Introduction
The heart is an uncommon organ for primary tumor
development. Primary rhabdomyosarcoma of the heart is a rare tumor;
however, it is the second most common cardiac sarcoma (1). Rhabdomyosarcoma arises in the right
atrium and is often complicated with heart failure, which is
serious and difficult to control (2). Early recognition and therapeutic
intervention is extremely important in the prevention of heart
failure. Surgery is the mainstay of treatment for non-metastatic
disease. If surgical intervention fails to completely resect the
tumor (3,4), radiotherapy is essential and effective;
however, the therapeutic success of chemotherapy and radiotherapy
is poor for metastatic disease (5,6).
Therefore, the improvement of radiotherapy success for
rhabdomyosarcoma has become a major focus of research.
Proteins of the centrosomal protein (CEP) family are
critical components of centrosomes and have vital roles in the
control of cell cycle progression. This protein family consists of
31 proteins, one of which is CEP164. CEP164 encodes a 180 kDa
protein. CEP164 may be phosphorylated by ataxia telangiectasia
mutated (ATM) kinase and Rad3-related protein (ATR) kinase in the
DNA damage response (DDR) pathway and has a key role in the gap
2/mitosis (G2/M) checkpoint (7,8).
Research has demonstrated that mutations in CEP164 are one cause of
nephronophthisis-related ciliopathies (NPHP-RC) (9). In the DDR, CEP164 is rapidly localized
to nuclear foci. Research has demonstrated that knockdown of CEP164
induces cell sensitivity to DNA-damaging agents and CEP164
knockdown in zebrafish results in deregulated DDR and an NPHP-RC
phenotype (4). Additionally, CEP164
modulates mediator of DNA damage checkpoint protein 1 (MDC1) and
checkpoint kinase 1 (CHEK1) to maintain genomic stability (10).
The importance of CEP164 in the DDR makes it a
notable target for enhancing the radiosensitivity of cells;
however, the effect of combining CEP164 inhibition with irradiation
(IR) in rhabdomyosarcoma cells is unknown. In the present study,
the role of CEP164 in the cell cycle, and the cell viability of the
rhabdomyosarcoma A-204 and A-673 cells lines following treatment
with IR were investigated. It was demonstrated that CEP164
expression levels are increased following treatment with IR and
depletion of CEP164 enhances cellular sensitivity to radiation,
resulting in decreased cell viability, promotion of apoptosis and
G2/M cell cycle arrest. In conclusion, the present study
demonstrated that CEP164 may be a candidate target gene for
rhabdomyosarcoma radiotherapy.
Materials and methods
Cell culture, small interfering
(si)RNA transfection and radiation exposure
Human rhabdomyosarcoma A-204 and A-673 cells
(American Type Culture Collection, Manassas, VA, USA) were cultured
in McCoy's 5A (Modified) Medium supplemented with 10% fetal bovine
serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA), incubated at 37°C in a 95% humidified incubator (5%
CO2) and irradiated with doses of 2, 4 or 6 Gy. Cells
were treated with Cobalt-60 γ-rays at 2 Gy/min in the Beijing
Radiation Center (Beijing Academy of Science and Technology,
Beijing, China). Cells were cultured to 80% confluence and were
subsequently transfected with siRNA against the negative control or
against CEP164 (Ambion; Thermo Fisher Scientific, Inc.). All
transfection was performed using Lipofectamine® RNAiMAX
reagent (Thermo Fisher Scientific, Inc.). Cells were exposed to IR
24 h after target gene knockdown.
