Introduction
Melanoma originates in pigmented melanocytes derived
from neural crest cells that are normally present in the epidermis
and dermis (1). Melanoma represents
<5% of skin tumors but carries the greatest mortality rate of
all skin neoplasms (2). The global
incidence of melanoma has increased markedly (3). Melanoma is able to metastasize through
the hematogenous or lymphatic system (4,5).
Advanced-stage, metastatic disease confers a poor prognosis, with a
median survival of less than one year (3,6).
The Wilms tumor 1 (WT1) protein is a transcription
factor that regulates the expression of genes involved in cell
proliferation and apoptosis (7–11). WT1
expression is essential for genitourinary development, and ~10% of
nephroblastomas exhibit WT1 mutations (12). By contrast, wild-type WT1 is
overexpressed in a variety of neoplasms, including lung, breast,
thyroid and melanoma (13–17).
WT1 is expressed in >80% of malignant melanoma
cells, but is not present in vivo in normal skin or benign
melanocytic nevi (17). Expression
of WT1 is associated with melanoma cell proliferation and is a
possible marker of melanocytic invasion into the dermis (18,19).
In vitro knockdown of WT1 induces apoptosis and increases
sensitivity to chemotherapy in B16F10 melanoma cells. Furthermore,
in vivo WT1 short hairpin RNA (shRNA) applied by aerosol
reduces the number and size of tumors (20,21).
Gene therapy using tumor-targeted liposomes as
delivery systems has opened a new era in cancer treatment (22). Liposomes are essentially phospholipid
bilayer envelopes capable of systemically delivering drugs or
genetic material. The advantage of using phospholipids for systemic
delivery is that they are biodegradable, minimally toxic and easily
removed (23,24). Recent studies show that the addition
of polyethylene glycol (PEG) to phospholipids increases the
half-life of liposomes in circulation; this modification also
improves the ability to incorporate ligand molecules that
specifically recognize superficial tumor targets (25–27).
Transferrin (Tf), a glycoprotein required for
cellular absorption of iron, is often used in gene or drug delivery
systems (28–37). As the Tf receptor (CD71) is
overexpressed on the surface of cancer cells, it is a useful target
for the delivery of therapeutic agents such as small-molecule drugs
and nucleic acids (26,31,37–40).
The aim of the present study was to analyze the
antitumor effects of Tf-conjugated PEG liposomes loaded with
WT1 shRNA applied systemically to a subcutaneous model of
melanoma in C57BL/6 mice.
Materials and methods
Materials
One-palmitoyl-2-oleoyl-sn-glycerol-3-phosphocholine (POPC),
dimethyldioctadecyl ammonium bromide (DDAB), pegylated
distearoylphosphatidylethanolamine (DSPE-PEG 2000) and DSPE-PEG
2000 conjugated to maleimide were purchased from Avanti Polar
Lipids, Inc. (Alabaster, AL, USA). DNase I was purchased from
Sigma-Aldrich (St. Louis, MO, USA). Polycarbonate membranes of 100
nm were purchased from Whatman International, Ltd. (Maidstone, UK).
Amicon Centriprep (molecular weight cut-off, 30 kDa) concentrator
was purchased from EMD Millipore (Billerica, MA, USA).
Holo-transferrin and 2-iminothiolane hydrochloride (Traut's
reagent) were purchased from Sigma-Aldrich. WT1.1 DNA
plasmid construction was performed as described by Zamora-Avila
et al (20). B16F10 murine
melanoma cells were obtained from American Type Culture Collection
(ATCC; Manassas, VA, USA). Rat anti-mouse CD71-(FITC) antibody
clone C2 was purchased from BD Biosciences (San Jose, CA, USA). The
pEGFP-N3 vector that expressed GFP protein was obtained from
Clontech Laboratories, Inc. (Mountain View, CA, USA). Anti-WT1 F6
antibody (sc-7385) was purchased from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). Anti-glyceraldehyde-3-phosphate
dehydrogenase (GAPDH; SAB2500451-100UG) antibody was purchased from
Sigma-Aldrich. Anti-goat secondary antibody (#170-6515) was
purchased from Bio-Rad Laboratories, Inc. (Hercules, CA, USA), and
anti-mouse secondary antibody (sc-516086) was purchased from Santa
Cruz Biotechnology, Inc.
Construction of WT1 shRNA plasmid
WT1 RNAi was designed using siRNA design
software available online (Ambion; Thermo Fisher Scientific, Inc.,
Austin, TX, USA), and synthesized at 0.05 mg (Ambion; Thermo Fisher
Scientific, Inc.) to create one recombinant plasmid (named WT1-1).
Oligonucleotides for the WT1-1 plasmid were
5′-GATCCGGCTGTCCCACTTACAGATGGAAGCTTGCATCTGTAAGTGGGACAGCTTTTTTGGAAG-3′
and
3′-GCCGACAGGGTGAATGTCTACCTTCGAACGTAGACATTCACCCTGTCGAAAAAACCTTCGCCGG-5′.
The oligonucleotides were resuspended to a final concentration of 1
mg/ml and annealed and ligated into pGSH1-GFP (Gene Therapy
Systems, Inc., San Diego, CA, USA).
Western blot analysis
Tumoral tissue samples (25 mg) were lysed with
TRIzol reagent according to manufacturer's instructions (Thermo
Fisher Scientific, Inc., Gaithersburg, MD, USA). Protein
concentration was determined using a DC protein assay kit. Proteins
(50 µg whole-cell lysates) was electrophoresed on 12%
SDS-polyacrylamide gels and transferred to nitrocellulose membranes
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Blocking was
realized with 5% lactose-free milk and Tween 20 in incubation for 1
h. Three washes were performed after each hybridizing with the
above antibodies. Monoclonal anti-WT1 F6 was used at a dilution of
1:2,500 in Tris-buffered saline (TBS) buffer, and secondary
antibody was used at 1:5,000 in TBS buffer. To control for protein
loading, anti-GAPDH was used at 1:10,000 in phosphate-buffered
saline (PBS), and anti-goat secondary antibody was used at 1:5,000.
All antibody incubations were 90 min at room temperature. Protein
bands were visualized by enhanced chemiluminescence using
Lumi-Light Western blotting substrate Roche Diagnostics
(Indianapolis, IN, USA) .
