Introduction
Vascular endothelial growth factor (VEGF) is
essential for tumor neovascularization, which leads to the
formation of blood vessels within a tumor and facilitates cancer
cell survival, local tumor growth and the development of distant
metastases (1). VEGF is also a
primary mediator of angiogenesis and vascular leakage in exudative
age-related macular degeneration (AMD) (2,3).
Additionally, VEGF is an autocrine survival factor for a number of
cell types and a critical factor for the retina during development
and maturity (4). VEGF binds to two
distinct receptors, VEGF receptor 1 (VEGFR1) and VEGF receptor 2
(VEGFR2); however, VEGFR2 is considered to be the dominant
signaling receptor for endothelial cell permeability,
differentiation and proliferation (5,6).
Wet AMD is characterized by the presence of
choroidal neovascularization (CNV) in which newly formed blood
vessels from choriocapillaris penetrate through Bruch's membrane
(BM) and retinal pigment epithelial (RPE) cells (7,8). Wet AMD
accounts for <10% of AMD, but serious complications may arise if
it is left untreated, leading to the development of retinal
detachment and blindness (9,10). Elevated levels of VEGF were
identified in vitreous samples from patients with retinal
neovascular diseases, indicating that VEGF serves a role in retinal
disorders (11). RPE cells, which
are located between the choroid and retina, serve a critical role
in the pathogenesis of AMD (12).
RPE cells also secrete various cytokines, chemokines and growth
factors, including interleukin (IL)-6, IL-11, chemokine (C-X-C
motif) ligand 9, platelet-derived growth factor, transforming
growth factor-β (TGF-β) and VEGF (13). Hypoxia results in the increased
expression of VEGF and IL-6, which are associated with increased
angiogenesis, cell migration and invasion (14). It has been demonstrated that TGF-β
and VEGF markedly upregulate Snail mRNA, which is associated with
the epithelial-mesenchymal transition (EMT), while no cytokines are
known to affect the expression of transcriptional factors
associated with EMT in adult retinal pigment epithelium-19 (ARPE19)
cells (15). Based on these results,
current treatments for AMD primarily focus on anti-VEGF therapies
due to the association of CNV and EMT with VEGF (16,17).
Vandetanib is a small molecule kinase inhibitor that
inhibits the tyrosine kinase activity of VEGFR2, as well as
angiogenesis (18). Vandetanib
regulates tumor growth and vasculature in lung cancer by inhibiting
epidermal growth factor receptor (EGFR) and VEGFR2 when
administered as a single agent or used in combination with
chemotherapy (19,20). Vandetanib inhibits not only growth
factor-induced phosphorylation of EGFR, mitogen-activated protein
kinase (MAPK), VEGFR2 and protein kinase B (Akt), but also colony
formation and invasion of breast cancer cells (21). Constitutive VEGF expression and
secretion decreases following p38 inhibition, while no clear effect
is observed following the inhibition of extracellular
signal-regulated kinase (ERK) or c-Jun N-terminal kinase (JNK)
(22). However, there is no specific
information regarding the use of vandetanib in preclinical or
clinical ocular pathological conditions.
A disintegrin and metalloproteinase (ADAM) 10 and
ADAM17 proteins are widely expressed in the differential layers of
the retina throughout the embryonic period and are essential for a
number of biological processes. These include cell fate
determination, angiogenesis, cell migration and wound healing
(23–25). VEGF enhances VEGFR2 shedding via
ADAM17, which is activated by the ERK and MAPK pathways (26). EBV-infected human corneal epithelial
cells (HCECs) have previously been generated, which cause EMT by
enhancing the secretion of IL-6, IL-8, VEGF, TGF-β1, tumor necrosis
factor (TNF)-α and monocyte chemoattractant protein-1, as well as
phosphoinositide 3-kinase (PI3K)/Akt and ERK signaling activation
(27). EMT is a characteristic
phenotypical change observed in patients with proliferative
vitreoretinopathy (PVR) (28). RPE
cells are crucial for the formation of fibrous tissue on the
detached retina and EMT in PVR (29); however, to the best of our knowledge,
there is currently no specific model for studying the effect of
vandetanib on ADAM expression and cellular regulatory mechanism in
RPE cells. In the present study, EBV-infected RPE cells
(ARPE19/EBV) were produced as an in vitro model of
EMT-related retinopathy, which expressed the mesenchymal
phenotypes. The effect of vandetanib on ADAM expression and EMT
using ARPE19/EBV that secreted VEGF and the signaling pathway that
is involved in EBV-infected ARPE cells as a model of CNV or PVR,
was investigated to assess their application in a clinical
setting.
Materials and methods
Cell culture and reagents
Cells from the human retinal pigment epithelial cell
line ARPE19 were purchased from the American Type Culture
Collection (ATCC; Manassas, VA, USA). The cells were maintained in
Dulbecco's modified Eagle's medium/F12 (DMEM/F12; HyClone; Logan,
UT, USA) supplemented with 10% fetal bovine serum (FBS; HyClone; GE
Healthcare Life Sciences) and penicillin (100 U/ml)-streptomycin
(100 µg/ml; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) under a humidified atmosphere with 5% CO2. For the
generation of EBV-infected ARPE19 cells, cell-free EBV virions were
prepared from the B95-8 cell line (EBV type I; ATCC), as previously
described (27). Briefly, the B95-8
cells were grown in RPMI-1640 medium (HyClone; GE Healthcare Life
Sciences) supplemented with streptomycin, glutamine, and 10% FBS
for 48 h at 37°C in an atmosphere containing 5% CO2. The
infectious culture supernatant was harvested, centrifuged (200 ×
g for 10 min at 25°C), and filtered using a 0.2-µm-pore
filter (Corning Inc., Corning, NY, USA) to remove cell debris.
Following the attachment of the ARPE19 cells (2×105
cells/T25 flask/4 ml media), an equal volume of EBV supernatant (4
ml, 73±11 CFU/ml) was added. EBV infection was confirmed using
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and immunoblotting to detect viral transcripts and
proteins, at four weeks following infection with EBV. Human
recombinant VEGF was purchased from Human Biosciences, Inc.
(Gaithersburg, MD, USA). Vandetanib was purchased from
SelleckChemicals (Houston, TX, USA). GI254023X, Marimastat and
ONO4817 were purchased from Tocris Bioscience (Bristol, UK). The
Ethics committee and Institutional Review Board of the College of
Medicine, Inje University (Busan, Korea) approved all protocols and
procedures used in the present study.
Cell viability assay
The effect of vandetanib on the cell viability of
ARPE19 and ARPE19/EBV cells was evaluated using an MTT assay (Sigma
Aldrich; Merck KGaA, Darmstadt, Germany). The cells were seeded at
a density of 1–2×104 cells/well in a 96-well plate and
treated with various concentrations (10, 50, 100, and 500 nM) of
vandetanib. Following 48 and 72 h incubation at 37°C, 30 µl MTT
solution (1 mg/ml) was added to each well. The formazan dye, part
of the MTT assay, was dissolved with dimethyl sulfoxide and the
optical density (OD) was measured using a Synergy™ HT
Multi-Detection Microplate Reader (Bio-Tek instruments Inc.,
Winooski, VT, USA) at 570 nm. Cell viability (%) was calculated as
follows:
[OD(vandetanib)-OD(blank)]/[OD(control)-OD(blank)]
×100.
Wound-healing assay
The migration ability of the ARPE19/EBV cells was
analyzed using a wound-healing assay. The cells (3×105/2
ml) were seeded into a 6-well plate. After incubation for 24 h at
37°C, the confluent cell monolayers were manually scratched with a
200 µl micropipette tip to create a cell-free area. The plates were
washed with phosphate-buffered saline (PBS) to remove all cell
debris and were then incubated with DMEM/F12 containing 10% FBS in
the presence or absence of vandetanib. Following 16 h treatment,
repopulation of the wounded areas was observed under an inverted
phase contrast microscope and the cells that migrated to the
wounded area were imaged at ×100 magnification. The number of cells
that migrated into the scratched area was analyzed using ImageJ
software (National Institutes of Health, Bethesda, MD, USA).
RNA isolation and RT-qPCR
analysis
Total RNA was prepared using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Briefly, the treated cells were harvested
and washed twice with PBS following exposure to 1 ml TRIzol and 0.2
ml chloroform. Samples were vigorously vortexed for 15 sec and
incubated at room temperature for 2–3 min. Following centrifugation
(12,000 × g for 15 min at 4°C), the colorless upper aqueous
phase was separated, mixed with 0.5 ml of isopropyl alcohol, and
centrifuged (12,000 × g for 10 min at 4°C). Following
removal of the supernatant, the RNA pellet was washed the twice
with 1 ml 75% ethanol and dried using a vacuum for 10 min at room
temperature. For further removal of residual DNA, dissolved RNA
samples in diethylpyrocarbonate-treated water were mixed with 2.5
µl DNase I stock solution (no. 79254; Qiagen GmbH, Hilden, Germany)
and incubated for 10 min at 25°C. Purity of RNA samples
(A260/A280 ratio >1.6) was monitored with
a Nanodrop 1000 spectrophotometer (Thermo Fisher Scientific, Inc.).
Each RNA sample was transcribed into cDNA using oligo (dT) (no.
N-7053; Bioneer Corporation, Daejeon, Korea) and AccuPower RT
Premix (no. K-2043; 10 mM dNTPs; 2.5 mM, each dNTP; final
concentration, 0.5 mM; Bioneer Corporation), oligo (dT)(5 µM),
RNase inhibitor (no. 2313A; 10 U/µl; Takara Bio, Inc., Otsu,
Japan), and reverse transcriptase (100 U/µl; Bioneer Corporation).
