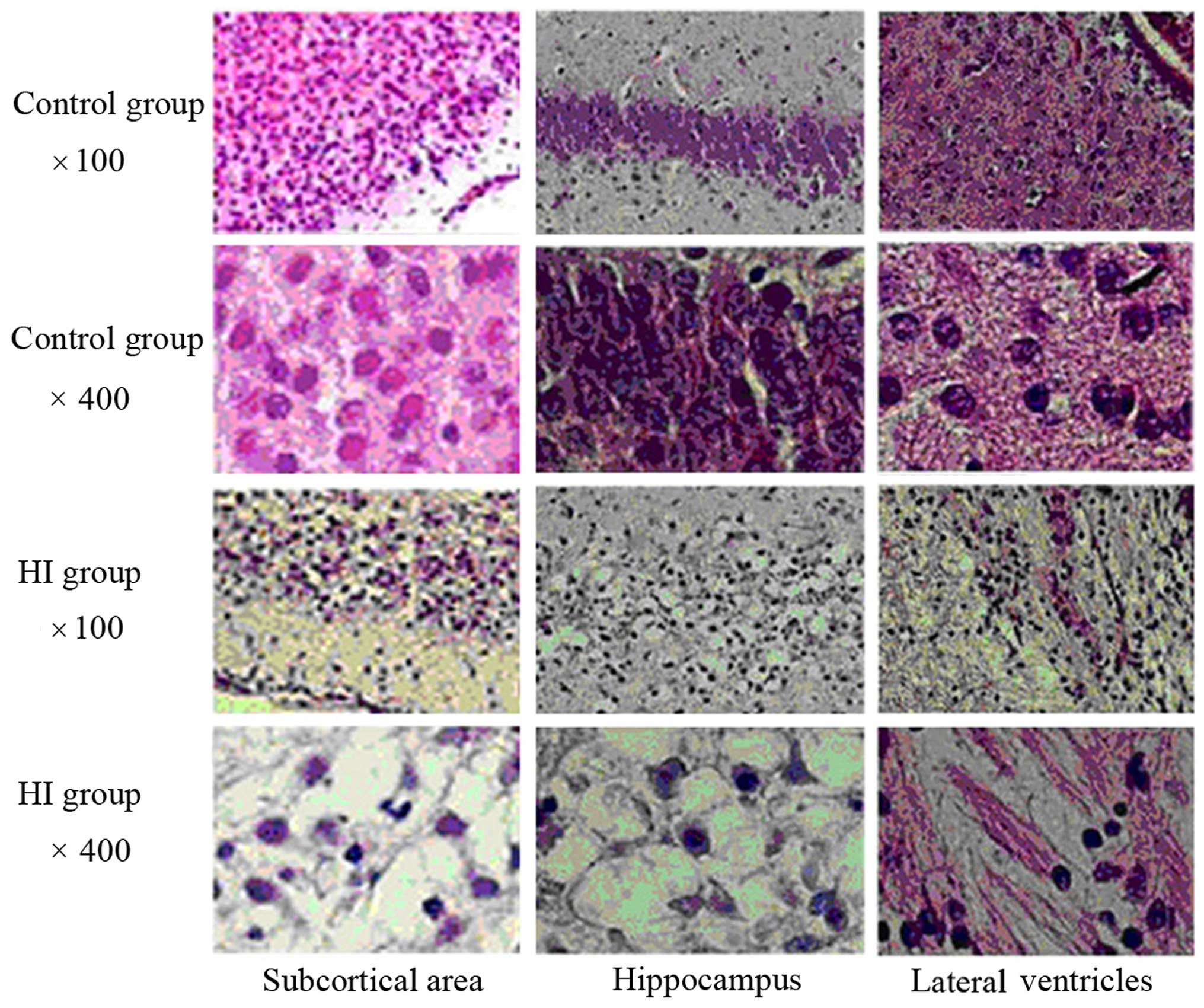
H&E staining of the rat brain
tissue sections
In the NC group, the brain tissues looked healthy,
with neatly ordered, normally shaped cells with a central nucleus
and clearly visible nucleoli (Fig.
1). However, in the HI group, a large number of cells in the
cortical, hippocampal and periventricular areas exhibited necrosis,
vacuolar degeneration, nuclear condensation and fragmentation. In
some cases, dissolution of the nucleus was observed.
Brain iron content
As shown in Table
II, there were significant differences in brain iron content
among the groups (F=5.853; P=0.003). Further analysis indicated
that the iron content in the anemia group was significantly lower
than that in the NC group (P=0.047); whereas the iron content of
the HI group was significantly higher compared with that in the NC
group (P=0.045). No significant differences in iron content were
observed between the HI + anemia group and the NC group (P=0.766).
The iron content of the HI group was significantly higher than that
of the HI + anemia group (P=0.024).
| Table II.Iron content of each group. |
Table II.
