Introduction
Hepatocellular carcinoma (HCC) is the third most
frequent cause of tumor-related deaths in China (1,2). Due to
the high metastatic potential of HCC (3,4),
progression is found in approximately 70% of patients within one
year of diagnosis (5), and it is
difficult to treat patients with advanced HCC (6,7). It is
therefore necessary to contrive novel anticancer reagents to
prevent the invasiveness of HCC.
Propofol is one of the most extensively used
intravenous anesthetic agents. Previous studies have confirmed that
propofol also has an antitumor effect (8–12).
Clinically relevant concentrations of propofol have the ability to
inhibit cell proliferation, motility and invasion, and induce
apoptosis of many cancer cells, such as HeLa, HT1080, HOS and
RPMI-7951 (9). Zhang et al
investigated the effect of propofol on liver cancer and found that
propofol can effectively inhibit the proliferation and invasiveness
of HCC cells, and induce apoptosis, in vitro (13,14). In
addition, Zhang et al demonstrated that propofol can inhibit
tumor growth in tumor-bearing mice by activation of macrophages,
thus exerting a therapeutic effect on HCC in vivo (15). However, the molecular mechanisms
underlying the antitumor function of propofol on HCC has not been
elucidated until now.
High Mobility Group AT-Hook 2 (HMGA2) is a member of
the high mobility group family. It is essential for tumor
progression and metastasis in vitro and in vivo
(16). The role of HMGA2 in normal
and HCC cell growth is well established (17,18).
HMGA2 was reported to serve important functions in
epithelial-mesenchymal transition (EMT) occurring in HCC,
contributing to tumor growth and invasion (18). Recently, it has been suggested that
HMGA2 acts as a transcriptional organizer of key signaling
molecules (19,20). Zha et al reported that the
Wnt/β-catenin pathway could be activated by HMGA2 (21).
To the best of our knowledge, the present study is
the first to investigate whether propofol affects cell
proliferation, apoptosis and invasion of HCC cells through the
HMGA2 mediated Wnt/β-catenin pathway, which may elucidate novel
mechanisms underlying the antitumor function of propofol on
HCC.
Materials and methods
Cell culture and drug treatment
The HepG2 cells (American Type Culture Collection,
Manassas, VA, USA) were cultured in Dulbecco's modified Eagle's
medium (HyClone; GE Healthcare, Logan, UT, USA) supplemented with
10% fetal bovine serum (FBS; HyClone), and maintained at 37°C in a
humidified atmosphere with 5% CO2. Following the use of
miRanda prediction software (accessible: http://www.microrna.org/microrna/home.do), it was
indicated that propofol may influence HMGA2 expression. Propofol
was obtained from Corden Pharma S.p.A. (Latina, Italy), and diluted
to concentrations of 5, 10 and 20 µg/ml to treat the HepG2 cells
for 24 h.
Cell transfection
Cell transfection of pcDNA3.1 and HMGA2-pcDNA3.1
plasmid was performed using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA),
according to the manufacturer's instructions. A negative control
group were treated with Lipofectamine® 2000 diluted in
Opti-MEM™ I (Gibco; Thermo Fisher Scientific, Inc.) only. Briefly,
5 µl Lipofectamine® 2000 and 100 ng HMGA2-pcDNA3.1
plasmid were diluted in 250 µl Opti-MEM™ I, respectively. After
incubation at room temperature for 5 min, the two solutions were
mixed and incubated for 20 min. Subsequently, the lipid-DNA
complexes were added to the cell plates. After incubation at 37°C
for 6 h, the culture medium was replaced with fresh medium. Cells
only treated with Lipofectamine® 2000 were used as
transfection control. After 48 h, the transfection efficiency was
assessed under the fluorescence microscope (N-STORM; Nikon
Corporation, Tokyo, Japan). The cells were harvested for analysis
when the transfection efficiency was >80%.
Western blot analysis
The HepG2 cells were harvested and lysed in RIPA
Lysis Buffer (Sangon Biotech, Shanghai, China). The protein
concentration was determined using a BCA Protein Assay kit (Sangon
Biotech). A total of 20 µg protein was loaded and separated by 10%
SDS-polyacrylamide gel. The proteins were then transferred onto
PVDF membranes (Merck Millipore, Billerica, MA, USA). After
blocking in 3% bovine serum albumin (Amresco, Inc., Solon, OH, USA)
at 37°C overnight, the membranes were incubated with the primary
antibodies at 37°C for 1 h. The primary antibodies were as follows:
Rabbit polyclonal to HMGA2 (ab52039; 1:800; Abcam, Cambridge, MA,
USA), mouse monoclonal to Wnt3a (sc-136163; 1:400; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), rabbit polyclonal to
β-catenin (sc-7199; 1:400; Santa Cruz Biotechnology, Inc.), mouse
monoclonal to Snail Family Zinc Finger 1 (Snail1) (#3895; 1:500;
Cell Signaling Technology, Inc., Beverly, MA, USA), rabbit
polyclonal to c-myc (ab52039; 1:800; Abcam) and rabbit polyclonal
to β-actin (#40552; 1:1,000; Signalway Antibody Inc., College Park,
MD, USA). The β-actin antibody was used as the internal control.