Western blot analysis
A-204 cells were lysed in RIPA lysis and extraction
buffer containing protease and phosphatase inhibitor (all Pierce;
Thermo Fisher Scientific, Inc.) for 20 min at 4°C. Protein
concentration was measured using a BCA Protein Assay kit (Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
A total of 30 µg of protein was separated by 10% Bis-Tris gels
(Thermo Fisher Scientific, Inc.) and transferred onto
nitrocellulose membranes for western blot analysis. Following
washing three times with Tris-buffered saline-Tween 20, the
membranes were blocked with 5% nonfat milk for 60 min and were
subsequently incubated with primary anti-CEP164 (1:1,000; GTX85298;
GeneTex, Inc., Irvine, CA, USA) or anti-β-actin antibodies
(1:1,000; ab8227; Abcam, Cambridge, UK) overnight at 4°C. Following
two washes with washing buffer (0.5% Tween in PBS), the membranes
were incubated with horseradish peroxidase-conjugated secondary
anti-rabbit antibody (1:3,000; ab6721; Abcam) for 1 h at room
temperature. Following this, protein bands were detected using
chemiluminescence liquid (Pierce; Thermo Fisher Scientific, Inc.),
according to the manufacturer's instructions, and analyzed using
ImageJ software (ImageJ 2x version 2.1.4.6, Bio-Rad Laboratories,
Inc., Hercules, CA, USA).
Cell cycle assay
A total of 1×106 cells were harvested and
resuspended in 1 ml McCoy's 5A (Modified) Medium supplemented with
10% FBS. Cells were stained with 2 µl Vybrant DyeCycle Green Stain
and incubated at 37°C for 30 min. The cell cycle was analyzed using
flow cytometry using 488 nm excitation and green emission.
Cell viability assays
Following treatment with IR, cells were mixed with
10% volume of alamarBlue reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and incubated for 30 min at 37°C, protected from
direct light. Results were recorded using florescence at 570/585 nm
(excitation/emission).
Cell apoptosis assay
Cells were harvested and washed twice with cold
phosphate-buffered saline. Subsequently, cells were resuspended in
1X binding buffer (BD Biosciences, San Jose, CA, USA) at a
concentration of 1×106 cells/ml and stained with PE
Annexin V with 7-AAD (BD Biosciences) in the dark for 15 min at
room temperature. Samples were analyzed using flow cytometry.
Statistical analysis
Statistical analysis was performed using Microsoft
Excel 2010 (Microsoft Corp., Redmond, WA, USA). Data are presented
as the mean ± standard error of the mean of three independent
experiments. Differences in mean values between groups were
determined by Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
γH2AX foci formation
A-204 cells were seeded onto glass slides. Following
transfection with siRNAs, the cells were irradiated with 4 Gy for
30 min, washed twice in PBS for 5 min and subsequently fixed for 10
min at room temperature using 4% paraformaldehyde. Cells were
washed twice in PBS for 5 min, permeated for 10 min at room
temperature using permeabilisation buffer (0.2% TritonX-100 in PBS)
and washed twice more for 5 min. Cells were subsequently blocked
for 1 h at room temperature with blocking buffer (0.05% FBS in
PBS), and incubated with anti-γH2AX (1:200; Merck KGaA, Darmstadt,
Germany; 05-636-AF488) antibody overnight at 4°C. The cells were
washed twice more in washing buffer (0.05% Tween in PBS) and the
coverslips were mounted with mounting medium (glycerol), stained
with DAPI and detected using a fluorescent microscope (Leica
Microsystems GmbH, Wetzlar, Germany). γH2AX foci were analyzed
using Image Pro Plus 6.0 software (Media Cybernetics, Inc.,
Rockville, MD, USA).