Flow cytometric analysis
B16F10 melanoma cells (1×106) were
resuspended in 200 µl PBS buffer and stained with anti-mouse CD71
(FITC) antibody. Cells were then incubated at 4°C for 30 min and
spun at 400 × g for 10 min at 4°C. Cells were then washed twice
with PBS buffer and spun at 1,600 rpm for 10 min at 4°C. Cells were
suspended in 200 µl PBS and analyzed using a Accuri C6 flow
cytometer (BD Biosciences) to assess cellular expression of
CD71.
Liposomal preparation
Liposomal formulation was performed according to Shi
and Pardridge (41). POPC (19.2
µmol), DDAB (0.2 µmol), DSPE-PEG 2000 (0.6 µmol), and DSPE-PEG
2000-maleimide (30 nmol) were dissolved in a mixture of
chloroform/methanol (2:1 in a total volume of 3 ml) and
subsequently evaporated using nitrogen gas. Lipids were then
suspended in 0.2 ml PBS buffer containing 200 µg plasmid DNA and
vortexed vigorously for 2 min. The liposome/DNA suspension was
frozen in ethanol/dry ice for 4 min and thawed at 40°C for 2 min in
a cycle 10 times. Adjustments to the size of the liposomes were
performed using polycarbonate membranes containing a 100-nm pore
size; this step was repeated five times. After extrusion, the
liposomes were graduated to 2 ml and stored in the dark at 4°C. To
remove unincorporated DNA inside the liposome, 5 U endonuclease I
and 5 mM MgCl2 were added to the mixture and incubated
for 1 h at 37°C. DNA concentration was calculated at ~50 and 60%
using NanoDrop 2000 (Thermo Fisher Scientific, Inc., Waltham, MA,
USA).
Anchoring of the protein to liposome was performed
using 10 nmol Tf protein (Sigma-Aldrich) and thiolated using 400
µmol 2-iminothiolane (Traut's reagent) in 100 µl borate-EDTA
buffer, pH 8.5 (0.15 M sodium borate and 0.1 mM EDTA). The mixture
was incubated for 2 h in the dark at room temperature with shaking.
Loaded Tf was washed with PBS in a concentration column (Centricon
30; Amicon) and concentrated to a final volume of 0.2 ml. The
loaded Tf was immediately added to the liposomes and incubated for
24 h at 4°C to allow reaction of the maleimide group to generate
PEG-immunoliposome complexes. The complexes were dialyzed using an
50-nm membrane to exclude non-incorporated materials. The mean
vesicle diameters were determined using a Zetasizer Nano ZS90
particle size analyzer (Malvern Instruments, Ltd., Malvern,
UK).
Subcutaneous in vivo model of
melanoma
Female C57BL/6 mice (age, 7–8 weeks; weight, 20–25
g) were obtained from Harlan Laboratories S.A. De C.V. (Distrito
Federal, Mexico). Seven mice in each group were used in the assay.
Animals were housed under a 12-h light/dark cycle and received an
autoclaved rodent diet and water ad libitum. All experiments
were performed with prior approval from the local animal ethics
committee. Four groups of seven mice were used for the in
vivo assay. A subcutaneous model of melanoma was developed
using the B16F10 cell line (CRL-6475; ATCC), which is derived from
the same mouse species. A suspension of 200 µl containing
5×105 cells was subcutaneously injected into the
posterior right flank of the mouse. Complete complex of
Tf-conjugated PEG liposomes (50 µl) were loaded with shRNA against
WT1 (Lip + RNAi + Tf), PEG liposomes (50 µl) and loaded with
shRNA against WT1 (Lip + RNAi), Tf-conjugated PEG liposomes (50 µl)
and loaded with pEGFP-N3 vector (Lip + GFP + Tf) and saline
solution (50 µl; untreated group) were delivered intravenously
through the tail vein on day 4 and every 5 days subsequently until
day 29. Surviving mice were sacrificed on day 30 by cervical
dislocation according the good management practices guidelines of
laboratory animals, and tumors were collected for further analyses.
Mouse weight, tumor weight and date of death were recorded.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from 25 mg tumoral tissue
using 1 ml TRIzol according to manufacturer's instructions. The
cDNA samples were obtained using 5 µg total RNA, 200 U Superscript
III and 0.5 µg oligo dT (12–17) at
42°C for 90 min, followed by heating at 70°C for 10 min.
TaqMan® One-Step RT-PCR Master Mix Reagents Kit
manufactured by Applied Biosystems (Thermo Fisher Scientific, Inc.,
Foster City, CA, USA).
Each qPCR reaction was performed with 2 µl cDNA and
WT1 forward primer, TCTGCGGAGCCCAATACAG, reverse primer,
CACATCCTGAATGCCTCTGAAGA, and probe FAM-CACCGTGCGTGTGTATT-NFQ. As an
endogenous control, a mouse β-actin primer set was used,
manufactured by Applied Biosystems. For each reaction, we used
Universal PCR Master Mix manufactured by Roche Molecular Systems,
Inc. (Branchburg, NJ, USA). The protocol was performed for 40
cycles at 94°C for 30 sec and 64°C for 30 sec using a Real-Time
Thermal cycler CROMO4 (Bio-Rad Laboratories, Inc.). Relative
quantification was performed using the Livak method (42). All samples were run in duplicate,
destilled water was including as negative control of reacction.
Statistical analysis
Significance of different treatments was determined
by analysis of variance by Dunnett's test. To survival rate were
performed Kaplan-Meier curves, using SPSS software, version 13
(SPSS, Inc., Chicago, IL, USA) All data are expressed as the mean ±
the standard errors of the mean. P<0.05 was considered to
indicate a statistically significant difference.