Following gentle agitation, the mixture was incubated at 42°C for 1
h, and then heated at 94°C for 5 min to inactivate the reverse
transcriptase. PCR was performed using Prime Taq Premix with Prime
Taq DNA Polymerase, 1 unit/10 µl; 2X reaction buffer; 4 mM MgCl2;
enzyme stabilizer; sediment; loading dye (pH 9.0) and 0.5 mM each
of dATP, dCTP, dGTP, dTTP (no. G-3002; GeNet Bio, Daejeon, Korea)
with the specified primers (Table I)
and TaKaRa PCR Thermal Cycler Dice (no. TP600; Takara Bio, Inc.).
The annealing temperature for specific target sequences and cycling
conditions are provided in Table I.
β-actin was used as a housekeeping gene. PCR products were analyzed
by agarose gel (1%) electrophoresis and visualized using ethidium
bromide under ultraviolet light using the multiple Gel DOC system
(Fujifilm, Tokyo, Japan). Each experiment was performed at least
three times. Results are representative of three independent
experiments.
 | Table I.Primer sequences used for reverse
transcriptase-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcriptase-quantitative polymerase chain reaction.
|
| Primers
(5′-3′) |
|
|---|
|
|
|
|
|---|
| Target | Forward | Reverse | Temp/cycles |
|---|
| VEGFR2 |
GTGACCAACATGGAGTCGTG |
TGCTTCACAGAAGACCATGC | 62°C/30 |
| VEGF |
AGGAGGGCAGAATCATCACG |
CAAGGCCCACAGGGATTTTCT | 66°C/30 |
| EGFR |
AGGCACGAGTAACAAGCTCAC |
ATGAGGACATAACCAGCCACC | 65°C/25 |
| EGF |
CAGTTCCCACCACTTCAGGT |
GCCAGCTGCACAAATACAGA | 58°C/25 |
| E-cadherin |
GACGCGGACGATGATGTGAAC |
TTGTACTGTTGTGGATTGAAG | 50°C/30 |
| N-cadherin |
CACCCAACATGTTTACAATCAACAATGAGAC |
CTGCAGCAACAGTAAGGACAAACATCCTATT | 60°C/30 |
| Vimentin |
GGAAGAGAACTTTGCCGTTGAA |
GTGACGAGCCATTTCCTCCTT | 60°C/30 |
| Snail |
CAGATGAGGACAGTGGGAAAGG |
ACTCTTGGTGCTTGTGGAGCAG | 66°C/30 |
| α-SMA |
ATCACCATCGGAAATGAACG |
CTGGAAGGTGGACAGAGAGG | 60°C/28 |
| β-actin |
CAGGCACCAGGGCGT |
ATGGCTGGGGTGTTGAAG | 60°C/25 |
Western blot analysis
The cells were washed with PBS and lysed in NP-40
cell lysis buffer (Thermo Fisher Scientific, Inc.) supplemented
with a protease inhibitor cocktail (AEBSF, Aprotinin, bestatin
hydrochloride, E-64, EDTA, and leupeptin hemisulfate salt; no.
P8340; Sigma-Aldrich; Merck KGaA). To assess phosphorylation
events, an additional set of phosphatase inhibitor Cocktail II
(sodium orthovanadate, sodium molybdata, sodium tartrate, and
imidazole; no. P5726; Sigma-Aldrich; Merck KGaA) was added to the
NP-40 buffer. Equal quantities of protein (10 µg/lane) were
separated using 8–12% SDS-PAGE, then transferred to nitrocellulose
membranes (Merck Millipore; Merck KGaA) at 340 mA for 2 h.
Following blocking with 5% nonfat skim milk for 1 h at room
temperature, the membrane was probed with primary antibodies
against Epstein-Barr virus nuclear antigen (EBNA) 2 (no. sc-17501;
1:100), EBNA3A (no. sc-23533; 1:100), latent membrane protein (LMP)
2A (no. sc-101314; 1:100), VEGFR2 (no. sc-6251; 1:500), ADAM10 (no.
sc-25578; 1:500), ADAM12 (no. sc-25579; 1:500), ADAM17 (no.
sc-25782; 1:500), E-cadherin (no. sc-7870; 1:500), β-actin (no.
sc-47778; 1:1,000; all Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA); EGFR (no. 2232; 1:1,000), E-cadherin (no. 3195; 1:1,000),
N-cadherin (no. 13116; 1:1000), Vimentin (no. 5741; 1:1,000), Snail
(no. 3879; 1:1,000), phospho-Akt (Ser473; no. 9271; 1:1,000), Akt
(no. 9272; 1:1,000), phospho-ERK1/2 (Thr202/Tyr204; no. 9101;
1:1,000), ERK1/2 (no. 9102; 1:1,000), phospho-JNK (Thr183/Tyr185;
no. 4671; 1:1,000), JNK (no. 9258; 1:1,000), phospho-p38-MAPK
(Thr180/Tyr182; no. 9211; 1:1,000), p38-MAPK (no. 9212; 1:1,000;
all Cell Signaling Technology, Inc., Danvers, MA, USA); paired box
protein (PAX) 2 (no. PAB24790; 1:500), LMP1 (no. MAB9772; 1:2,000;
both Abnova, Taipei, Taiwan); EBNA1 (no. MA1-7271; 1:50; Thermo
Fisher Scientific, Inc.); and α-smooth muscle actin (α-SMA; no.
bs-10196R; 1:1,000; Bioss, Inc., Woburn, MA, USA), followed by the
following specific secondary antibodies: anti-mouse-horseradish
peroxidase (HRP; no. K0211589; 1:3,000) or anti-rabbit-HRP (no.
K0211708; 1:3,000; KOMABiotech, Seoul, Korea). Chemiluminescence
was detected using a WesternBright electrochemiluminescence HRP
substrate (no. K-12045-D50; Advansta, Inc., Menlo Park, CA, USA)
and a Gel Doc system (Fujifilm). Each experiment was performed at
least three times. Results are representative of three independent
experiments. Quantifications of western blots were analyzed using
ImageJ 1.38 software. Relative intensity of bands was calculated by
Image J and expressed as relative values to β-actin.
Immunofluorescence assay using
confocal microscopy
The cells (3×105/ml) were seeded and
treated with vandetanib and ADAM inhibitors. Following washing with
PBS, the cells were fixed in 4% methanol-free formaldehyde solution
(pH 7.4) at 4°C for 10 min, permeabilized with 0.1% saponin in PBS
for 10 min and blocked in blocking buffer at room temperature (5%
bovine serum albumin, 0.5% Tween-20 in PBS; no. A2058;
Sigma-Aldrich; Merck KGaA) for 1 h. The cells were then incubated
with anti-E-cadherin antibody (no. sc-7870; 1:500; Santa Cruz
Biotechnology, Inc.) for 24 h at room temperature, washed with PBS
and incubated with the fluorescein isothiocyanate-labeled secondary
antibody (no. F0382; 1:80; Sigma-Aldrich; Merck KGaA) for 1 h at
room temperature. The cells were stained with a propidium iodide
solution, mounted and visualized under a Zeiss LSM 510 Meta
confocal laser-scanning microscope (Carl Zeiss AG, Oberkochen,
Germany).
Statistical analysis
All data are presented as the mean ± standard
deviation from ≥3 independent experiments. All statistics were
calculated with the Student's t-test using SigmaPlot
software (version 10.0; Systat Software, Inc., San Jose, CA, USA)
and P<0.05 was considered to represented a statistically
significant difference.
Results
Cell characterization based on the
expression of EMT-related molecules
The present study aimed to determine whether
ARPE19/EBV cells exhibit fibroblast-like morphology and express
mesenchymal cell markers. The ARPE19/EBV cells developed a
characteristic small and spindle-like mesenchymal cell shape, and
stably expressed the EBV-related viral proteins, including
Epstein-Barr virus nuclear antigen EBNA1, EBNA2, EBNA3A, LMP1 and
LMP2A (Fig. 1A). It was then
investigated whether ARPE19/EBV cells exhibited EMT-related
molecules, including mRNA and protein. Although EGF and VEGF mRNA
were detected in the ARPE19 cells ARPE19/EBV cells, the levels of
EGF, VEGF, EGFR and VEGFR2 mRNA were markedly increased in
ARPE19/EBV cells (Fig. 1B).
Increased expression of EGFR and VEGFR2 proteins were also detected
in the ARPE19/EBV cells compared with the ARPE19 cells (Fig. 1C). ARPE19/EBV cells exhibited
mesenchymal-like cell features, including upregulated expression of
vimentin, Snail and α-SMA mRNA and protein. The cells also
exhibited decreased E-cadherin and N-cadherin expression consistent
with the loss of epithelial characteristics (Fig. 1B and C). In the ARPE19/EBV cells, the
expression of ADAM10 and ADAM17 was upregulated compared with
ARPE19 cells (Fig. 1C), however
ADAM12 expression was not. Compared with the ARPE19 cells,
increased expression of paired bow gene 2 (PAX2) protein, a
regulatory transcriptional factor for ADAM10, was observed in the
ARPE19/EBV cells (Fig. 1C). It was
them determined whether exposure to vandetanib had an effect on
cell viability of the ARPE19 and ARPE19/EBV cells. Although
ARPE19/EBV cells were more sensitive to vandetanib, following 48 h
vandetanib (500 nM) treatment, both drug-treated ARPE19 and
ARPE19/EBV cells had a survival rate >80%. However, the survival
rate of vandetanib-treated ARPE19/EBV slightly decreased at 72 h
(Fig. 1D). Based on these findings,
the effect of vandetanib on ARPE19/EBV as a model of CNV to develop
new treatment methods for retinal pathological conditions was
evaluated.