Iron content of each group.
| Group | Iron (µg/g) |
|---|
| NC group |
17.45±1.21 |
| Anemia group |
14.29±0.97a,b |
| HI group |
20.62±5.11a,c |
| HI + anemia
group |
16.99±2.89 |
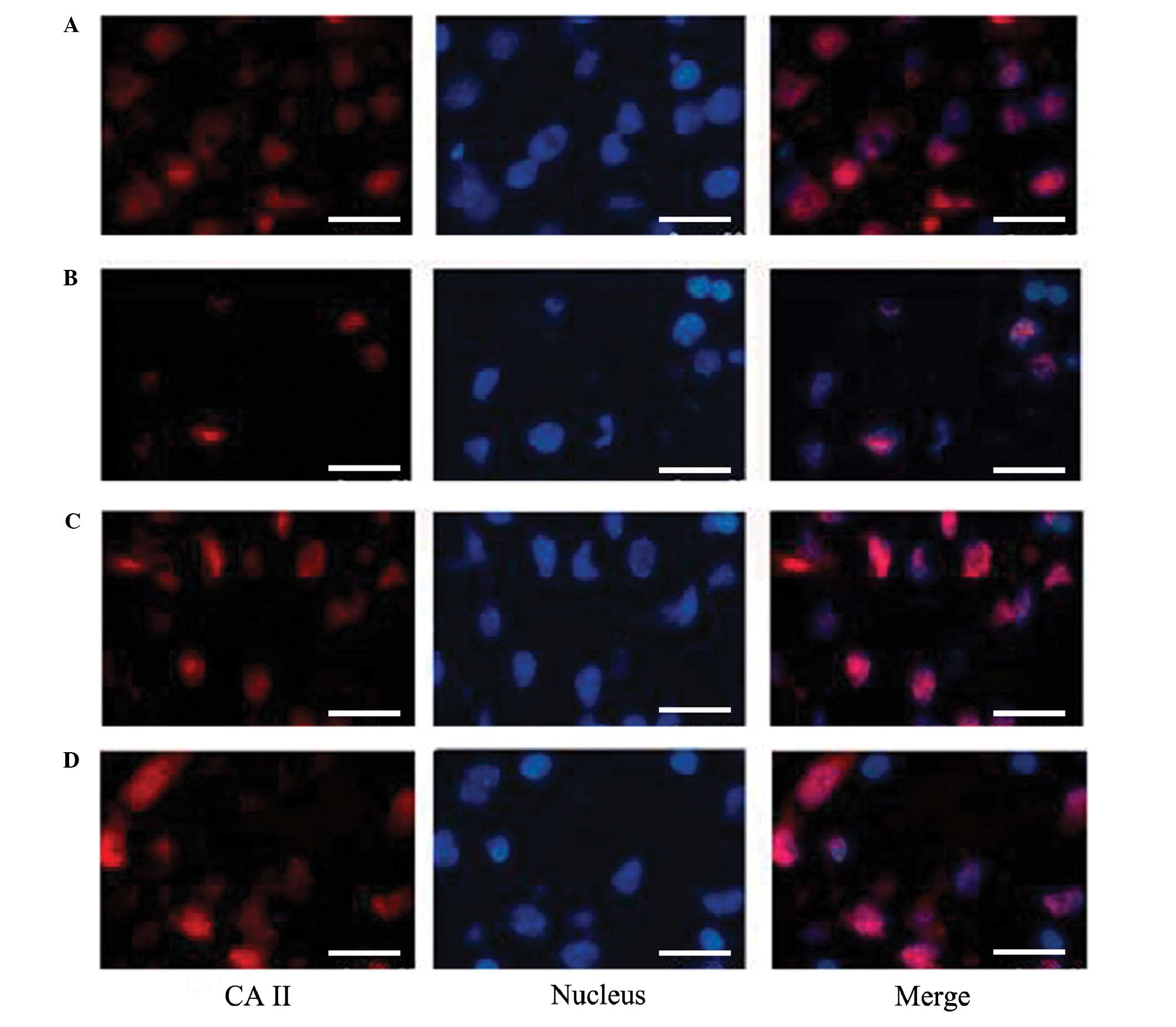
Immunofluorescence of
oligodendrocytes
As depicted in Fig.
2, the number of oligodendrocytes around the ventricles in the
HI group was markedly decreased compared with that in the NC group.
Furthermore, the number of viable oligodendrocyte glial cells in
the ventricles of the anemia group was slightly increased compared
with that in the NC group, and the number of viable
oligodendrocytes in the HI + anemia group was increased compared
with that in the HI group.
mRNA expression levels of IRP2 and TFR
in the rat brains
As shown in Tables
III and IV, there were
significant differences in the mRNA expression levels of IRP2 and
TFR in the rat brains among the groups (IRP2: F=7.220, P=0.003;
TFR: F=5.169, P=0.011). Further analysis suggested that the mRNA
expression levels of IRP2 were significantly decreased in the
anemia group compared with those in the NC group (P=0.05), and that
they were significantly increased in the HI group (P=0.023)
compared with the NC group. There was no significant difference in
IRP2 mRNA expression between the HI + anemia and NC groups
(P=0.851). The mRNA expression levels of IRP2 were significantly
increased in the HI group compared with those in the HI + anemia
group (P=0.033). The changes in the mRNA expression levels of TFR
were similar to those of the mRNA expression levels of IRP2 in each
group. The mRNA expression levels of TFR in the anemia group were
markedly lower than those in the normal control group, but were not
significantly different (P=0.128). The mRNA expression levels of
TFR were significantly higher in the HI group than in the NC group
(P=0.041) and the HI + anemia group (P=0.016). There was no
significant difference between the HI + anemia and NC groups
(P=0.643).