Results
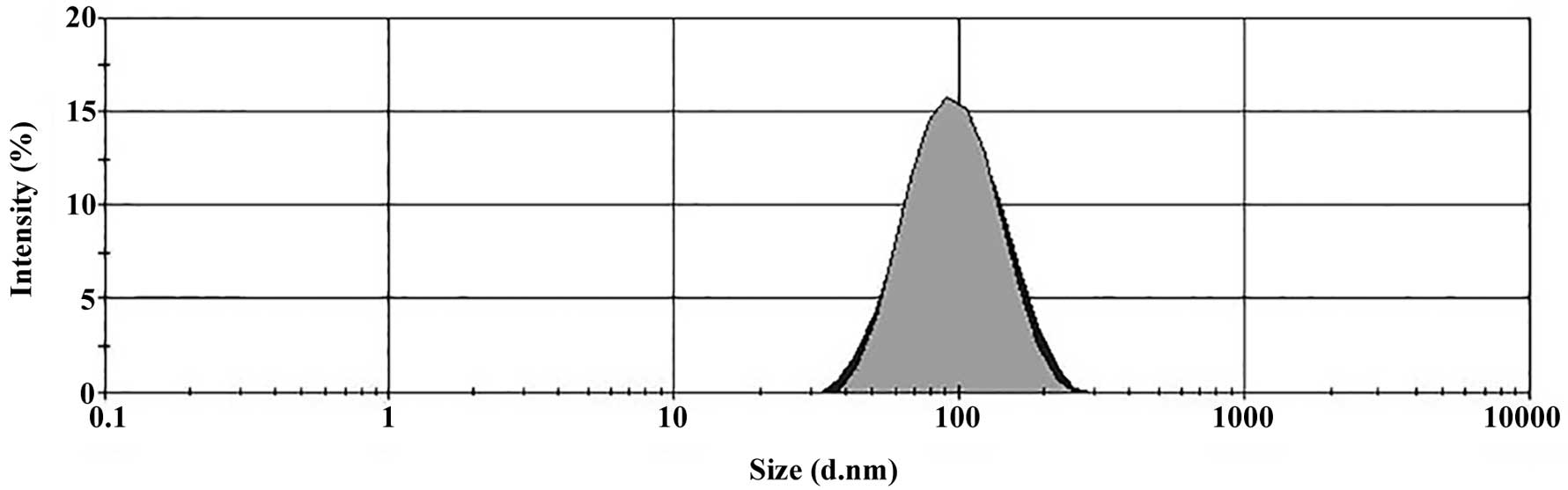
Size distribution according to
intensity of liposomes
Sizes of the Tf-conjugated PEG liposomes loaded with
WT1 shRNA are shown in Fig.
1. The mean diameter of the PEGylated Tf-conjugated PEG
liposomes loaded with shRNA against WT1 was 86 nm. The
poly-dispersity index (PDI) was 0.154, indicating that the vesicles
have a high level of homogeneity. Liposome formulation by vortexing
and repeated cycles of freezing-thawing allowed the formation of
unilamellar liposomes. Liposomes contained a high concentration of
the neutral phospholipid POPC and a minor quantity of cationic
lipid DDAB (0.2 mol), which improved the incorporation of nucleic
acid into the aqueous liposome center. A smaller quantity of
DSPE-PEG 2000 (0.6 mol) and DSPE-PEG 2000-maleimide (30 nmol)
provided steric properties, and the maleimide group allowed the
addition of thiolated transferrin.
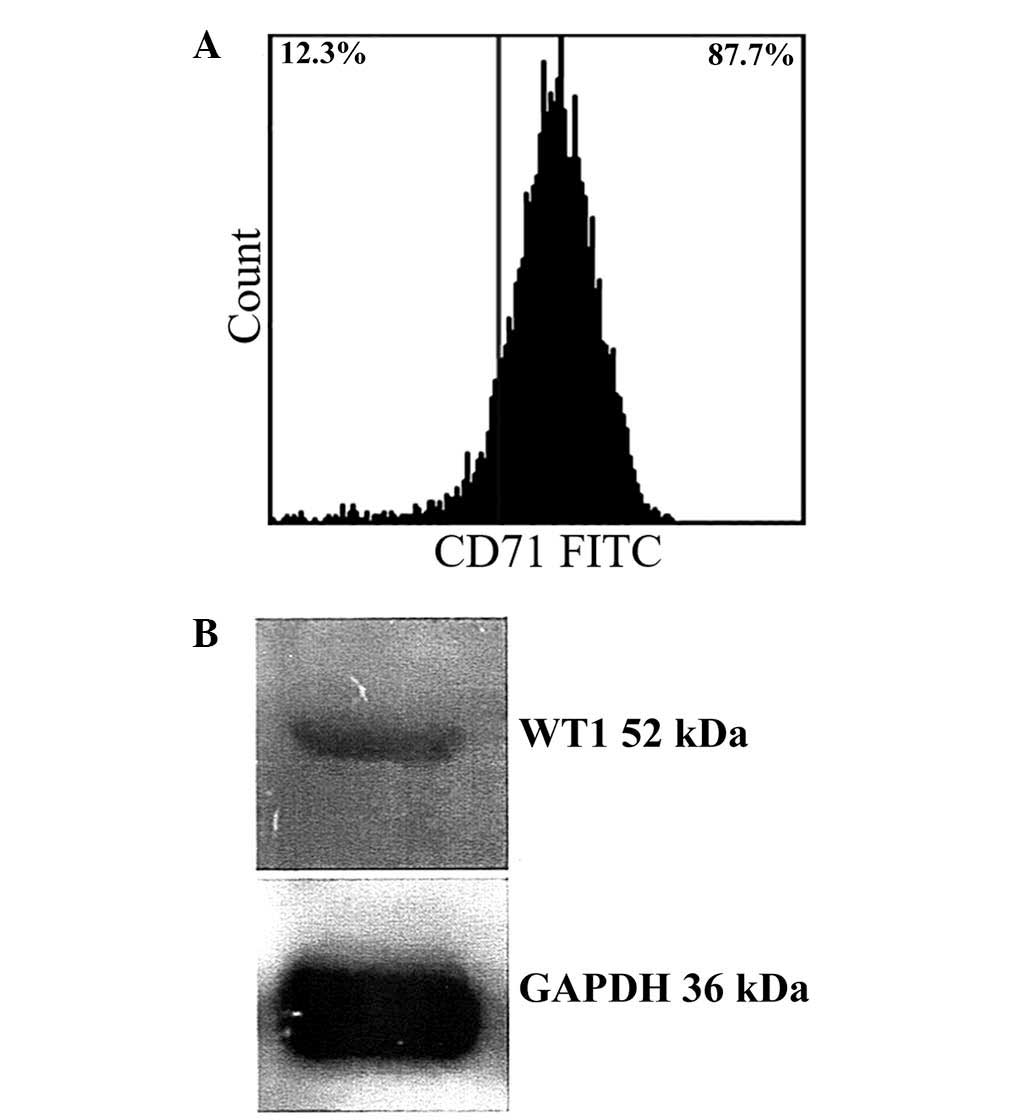
Transferrin receptor and WT1
expression in the Bl6F10 cell line
B16F10 cells showed 87% positivity for the Tf
receptor according to flow cytometry results (Fig. 2A). In addition, western blotting
confirmed expression of WT1 in B16F10 cells (Fig. 2B). Thus, B16F10 cells were selected
as a suitable model to test Tf-conjugated PEG liposomes loaded with
WT1 shRNA.