After washing with Tris-buffered saline with Tween (Sangon Biotech)
three times, the membranes were incubated with the secondary
antibodies [goat anti-mouse IgG HRP conjugated antibody (#L3032-2;
1:2,000; Signalway Antibody, Inc.), goat anti-rabbit IgG HRP
conjugated antibody (ab6721; 1:5,000; Abcam)] at 37°C for 1 h. The
blots were detected using an ECL western blotting kit (Pierce
Protein Biology; Thermo Fisher Scientific, Inc., Rockford, IL, USA)
and analyzed using Image-Pro Plus version 6.0 software (Media
Cybernetics, Inc., Rockville, MD, USA).
Cell proliferation assay
To determine cell proliferation, the
3-(4,5-dimethylthiazole-2-yl)-2,5-biphenyl tetrazolium bromide
(MTT) assay was performed. The HepG2 cells were suspended into a
density of 2×104 cells/ml, and 100 µl cell suspension
was added into each well of the 96-well plates. After incubation at
37°C for 0, 24, 48, 72 and 96 h, 10 µl MTT solution [5 mg/ml in
phosphate-buffered saline (PBS); Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany] was added into each well and incubated for 4 h.
The reaction product, formazan crystals, was dissolved by the
addition of dimethyl sulfoxide. The absorbance at 570 nm was
measured using a microplate reader (BioTek Instruments, Inc.,
Winooski, VT, USA).
Flow cytometry (FCM)
Cell apoptosis rate was determined using an Annexin
V-FITC/Propidium Iodide Apoptosis Detection kit (Kaiji Biological
Inc., Nanjing, China) according to the manufacturer's instructions.
Briefly, 5×105 cells were harvested and washed with PBS.
Binding Buffer was added to resuspend the cells, and then the cells
were stained with Annexin V and propidium iodide for 15 min. Cell
apoptosis was analyzed using FCM (Beckman Counter, Inc., Miami, FL,
USA).
Cell invasion assay
Matrigel matrix (BD Biosciences, Franklin Lakes, NJ,
USA) was added to the Transwell inserts (Corning, New York, NY,
USA) at a final concentration of 200 µg/ml and allowed to
polymerize at 37°C for 1 h. The HepG2 cells were suspended in
serum-free medium into a density of 5×104 cells/ml, and
added to the upper chambers. The cell medium containing 10% FBS was
added to the lower chambers. The cell plates were then incubated at
37°C in a humidified atmosphere with 5% CO2 overnight.
After that, the non-invaded cells were removed by a cotton swab,
and the invaded cells in the lower surface of the membrane were
fixed with ethanol and stained with hematoxylin for 15 min. The
stained cells were counted under a microscope (Nikon
Corporation).
Statistical analysis
All the data were analyzed using SPSS 19.0
statistical software (IBM SPSS, Armonk, NY, USA) and were presented
as the mean ± standard deviation. Statistical differences between
the two groups were analyzed with the Student's t-test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Effect of propofol on the expression
of HMGA2
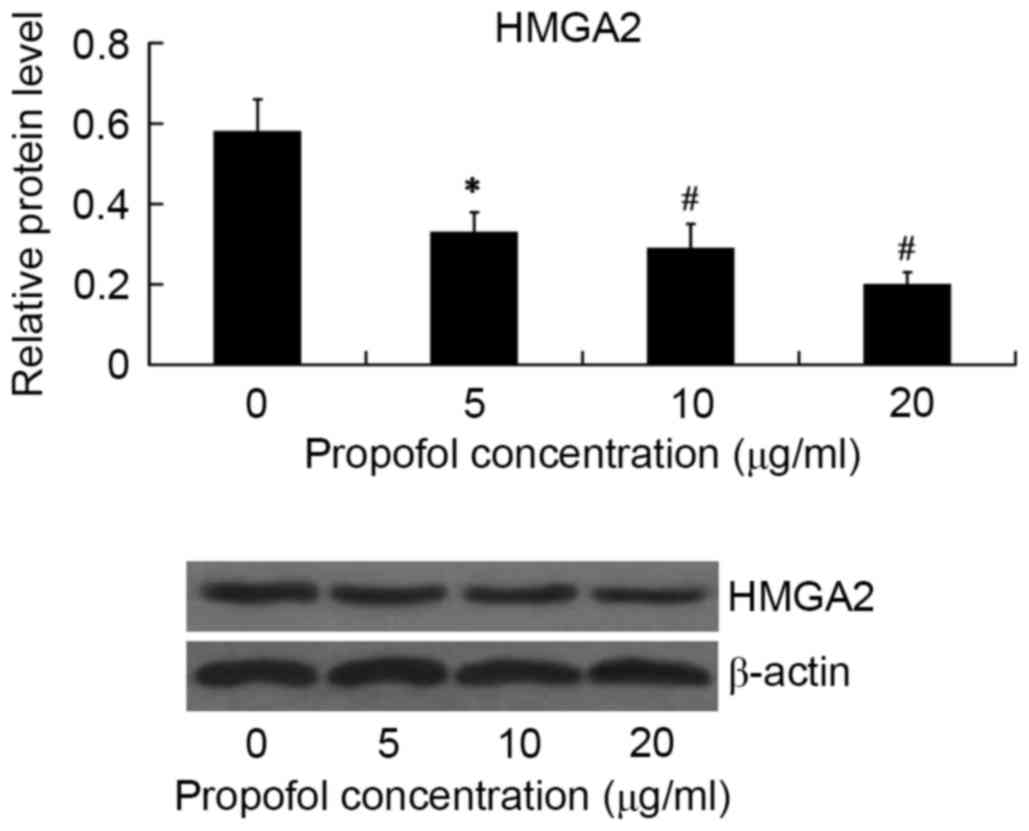
The effect of propofol on HMGA2 protein expression
in HepG2 cells was examined by western blot analysis. The HepG2
cells were treated with 5, 10 and 20 µg/ml propofol for 24 h, and
the relative protein level of HMGA2 was then examined. As shown in
Fig. 1, HMGA2 expression was
significantly inhibited by propofol in a dose-dependent manner.