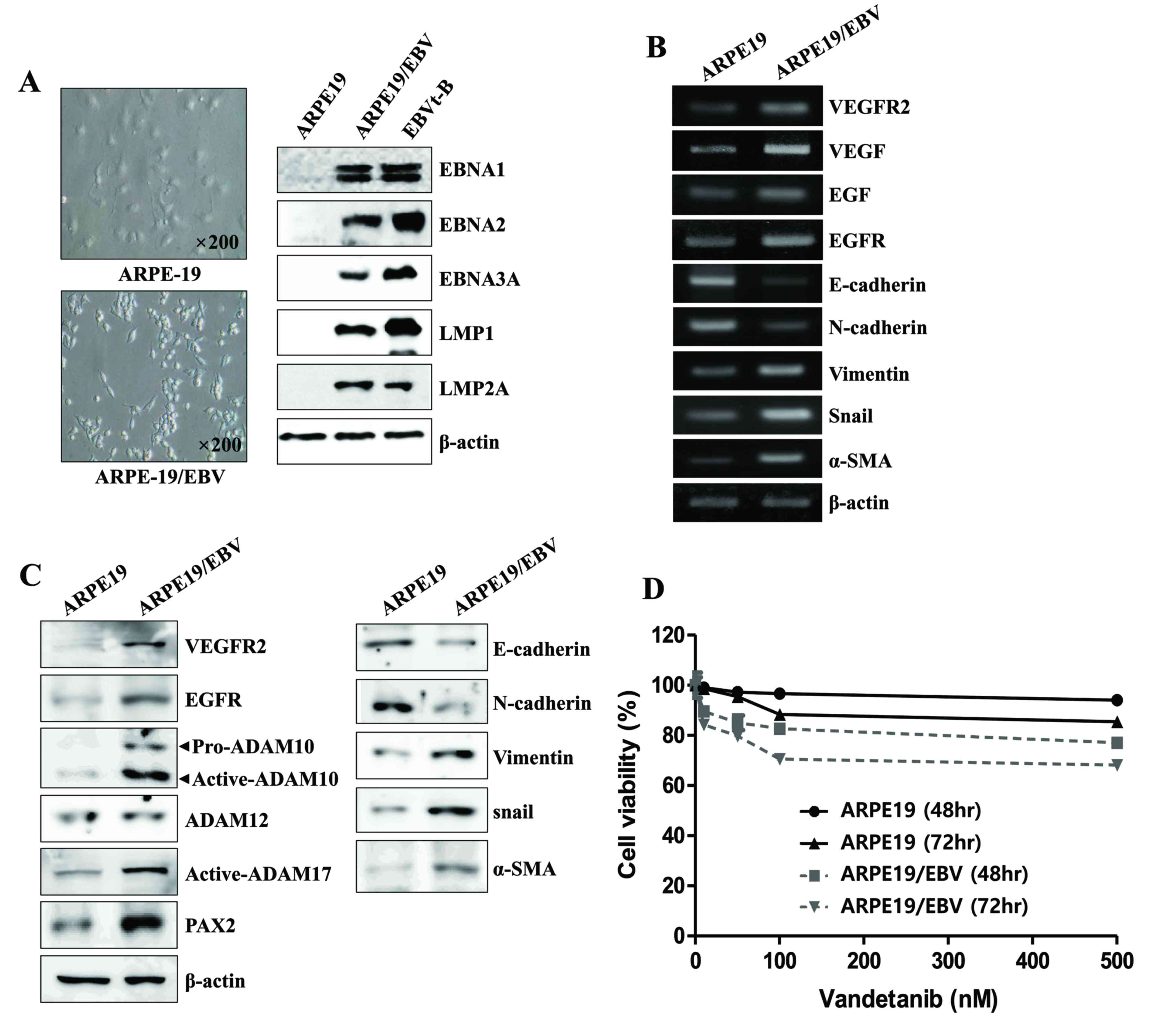 | Figure 1.Mesenchymal morphology of ARPE19
cells elicited by EBV infection. (A) Mesenchymal cell-like
transformation was observed under an inverted phase-contrast
microscope and the level of EBV-related gene protein in
EBV-infected ARPE19 cells was measured using western blotting. (B)
Levels of VEGFR2, VEGF, EGFR, EGF, E-cadherin, N-cadherin,
Vimentin, Snail, α-SMA and β-actin mRNA in ARPE19 and ARPE19/EBV
cells were determined by reverse transcription-polymerase chain
reaction. (C) The levels of VEGFR2, EGFR, E-cadherin, N-cadherin,
Vimentin, Snail, α-SMA, pro-ADAM10, active-ADAM10, active-ADAM12,
active-ADAM17, PAX2 and β-actin expression in ARPE19 and ARPE19/EBV
cells were determined by western blot analysis. (D) ARPE19 and
ARPE19/EBV cells were treated with various concentrations of
vandetanib (0, 10, 50, 100 or 500 nM) for 48 and 72 h. Cell
viability was analyzed by MTT assay. The results are representative
of three independent experiments. ARPE19, adult retinal pigment
epithelium-19 cells; EBV, Epstein-Barr virus; VEGF, vascular
endothelial growth factor; VEGFR2, VEGF receptor 2; EGF, epidermal
growth factor; EGFR, EGF receptor; α-SMA, α-smooth muscle actin;
ADAM, disintegrin and metalloproteinase protein; PAX2, paired box
gene 2; EBNA, Epstein-Barr virus nuclear antigen; LMP, latent
membrane protein. |
Modulation of mesenchymal
characteristics of ARPE19/EBV cells following treatment with
vandetanib
The alteration of mesenchymal features in the
ARPE19/EBV cells at 48 and 72 h following treatment with vandetanib
was assessed. After 48 h treatment, the expression levels of VEGFR2
and EGFR in the ARPE19/EBV cells were attenuated compared with the
untreated group following treatment with low (50 nM) or high (500
nM) doses of vandetanib (Fig. 2A).
Epithelial cell markers of retina cells (E-cadherin and N-cadherin)
were also identified in the ARPE19/EBV cells treated with
vandetanib (Fig. 2A). By contrast,
the expression of proteins associated with mesenchymal
characteristics (vimentin, Snail and α-SMA) of ARPE19/EBV cells
decreased in a dose-dependent manner following treatment with
vandetanib (Fig. 2A). Similarly,
treatment of ARPE19/EBV cells with vandetanib led to downregulation
of ADAM10, ADAM17 and PAX2 expression in a dose-dependent manner
(Fig. 2B). The mesenchymal
characteristics of ARPE19/EBV cells were also considerably
suppressed at 72 h after treatment with low or high doses of
vandetanib (Fig. 2C and D). To
determine the effect of vandetanib on migratory or invasive
activity of the ARPE19/EBV cells, a wound healing assay was
performed. Treatment with 500 nM vandetanib significantly inhibited
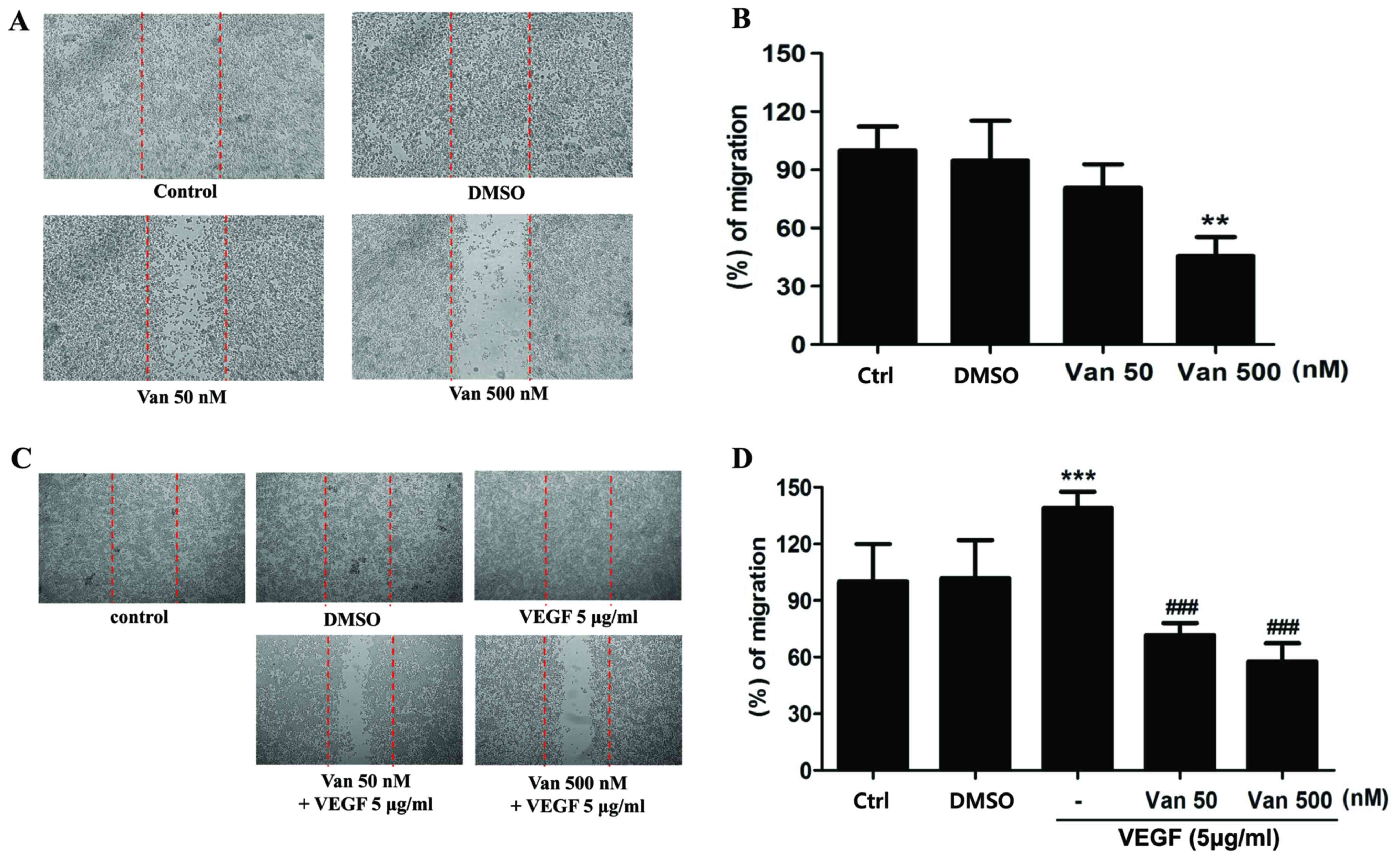
the migratory activity of the ARPE19/EBV cells (P<0.01; Fig. 3A and B). Furthermore,
vandetanib-treated ARPE19/EBV cells exhibited decreased wound
healing capacity compared with the cells cultured in normal media
or VEGF-supplemented conditioned media (P<0.001; Fig. 3C and D). These results suggest that
vandetanib may recover the epithelial characteristics of
EBV-infected ARPE19 cells and inhibit the migratory or invasive
activity of RPE cells in a retina pathological condition.