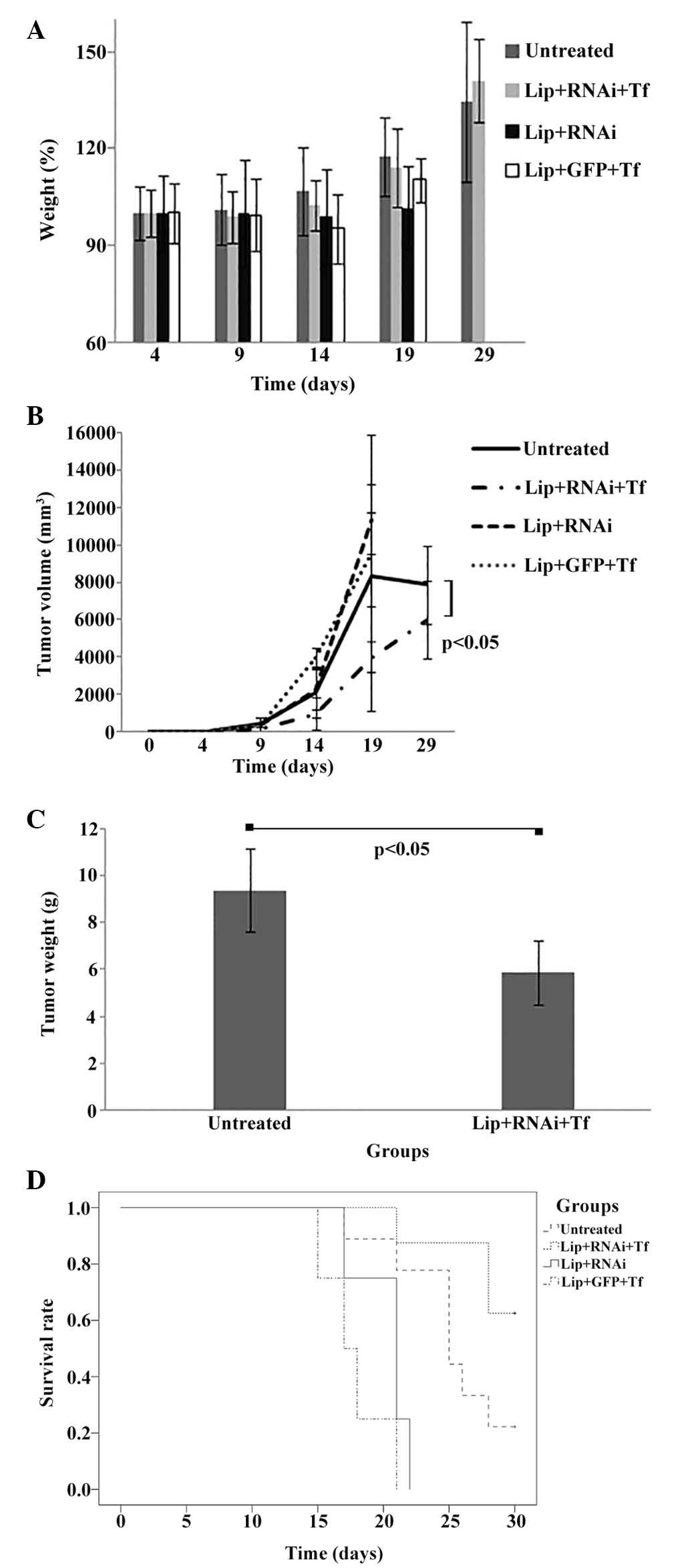
In vivo assay
Administration of different treatments Lip + RNAi +
Tf, Lip + RNAi, Lip + GFP + Tf and untreated was initiated four
days after cells were injected subcutaneously into mice. Treatments
were administered six times through intravenous tail vein injection
during a period of five days.
The mean percentage of weights of mice previously
inoculated with B16F10 cells and treated with Lip + RNAi + Tf, Lip
+ RNAi, Lip + GFP + Tf and untreated is shown in Fig. 3A. Untreated and Lip + RNAi + Tf
groups showed increases of 34% (SD±24.92) and 41% (SD±12.78)
respectively, without significant differences between groups. In
Lip + RNAi and Lip + GFP + Tf groups showed a low increase of 1.5%
(SD±12.97) and 10% (SD±6.87) respectively, against final reading on
day 19.
Fig. 3B we found that
Lip + RNAi and Lip + GFP + Tf groups grew, and there was a decrease
in survival. For this reason, we were only able to record
measurements to day 19 post-implant. The Lip + RNAi + Tf treatment
showed a significant decrease (P<0.05) in tumor volume compared
to the untreated group. The final readings presented a mean of
7,871 mm3 (SD±2,087) for the untreated group and 5,981
mm3 (SD±2,099) for Lip + RNAi + Tf group. This change
represents a reduction in tumor volume of 24%.
Fig. 3C shows the
mean final tumors weight of the groups of mice sacrificed at the
end of the trial. Only the untreated and Lip + RNAi + Tf groups
were analyzed because the mice of the other groups died after day
30. The results showed a reduction in tumor mass in the Lip + RNAi
+ Tf group and a mean tumor weight of 5.5 g (SD±0.87) compared with
the untreated group that showed to mean tumor weight of 8.8 g
(SD±0.30). The decrease of 34% in weight of the tumors collected
shows that tumor size was significantly reduced (P<0.05) in Lip
+ RNAi + Tf group.
Fig. 3D the survival
rate of C57BL/6 mice with implanted B16F10 melanoma cells
subcutaneously into the right thigh of the hind limb was 25.22 days
(SD±1.31). The untreated group presented a survival rate of 22.2%
and Lip + RNAi + Tf group showed a survival rate of 62.5%,
equivalent to an increase of 37%. This difference suggests that
treatment with Lip + RNAi + Tf significantly increases survival in
melanoma model employed (P<0.060). The Lip + RNAi and Lip + GFP
+ Tf groups showed a reduced survival compared to the untreated
group, with a mean survival estimated at 20.25 and 17.75 days,
respectively. The surviving mice were sacrificed on day 30 for
humane reasons (+ Censored groups).