| Table III.mRNA expression levels of IRP2 in each
group (relative levels vs. β-actin). |
Table III.
mRNA expression levels of IRP2 in each
group (relative levels vs. β-actin).
| Group | IRP2 |
| NC group |
1.11±0.13 |
| Anemia group |
0.87±0.25a,b |
| HI group |
1.40±0.17a,c |
| HI + anemia
group |
1.13±0.15 |
| Table IV.mRNA expression levels of TFR in each
group (relative levels vs. β-actin). |
Table IV.
mRNA expression levels of TFR in each
group (relative levels vs. β-actin).
| Group | TFR |
|---|
| NC group |
1.11±0.19 |
| Anemia group |
0.92±0.27a,b |
| HI group |
1.37±0.12b,c |
| HI + anemia
group |
1.05±0.24 |
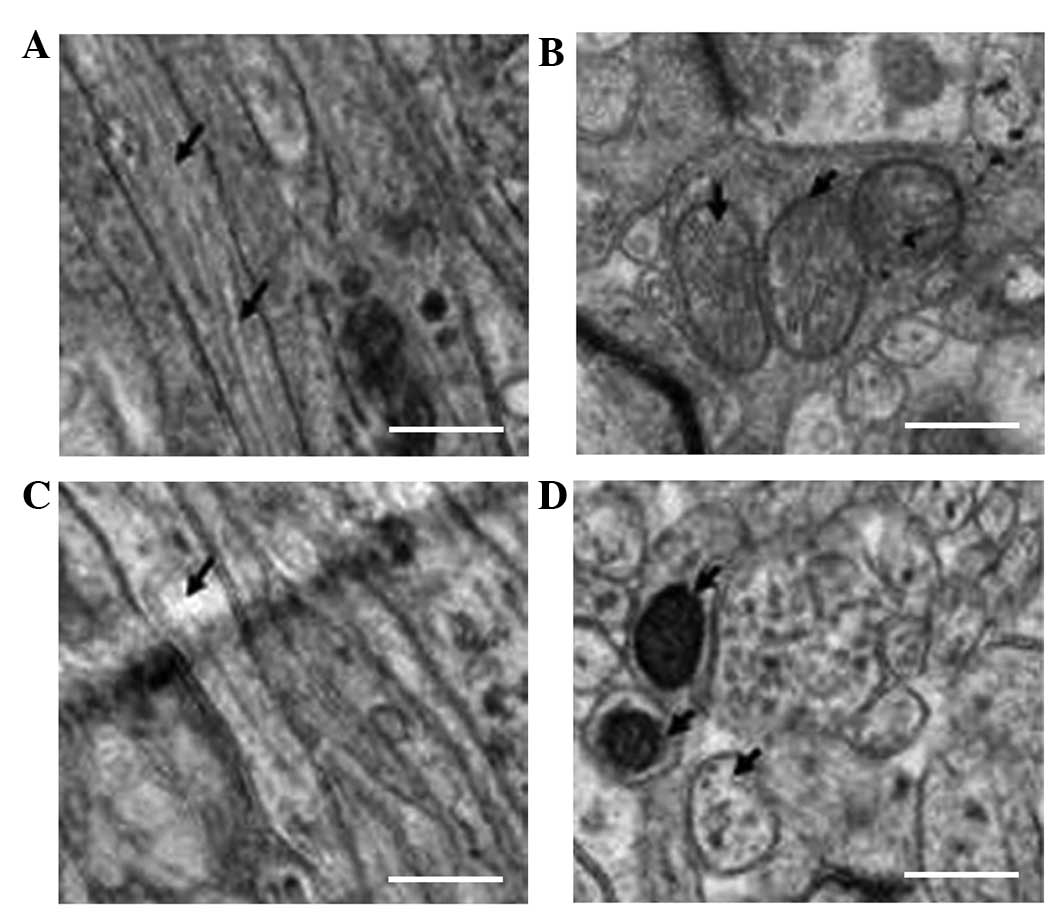
Electron microscopy results