Propofol showed the maximum inhibitory effect on HMGA2 expression
at the concentration of 20 µg/ml (P<0.01).
Effect of propofol on the activation
of Wnt/β-catenin signaling
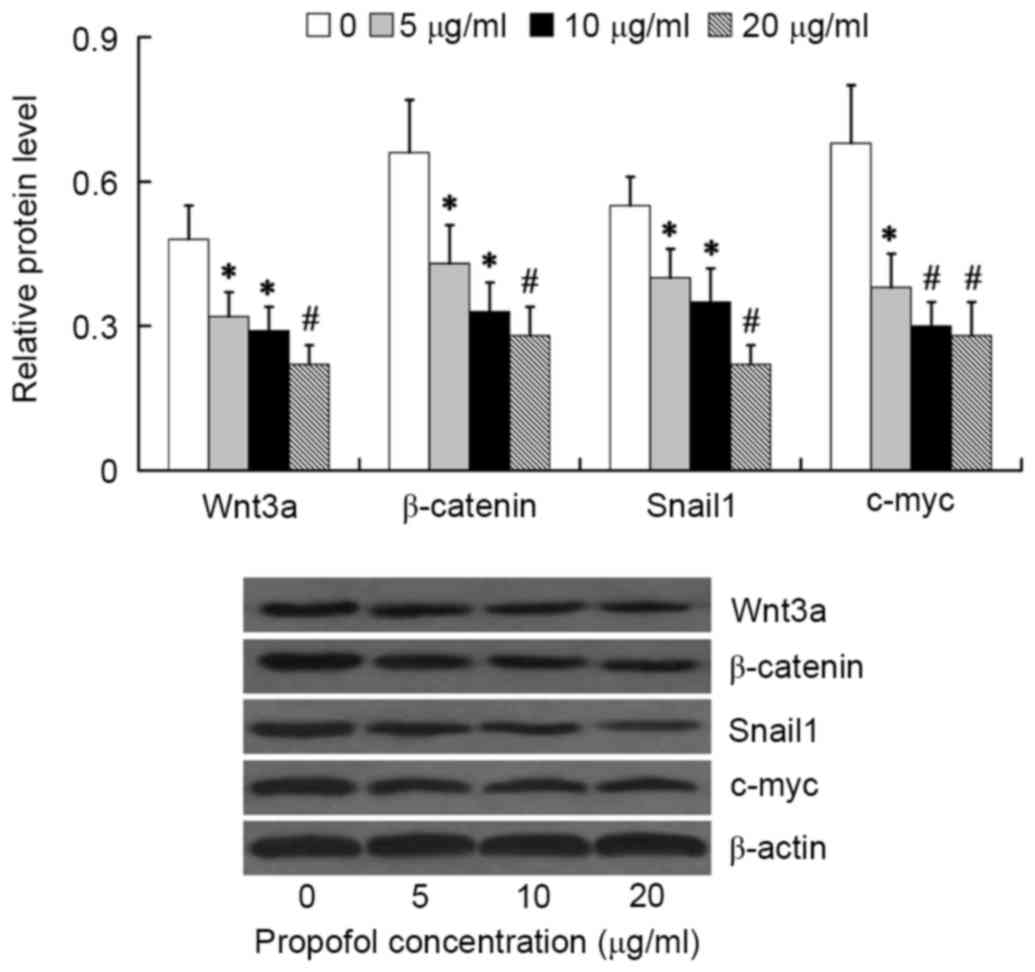
To investigate the effect of propofol on
Wnt/β-catenin signaling activation, the relative protein levels of
Wnt3a, β-catenin, Snail1 and c-myc were examined in HepG2 cells
following treatment with various concentrations of propofol for 24
h. We found that the Wnt/β-catenin signaling was significantly
inhibited by propofol treatment at concentrations between 5 and 20
µg/ml, as evidenced by decreased expression of Wnt3a, β-catenin,
Snail1 and c-myc in the propofol treatment groups compared with the
β-actin control group. The lowest expression values were observed
at propofol concentrations of 20 µg/ml (P<0.01; Fig. 2) thus, cells were treated with 20
µg/ml propofol in all subsequent experiments.