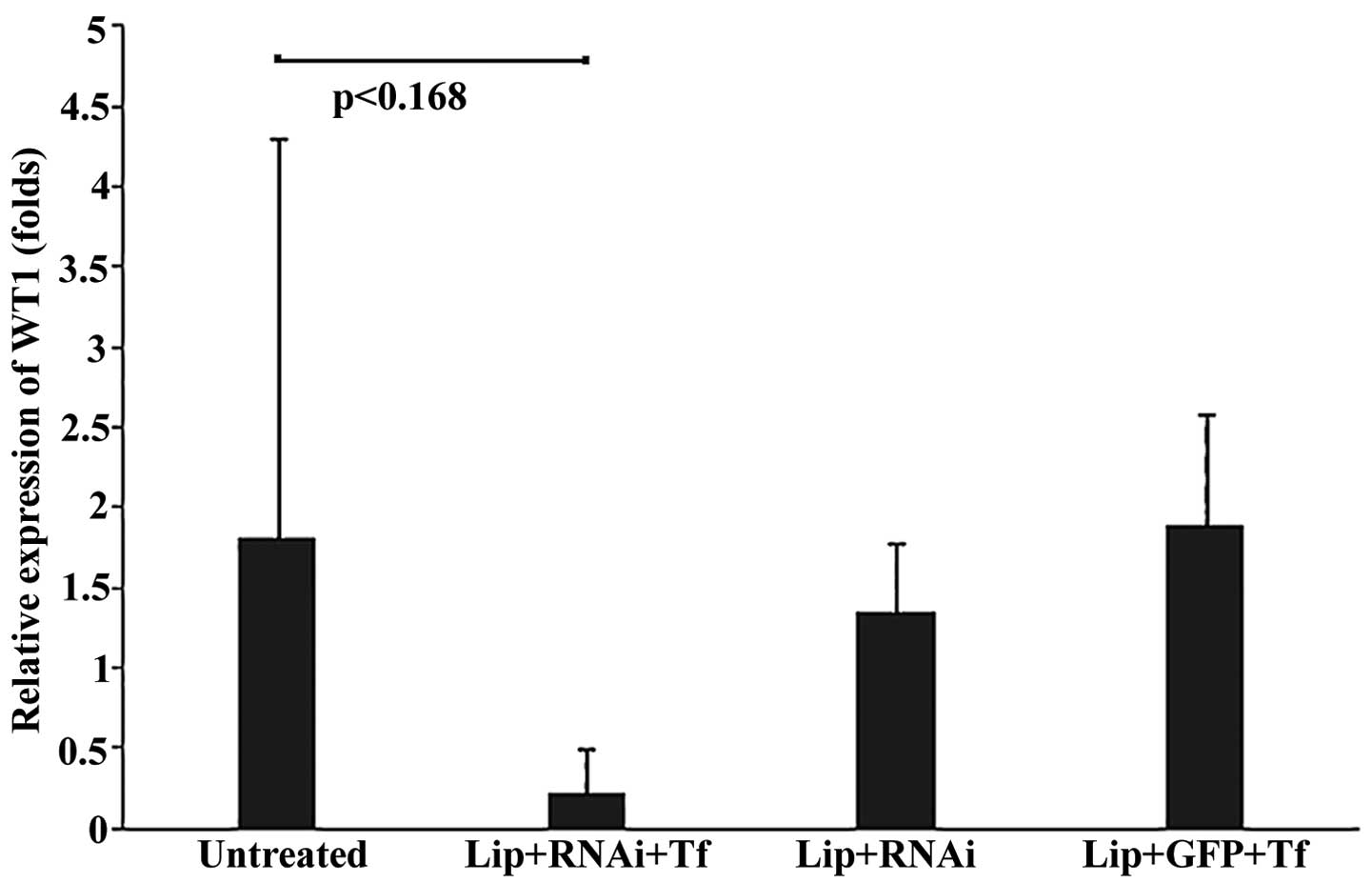
Analysis of WT1 expression in mouse
tumor tissues
The WT1 expression levels in all groups of
mice were analyzed using RT-qPCR analysis. The samples analyzed of
Lip + RNAi and Lip + GFP + Tf groups were extracted soon after
death of the mouse. In Fig. 4 show
the relative expression of WT1 standardized according to the Livak
method. To accomplish this, the average obtained from samples of
untreated group was established as a calibrator and β-actin was
used as endogenous control. Fig. 4
shows that the Lip + RNAi + Tf group showed a mean relative
expression level of 0.21 (SD±0.28)-fold, representing a decrease in
expression of WT1, compared with the other groups of mice
which presented mean relative expression levels of 1.8
(SD±2.49)-fold for the untreated group of mice, 1.34 (SD±0.43)-fold
for the Lip + RNAi group, and 1.89 (SD±0.69)-fold for the Lip + GFP
+ Tf group. The reduction in the expression levels of WT1 in
the Lip + RNAi + Tf group suggests that tumor samples decrease in
the expression of WT, which in turn suggests internalization
directed by Tf.
Discussion
During the last three decades, global incidence of
melanoma has rapidly increased, particularly among Caucasian
populations (3). The increased
mortality rate from melanoma is associated with a relatively
late-stage diagnosis and resistance to chemotherapeutic agents
(3).
The search for new therapeutic strategies has helped
identify new tumor-specific molecules suitable for gene therapy
(9). Restoration of tumor suppressor
genes, such as p53, is a major strategy used for tumor
reduction (43). Antisense and RNA
interference are additional strategies for targeting genes involved
in cancer. Well-defined tumor markers in melanoma include mutations
in the oncogenes NRAS, BRAF, c-KIT, GNAQ and GNA11 or
reduced function of the tumor suppressor genes PTEN and
p53 (44–48). Between 40 and 60% of melanomas
exhibit mutations in BRAF, and 90% have mutations in
p53, although only 10% of these mutations actually disable
the tumor suppressor activity of p53 (49). Recent studies show high expression of
wild-type WT1 in a large proportion of solid and non-solid tumors,
including melanoma (13–16,50). WT1
is a multifaceted protein involved in cell proliferation, cell
death and angiogenesis. WT1 expression is found in 39% of
melanomas, and >80% of WT1-positive melanomas is diagnosed in
advanced stages of the disease. Thus, WT1 is a potential
therapeutic target in melanoma. Antisense oligonucleotide and RNA
interference strategies that decrease the expression of WT1
decreased the proliferation of cancer cells (51–54).
One of the limitations of gene therapy is
tumor-specific delivery. The use of viral vectors is a good method
for delivering genetic material in vitro (46–48);
this strategy is limited by the host immune response in vivo
(55–57).