In the rat brains of the NC group a uniform
thickness and density, as well as a regular shape and clear
boundaries of the myelin sheath, were observed in the intact
structure. Furthermore, the nerve cells had a refined structure,
the organelles in the cytoplasm appeared to have a normal
morphology, the mitochondrial structure was intact, the
distribution of chromatin in the nucleus was uniform, the axonal
microtubules and filaments were well-organized and integrity of
structure was detected. In addition, no pathological changes were
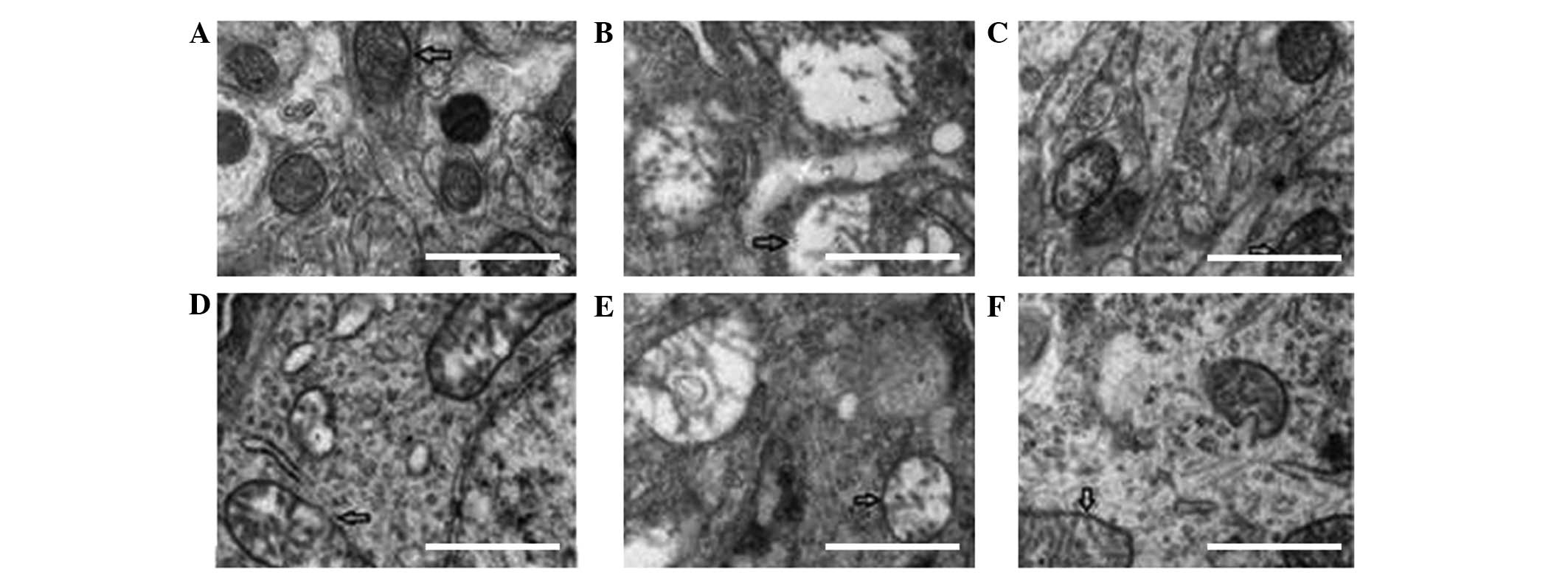
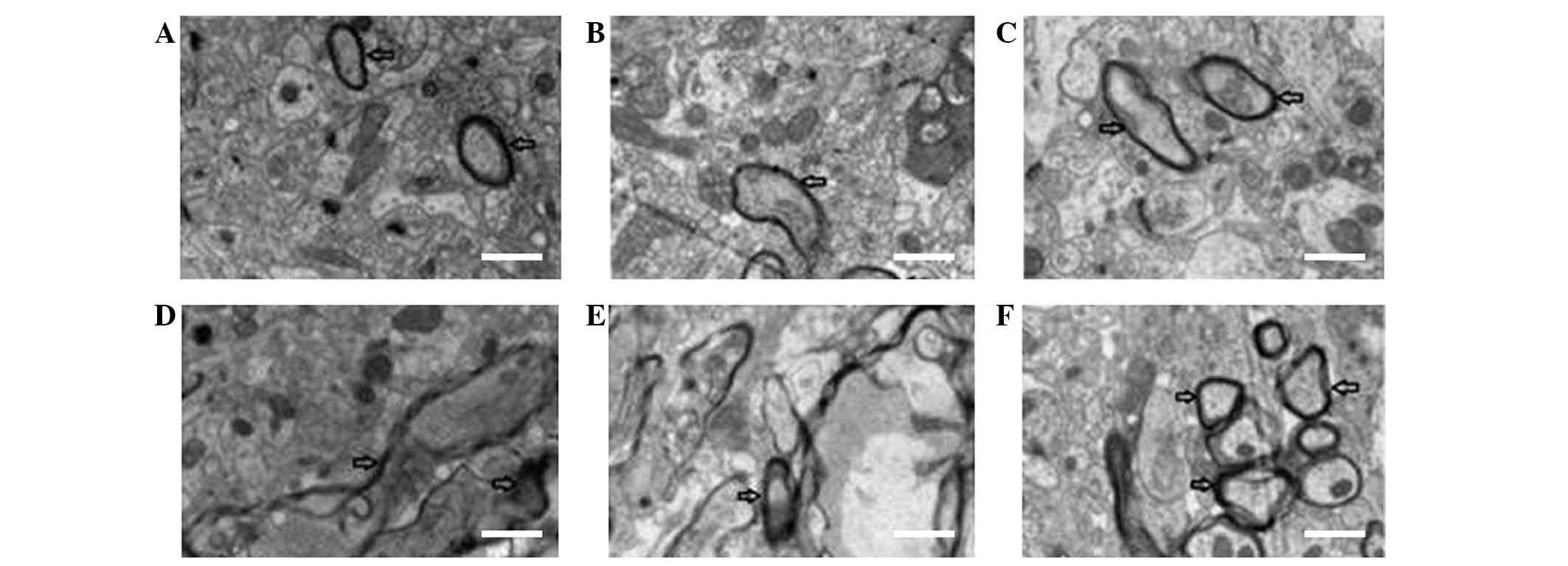
observed in the surrounding areas of the brain (Fig. 3A and B).