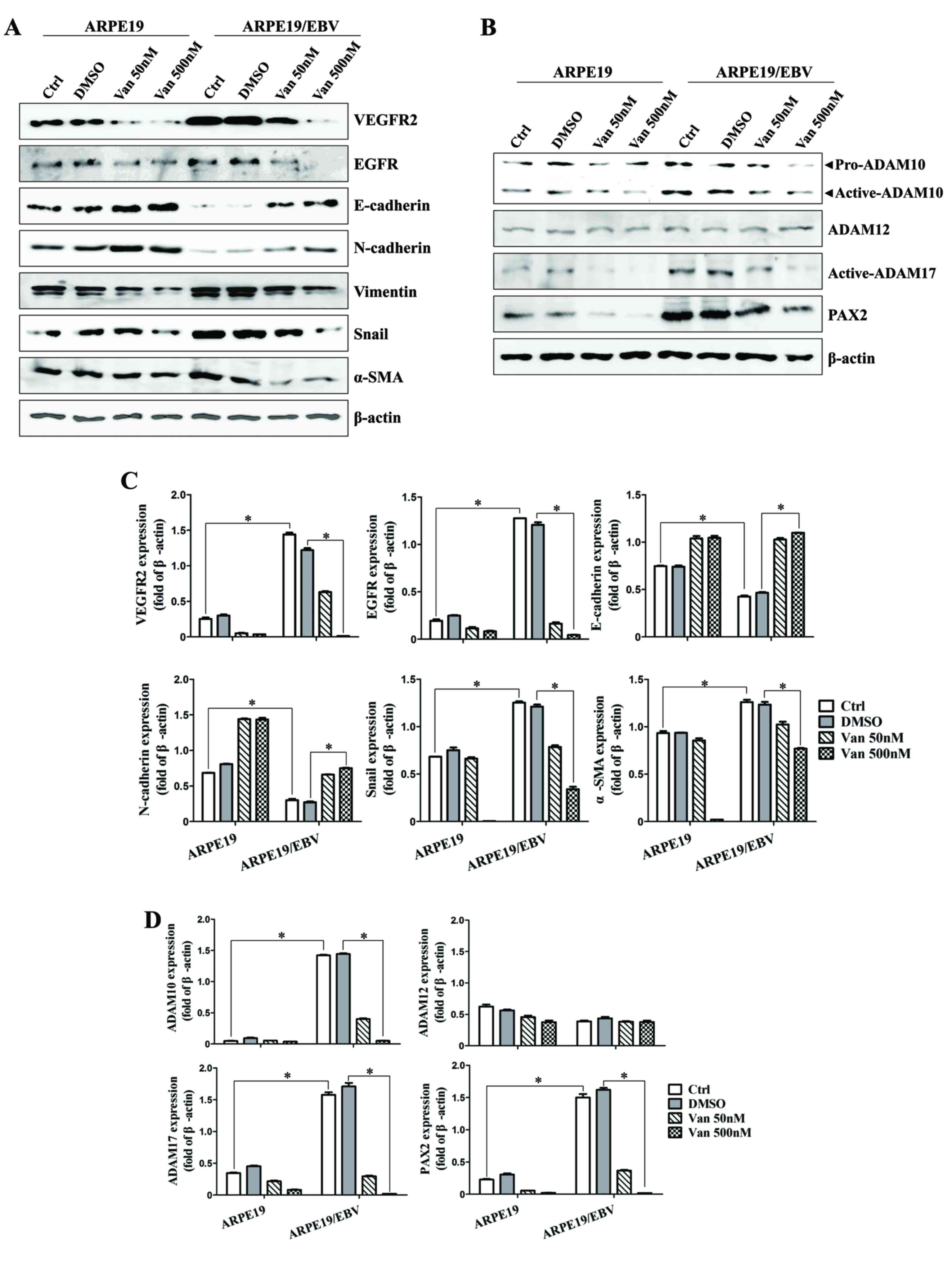 | Figure 2.Van decreases the expression of
VEGFR2, EGFR, ADAM family and EMT markers in ARPE19 and ARPE19/EBV
cells. ARPE19 and ARPE19/EBV cells were treated with 0, 50 and 500
nM Van for (A and B) 48 h or (C and D) 72 h. Whole cell lysates
were analyzed by western blot analysis using antibodies against
VEGFR2, EGFR, E-cadherin, N-cadherin, Vimentin, Snail, α-SMA,
pro-ADAM10, active-ADAM10, active-ADAM12, active-ADAM17, and PAX2.
β-actin was used as an internal control. The results are
representative of three independent experiments. (C and D) The
protein concentration in western blots was calculated using ImageJ
software and expressed relative to β-actin. *P<0.01. VEGFR2,
vascular endothelial growth factor receptor 2; EGFR, epidermal
growth factor receptor; ADAM, disintegrin and metalloproteinase
protein; ARPE19, adult retinal pigment epithelium-19 cells; α-SMA,
α-smooth muscle actin; PAX2, paired box gene 2; DMSO, dimethyl
sulfoxide; Van, vandetanib; Ctrl, control; EBV, Epstein-Barr
virus. |
Synergistic effect of vandetanib and
ADAM inhibitors on mesenchymal features and migratory activity in
ARPE19/EBV cells
Vandetanib affected the expression of ADAM family
proteins in the ARPE19/EBV cells. To examine the effect of ADAM
inhibitor on the expression of mesenchymal marker and invasion
ability in the ARPE19/EBV cells, vandetanib was combined with ADAM
inhibitors or a matrix metalloproteinase (MMP) inhibitor. In the
ARPE19/EBV cells, it was demonstrated that ADAM10 inhibitor
(GI254023X) and ADAM17 inhibitor (Marimastat) significantly
downregulated the expression of ADAM10 and ADAM17 respectively
(Fig. 4A; P<0.01). The MMP
inhibitor (ONO4817) also slightly decreased the expression of ADAM
17 (Fig. 4A). The combination of low
dose vandetanib and each inhibitor clearly reduced the expression
of VEGFR2, EGFR and PAX2 (Fig. 4B).
Furthermore, the expression of ADAM10 and ADAM17 in the ARPE19/EBV
cells co-treated with vandetanib and each inhibitor was also
downregulated, compared with cells treated with vandetanib alone
(Fig. 4B). The expression of
epithelial markers (E-cadherin and N-cadherin) markedly increased
following combination treatment with vandetanib and each inhibitor.
Vimentin, Snail and α-SMA expression were all decreased following
co-treatment with vandetanib and each inhibitor (Fig. 4B). In addition, the combination
treatment with vandetanib and each ADAM inhibitor markedly
inhibited the migratory activity of the ARPE19/EBV cells compared
with that of single drug treatment (Fig.
4C and D). To determine the recovery of epithelial markers
(E-cadherin) in the ARPE19/EBV cells, an immunofluorescence
analysis was performed and visualized using confocal microscopy.
E-cadherin expression in the ARPE19/EBV cells was upregulated
following treatment with vandetanib in a dose-dependent manner
(Fig. 5A). Similarly, GI254023X,
Marimastat and ONO4817 induced the expression of E-cadherin in the
ARPE19/EBV cells (Fig. 5B).
Furthermore, an evident increase in E-cadherin was observed in the
ARPE19/EBV cells co-treated with vandetanib and each inhibitor
compared with the cells treated with each inhibitor alone (Fig. 5C). These results suggest that a
combination treatment with vandetanib and ADAM inhibitor may
efficiently control the expression of mesenchymal features in RPE
cells.