Role of HMGA2 in mediating the effect
of propofol on cell proliferation
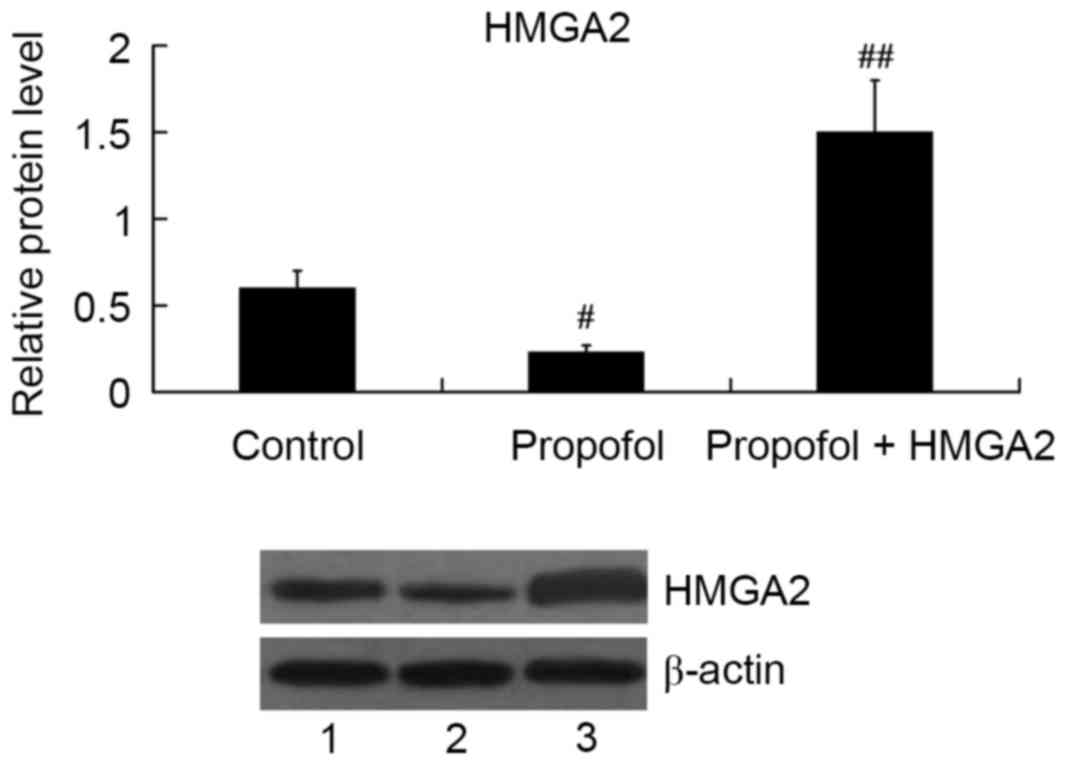
HMGA2-pcDNA3.1 plasmid was transfected into HepG2
cells to overexpress HMGA2, and then the cells were treated with 20
µg/ml propofol for 24 h. As expected, propofol inhibited HMGA2
expression; however, the relative protein level of HMGA2 was
significantly increased in the propofol + HMGA2 group compared with
the propofol group (P<0.01; Fig.
3).
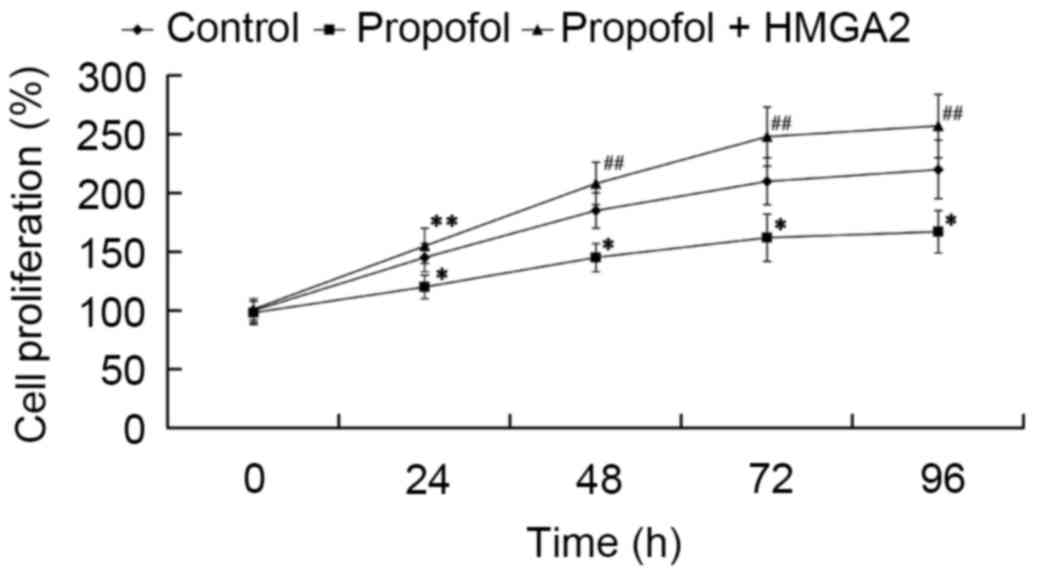
Subsequently, we investigated whether HMGA2 mediates
the effect of propofol on cell proliferation. Cells were
transfected with HMGA2-pcDNA3.1 plasmid and treated with 20 µg/ml
propofol for 24, 48, 72 and 96 h, then an MTT assay was performed.
As shown in Fig. 4, cell
proliferation was significantly inhibited by propofol compared to
the control group (P<0.05). Furthermore, it was found that HMGA2
overexpression reversed the inhibitory effect of propofol, as cell
proliferation significantly increased compared to the propofol
group (P<0.05 and P<0.01).