Results
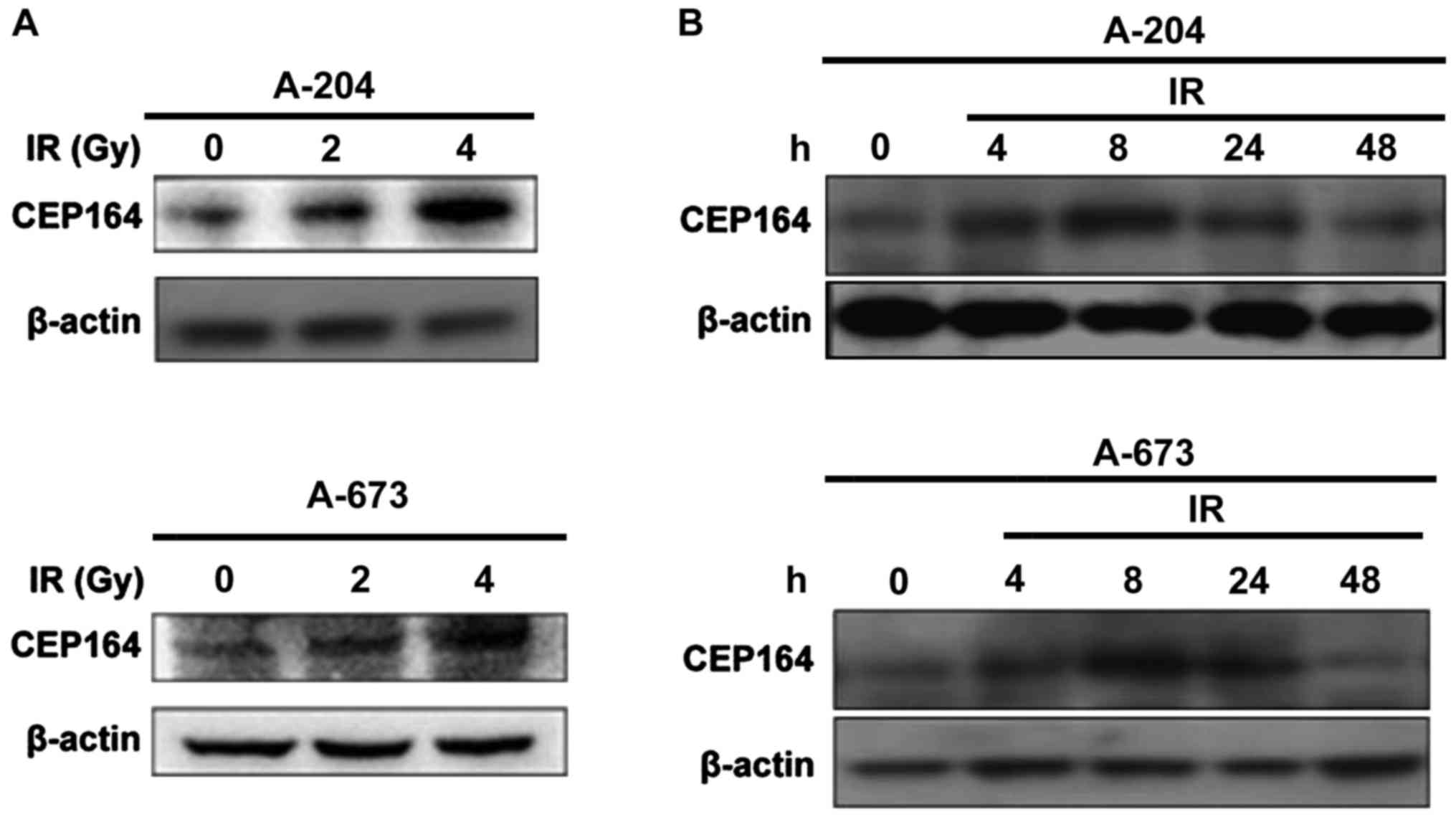
Radiation induces CEP164 protein
expression
To determine whether CEP164 is involved in the DDR,
cells were γ-irradiated with 2, 4, 6 Gy and, following this, CEP164
expression levels were measured. Results demonstrated that CEP164
protein expression levels increased following IR with 2 and 4 Gy
and the expression levels peaked at 24 h following treatment with 2
Gy IR in A-204 and A-673 cells, respectively (Fig. 1).