The number of myelin sheaths in the rat brains of
the HI group was significantly reduced compared with the number in
the NC group. Furthermore, incomplete structures, irregular
organelle morphologies and a disrupted arrangement were observed.
Edema was evident in the cytoplasmic organelles of the neurons,
including a reduced number of mitochondria in comparison with the
NC group. Similar results were observed for other characteristics,
including edema morphology, ridge breaks and even disappearance,
severe vacuolization of some mitochondria, chromatin condensation
in the nucleus, disappearance of the cytoplasm and nucleus,
swelling of the surrounding nerve fibers and the fracture and
reduction of axonal microtubules and filaments (Fig. 3C and D).
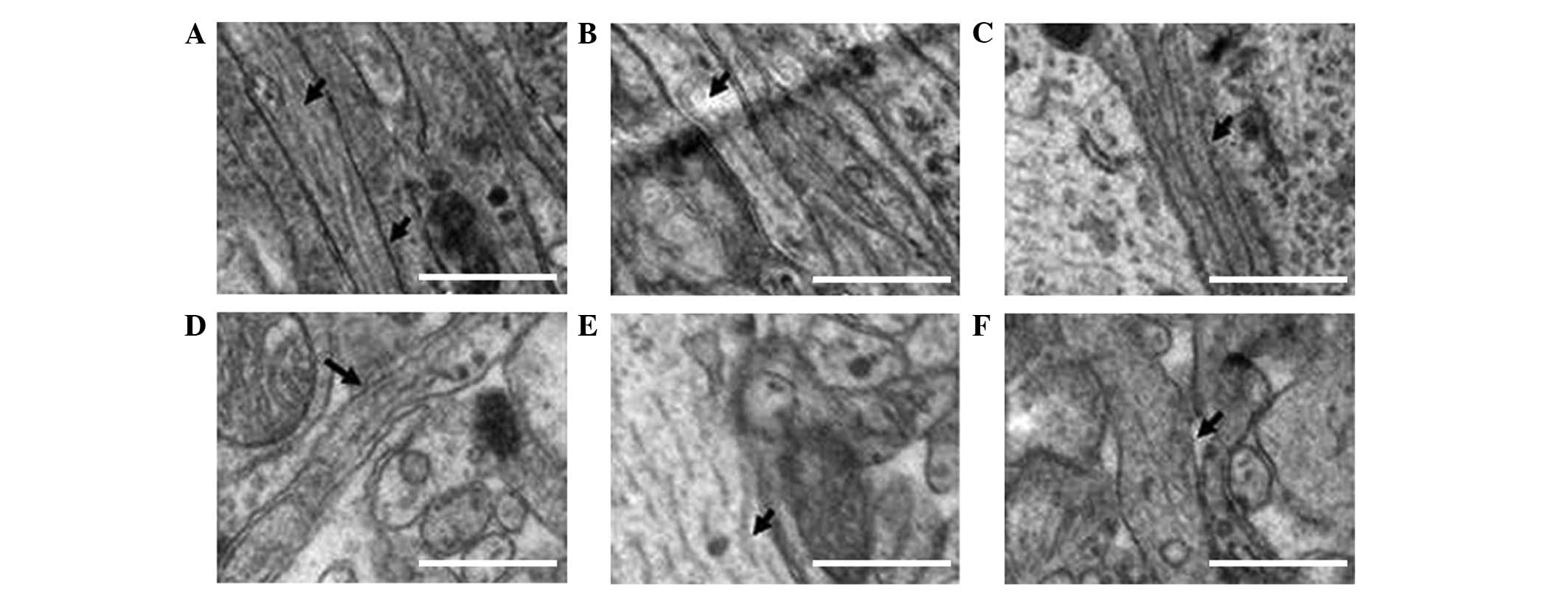
The number of myelin sheaths was markedly decreased
in the anemia group compared with the NC group. Furthermore,
partial incomplete structures and irregular morphology were
observed in the anemia group. Slight edema was detected in the
cytoplasmic organelles of some nerve cells, the number of
mitochondria was decreased, the ridge structure was partially
broken, and there was an even distribution of chromatin in the
nucleus. The filaments, including the microtubules, in axons were
well arranged and their structures appeared to be intact (Figs. 4–6).
As compared with the HI group, the number of myelin
sheaths was markedly increased in the HI + anemia group, and this
was associated with structural improvements. Extended edema of the
neuron organelles and mitochondria was alleviated. In addition, the
ridge structure was well maintained and chromatin condensation in
the nucleus was markedly reduced. Similar to the HI group, only
slight breaks were observed in the filaments and microtubules of
the axons in the HI + anemia group (Figs. 4–6).