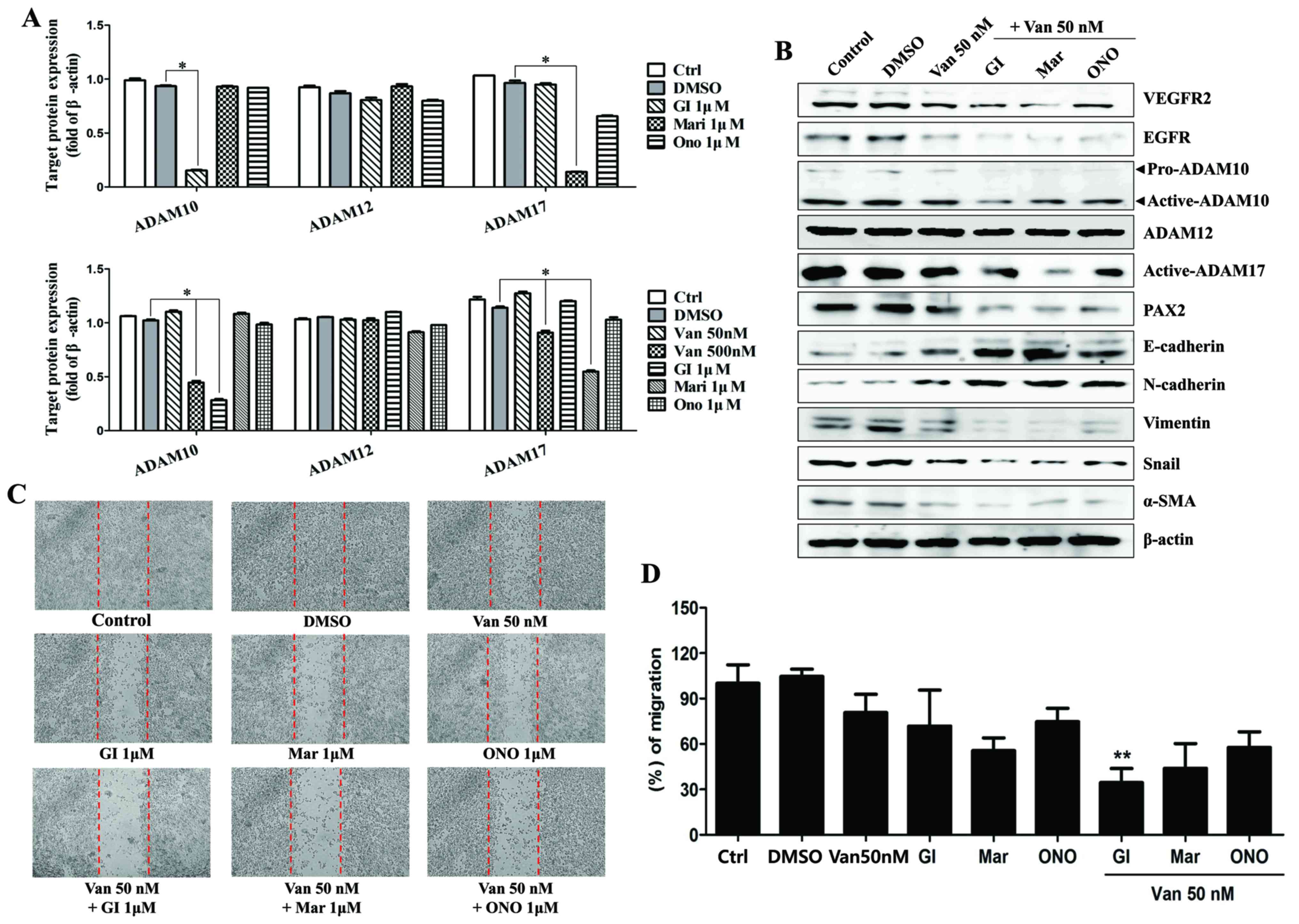 | Figure 4.Inhibition of ADAMs enhances the
inhibitory effects of vandetanib in ARPE19/EBV cells. (A)
ARPE19/EBV cells were treated with various ADAM inhibitors to
examine the effect against ADAM activity. ARPE19/EBV cells were
treated with 0, 50 or 500 nM Van for 3 h to compare the blocking
effect on ADAM activity. The cells were also treated with 1 µM GI
(ADAM10 inhibitor), 1 µM Mari (ADAM17 inhibitor) or 1 µM ONO
(matrix metalloproteinase inhibitor) for 1 h. Following washing
with phosphate-buffered saline, the cells were incubated for 24 h
in Dulbecco's modified Eagle's medium/F12 containing 10% fetal
bovine serum. Whole cell lysates were analyzed by western blot
analysis using antibodies against pro-ADAM10, active-ADAM10,
active-ADAM12 and active-ADAM17. The protein concentration in
western blots was calculated using Image J software and expressed
relative to β-actin. *P<0.01. (B) ARPE19/EBV cells were
pre-treated with 50 nM Van for 3 h and then exposed to 1 µM GI,
Mari or ONO for 1 h. Following washing with phosphate-buffered
saline, cells were maintained for 48 h Dulbecco's modified Eagle's
medium/F12 containing 10% fetal bovine serum. Whole cell lysates
were analyzed by western blot analysis using antibodies against
VEGFR2, EGFR, E-cadherin, N-cadherin, Vimentin, Snail, α-SMA,
pro-ADAM10, active-ADAM10, active-ADAM12, active-ADAM17, and PAX2.
β-actin was used as an internal control. (C and D) Cell motility
was measured by a wound-healing assay. Dotted lines indicate the
initial wounded area. Wound closure was slower in cells treated
with vandetanib and ADAM inhibitors compared with those treated
with DMSO or a single drug. The migratory distance of the cells
toward the scratched area was imaged and calculated as a percentage
of migration. **P<0.01 vs. Van 50 nM. The results are
representative of three independent experiments. ADAM, disintegrin
and metalloproteinase protein; ARPE19, adult retinal pigment
epithelium-19; EBV, Epstein-Barr virus; GI, GI254023X; Mari,
Marimastat; ONO, ONO4817; Van, vandetanib; DMSO, dimethyl
sulfoxide; VEGFR2, vascular endothelial growth factor receptor 2;
EGFR, epidermal growth factor receptor; α-SMA, α-smooth muscle
actin; PAX2, paired box gene 2; Ctrl, control. |
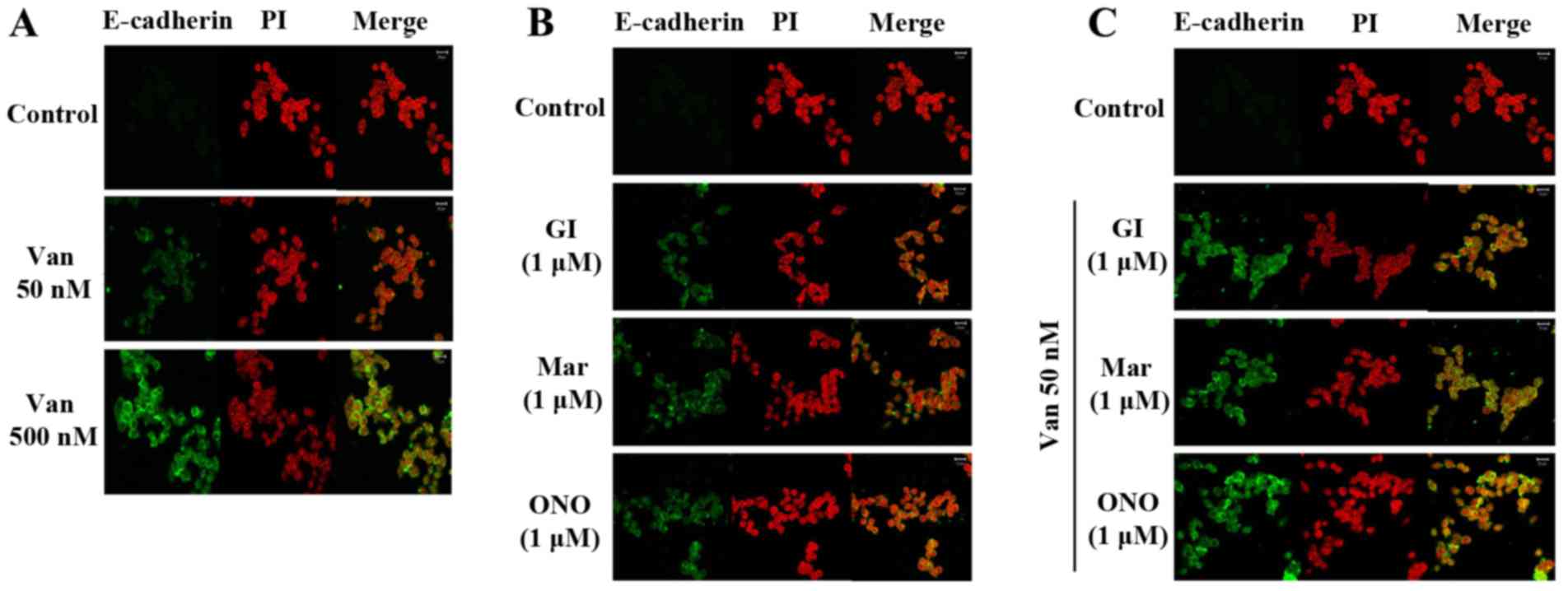 | Figure 5.Van and ADAM inhibitors significantly
increase E-cadherin in ARPE19/EBV cells. (A) ARPE19/EBV cells were
treated with 0, 50 or 500 nM Van for 3 h and then maintained for 8
h in DMEM/F12 containing 10% FBS (B) ARPE19/EBV cells were treated
with ADAM inhibitors for 1 h and then maintained for 8 h in
DMEM/F12 containing 10% FBS. (C) ARPE19/EBV cells were pre-treated
with 50 nM Van for 3 h and then treated with ADAM inhibitors (GI,
Mari, ONO) for 1 h. Following washing with phosphate-buffered
saline, the cells were maintained for 8 h in DMEM/F12 containing
10% FBS. The nucleus was stained with PI. The cells were visualized
under a confocal microscope (magnification, ×200). Green
fluorescence indicates E-cadherin and red fluorescence indicates
the nucleus (scale bar, 20 µm). The results are representative of
three independent experiments. ARPE19, adult retinal pigment
epithelium-19; Van, vandetanib; ADAM, disintegrin and
metalloproteinase protein; DMEM, Dulbecco's modified Eagle's
medium; FBS, fetal bovine serum; GI, GI254023X; Mari, Marimastat;
ONO, ONO4817; PI, propidium iodide. |
Effect of combining vandetanib and
ADAM inhibitors on the MAPK signaling pathway to control EMT
characteristics in ARPE19/EBV cells
Finally, the effect of co-treatment with vandetanib
and an ADAM inhibitor on inhibiting the signaling pathway induced
by VEGF in the ARPE19/EBV cells was evaluated. The expression
levels of phosphorylated JNK and p38 MAPK were increased in the
ARPE19/EBV cells compared with ARPE19 cells but expression of
phosphorylated ERK was decreased in the ARPE19/EBV cells (Fig. 6A). Although the level of
phosphorylated ERK was clearly increased following treatment with
high doses of vandetanib, there was no effect on the levels of
phosphorylated JNK, p38 MAPK and Akt in ARPE19/EBV cells (Fig. 6A); however, co-treatment with a low
dose of vandetanib and an ADAM or MMP inhibitor upregulated levels
of phosphorylated ERK and attenuated the signaling pathway by
blocking the phosphorylation of JNK, p38 MAPK and Akt (Fig. 6B). The upregulation of the
VEGF-induced mesenchymal markers (vimentin, Snail and α-SMA) and
ADAM proteins was effectively blocked following co-treatment with
low dose vandetanib and each inhibitor (Fig. 6C). In addition, the migratory
activity of VEGF-treated ARPE19/EBV cells significantly decreased
(P<0.01) following combination treatment with low dose of
vandetanib and ADAM or MMP inhibitor (Fig. 6D and E). These results suggest that
combination treatment with vandetanib and ADAM inhibitors regulate
the VEGF-mediated migratory activity of ARPE19/EBV cells through
modulation of the MAPK signaling pathway.