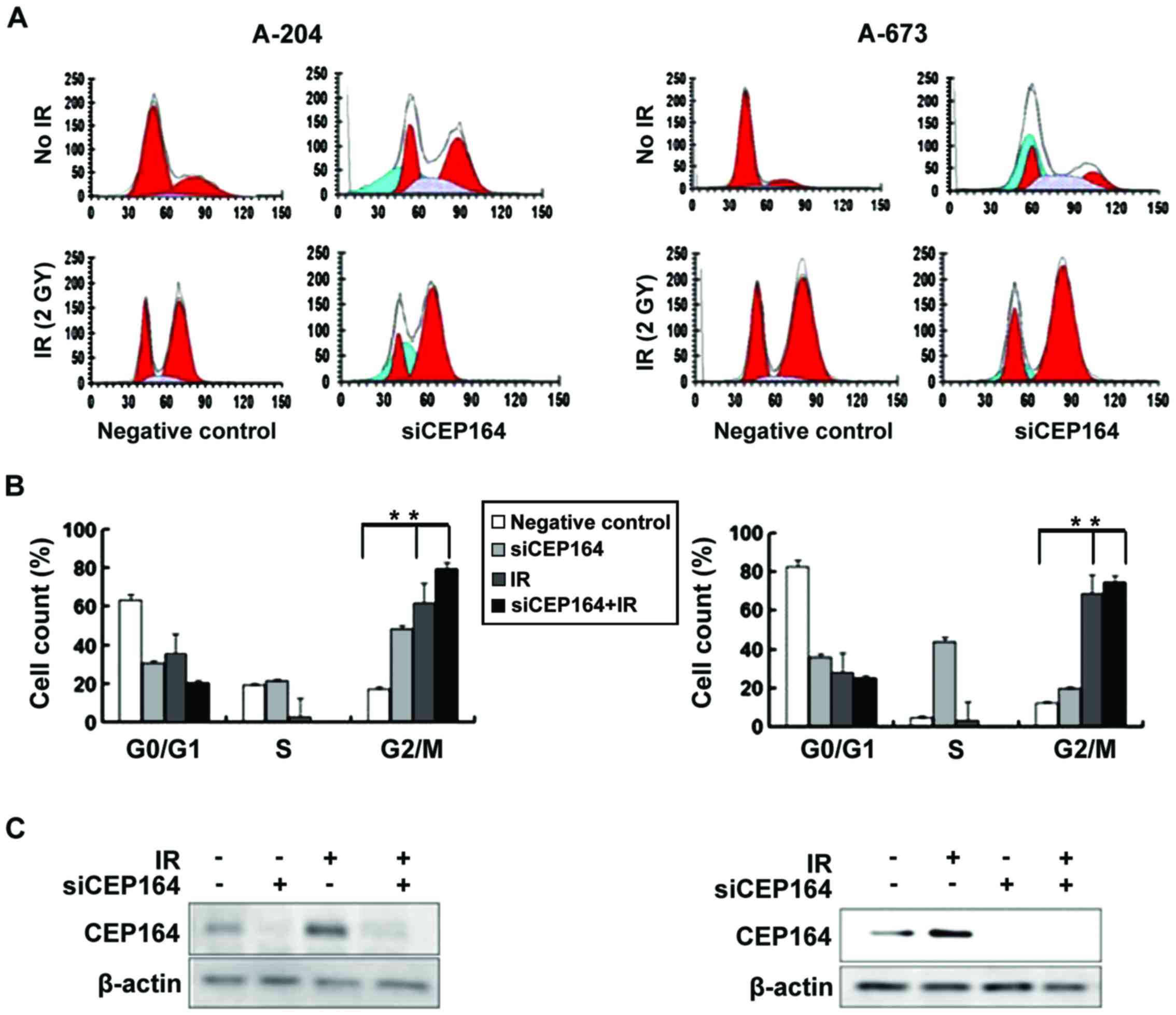
CEP164 inhibition induces G2/M arrest
of the cell cycle
In order to determine whether CEP164 has a
regulatory role in cell cycle progression, siRNA was utilized to
knockdown CEP164 expression levels in A-204 and A-673 cells and,
following 24 h, G2/M cell cycle arrest was measured. Results
demonstrated that CEP164 delayed S-phase exit. Furthermore, to
determine whether inhibition of CEP164 enhances the
radiosensitivity of A-204 and A-673 cells, the cell cycle of
CEP164-depleted cells following treatment with IR was analyzed.