The main advantages of liposomes include
considerable concentration of large quantities of antitumor agent
and intracellular delivery, increased half-life of tumor agents due
to reduced degradation, gradual delivery of antineoplastic
molecules, easy removal and reduced toxicity (24).
Phospholipid derivatives are currently used to
improve circulating of liposomes and reduce their elimination by
the liver and macrophages (58). The
incorporation of thiol groups allows crosslinking with lysines in
proteins, which can yield liposomes targeted to the
ligand-receptor. The Tf receptor is widely used as delivery vehicle
targeting tumor cells (41). The Tf
receptor, CD71, is involved in the intracellular uptake of iron.
The high expression of CD71 in many tumors, including melanoma, and
the association with high iron requirements by dividing cells,
makes this a good target (59). Tf
is a serum protein found in high concentrations in the blood and is
the natural ligand for the Tf receptor. Due to the versatility of
the Tf receptor, it is useful for the delivery of transferrin
antitumor agents, such as toxins, proteins and genetic materials
(28,29,32,33,40).
In a previous study, we demonstrated the ability of
shRNA to silence the WT1 gene and induce apoptosis in B16F10
murine melanoma cells (41). In a
model of lung metastasis, application of this shRNA via aerosol was
successful in reducing the size and number of tumors (60). The delivery system was effectively
applied using PEI-DNA complexes, which reached high concentrations
in the lung; however, this form application works exclusively in
the lung (60). Systemic application
of targeted liposomes opens the possibility of targeting WT1 in
tumors in other parts of body (60).
Liposome therapy would not be exclusive for melanoma, as other
types of tumors express WT1 and the Tf receptor. The present
results showed a 24% decrease in tumor size without changes in body
weight. The final weight of the tumor for the experimental group
differed by 34% from that of the control mice (20). In addition, an improvement in
survival of 37% was observed in mice treated with Tf-conjugated PEG
liposomes loaded with WT1 shRNA. These are important
results, considering that B16F10 cells form very aggressive tumors
in C57BL/6 mice (14).
A reduction in WT1 expression was noted in
the Lip + RNAi + Tf experimental treatment group. Although
transfection efficiency was not analyzed, RT-qPCR suggests that
decreased expression of WT1 was associated with a reduction in
tumor mass. Tumors were collected within 24 h of the last
treatment. In order to identify transfected cells with decreased
expression of WT1, we would have to assess expression of
beta-galactosidase about 2–6 h after in vivo inoculation
(41).
Complete tumor eradication was not possible; thus,
further studies should examine increased concentrations of liposome
complexes or additional anticancer molecules. Previous studies have
shown that WT1 silencing sensitizes cells to
chemotherapeutic agents, such as cisplatin, doxorubicin and
radiation. Increased sensitivity to apoptotic stimuli in response
to silencing of WT1 is likely due to the fact that WT1
regulates Bcl-2, multi-drug resistance 1 (MDR1) and P-glycoprotein
expression, which are involved in chemoresistance (21,61,62).
Acknowledgements
This study was supported by the Microbiology and
immunology Department, Biological Sciences Faculty, Autonomous
University of Nuevo León (UANL).
References
|
1
|
Cichorek M, Wachulska M, Stasiewicz A and
Tymińska A: Skin melanocytes: Biology and development. Postepy
Dermatol Alergol. 30:30–41. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Situm M, Buljan M, Bulić SO and Simić D:
The mechanisms of UV radiation in the development of malignant
melanoma. Coll Antropol 31 (Suppl 1). 13–16. 2007.
|
|
3
|
Nikolaou V and Stratigos AJ: Emerging
trends in the epidemiology of melanoma. Br J Dermatol. 170:11–19.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Leiter U, Buettner PG, Eigentler TK and
Garbe C: Prognostic factors of thin cutaneous melanoma: An analysis
of the central malignant melanoma registry of the german
dermatological society. J Clin Oncol. 22:3660–3667. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Leiter U, Meier F, Schittek B and Garbe C:
The natural course of cutaneous melanoma. J Surg Oncol. 86:172–178.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee ML, Tomsu K and Von Eschen KB:
Duration of survival for disseminated malignant melanoma: Results
of a meta-analysis. Melanoma Res. 10:81–92. 2000.PubMed/NCBI
|
|
7
|
Hewitt SM, Hamada S, McDonnell TJ,
Rauscher FJ III and Saunders GF: Regulation of the proto-oncogenes
bcl-2 and c-myc by the Wilms' tumor suppressor gene WT1. Cancer
Res. 55:5386–5389. 1995.PubMed/NCBI
|
|
8
|
Englert C, Hou X, Maheswaran S, Bennett P,
Ngwu C, Re GG, Garvin AJ, Rosner MR and Haber DA: WT1 suppresses
synthesis of the epidermal growth factor receptor and induces
apoptosis. EMBO J. 14:4662–4675. 1995.PubMed/NCBI
|
|
9
|
Maheswaran S, Englert C, Bennett P,
Heinrich G and Haber DA: The WT1 gene product stabilizes p53 and
inhibits p53-mediated apoptosis. Genes Dev. 9:2143–2156. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Haber DA, Englert C and Maheswaran S:
Functional properties of WT1. Med Pediatr Oncol. 27:453–455. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Englert C, Maheswaran S, Garvin AJ,
Kreidberg J and Haber DA: Induction of p21 by the Wilms' tumor
suppressor gene WT1. Cancer Res. 57:1429–1434. 1997.PubMed/NCBI
|
|
12
|
Scholz H and Kirschner KM: A role for the
Wilms tumor Protein WT1 in organ development. Physiology
(Bethesda). 20:54–59. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Oji Y, Miyoshi S, Maeda H, Hayashi S,
Tamaki H, Nakatsuka S, Yao M, Takahashi E, Nakano Y, Hirabayashi H,
et al: Overexpression of the Wilms' tumor gene WT1 in de novo lung
cancers. Int J Cancer. 100:297–303. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Loeb DM, Evron E, Patel CB, Sharma PM,
Niranjan B, Buluwela L, Weitzman SA, Korz D and Sukumar S: Wilms'
tumor suppressor gene (WT1) is expressed inprimary breast tumors
despite tumor-specific promoter methylation. Cancer Res.