As compared with the HI + anemia group, the number
of myelin sheaths was markedly reduced in the early iron treatment
group. This was accompanied by structural disorder, partial
fracture of myelin sheaths and a disordered arrangement. Nerve cell
edema was largely aggregated, with marked reductions in the number
of mitochondria and vacuoles. Furthermore, chromatin condensation
and shrinkage of the nucleus were detected in the nerve cells.
Similar to the HI group, more severe symptoms were observed in the
early iron treatment group, including breakages and a reduced
number of filaments and microtubules in axons, when compared with
the HI + anemia group (Figs.
4–6).
The number of myelin sheaths was markedly increased
and the structure was improved in the late iron treatment group
compared with the HI + anemia group. Alleviation of edema was
detected in the intracellular organelles and mitochondria, and the
ridge structure was well maintained and chromatin condensation in
the nucleus of nerve cells was markedly reduced. However, similar
to the observations for the HI group, there was a significant
improvement in the damaged filaments and microtubules of the axon,
which were decreased in number (Figs.
4–6).
As shown in Table V,
there was a significant difference in the number of myelin sheaths
among the different groups, when observed by low-resolution
electron microscopy (F=118.429, P<0.001). Further analysis
indicated that the number of myelin sheaths in the HI + anemia
group and the HI group were markedly lower than the number in the
NC group (t=12.691, P<0.001; t=6.091, P=0.001, respectively).
The number of myelin sheaths in the early iron treatment group was
significantly reduced compared with that in the HI + anemia group
(t=10.928, P<0.001). The number of myelin sheaths in the late
iron treatment group was significantly lower than that in the NC
group (t=8.356, P<0.001); however, it was significantly higher
than that in the HI + anemia group (t=4.811, P=0.003). As compared
with the HI group, a higher number of myelin sheaths was observed
in the HI + anemia group (t=2.524, P=0.045), although the number of
myelin sheaths in these two groups was significantly lower than the
number in the NC group (t=12.277, P<0.001; t=12.691, P<0.001,
respectively).
| Table V.Number of myelin sheaths in each
group. |
Table V.
Number of myelin sheaths in each
group.
| Group | Myelin count |
|---|
| NC group |
75.00±4.55 |
| HI group |
38.00±3.65a |
| Anemia group |
58.25±3.09a |
| HI + anemia
group |
43.50±2.38a,b |
| Early iron
treatment |
21.25±3.30a,c |
| Late iron
treatment |
52.50±2.88a,c |
Discussion
The production, differentiation and maturation of
oligodendrocyte precursor cells differ between humans and rats; in
particular, human oligodendrocyte precursor cells arise during
early pregnancy (<32 weeks) and gradually differentiate into
mature oligodendrocytes. In addition, the fastest period of axon
myelination occurs during the first year after birth. Conversely,
rat oligodendrocyte precursor cells increase in numbers during the
first week post-birth, and the myelin sheaths are formed during the
first 2 weeks, with the fastest period of myelin formation
occurring during the first 3 weeks after birth (16). The nervous system of a 7-day-old
newborn rat is equivalent to that at 32 weeks of human pregnancy,
and that of a 2–4-day-old newborn rat is similar to that of a human
at middle pregnancy (28–31 weeks) (16). Therefore, the 3-day-old newborn rat
model of HI-induced brain injury established in the present study
is suitable for use as a reference for pre-term infants with
HI-induced brain injury that can be used for the investigation of
potential clinical treatments.
The results of the present study suggested that HI
may lead to an increased postnatal brain iron content. Under an
electron microscope, an incomplete structure, loosely arranged
cells, oligodendrocyte and mitochondria abnormalities, myelin
damage, lamellar separation and a reduced number of myelin sheaths
were observed in the HI group, as compared with 28-day-old rats in
the NC group. These results indicated that the early occurrence of
HI in immature rats may affect the development of the nervous
system; the time following birth for immature rats is a critical
period for oligodendrocyte and myelin development, which causes
oligodendrocyte precursor cells to be highly sensitive to the
effects of adverse factors, such as hypoxia (17). Therefore, myelin damage is more
severe at this time.
When HI was accompanied by anemia, the iron content
of brain tissues was significantly reduced compared with that in
the HI group. Similar pathological changes were observed under the
light microscope in the HI + anemia and HI groups. However, under
the electron microscope, ultrastructural changes were significantly
reduced in the HI + anemia group when compared with those in the HI
group, and suggested that the number of myelin sheaths was
significantly higher in the HI + anemia group compared with the HI
group. These results suggested that anemia was able to reduce the
iron content of the brain and concomitantly attenuate HI-induced
brain injury. Therefore, early anemia may protect against
HI-induced brain injury. However, significant ultrastructural
changes were observed in both the HI + anemia and anemia groups
after 28 days. In addition, the number of myelin sheaths, as
detected using low-resolution electron microscopy, was markedly
lower in the two groups, as compared with the NC group. Therefore,
it may be beneficial to develop a novel intervention strategy for
neonatal anemia, although the time point of intervention may
influence the prognosis if the pre-term infant has suffered from
HI-induced brain injury.