Treatment of A-204 cells with 2 Gy of IR induced entry into G2/M
phase and a lower population in the synthesis (S) phase; however,
combined radiation and CEP164 inhibition resulted in G2/M cell
cycle arrest. These experiments were also conducted on A-673 cells,
and similar results were found in CEP164 silencing A-673 cells
(Fig. 2).
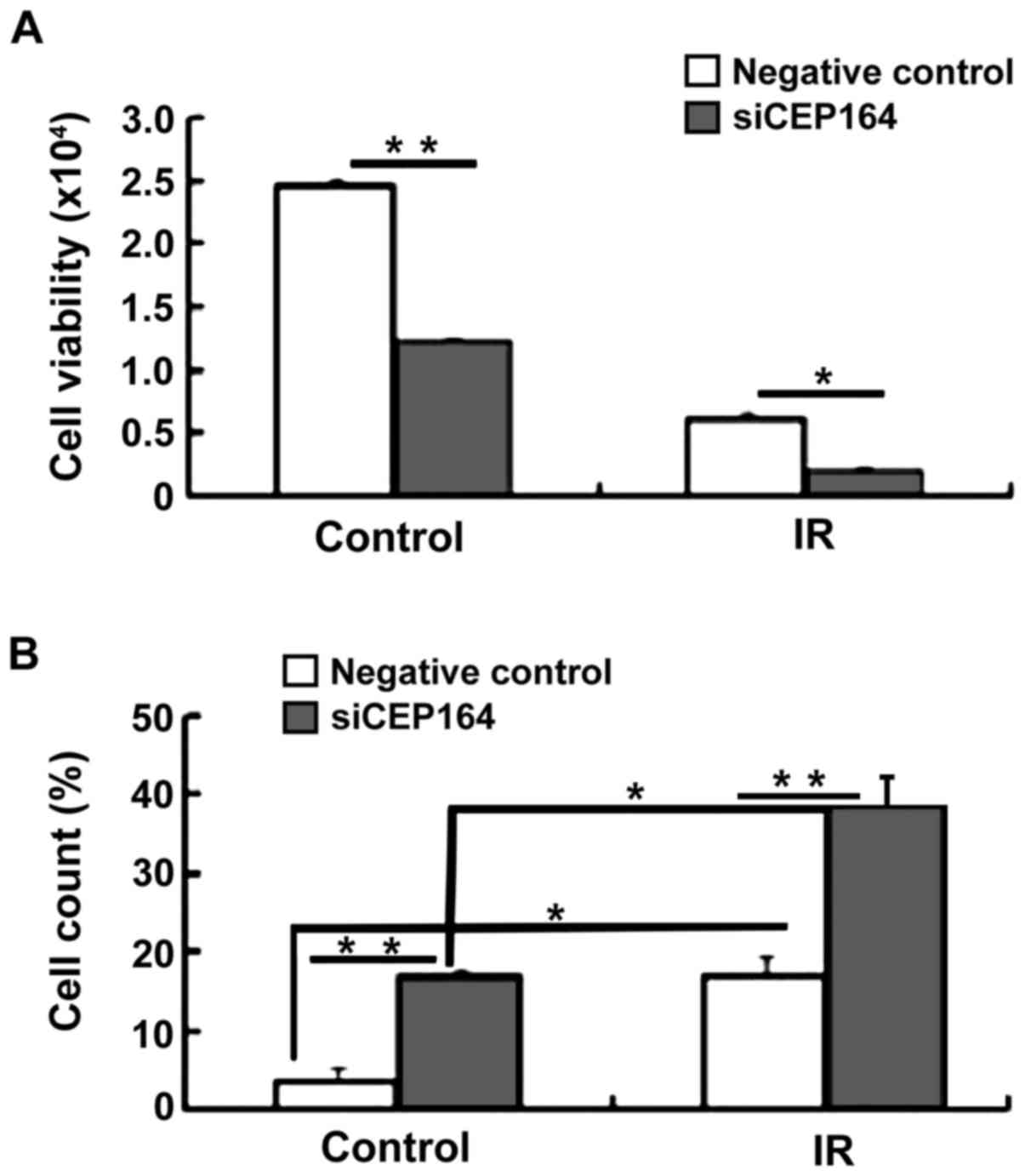
CEP164 silencing increases cell
apoptosis and decreases cell viability upon IR
Following the result that CEP164 inhibition is able
to arrest cells in the S phase of the cell cycle, colony formation
assays were conducted in order to determine whether CEP164 has a
role in cell proliferation and apoptosis. Cell viability assays
indicated that CEP164 knockdown significantly decreased cell
viability following treatment with IR compared with the negative
control IR-treatment group (P<0.05). In order to determine
whether CEP164 knockdown enhances IR-induced apoptosis, four
different experimental conditions were investigated, including: No
IR and no CEP164 knockdown; IR only (4 Gy); CEP164 knockdown only;
and combined CEP164 knockdown and IR (4 Gy). Results demonstrated
that the combined treatment of CEP164-silencing and IR produced a
significant increase in apoptosis compared with either of the
treatments alone (P<0.05, P<0.01; Fig. 3). These results indicate that CEP164
is able to facilitate cellular proliferation and inhibit radiation