61:921–925. 2001.PubMed/NCBI
|
|
15
|
Miyoshi Y, Ando A, Egawa C, Taguchi T,
Tamaki Y, Tamaki H, Sugiyama H and Noguchi S: High expression of
Wilms' tumor suppressor gene predicts poor prognosis in breast
cancer patients. Clin Cancer Res. 8:1167–1171. 2002.PubMed/NCBI
|
|
16
|
Oji Y, Miyoshi Y, Koga S, Nakano Y, Ando
A, Nakatsuka S, Ikeba A, Takahashi E, Sakaguchi N, Yokota A, et al:
Overexpression of the Wilms' tumor gene WT1 in primary thyroid
cancer. Cancer Sci. 94:606–611. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wagner N, Panelos J, Massi D and Wagner
KD: The Wilms' tumor suppressor WT1 is associated with melanoma
proliferation. Pflugers Arch. 455:839–847. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Garrido-Ruiz MC, Rodriguez-Pinilla SM,
Pérez-Gómez B and Rodriguez-Peralto JL: WT 1 expression in nevi and
melanomas: A marker of melanocytic invasion into the dermis. J
Cutan Pathol. 37:542–548. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wilsher M and Cheerala B: WT1 as a
complementary marker of malignant melanoma: An immunohistochemical
study of whole sections. Histopathology. 51:605–610. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zamora-Avila DE, Franco-Molina MA,
Trejo-Avila LM, Rodríguez-Padilla C, Reséndez-Pérez D and
Zapata-Benavides P: RNAi silencing of the WT1 gene inhibits cell
proliferation and induces apoptosis in the B16F10 murine melanoma
cell line. Melanoma Res. 17:341–348. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zapata-Benavides P, Manilla-Muñoz E,
Zamora-Avila DE, Saavedra-Alonso S, Franco-Molina MA, Trejo-Avila
LM, Davalos-Aranda G and Rodríguez-Padilla C: WT1 silencing by RNAi
synergizes with chemotherapeutic agents and induces
chemosensitization to doxorubicin and cisplatin in B16F10 murine
melanoma cells. Oncol Lett. 3:751–755. 2012.PubMed/NCBI
|
|
22
|
Yu KF, Zhang WQ, Luo LM, Song P, Li D, Du
R, Ren W, Huang D, Lu WL, Zhang X and Zhang Q: The antitumor
activity of a doxorubicin loaded, iRGD-modified
sterically-stabilized liposome on B16-F10 melanoma cells: In vitro
and in vivo evaluation. Int J Nanomedicine. 8:2473–2485.
2013.PubMed/NCBI
|
|
23
|
Maruyama K, Ishida O, Kasaoka S, Takizawa
T, Utoguchi N, Shinohara A, Chiba M, Kobayashi H, Eriguchi M and
Yanagie H: Intracellular targeting of sodium
mercaptoundecahydrododecaborate (BSH) to solid tumors by
transferrin-PEG liposomes, for boron neutron-capture therapy
(BNCT). J Control Release. 98:195–207. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Torchilin V: Tumor delivery of
macromolecular drugs based on the EPR effect. Adv Drug Deliv Rev.
63:131–135. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Boerman OC, Oyen WJ, van Bloois L,
Koenders EB, van der Meer JW, Corstens FH and Storm G: Optimization
of technetium-99m-labeled PEG liposomes to image focal infection:
Effects of particle size and circulation time. J Nucl Med.
38:489–493. 1997.PubMed/NCBI
|
|
26
|
Derycke AS and De Witte PA:
Transferrin-mediated targeting of hypericin embedded in sterically
stabilized PEG-liposomes. Int J Oncol. 20:181–187. 2002.PubMed/NCBI
|
|
27
|
Papahadjopoulos D, Allen TM, Gabizon A,
Mayhew E, Matthay K, Huang SK, Lee KD, Woodle MC, Lasic DD,
Redemann C, et al: Sterically stabilized liposomes: Improvements in
pharmacokinetics and antitumor therapeutic efficacy. Proc Natl Acad
Sci USA. 88:11460–11464. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stavridis JC, Deliconstantinos G,
Psallidopoulos MC, Armenakas NA, Hadjiminas DJ and Hadjiminas J:
Construction of transferrin-coated liposomes for in vivo transport
of exogenous DNA to bone marrow erythroblasts in rabbits. Exp Cell
Res. 164:568–572. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Vidal M, Sainte-Marie J, Philippot JR and
Bienvenue A: The influence of coupling transferrin to liposomes or
minibeads on its uptake and fate in leukemic L2C cells. FEBS Lett.
216:159–163. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Di Giulio A, D'Andrea G, Saletti MA,
Impagnatiello A, D'Alessandro AM and Oratore A: The binding of
human serum transferrin to its specific receptor reconstituted into
liposomes. Cell Signal. 6:83–90. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Egea MA, García ML, Alsina MA and Reig F:
Coating of liposomes with transferrin: Physicochemical study of the
transferrin-lipid system. J Pharm Sci. 83:169–173. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Corley P and Loughrey HC: Targeting of
doxorubicin loaded liposomes to T-cells via the transferrin
receptor. Biochem Soc Trans. 26:S371998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Simões S, Slepushkin V, Gaspar R, de Lima
MC and Düzgüneş N: Gene delivery by negatively charged ternary
complexes of DNA, cationic liposomes and transferrin or fusigenic
peptides. Gene Ther. 5:955–964. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Singh M: Transferrin as a targeting ligand
for liposomes and anticancer drugs. Curr Pharm Des. 5:443–451.