Role of HMGA2 in mediating the effect
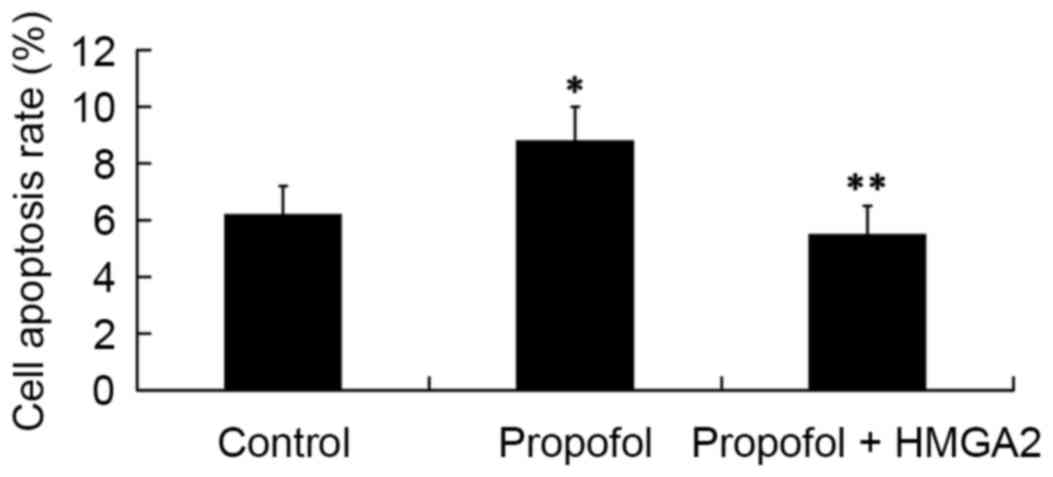
of propofol on cell apoptosis
Next, we investigated whether HMGA2 mediates the
effect of propofol on cell apoptosis by FCM analysis. The results
revealed that compared with the cell apoptosis rate in the control
group (6.2±1.0%), treatment of 20 µg/ml propofol for 24 h
significantly increased cell apoptosis rate to 8.8±1.2%
(P<0.05). However, the cell apoptosis rate was significantly
decreased to 5.5±1.0% by transfection with the HMGA2-pcDNA3.1
plasmid (P<0.05; Fig. 5).
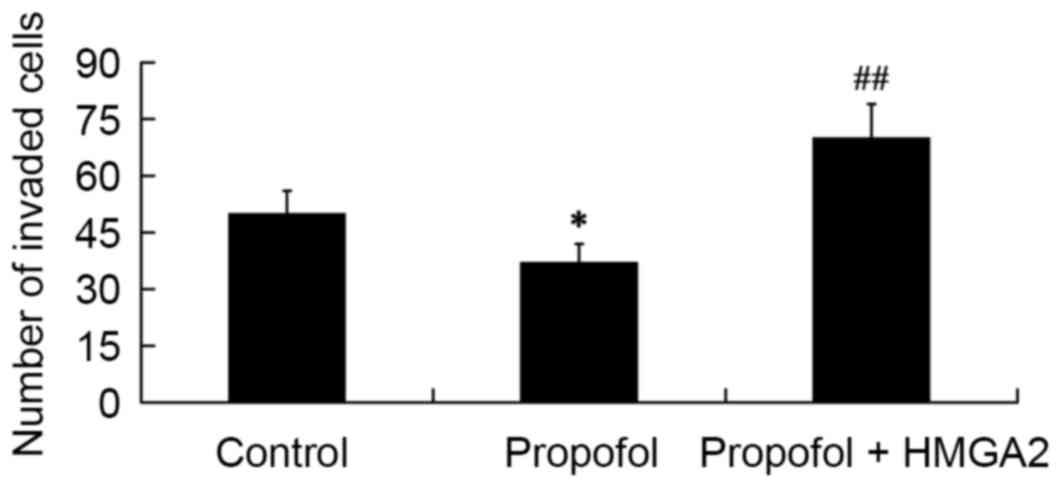
Role of HMGA2 in mediating the effect
of propofol on cell invasion
To determine the role of HMGA2 in mediating the
effect of propofol on cell invasion, a Transwell-matrigel cell
invasion assay was performed. The results showed that compared with
the control group, the number of invaded cells was significantly
decreased in the propofol group (P<0.05); however, the
inhibitory effect of propofol on cell invasion was reversed by
transfection of the HMGA2-pcDNA3.1 plasmid (P<0.01; Fig. 6).