injury.
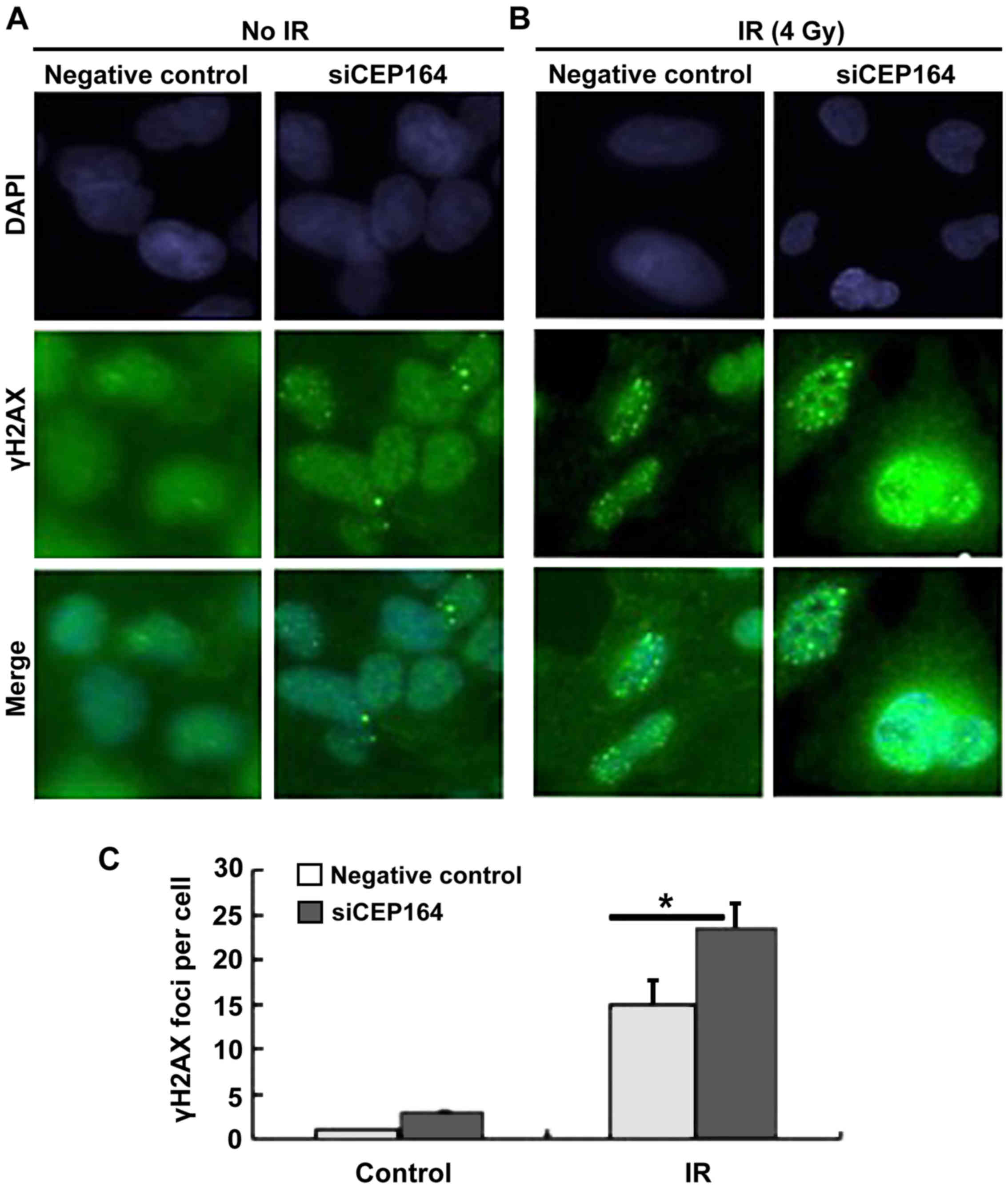
CEP164 knockdown upregulates
IR-induced γH2A histone family member X (γH2AX) foci formation
In order to confirm whether CEP164 is able to
enhance the DDR, the γH2AX foci assembly in A-204 cells with or
without CEP164 silencing was investigated. It was demonstrated that
CEP164 depletion significantly increased γH2AX foci assembly
compared with the negative control cells during 4 Gy IR
(P<0.05), indicating that CEP164 repression may enhance the
recruitment of γH2AX and its associated repair proteins to DNA
damage sites (Fig. 4).
Discussion
Cardiac tumors are classified as primary benign and
malignant tumors if they arise in the heart, or as secondary tumors
if they metastasize to the heart. In children, 90% of cardiac
tumors are benign, whereas in adults 75% of cardiac tumors are
benign (1,11–13).
Rhabdomyosarcoma is the most common soft tissue malignant tumor in
infants and children (14). Surgical
resection of rhabdomyosarcoma is technically challenging and many
malignant tumors cannot be resected completely. Chemotherapy and
radiation therapy are able to relieve symptoms and prolong
survival. A multi-treatment approach, including chemotherapy,
radiation and novel, evolving approaches, such as gene therapy, may
provide a more effective palliative and curative result.
CEP164 protein expression levels and activity are
up-regulated in various types of cancer, and the protein is often
overexpressed in tumors and is associated with poor prognosis
(10). Additionally, CEP164 has a
key role in primary cilia formation, which is a marker for distal
appendages on mature centrioles or basal bodies (3,6). As a
novel mediator protein in DDR, CEP164 is required for the DNA
damage-activated signaling cascade (4,10).
CEP164 interacts with both ATR and ATM, which are able to
phosphorylate CEP164 under conditions of DNA damage and replication
stress. Following phosphorylation, CEP164 is able to phosphorylate
H2AX and CHEK2 (15). In the present
study, the association of CEP164 knockdown with radiosensitivity in
the cells was investigated. Results demonstrated that CEP164
protein expression levels increased following IR and inhibition of
CEP164 resulted in the increased radiosensitivity of
rhabdomyosarcoma cells. Furthermore, the results demonstrated that
CEP164 promotes cell survival and decreases cell viability upon IR,
indicating that CEP164 has an important role in cellular
proliferation in response to cellular stress.
CEP164 is required for genomic stability and CEP164
expression is cell cycle stage-dependent, being expressed at the
end of the S phase and the beginning of the G2/M phase (16). CEP164 is essential for G2/M
checkpoint activation through the phosphorylation of CHEK proteins.