1999.PubMed/NCBI
|
|
35
|
Eavarone DA, Yu X and Bellamkonda RV:
Targeted drug delivery to C6 glioma by transferrin-coupled
liposomes. J Biomed Mater Res. 51:10–14. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ishida O, Maruyama K, Tanahashi H,
Iwatsuru M, Sasaki K, Eriguchi M and Yanagie H: Liposomes bearing
polyethyleneglycol-coupled transferrin with intracellular targeting
property to the solid tumors in vivo. Pharm Res. 18:1042–1048.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Voinea M, Dragomir E, Manduteanu I and
Simionescu M: Binding and uptake of transferrin-bound liposomes
targeted to transferrin receptors of endothelial cells. Vascul
Pharmacol. 39:13–20. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Anabousi S, Laue M, Lehr CM, Bakowsky U
and Ehrhardt C: Assessing transferrin modification of liposomes by
atomic force microscopy and transmission electron microscopy. Eur J
Pharm Biopharm. 60:295–303. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sharma G, Modgil A, Layek B, Arora K, Sun
C, Law B and Singh J: Cell penetrating peptide tethered bi-ligand
liposomes for delivery to brain in vivo: Biodistribution and
transfection. J Control Release. 167:1–10. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kobayashi T, Ishida T, Okada Y, Ise S,
Harashima H and Kiwada H: Effect of transferrin receptor-targeted
liposomal doxorubicin in P-glycoprotein-mediated drug resistant
tumor cells. Int J Pharm. 329:94–102. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shi N and Pardridge WM: Noninvasive gene
targeting to the brain. Proc Natl Acad Sci USA. 97:7567–7572. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C (T)) Method. Methods. 25:402–418. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xu L, Pirollo KF and Chang EH:
Tumor-targeted p53-gene therapy enhances the efficacy of
conventional chemo/radiotherapy. J Control Release. 74:115–128.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Carlino MS, Haydu LE, Kakavand H, Menzies
AM, Hamilton AL, Yu B, Ng CC, Cooper WA, Thompson JF, Kefford RF,
et al: Correlation of BRAF and NRAS mutation status with outcome,
site of distant metastasis and response to chemotherapy in
metastatic melanoma. Br J Cancer. 111:292–299. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pilloni L, Bianco P, Difelice E, Cabras S,
Castellanos ME, Atzori L, Ferreli C, Mulas P, Nemolato S and Faa G:
The usefulness of c-Kit in the immunohistochemical assessment of
melanocytic lesions. Eur J Histochem. 55:105–111. 2011. View Article : Google Scholar
|
|
46
|
Khalili JS, Yu X, Wang J, Hayes BC, Davies
MA, Lizee G, Esmaeli B and Woodman SE: Combination small molecule
MEK and PI3K inhibition enhances uveal melanoma cell death in a
mutant GNAQ and GNA11 dependent manner. Clin Cancer Res.
18:4345–4355. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou X-P, Gimm O, Hampel H, Niemann T,
Walker MJ and Eng C: Epigenetic PTEN silencing in Malignant
Melanomas without PTEN Mutation. Am J Pathol. 157:1123–1129. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Terzian T, Torchia EC, Dai D, Robinson SE,
Murao K, Stiegmann RA, Gonzalez V, Boyle GM, Powell MB, Pollock PM,
et al: p53 prevents progression of nevi to melanoma predominantly
through cellcycle regulation. Pigment Cell Melanoma Res.
23:781–794. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Box NF, Vukmer TO and Terzian T: Targeting
p53 in melanoma. Pigment Cell Melanoma Res. 27:8–10. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Miyagi T, Ahuja H, Kubota T, Kubonishi I,
Koeffler HP and Miyoshi I: Expression of the candidate Wilm's tumor
gene, WT1, in human leukemia cells. Leukemia. 7:970–977.
1993.PubMed/NCBI
|
|
51
|
Zapata-Benavides P, Tuna M,
Lopez-Berestein G and Tari AM: Downregulation of Wilms' tumor 1
protein inhibits breast cancer proliferation. Biochem Biophys Res
Commun. 295:784–790. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chen MY, Clark AJ, Chan DC, Ware JL, Holt
SE, Chidambaram A, Fillmore HL and Broaddus WC: Wilms' tumor 1
silencing decreases the viability and chemoresistance of
glioblastoma cells in vitro: A potential role for IGF-1R
de-repression. J Neurooncol. 103:87–102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Huo X, Ren L, Shang L, Wang X and Wang J:
Effect of WT1 antisense mRNA on the induction of apoptosis in
ovarian carcinoma SKOV3 cells. Eur J Gynaecol Oncol. 32:651–656.
2011.PubMed/NCBI
|
|
54
|
Chen Y, Bathula SR, Yang Q and Huang L:
Targeted nanoparticles deliver siRNA to melanoma. J Invest
Dermatol. 130:2790–2798. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhang X, Jin Y, Zhang B and Chen H:
Preparation of recombinant adenovirus Ad5/F35 containing human WT1
and identification after recombinant adenovirus infected dendritic
cells. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 30:135–138. 2014.(In
Chinese). PubMed/NCBI
|
|
56
|
Sundarasetty BS, Singh VK, Salguero G,
Geffers R, Rickmann M, Macke L, Borchers S, Figueiredo C, Schambach
A, Gullberg U, et al: Lentivirus-induced dendritic cells for
immunization against high-risk WT1(+) acute myeloid leukemia. Hum
Gene Ther. 24:220–237. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Shan XY, Liu ZL, Wang B, Guo GX, Wang MS,
Zhuang FL, Cai CS, Zhang MF and Zhang YD: Construction of
recombinant lentiviral vector of Tie2-RNAi and its influence on
malignant melanoma cells in vitro. Zhonghua Zheng Xing Wai Ke Za
Zhi. 27:277–283. 2011.(In Chinese). PubMed/NCBI
|
|
58
|
Drummond DC, Meyer O, Hong K, Kirpotin DB
and Papahadjopoulos D: Optimizing liposomes for delivery of
chemotherapeutic agents to solid tumors. Pharmacol Rev. 51:691–743.
1999.PubMed/NCBI
|
|
59
|
Cheever MA, Allison JP, Ferris AS, Finn
OJ, Hastings BM, Hecht TT, Mellman I, Prindiville SA, Viner JL,
Weiner LM and Matrisian LM: The prioritization of cancer antigen: A
national cancer institute pilot project for the acceleration of
translational research. Clin Cancer Res. 15:5323–5337. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zamora-Avila DE, Zapata-Benavides P,
Franco-Molina MA, Saavedra-Alonso S, Trejo-Avila LM, Reséndez-Pérez
D, Méndez-Vázquez JL, Isaias-Badillo J and Rodríguez-Padilla C: WT1
gene silencing by aerosol delivery of PEI:RNAi complexes inhibits
B16-F10 lung metastases growth. Cancer Gene Ther. 16:892–899. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Clark AJ, Chan DC, Chen MY, Fillmore H,
Dos Santos WG, Van Meter TE, Graf MR and Broaddus WC:
Down-regulation of Wilms' tumor 1 expression in glioblastoma cells
increases radiosensitivity independently of p53. J Neurooncol.
83:163–172. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Shen H, Xu W, Wu Z, Tang H, Xie Y and
Zhong X: Down-regulation of WT1/+17AA gene expression using RNAi
and modulating leukemia cell chemotherapy resistance.
Haematologica. 92:1270–1272. 2007. View Article : Google Scholar : PubMed/NCBI
|