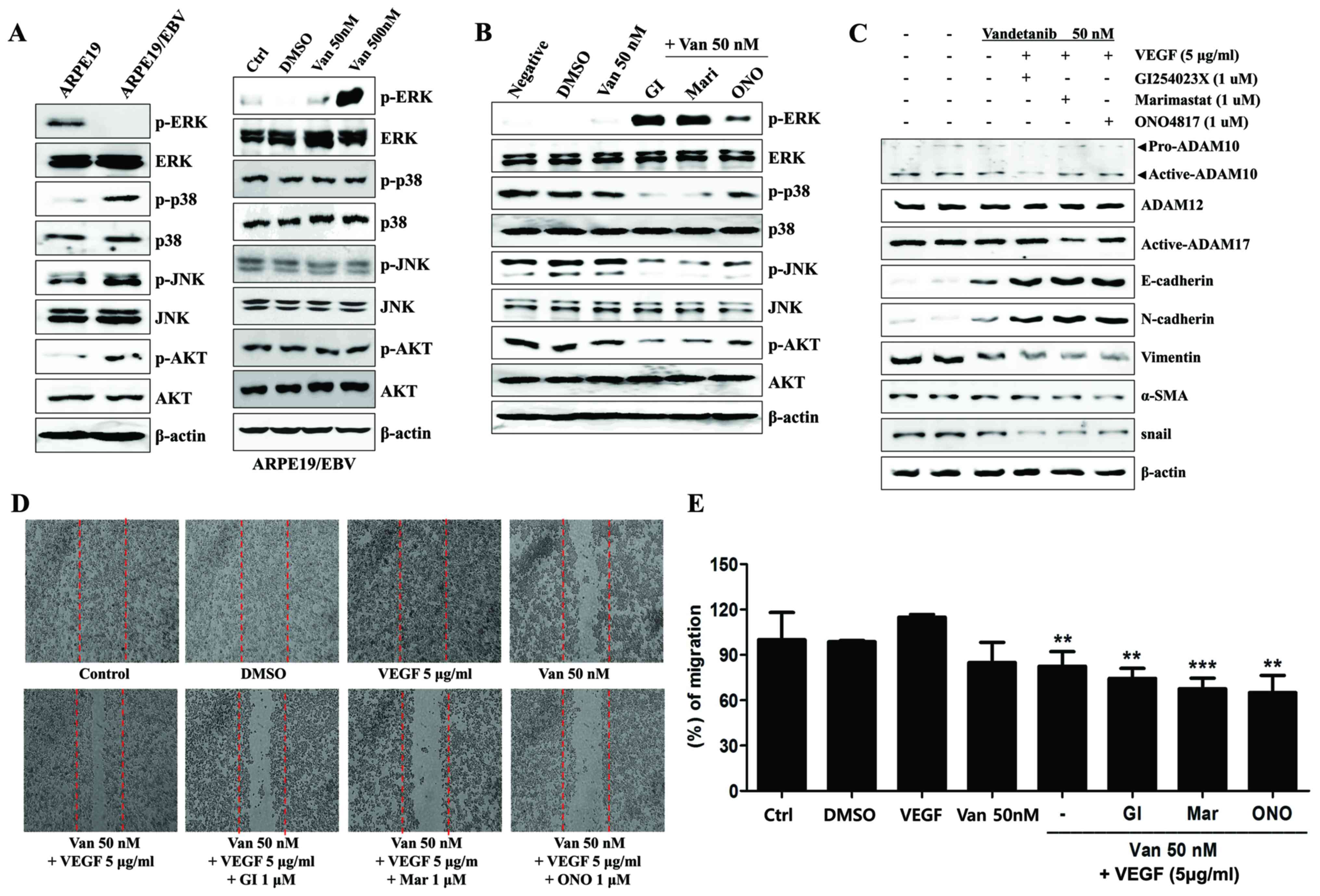 | Figure 6.Combined treatment with Van and ADAM
inhibitors attenuates the MAPK signaling pathway and reverses
epithelial-mesenchymal transition markers in ARPE19/EBV cells. (A)
Protein levels of p-ERK, ERK, p-p38, p38, p-JNK, JNK, p-AKT, and
β-actin in ARPE19 and ARPE19/EBV cells. ARPE19/EBV cells treated
with various concentrations of Van (0, 50, 500 nM) for 3 h in order
to analyze MAPK expression. (B) ARPE19/EBV cells were pre-treated
with 50 nM Van for 3 h and exposed to 1 µM GI, Mari or ONO for 1 h.
Following washing with phosphate buffered saline, the cells were
maintained for 24 h in DMEM/F12 containing 10% FBS. Whole cell
lysates were prepared and used in western blot analysis to
determine the expression of p-ERK, ERK, p-p38, p38, p-JNK, JNK,
p-AKT, AKT and β-actin, which served as an internal control. (C)
ARPE19/EBV cells were pre-treated with Van (50 nM) for 3 h and then
treated with recombinant VEGF (5 µg/ml) for 30 min. Following VEGF
treatment; the cells were treated with GI, Mari and ONO for 1 h and
incubated for 8 h in DMEM/F12 containing 10% FBS. Whole cell
lysates were prepared and used for western blot analysis to
determine the expression of pro-ADAM10, active-ADAM10,
active-ADAM12, active-ADAM17, E-cadherin, N-cadherin, Vimentin,
α-SMA, snail and β-actin, which served as an internal control. (D
and E) The migration ability of the cells was analyzed using a
wound healing assay. Dotted lines indicate the initial wounded
area. The length of the cells that migrated into the scratched area
was imaged (D) and calculated as a percentage of migration (E).
**P<0.01, ***P<0.001 vs. VEGF. The results are representative
of three independent experiments. Van, Vandetanib; ADAM,
disintegrin and metalloproteinase protein; MAPK, mitogen-activated
protein kinase; ARPE19, adult retinal pigment epithelium-19; ERK,
extracellular signal-regulated kinase; JNK, c-Jun N-terminal
kinase; AKT, protein kinase B; DMEM, Dulbecco's modified Eagle's
medium; FBS, fetal bovine serum; DMSO, dimethyl sulfoxide; GI,
GI254023X; Mari, Marimastat; ONO, ONO4817; VEGF, vascular
endothelial growth factor; α-SMA, α-smooth muscle actin; Ctrl,
control; p-, phosphorylated. |
Discussion
VEGF expression is increased in RPE cells of the
macula in patients with AMD, a condition associated with a high
risk of CNV (2). CNV is a serious
vision-limiting complication in which the integrity of the BM is
compromised, allowing leaky tortuous vessels to sprout from the
choriocapillaris into the subretinal pigment epithelium and the
subretinal spaces (7,8). In humans, it has been demonstrated that
the vitreous level of VEGF is increased in patients with
proliferative diabetic retinopathy (30). Therefore, intravitreal administration
of neutralizing anti-VEGF monoclonal antibody is currently the
primary treatment for AMD. Bevacizumab (Avastin®) is a
full-length antibody and ranibizumab (Lucentis™) is an antibody
fragment. Both bind all isoforms of VEGF. Unfortunately,
Bevacizumab increases the expression of α-SMA and decreases the
expression of zonula occludens-1 by stimulating the secretion of
connective tissue growth factor (CTGF) in ARPE19 cells (31). CTGF is involved in the pathogenesis
of PVR and retinal fibrosis, and serves an important role in EMT of
RPE (32–34). In addition, AMD treated with
anti-VEGF monoclonal antibodies generated undesirable additional
hemorrhagic retinal lesions, which develop ≤3.5 years following the
initiation of therapy (35).