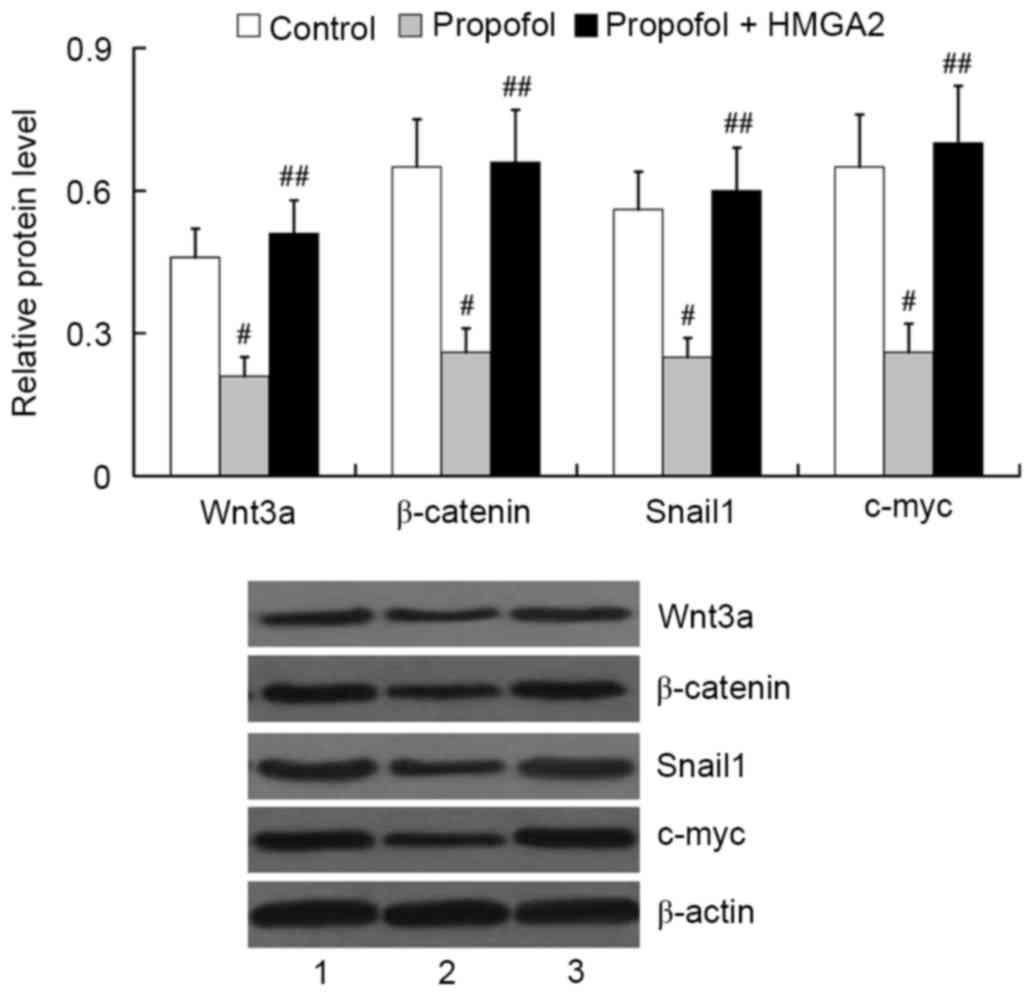
Role of HMGA2 in mediating the effect
of propofol on Wnt/β-catenin signaling activation
To investigate whether HMGA2 mediates the effect of
propofol on Wnt/β-catenin signaling activation, we examined the
expression of Wnt3a, β-catenin, Snail1 and c-myc in HepG2 cells
following transfection with the HMGA2-pcDNA3.1 plasmid and
treatment with 20 µg/ml propofol for 24 h. The results from western
blot analysis showed that propofol treatment induced significant
downregulation of Wnt3a, β-catenin, Snail1 and c-myc in HepG2 cells
(P<0.01). However, overexpression of HMGA2 was able to attenuate
the inhibitory effect of propofol on Wnt/β-catenin signaling
activation. The relative protein levels of Wnt3a, β-catenin, Snail1
and c-myc in the propofol + HMGA2 group were significantly higher
than that in the propofol group (P<0.01; Fig. 7).
Discussion
Propofol is a commonly used intravenous anesthetic
in tumor surgery. Increasing evidence has shown that propofol can
effectively modulate the behavior of numerous human cancer cell
types in vitro and in vivo (8–10). For
example, propofol can effectively inhibit cell proliferation and
invasion of cervical cancer, pancreatic cancer (11), gastric cancer (12) and colon carcinoma (10). Propofol treatment also resulted in
the apoptosis of pancreatic cancer cells (11), gastric cancer cells (12), and promyelocytic leukemia cells
(22). Animal studies have shown
that propofol can modulate the immune reaction to exert its
antitumor activity (23). Consistent
with the previous studies (13,14), in
the present study, it was demonstrated that propofol could inhibit
cell proliferation and invasion, and induce cell apoptosis of HCC
cells, indicating that propofol has an anti-HCC effect.
Previous studies have suggested that microRNAs
(miRNAs) are involved in mediating the effects of propofol on
anti-cancers, including miR-199a, miR-142-3p, miR-143 and let-7
(13–15,24,25).
Propofol can influence the expression of these miRNAs, which in
turn are able to regulate their target mRNAs to affect the behavior
of cancer cells. Using miRanda prediction software, it was
indicated that HMGA2 is a potential target of miR-142-3p and let-7.
Based on this, the present study investigated whether propofol
could influence the expression of HMGA2. The results from western
blot analysis demonstrated that HMGA2 protein expression was
significantly inhibited by propofol at concentrations between 5 and
20 µg/ml, and the inhibitory effect of propofol on HMGA2 expression
was dose-dependent.
HMGA2 is an architectural transcriptional regulator
that binds AT-rich DNA sequences. In a prior study, HMGA2 could not
be detected in normal adult tissues (26); however, elevated expression of HMGA2
is found in a variety of human cancers, such as lung cancer,
colorectal cancer and breast cancer (27–29). The
role of HMGA2 in normal and HCC cell growth is well established
(17,18). HMGA2 is a marker for hepatic
progenitor cells. It is expressed in all hepatoblastomas and a
subset of HCCs (17). Expression of
HMGA2 is associated with poor survival in patients with HCC
(30). The present gain-of-function
experiments found that the effect of propofol on HepG2 cell
proliferation, apoptosis and invasion could be attenuated by HMGA2
overexpression. These results suggested that HMGA2 mediates the
effect of propofol on HCC.