Considerable ultrastructural changes in the brains
of the rats in the HI + anemia group, including changes within the
myelin, synapses and mitochondria, were observed by electron
microscopy. In the early iron treatment group, iron gavage was
commenced 4 days after birth (1 day following the occurrence of HI
and anemia), and continued until 28 days. When observed under an
electron microscope, the number of myelin sheaths was found to be
markedly lower in the early iron treatment group, as compared with
that in the NC, HI + anemia, HI and anemia groups. These results
suggested that iron treatment during the early postnatal stages may
exacerbate brain injury or hinder the repair of brain damage.
Therefore, in cases where HI-induced brain injury is accompanied by
anemia in preterm children, the supplementation of iron during the
early stages of development may not be beneficial.
In the HI-exposed immature rats that had anemia for
a prolonged time period, such as until 10 days post-birth (7 days
following the occurrence of HI and anemia) and were then treated
with iron by gavage until 28 days (HI + anemia group, with late
treatment), the brain injury was markedly alleviated when observed
under an electron microscope. Beneficial effects of late iron
treatment were observed in, for example, the ultrastructure of
myelin, synapses and mitochondria.
The number of myelin sheaths was significantly
higher in the late iron treatment group compared with the early
iron treatment and HI + anemia groups. This suggested that late
iron treatment may be beneficial for repairing the damaged brain,
without incurring any adverse effects. Therefore, late iron
treatment for HI-induced brain injury accompanied by anemia may
facilitate the repair of brain injury.
There are two forms of IRP, namely IRP1 and IRP2
(18–20). When the existence of cytoplasmic RNA
binding proteins, also known as IRPs, was discovered, it was
demonstrated that the proteins were able to regulate the expression
levels of ferritin and transferrin at the post-transcriptional
level, thus suggesting that they may have an important role in
regulating the cellular metabolism of iron (21,22).
Subsequently, IRP2 was discovered, and was shown to have a very
similar structure and function to IRP1 (23–26). In
cells, IRP1 is predominantly regulated by the intracellular iron
concentration; when there is an iron deficiency, the binding
affinity of IRP1 to the iron-responsive element (IRE) in ferritin
RNA is increased, leading to the inhibition of ferritin mRNA
translation. This in turn stabilizes the TFR mRNA, thereby enabling
the cellular uptake of iron and preventing the occurrence of iron
storage (27). Conversely, when the
intracellular iron concentration is increased, the binding affinity
of IRP1 to the IRE in ferritin mRNA is reduced, leading to the
reduced cellular uptake of iron and an increased rate of iron
storage (27). IRP2 is also
regulated by the intracellular iron concentration, although by a
different mechanism. When the intracellular iron concentration is
increased, the binding affinity of IRP2 with mRNA decreases, which
is accompanied by decreased levels of protein expression. This in
turn results in iron-mediated protein hydrolysis (28). The results of the present study
suggested that the mRNA expression levels of IRP2 in the rats of
the HI model group was significantly increased as compared with
that in the NC group, thus suggesting that HI may increase the mRNA
expression levels of IRP2 in the brain, leading to the increased
iron content associated with HI-induced brain damage.
In conclusion, anemia reduced the rate of increase
in iron content in the brain tissues of hypoxic-ischemic
brain-injured newborn immature rats. Iron supplementation during
the early stage of newborn infancy may result in aggravation of
brain injury or affect the recovery of brain damage. However, iron
supplementation at a later stage after birth appears to result in a
significant alleviation of brain injury.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81370741 and
81401245), the National Natural Science Foundation of Beijing and
Key Technology Program of Beijing Education Committee (grant no.
KZ201410025025).
References
|
1
|
Rahman A, Abdellatif M, Sharef SW,
Fazalullah M, Al-Senaidi K, Khan AA, Ahmad M, Kripail M, Abuanza M
and Bataclan F: Changing survival rate of infants born before 26
gestational weeks: Single-centre study. Sultan Qaboos Univ Med J.
15:e351–e356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Olivieri I, Bova SM, Urgesi C, Ariaudo G,
Perotto E, Fazzi E, Stronati M, Fabbro F, Balottin U and Orcesi S:
Outcome of extremely low birth weight infants: What's new in the
third millennium? Neuropsychological profiles at four years. Early
Hum Dev. 88:241–250. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Skiöld B, Vollmer B, Böhm B, Hallberg B,
Horsch S, Mosskin M, Lagercrantz H, Ådén U and Blennow M: Neonatal
magnetic resonance imaging and outcome at age 30 months in
extremely preterm infants. J Pediatr. 160:559–566, e1. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Madan N, Rusia U, Sikka M, Sharma S and
Shankar N: Developmental and neurophysiologic deficits in iron
deficiency in children. Indian J Pediatr. 78:58–64. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Connor JR: Iron acquisition and expression
of iron regulatory proteins in the developing brain: Manipulation
by ethanol exposure, iron deprivation and cellular dysfunction. Dev
Neurosci. 16:233–247. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Grantham-Mcgregor S and Baker-Henningham
H: Iron deficiency in childhood: Causes and consequences for child
development. Annales Nestlé. 68:105–119. 2010. View Article : Google Scholar
|
|
7
|
Ong WY, Tanaka K, Dawe GS, Ittner LM and
Farooqui AA: Slow excitotoxicity in Alzheimer's disease. J
Alzheimers Dis. 35:643–668. 2013.PubMed/NCBI
|
|
8
|
Suttkus A, Rohn S, Jäger C, Arendt T and
Morawski M: Neuroprotection against iron-induced cell death by
perineuronal nets - an in vivo analysis of oxidative stress. Am J
Neurodegener Dis. 1:122–129. 2012.PubMed/NCBI
|
|
9
|
Bartzokis G, Lu PH, Tishler TA, Fong SM,
Oluwadara B, Finn JP, Huang D, Bordelon Y, Mintz J and Perlman S:
Myelin breakdown and iron changes in Huntington's disease:
Pathogenesis and treatment implications. Neurochem Res.