Furthermore, VEGF depletion in adult mouse RPE cells rapidly leads
to vision loss and dysfunction of cone photoreceptors in
physiological and pathological states (36). Based on these results, novel targets
or drugs to prevent the progression of AMD through additional
regulation are required and were assessed in the present study. The
results of the current study suggest that the combination of
vandetanib and ADAM10 or ADAM17 inhibitors may synergistically
control the AMD-related EMT of RPE cells by upregulating epithelial
markers.
EBV infection induces the expression of a series of
cell-invasiveness and angiogenic factors, including MMP9 (37), MMP1, MMP3 (38) and VEGF (39), regardless of the LMP-1 expression of
EBV (40). It has been demonstrated
that EBV-infected HCECs exhibit increased migration and
invasiveness due to the upregulation of MMP2 and MMP9 (27). Similarly, the present study indicated
that EBV induced the loss of the epithelial marker E-cadherin and
induced the upregulation of the mesenchymal markers (vimentin,
Snail, and α-SMA) in ARPE19 cells. Treatment with vandetanib also
enhanced the expression of E-cadherin in epithelial cells.
Furthermore, mesenchymal cells exhibit decreased vimentin
expression following treatment with vandetanib in the presence of
EGF and VEGF (41). Vandetanib is
considered to be a potential postoperative adjuvant therapy to
inhibit the proliferation, progression, migration and survival of
cancer cells, as it serves a role in the regulation of the VEGFR
and EGFR signaling pathways (42).
However, this drug also induces various side-effects including
diarrhea, hypertension and prolongation of the QT interval
(43). It has been suggested that
the PI3K/Akt pathway may be activated in a VEGFR1-dependent manner
in certain cells (44,45). Proteomic analysis has indicated that
phosphorylated-EGFR and VEGFR2 are significantly inhibited by
vandetanib in tumor tissues (46).
In addition, vandetanib inhibits the phosphorylation of ERK1/2
(47). Thus, in the present study it
was investigated whether the combination of vandetanib with an ADAM
inhibitor influences the signaling pathway induced by VEGF.
Phosphorylated ERK was increased following combination treatment
with vandetanib and ADAM inhibitor or co-treatment with vandetanib
and MMP inhibitor. Additionally, the ARPE19/EBV cells co-treated
with vandetanib and an ADAM inhibitor more efficiently reduced the
expression of VEGFR and suppressed the signal transduction by
inhibiting the phosphorylation of JNK, p38-MAPK and Akt compared
with cells treated with vandetanib alone. The results of the
current study suggest that co-treatment with vandetanib and an ADAM
inhibitor more effectively prevents the signal transduction
underlying pathological migration of RPE cells, caused by
VEGF/VEGFR2 in retinal diseases.
ADAM family proteins have emerged as major
proteinases that mediate ectodomain shedding. The dysregulation of
ectodomain shedding is associated with autoimmune and
cardiovascular diseases, cancer, inflammation, infection and
neurodegeneration (25). Inhibition
of ADAM10 may inhibit Pax6 expression and N-cadherin ectodomain
shedding in retinal cells, potentially affecting neurite outgrowth
and differentiation of ganglion cells (48). Furthermore, the ectodomain shedding
activity of ADAM17 demonstrates wide substrate specificity
including cytokine receptors, TNF receptor, VEGFR2, adhesion
molecules (such as L-selectin) and transforming growth factor-α
(26,49). Inhibition of ADAM17 induces the
expression of thrombospondin 1, a naturally occurring inhibitor of
angiogenesis, whereas ADAM10 inhibition does not (50). Increased production of MMPs by TNF-α
and ADAM17 activated by VEGF in the vascular endothelial cells
serves a role in the regulation of retinal neovascularization
(51). The endothelial-specific
reduction of ADAM10 and inhibition of γ-secretase increases the
collateral formation and endothelial-specific knockdown of ADAM17
reduced collateral formation (52);
however, ADAM10 or ADAM17 inhibition exhibited a comparable
influence on the invasion activity of the ARPE19/EBV cells in the
current study. Furthermore, it was demonstrated that the
combination of low dose vandetanib with an ADAM10 or ADAM17
inhibitor synergistically attenuated the migratory capacity of
EBV-infected ARPE19 cells in an in vitro model of CNV or
PVR. In conclusion, the results of the present study suggest that
the inhibition of VEGF-mediated EMT signaling through co-treatment
with vandetanib and an ADAM inhibitor provides a novel and
promising therapeutic measure for neovascular AMD or other retina
pathological conditions.
Acknowledgements
The present study was supported by a grant from the
Korea Healthcare Technology R&D Project of the Ministry of
Health and Welfare Affairs, Republic of Korea (no. HI12C0005).
References
|
1
|
Kerbel RS: Tumor angiogenesis: Past,
present and the near future. Carcinogenesis. 21:505–515. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kliffen M, Sharma HS, Mooy CM, Kerkvliet S
and de Jong PT: Increased expression of angiogenic growth factors
in age-related maculopathy. Br J Ophthalmol. 81:154–162. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ida H, Tobe T, Nambu H, Matsumura M, Uyama
M and Campochiaro PA: RPE cells modulate subretinal
neovascularization, but do not cause regression in mice with
sustained expression of VEGF. Invest Ophthalmol Vis Sci.
44:5430–5437. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Penn JS, Madan A, Caldwell RB, Bartoli M,
Caldwell RW and Hartnett ME: Vascular endothelial growth factor in
eye disease. Prog Retin Eye Res. 27:331–371. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ferrara N, Houck K, Jakeman L and Leung
DW: Molecular and biological properties of the vascular endothelial
growth factor family of proteins. Endocr Rev. 13:18–32. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tuccillo C, Romano M, Troiani T,
Martinelli E, Morgillo F, De Vita F, Bianco R, Fontanini G, Bianco
RA, Tortora G and Ciardiello F: Antitumor activity of ZD6474, a
vascular endothelial growth factor-2 and epidermal growth factor
receptor small molecule tyrosine kinase inhibitor, in combination
with SC-236, a cyclooxygenase-2 inhibitor. Clin Cancer Res.
11:1268–1276. 2005.PubMed/NCBI
|
|
7
|
Ambati J and Fowler BJ: Mechanisms of
age-related macular degeneration. Neuron. 75:26–39. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Swaroop A, Chew EY, Rickman CB and
Abecasis GR: Unraveling a multifactorial late-onset disease: From
genetic susceptibility to disease mechanisms for age-related
macular degeneration. Annu Rev Genomics Human Genet. 10:19–43.
2009. View Article : Google Scholar
|
|
9
|
Campochiaro PA: Ocular neovascularization.
J Mol Med (Berl). 91:311–321. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zarbin MA: Current concepts in the
pathogenesis of age-related macular degeneration. Arch Ophthalmol.
122:598–614. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Aiello LP, Avery RL, Arrigg PG, Keyt BA,
Jampel HD, Shah ST, Pasquale LR, Thieme H, Iwamoto MA, Park JE, et
al: Vascular endothelial growth factor in ocular fluid of patients
with diabetic retinopathy and other retinal disorders. New Engl J
Med. 331:1480–1487. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Young RW: Pathophysiology of age-related
macular degeneration. Sur Ophthalmol. 31:291–306. 1987. View Article : Google Scholar
|
|
13
|
Nagineni CN, Kommineni VK, William A,
Detrick B and Hooks JJ: Regulation of VEGF expression in human
retinal cells by cytokines: Implications for the role of
inflammation in age-related macular degeneration. J Cell Physiol.
227:116–126. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bao B, Ali S, Ahmad A, Azmi AS, Li Y,
Banerjee S, Kong D, Sethi S, Aboukameel A, Padhye SB and Sarkar FH:
Hypoxia-induced aggressiveness of pancreatic cancer cells is due to
increased expression of VEGF, IL-6 and miR-21, which can be
attenuated by CDF treatment. PLoS One. 7:e501652012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hirasawa M, Noda K, Noda S, Suzuki M,
Ozawa Y, Shinoda K, Inoue M, Ogawa Y, Tsubota K and Ishida S:
Transcriptional factors associated with pithelial-mesenchymal
transition in choroidal neovascularization. Mol Vis. 17:1222–1230.
2011.PubMed/NCBI
|
|
16
|
Noël A, Jost M, Lambert V, Lecomte J and
Rakic JM: Anti-angiogenic therapy of exudative age-related macular
degeneration: Current progress and emerging concepts. Trend Mol
Med. 13:345–352. 2007. View Article : Google Scholar
|
|
17
|
Zhang SX and Ma JX: Ocular
neovascularization: Implication of endogenous angiogenic inhibitors
and potential therapy. Prog Ret Eye Res. 26:1–37. 2007. View Article : Google Scholar
|
|
18
|
Wedge SR, Ogilvie DJ, Dukes M, Kendrew J,
Chester R, Jackson JA, Boffey SJ, Valentine PJ, Curwen JO, Musgrove
HL, et al: ZD6474 inhibits vascular endothelial growth factor
signaling, angiogenesis, and tumor growth following oral
administration. Cancer Res. 62:4645–4655. 2002.PubMed/NCBI
|
|
19
|
Heymach JV, Johnson BE, Prager D, Csada E,
Roubec J, Pesek M, Spásová I, Belani CP, Bodrogi I, Gadgeel S, et
al: Randomized, placebo-controlled phase II study of vandetanib
plus docetaxel in previously treated non-small cell lung cancer. J
Clin Oncol. 25:4270–4277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Heymach JV, Paz-Ares L, De Braud F,
Sebastian M, Stewart DJ, Eberhardt WE, Ranade AA, Cohen G, Trigo
JM, Sandler AB, et al: Randomized phase II study of vandetanib
alone or with paclitaxel and carboplatin as first-line treatment
for advanced non-small-cell lung cancer. J Clin Oncol.