Wnt/β-catenin pathway is the downstream pathway of
HMGA2 (21). Aberrant Wnt/β-catenin
signaling has been implicated in the pathogenesis of multiple
tumors (31,32). Snail1 and c-myc are the downstream
effectors of the Wnt/β-catenin signaling (33,34).
Snail1 is involved in the induction of EMT, contributing to the
invasion and migration of cancer cells (35), and c-myc is a transcription factor
that regulates cell cycle, apoptosis and growth (36). Lee et al demonstrated that the
expression of HMGA2 and β-catenin correlated positively in
hepatoblastoma (17). In the present
study, the effect of propofol on Wnt/β-catenin pathway was
investigated. We demonstrated for the first time that propofol
inhibits the Wnt/β-catenin pathway in a dose-dependent manner, as
evidenced by the decreased expression of Wnt3a, β-catenin, Snail1
and c-myc in HepG2 cells following treatment with propofol at
concentrations between 5 and 20 µg/ml. Furthermore, we found that
the effect of propofol on the Wnt/β-catenin pathway was attenuated
by HMGA2 overexpression. These results indicated that HMGA2 was
involved in mediating the effect of propofol on Wnt/β-catenin
pathway in HepG2 cells.
Collectively, the present results provide the first
evidence, to our knowledge, that HMGA2 mediates the effect of
propofol on HepG2 cells. Propofol was able to downregulate the
expression of HMGA2, which inhibited the Wnt/β-catenin pathway,
thus leading to the inhibition of cell proliferation and invasion,
as well as induction of cell apoptosis of HepG2 cells. These
findings suggest a novel molecular mechanism underlying the
therapeutic effect of propofol on HCC. Further studies in
vivo are required to validate this molecular mechanism and its
clinical relevance.
References
|
1
|
Gomes MA, Priolli DG, Tralhão JG and
Botelho MF: Hepatocellular carcinoma: Epidemiology, biology,
diagnosis, and therapies. Rev Assoc Med Bras. 1992.59:514–524.
2013. View Article : Google Scholar
|
|
2
|
Herszényi L and Tulassay Z: Epidemiology
of gastrointestinal and liver tumors. Eur Rev Med Pharmacol Sci.
14:249–258. 2010.PubMed/NCBI
|
|
3
|
Ribatti D, Vacca A, Nico B, Sansonno D and
Dammacco F: Angiogenesis and anti-angiogenesis in hepatocellular
carcinoma. Cancer Treat Rev. 32:437–444. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pang RW and Poon RT: From molecular
biology to targeted therapies for hepatocellular carcinoma: The
future is now. Oncology. 72 Suppl 1:S30–S44. 2007. View Article : Google Scholar
|
|
5
|
Castroagudín JF, Delgado M, Villanueva A,
Bustamante M, Martínez J, Otero E, Tomé S, Martínez SM, Segade FR,
Conde R, et al: Safety of percutaneous ethanol injection as
neoadjuvant therapy for hepatocellular carcinoma in waiting list
liver transplant candidates. Transplant Proc. 37:3871–3873. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gluer AM, Cocco N, Laurence JM, Johnston
ES, Hollands MJ, Pleass HC, Richardson AJ and Lam VW: Systematic
review of actual 10-year survival following resection for
hepatocellular carcinoma. HPB (Oxford). 14:285–290. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou XD: Recurrence and metastasis of
hepatocellular carcinoma: Progress and prospects. Hepatobiliary
Pancreat Dis Int. 1:35–41. 2002.PubMed/NCBI
|
|
8
|
Altenburg JD, Harvey KA, McCray S, Xu Z
and Siddiqui RA: A novel 2,6-diisopropylphenyl-docosahexaenoamide
conjugate induces apoptosis in T cell acute lymphoblastic leukemia
cell lines. Biochem Biophys Res Commun. 411:427–432. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mammoto T, Mukai M, Mammoto A, Yamanaka Y,
Hayashi Y, Mashimo T, Kishi Y and Nakamura H: Intravenous
anesthetic, propofol inhibits invasion of cancer cells. Cancer
Lett. 184:165–170. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miao Y, Zhang Y, Wan H, Chen L and Wang F:
GABA-receptor agonist, propofol inhibits invasion of colon
carcinoma cells. Biomed Pharmacother. 64:583–588. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang ZT, Gong HY, Zheng F, Liu DJ and Dong
TL: Propofol suppresses proliferation and invasion of pancreatic
cancer cells by upregulating microRNA-133a expression. Genet Mol
Res. 14:7529–7537. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang ZT, Gong HY, Zheng F, Liu DJ and Yue
XQ: Propofol suppresses proliferation and invasion of gastric
cancer cells via downregulation of microRNA-221 expression. Genet
Mol Res. 14:8117–8124. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang J, Zhang D, Wu GQ, Feng ZY and Zhu
SM: Propofol inhibits the adhesion of hepatocellular carcinoma
cells by upregulating microRNA-199a and downregulating MMP-9
expression. Hepatobiliary Pancreat Dis Int. 12:305–309. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Wu GQ, Zhang Y, Feng ZY and Zhu
SM: Propofol induces apoptosis of hepatocellular carcinoma cells by
upregulation of microRNA-199a expression. Cell Biol Int.