32:1655–1664. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tanskanen M, Mäkelä M, Myllykangas L,
Rastas S, Sulkava R and Paetau A: Intracerebral hemorrhage in the
oldest old: A population-based study (Vantaa 85+). Front Neurol.
3:1032012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dong M, Xi G, Keep RF and Hua Y: Role of
iron in brain lipocalin 2 upregulation after intracerebral
hemorrhage in rats. Brain Res. 1505:86–92. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo C, Wang P, Zhong ML, Wang T, Huang XS,
Li JY and Wang ZY: Deferoxamine inhibits iron induced hippocampal
tau phosphorylation in the Alzheimer transgenic mouse brain.
Neurochem Int. 62:165–172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kostic M, Zivkovic N and Stojanovic I:
Multiple sclerosis and glutamate excitotoxicity. Rev Neurosci.
24:71–88. 2013.PubMed/NCBI
|
|
14
|
Fitsanakis V A, Zhang N, Anderson J G,
Erikson KM, Avison MJ, Gore JC and Aschner M: Measuring brain
manganese and iron accumulation in rats following 14 weeks of
low-dose manganese treatment using atomic absorption spectroscopy
and magnetic resonance imaging. Toxicol Sci. 103:116–124. 2008.
View Article : Google Scholar
|
|
15
|
Arocho A, Chen B, Ladanyi M and Pan Q:
Validation of the 2-DeltaDeltaCt calculation as an alternate method
of data analysis for quantitative PCR of BCR-ABL P210 transcripts.
Diagn Mol Pathol. 15:56–61. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Back SA, Luo NL, Borenstein NS, Levine JM,
Volpe JJ and Kinney HC: Late oligodendrocyte progenitors coincide
with the developmental window of vulnerability for human perinatal
white matter injury. J Neurosci. 21:1302–1312. 2001.PubMed/NCBI
|
|
17
|
Khwaja O and Volpe JJ: Pathogenesis of
cerebral white matter injury of prematurity. Arch Dis Child-Fetal
Neonatal Ed. 93:F153–F161. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Muckenthaler MU, Galy B and Hentze MW:
Systemic iron homeostasis and the iron-responsive
element/iron-regulatory protein (IRE/IRP) regulatory network. Annu
Rev Nutr. 28:197–213. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wallander ML, Leibold EA and Eisenstein
RS: Molecular control of vertebrate iron homeostasis by iron
regulatory proteins. Biochim Biophys Acta. 1763:668–689. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ganz T: Hepcidin and iron regulation, 10
years later. Blood. 117:4425–4433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Davis M and Clarke S: Influence of
microRNA on the maintenance of human iron metabolism. Nutrients.
5:2611–2628. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Theil EC: Ferritin: The protein nanocage
and iron biomineral in health and in disease. Inorg Chem.
52:12223–12233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sanchez M, Galy B, Schwanhaeusser B, Blake
J, Bähr-Ivacevic T, Benes V, Selbach M, Muckenthaler MU and Hentze
MW: Iron regulatory protein-1 and-2: Transcriptome-wide definition
of binding mRNAs and shaping of the cellular proteome by iron
regulatory proteins. Blood. 118:e168–e179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shpyleva SI, Tryndyak VP, Kovalchuk O,
Starlard-Davenport A, Chekhun VF, Beland FA and Pogribny IP: Role
of ferritin alterations in human breast cancer cells. Breast Cancer
Res Treat. 126:63–71. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang W, Deng Z, Hatcher H, Miller LD, Di
X, Tesfay L, Sui G, D'Agostino RB Jr, Torti FM and Torti SV: IRP2
regulates breast tumor growth. Cancer Res. 74:497–507. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
do Nascimento PR, Martins DR, Monteiro GR,
Queiroz PV, Freire-Neto FP, Queiroz JW, Morais Lima AL and Jeronimo
SM: Association of pro-inflammatory cytokines and iron regulatory
protein 2 (IRP2) with Leishmania burden in canine visceral
leishmaniasis. PLOS One. 8:e738732013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Khan MA, Ma J, Walden WE, Merrick WC,
Theil EC and Goss DJ: Rapid kinetics of iron responsive element
(IRE) RNA/iron regulatory protein 1 and IRE-RNA/eIF4F complexes
respond differently to metal ions. Nucleic Acids Res. 42:6567–6577.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hausmann A, Lee J and Pantopoulos K: Redox
control of iron regulatory protein 2 stability. FEBS Lett.
585:687–692. 2011. View Article : Google Scholar : PubMed/NCBI
|