The results of the present study indicated that CEP164 engaged in
cellular G2/M arrest, demonstrating that CEP164 has a function in
cell cycle switching.
In conclusion, the present study demonstrated that
CEP164 deletion is involved in the IR-induced cellular response and
enhances the radiosensitivity of rhabdomyosarcoma cells. This,
therefore, indicates that CEP164 may be a potential target to
improve the outcome of radiotherapy in heart rhabdomyosarcoma
treatment.
References
|
1
|
Yilmaz M, Kehlibar T, Arslan IY, Yilmaz
HY, Tarhan IA and Ozler A: A case of primary cardiac
rhabdomyosarcoma with surgical removal and mitral valve repair.
Heart Surg Forum. 16:E164–E166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Abraham R: Cell cycle checkpoint signaling
through the ATM and ATR kinases. Genes Dev. 15:2177–2196. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Čajánek L and Nigg EA: Cep164 triggers
ciliogenesis by recruiting Tau tubulin kinase 2 to the mother
centriole. Proc Natl Acad Sci USA. 111:E2841–E2850. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chaki M, Airik R, Ghosh AK, Giles RH, Chen
R, Slaats GG, Wang H, Hurd TW, Zhou W, Cluckey A, et al: Exome
capture reveals ZNF423 and CEP164 mutations, linking renal
ciliopathies to DNA damage response signaling. Cell. 150:533–548.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen K: Rhabdomyosarcoma in an adult
presenting with nodal metastasis: A pitfall in fine-needle
aspiration cytology of lymph nodes. Diagn Cytopathol. 32:303–306.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Graser S, Stierhof YD, Lavoie SB, Gassner
OS, Lamla S, Le Clech M and Nigg EA: Cep164, a novel centriole
appendage protein required for primary cilium formation. J Cell
Biol. 179:321–330. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo L, Wang ZY, Zou YB and Bi LR: Primary
cardiac embryonal rhabdomyosarcoma: Report of a case. Zhonghua Bing
Li Xue Za Zhi. 42:621–622. 2013.(In Chinese). PubMed/NCBI
|
|
8
|
Ma J, Sun JP, Chen M, Zhang L, Xu N, Wang
J, Pui-wai Lee A and Yu CM: Left atrial rhabdomyosarcoma.
Circulation. 129:e503–505. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Merlino G and Helman LJ:
Rhabdomyosarcoma-working out the pathways. Oncogene. 18:5340–5348.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pan YR and Lee EY: UV-dependent
interaction between Cep164 and XPA mediates localization of Cep164
at sites of DNA damage and UV sensitivity. Cell Cycle. 8:655–664.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Paulino AC and Okcu MF: Rhabdomyosarcoma.
Curr Probl Cancer. 32:7–34. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sivasubramaniam S, Sun X, Pan YR, Wang S
and Lee EY: Cep164 is a mediator protein required for the
maintenance of genomic stability through modulation of MDC1, RPA,
and CHK1. Genes Dev. 22:587–600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang S, Gao Y, Zhao H and Cao D: Cardiac
embryonal rhabdomyosarcoma in an adult. Eur J Cardiothorac Surg.
45:e2332014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Raphael S, Yusuf I and Imam I: Childhood
rhabdomyosarcoma in Kano, Nigeria: A retrospective analysis of 52
cases. Niger J Med. 24:32–36. 2015.PubMed/NCBI
|
|
15
|
Nigg EA, Čajánek L and Arquint C: The
centrosome duplication cycle in health and disease. FEBS Lett.
588:2366–2372. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Slaats GG, Ghosh AK, Falke LL, Le Corre S,
Shaltiel IA, van de Hoek G, Klasson TD, Stokman MF, Logister I,
Verhaar MC, et al: Nephronophthisis-associated CEP164 regulates
cell cycle progression, apoptosis and epithelial-to-mesenchymal
transition. PLoS Genet. 10:e10045942014. View Article : Google Scholar : PubMed/NCBI
|