37:227–232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang J, Shan WF, Jin TT, Wu GQ, Xiong XX,
Jin HY and Zhu SM: Propofol exerts anti-hepatocellular carcinoma by
microvesicle-mediated transfer of miR-142-3p from macrophage to
cancer cells. J Transl Med. 12:2792014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Morishita A, Zaidi MR, Mitoro A,
Sankarasharma D, Szabolcs M, Okada Y, D'Armiento J and Chada K:
HMGA2 is a driver of tumor metastasis. Cancer Res. 73:4289–4299.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee CT, Zhang L, Mounajjed T and Wu TT:
High mobility group AT-hook 2 is overexpressed in hepatoblastoma.
Hum Pathol. 44:802–810. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marijon H, Dokmak S, Paradis V, Zappa M,
Bieche I, Bouattour M, Raymond E and Faivre S:
Epithelial-to-mesenchymal transition and acquired resistance to
sunitinib in a patient with hepatocellular carcinoma. J Hepatol.
54:1073–1078. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tan EJ, Thuault S, Caja L, Carletti T,
Heldin CH and Moustakas A: Regulation of transcription factor Twist
expression by the DNA architectural protein high mobility group A2
during epithelial-to-mesenchymal transition. J Biol Chem.
287:7134–7145. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thuault S, Tan EJ, Peinado H, Cano A,
Heldin CH and Moustakas A: HMGA2 and Smads co-regulate SNAIL1
expression during induction of epithelial-to-mesenchymal
transition. J Biol Chem. 283:33437–33446. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zha L, Zhang J, Tang W, Zhang N, He M, Guo
Y and Wang Z: HMGA2 elicits EMT by activating the Wnt/β-catenin
pathway in gastric cancer. Dig Dis Sci. 58:724–733. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsuchiya M, Asada A, Arita K, Utsumi T,
Yoshida T, Sato EF, Utsumi K and Inoue M: Induction and mechanism
of apoptotic cell death by propofol in HL-60 cells. Acta
Anaesthesiol Scand. 46:1068–1074. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Inada T, Kubo K and Shingu K: Possible
link between cyclooxygenase-inhibiting and antitumor properties of
propofol. J Anesth. 25:569–575. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Su Z, Hou XK and Wen QP: Propofol induces
apoptosis of epithelial ovarian cancer cells by upregulation of
microRNA let-7i expression. Eur J Gynaecol Oncol. 35:688–691.
2014.PubMed/NCBI
|
|
25
|
Ye Z, Jingzhong L, Yangbo L, Lei C and
Jiandong Y: Propofol inhibits proliferation and invasion of
osteosarcoma cells by regulation of microRNA-143 expression. Oncol
Res. 21:201–207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou X, Benson KF, Ashar HR and Chada K:
Mutation responsible for the mouse pygmy phenotype in the
developmentally regulated factor HMGI-C. Nature. 376:771–774. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Langelotz C, Schmid P, Jakob C, Heider U,
Wernecke KD, Possinger K and Sezer O: Expression of
high-mobility-group-protein HMGI-C mRNA in the peripheral blood is
an independent poor prognostic indicator for survival in metastatic
breast cancer. Br J Cancer. 88:1406–1410. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang X, Liu X, Li AY, Chen L, Lai L, Lin
HH, Hu S, Yao L, Peng J, Loera S, et al: Overexpression of HMGA2
promotes metastasis and impacts survival of colorectal cancers.
Clin Cancer Res. 17:2570–2580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sarhadi VK, Wikman H, Salmenkivi K, Kuosma
E, Sioris T, Salo J, Karjalainen A, Knuutila S and Anttila S:
Increased expression of high mobility group A proteins in lung
cancer. J Pathol. 209:206–212. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu L, Wang Z, Lu R and Jiang W: Expression
of high mobility group A2 is associated with poor survival in
hepatocellular carcinoma. Pathol Oncol Res. 18:983–987. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gupta A, Verma A, Mishra AK, Wadhwa G,
Sharma SK and Jain CK: The Wnt pathway: Emerging anticancer
strategies. Recent Pat Endocr Metab Immune Drug Discov. 7:138–147.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rosenbluh J, Wang X and Hahn WC: Genomic
insights into WNT/β-catenin signaling. Trends Pharmacol Sci.
35:103–109. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dai C, Stolz DB, Kiss LP, Monga SP,
Holzman LB and Liu Y: Wnt/beta-catenin signaling promotes podocyte
dysfunction and albuminuria. J Am Soc Nephrol. 20:1997–2008. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Breuhahn K, Longerich T and Schirmacher P:
Dysregulation of growth factor signaling in human hepatocellular
carcinoma. Oncogene. 25:3787–3800. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Baulida J and García de Herreros A:
Snail1-driven plasticity of epithelial and mesenchymal cells
sustains cancer malignancy. Biochim Biophys Acta. 1856:55–61.
2015.PubMed/NCBI
|
|
36
|
Vita M and Henriksson M: The Myc
oncoprotein as a therapeutic target for human cancer. Semin Cancer
Biol. 16:318–330. 2006. View Article : Google Scholar : PubMed/NCBI
|