26:5407–5415. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sarkar S, Mazumdar A, Dash R, Sarkar D,
Fisher PB and Mandal M: ZD6474, a dual tyrosine kinase inhibitor of
EGFR and VEGFR-2, inhibits MAPK/ERK and AKT/PI3-K and induces
apoptosis in breast cancer cells. Cancer Biol Ther. 9:592–603.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Klettner A and Roider J: Constitutive and
oxidative-stress-induced expression of VEGF in the RPE are
differently regulated by different Mitogen-activated protein
kinases. Graefes Arch Clin Exp Ophthalmol. 247:1487–1492. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yan X, Lin J, Rolfs A and Luo J:
Differential expression of the ADAMs in developing chicken retina.
Dev Growth Differ. 53:726–739. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sel S, Kalinski T, Enssen I, Kaiser M,
Nass N, Trau S, Wollensak G, Bräuer L, Jäger K and Paulsen F:
Expression analysis of ADAM17 during mouse eye development. Ann
Anat. 194:334–338. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saftig P and Reiss K: The ‘A Disintegrin
and Metalloproteases’ ADAM10 and ADAM17: Novel drug targets with
therapeutic potential? Eur J Cell Biol. 90:527–535. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Swendeman S, Mendelson K, Weskamp G,
Horiuchi K, Deutsch U, Scherle P, Hooper A, Rafii S and Blobel CP:
VEGF-A stimulates ADAM17-dependent shedding of VEGFR2 and crosstalk
between VEGFR2 and ERK signaling. Circ Res. 103:916–918. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park GB, Kim D, Kim YS, Kim S, Lee HK,
Yang JW and Hur DY: The Epstein-Barr virus causes
epithelial-mesenchymal transition in human corneal epithelial cells
via Syk/src and Akt/Erk signaling pathways. Invest Ophthalmol Vis
Sci. 55:1770–1779. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li H, Li M, Xu D, Zhao C, Liu G and Wang
F: Overexpression of Snail in retinal pigment epithelial triggered
epithelial-mesenchymal transition. Biochem Biophys Res Commun.
446:347–351. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Saika S, Kono-Saika S, Tanaka T, Yamanaka
O, Ohnishi Y, Sato M, Muragaki Y, Ooshima A, Yoo J, Flanders KC and
Roberts AB: Smad3 is required for dedifferentiation of retinal
pigment epithelium following retinal detachment in mice. Lab
Invest. 84:1245–1258. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Adamis AP, Miller JW, Bernal MT, D'Amico
DJ, Folkman J, Yeo TK and Yeo KT: Increased vascular endothelial
growth factor levels in the vitreous of eyes with proliferative
diabetic retinopathy. Am J Ophthalmol. 118:445–450. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen CL, Liang CM, Chen YH, Tai MC, Lu DW
and Chen JT: Bevacizumab modulates epithelial-to-mesenchymal
transition in the retinal pigment epithelial cells via connective
tissue growth factor up-regulation. Acta Ophthalmol. 90:e389–e398.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kon CH, Occleston NL, Aylward GW and Khaw
PT: Expression of vitreous cytokines in proliferative
vitreoretinopathy: A prospective study. Invest Ophthalmol Vis Sci.
40:705–712. 1999.PubMed/NCBI
|
|
33
|
Leask A and Abraham DJ: The role of
connective tissue growth factor, a multifunctional matricellular
protein, in fibroblast biology. Biochem Cell Biol. 81:355–363.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Parapuram SK, Chang B, Li L, Hartung RA,
Chalam KV, Nair-Menon JU, Hunt DM and Hunt RC: Differential effects
of TGFbeta and vitreous on the transformation of retinal pigment
epithelial cells. Invest Ophthalmol Vis Sci. 50:5965–5974. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tanaka E, Chaikitmongkol V, Bressler SB
and Bressler NM: Vision-threatening lesions developing with
longer-term follow-up after treatment of neovascular age-related
macular degeneration. Ophthalmology. 122:153–161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kurihara T, Westenskow PD, Bravo S,
Aguilar E and Friedlander M: Targeted deletion of Vegfa in adult
mice induces vision loss. J Clin Invest. 122:4213–4217. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yoshizaki T, Sato H, Furukawa M and Pagano
JS: The expression of matrix metalloproteinase 9 is enhanced by
Epstein-Barr virus latent membrane protein 1. Proc Natl Acad Sci
USA. 95:3621–3626. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kondo S, Wakisaka N, Schell MJ, Horikawa
T, Sheen TS, Sato H, Furukawa M, Pagano JS and Yoshizaki T:
Epstein-Barr virus latent membrane protein 1 induces the matrix
metalloproteinase-1 promoter via an Ets binding site formed by a
single nucleotide polymorphism: Enhanced susceptibility to
nasopharyngeal carcinoma. Int J Cancer. 115:368–376. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Murono S, Inoue H, Tanabe T, Joab I,
Yoshizaki T, Furukawa M and Pagano JS: Induction of
cyclooxygenase-2 by Epstein-Barr virus latent membrane protein 1 is
involved in vascular endothelial growth factor production in
nasopharyngeal carcinoma cells. Proc Natl Acad Sci USA.
98:6905–6910. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lin JC, Liao SK, Lee EH, Hung MS, Sayion
Y, Chen HC, Kang CC, Huang LS and Cherng JM: Molecular events
associated with epithelial to mesenchymal transition of
nasopharyngeal carcinoma cells in the absence of Epstein-Barr virus
genome. J Biomed Sci. 16:1052009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li Y, Yang X, Su LJ and Flaig TW: VEGFR
and EGFR inhibition increases epithelial cellular characteristics
and chemotherapy sensitivity in mesenchymal bladder cancer cells.
Oncol Rep. 24:1019–1928. 2010.PubMed/NCBI
|
|
42
|
Yoshikawa D, Ojima H, Kokubu A, Ochiya T,
Kasai S, Hirohashi S and Shibata T: Vandetanib (ZD6474), an
inhibitor of VEGFR and EGFR signalling, as a novel
molecular-targeted therapy against cholangiocarcinoma. Br J Cancer.
100:1257–1266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chau NG and Haddad RI: Vandetanib for the
treatment of medullary thyroid cancer. Clin Cancer Res. 19:524–529.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Matsuzaki H, Tamatani M, Yamaguchi A,
Namikawa K, Kiyama H, Vitek MP, Mitsuda N and Tohyama M: Vascular
endothelial growth factor rescues hippocampal neurons from
glutamate-induced toxicity: Signal transduction cascades. FASEB J.
15:1218–1220. 2001.PubMed/NCBI
|
|
45
|
Roberts DM, Kearney JB, Johnson JH,
Rosenberg MP, Kumar R and Bautch VL: The vascular endothelial
growth factor (VEGF) receptor Flt-1 (VEGFR-1) modulates Flk-1
(VEGFR-2) signaling during blood vessel formation. Am J Pathol.
164:1531–1535. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Inoue K, Torimura T, Nakamura T, Iwamoto
H, Masuda H, Abe M, Hashimoto O, Koga H, Ueno T, Yano H and Sata M:
Vandetanib, an inhibitor of VEGF receptor-2 and EGF receptor,
suppresses tumor development and improves prognosis of liver cancer
in mice. Clin Cancer Res. 18:3924–3933. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Giannelli G, Azzariti A, Sgarra C,
Porcelli L, Antonaci S and Paradiso A: ZD6474 inhibits
proliferation and invasion of human hepatocellular carcinoma cells.
Biochem Pharmacol. 71:479–485. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Paudel S, Kim YH, Huh MI, Kim SJ, Chang Y,
Park YJ, Lee KW and Jung JC: ADAM10 mediates N-cadherin ectodomain
shedding during retinal ganglion cell differentiation in primary
cultured retinal cells from the developing chick retina. J Cell
Biochem. 114:942–954. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Peschon JJ, Slack JL, Reddy P, Stocking
KL, Sunnarborg SW, Lee DC, Russell WE, Castner BJ, Johnson RS,
Fitzner JN, et al: An essential role for ectodomain shedding in
mammalian development. Science. 282:1281–1284. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Caolo V, Swennen G, Chalaris A, Wagenaar
A, Verbruggen S, Rose-John S, Molin DG, Vooijs M and Post MJ:
ADAM10 and ADAM17 have opposite roles during sprouting
angiogenesis. Angiogenesis. 18:13–22. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Majka S, McGuire PG and Das A: Regulation
of matrix metalloproteinase expression by tumor necrosis factor in
a murine model of retinal neovascularization. Invest Ophthalmol Vis
Sci. 43:260–266. 2002.PubMed/NCBI
|
|
52
|
Lucitti JL, Mackey JK, Morrison JC, Haigh
JJ, Adams RH and Faber JE: Formation of the collateral circulation
is regulated by vascular endothelial growth factor-A and a
disintegrin and metalloprotease family members 10 and 17. Circ Res.
111:1539–1550. 2012. View Article : Google Scholar : PubMed/NCBI
|