Introduction
Endothelial progenitor cells (EPCs) are a population
of cells that participate in vessel formation in both physiological
and pathological processes, and demonstrate characteristics of both
endothelial and progenitor cells (1). EPCs have great potential as a source of
cells for the repair of vasculogenic injuries (2). Endothelial colony-forming cells
(ECFCs), which typically possess a cobblestone appearance and are
also referred to as blood outgrowth endothelial cells or late
outgrowth cells, are a rare putative type of EPC that have the
ability to produce endothelial cells and contribute to the
functional endothelial lining of injured vascular structures in
vivo at the colony level (3).
EPCs were first isolated from adult human peripheral blood in 1997
(3,4). Since then, they have been successfully
isolated from bone marrow (5),
peripheral blood (6) and umbilical
cord (UC) blood (7). Bone marrow is
considered the original source of EPCs. Depletion of or increases
in the number of EPCs in peripheral blood has been reported in
various pathologic conditions, such as cardiovascular diseases,
hypertension, type 2 diabetes mellitus, rheumatoid arthritis, aging
and hematological diseases (8,9). EPCs
have been used to repair ischemic or damaged cardiac tissue in
animal models (8,9), and these cells have also been explored
for the creation of living blood vessels (10,11). The
promising results obtained in these studies suggest that these
cells have the potential to be employed in clinical trials
(12).
Although transplantation of autologous bone
marrow-derived or peripheral blood-derived ECFCs has been
demonstrated to be safe, the utility of these cells is limited due
to the significant drop in cell number and
proliferative/differentiation capacity with age (13). A study by Mandraffino et al
(14) reported that, in elderly
patients, the peripheral cell count is not necessarily weakened and
the cluster of differentiation (CD)34+ cells maintain
their role in predicting mortality. CD34+ cells may
therefore be considered as a biomarker of longevity, not EPCs. Two
possible ideal sources of ECFCs are cord blood and UC, which were
discarded as medical waste in the past (15). However, the number of nucleated cells
in cord blood is limited, which is thought to be a serious
limitation to their utility for transplantation (16). In addition, a previous study
demonstrated that human placenta-derived ECFCs have greater
vasculogenic potential than cord blood-derived ECFCs in vivo
(9).
Numerous previous studies that have aimed to
identify EPCs have focused on simultaneous expression of CD34,
CD117, CD133, CD105, CD144, CD184, CD309 [kinase insert domain
receptor (KDR) or vascular endothelial growth factor receptor 2
(VEGFR2)], acetylated low-density lipoprotein and various plant
lectins (14,17–19).
Isolating sufficient EPCs is a major limitation to
clinical applications, as the number of EPCs obtained from bone
marrow, peripheral blood, adipose tissue and cord blood is limited.
To the best of our knowledge, there are no published studies aimed
at obtaining EPCs from the umbilical vein by direct enzymatic
digestion. The purpose of the present investigation was to isolate
and characterize the population of resident ECFCs from UCs and
explore it as an optimized source of ECFCs.
Materials and methods
Isolation and culture of human
UC-ECFCs
A total of 10 human UCs were obtained between
January 2012 and June 2015 from the General Hospital of the Chinese
People's Liberation Army (Beijing, China). The present study was
conducted in accordance with the Declaration of Helsinki, with
approval from the Ethics Committee of the Affiliated Hospital of
Academy of Military Medical Sciences (Beijing, China). All
newborns' mothers provided written informed consent.
The median cell yield was 4.2×105
cells/cm of UC [number of isolated cells (mean ± standard deviation
(SD)): 5.22×105±2.09×105 cells/10 cm; n=10;
length of UC (mean ± SD): 20.67±2.75 cm; n=10]. The characteristics
and functions, including growth kinetics, cell cycle,
colony-forming ability and tube formation capability, of the
isolated cells were similar among all samples. UCs were obtained by
cesarean section after normal deliveries and were flushed
repeatedly with phosphate-buffered saline (PBS; pH=7.0) containing
2% gentamicin (Thermo Fisher Scientific, Inc., Waltham MA, USA).
Following the removal of the residual blood from the UC
(particularly the umbilical vein cavity), the umbilical vein was
injected with 5–10 ml 0.1% collagenase type II (Gibco; Thermo
Fisher Scientific, Inc.) with dual-port ligation. Subsequently, the
UCs were placed in containers with PBS and incubated for 1 h at
37°C. The digested umbilical vein was washed five times, for 2 min
with PBS and the digested cells were collected by centrifugation at
256 × g at 4°C for 10 min. The resuspended cells were plated at a
density of 2×104 cells/cm2 in
fibronectin-coated T75 culture flasks containing
complete endothelial cell growth medium (EGM)-2 medium supplemented
with 10% fetal bovine serum (FBS; Lonza, Basel, Switzerland) and
incubated in a humidified incubator at 37°C under 5%
CO2. After 6 days, the medium was replaced to remove
non-adherent cells and debris. The EPCs may be further purified by
attachment-changing culture methods and subculturing for three
passages, which eliminates contamination with digested smooth
muscle cells for the following two reasons: First, the smooth
muscle cells do not adhere to the culture container; and second,
the markers detected by flow cytometry and reverse
transcription-polymerase chain reaction in the present study
(RT-PCR) are only expressed in EPCs. Adherent cells were passaged
at a ratio of 1:3. ECFCs clustered as colonies after 5–22 days.
ECFCs from passage three were used for the following experiments.
Four replicates were performed.
RT-PCR
Total RNA was extracted from 5×106 ECFCs
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
and reverse transcribed using a RevertAid First-Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. To evaluate the gene expression of human
endothelial-nitric oxide synthase (ec-NOS), Fms related
tyrosine kinase 1 (Flt-1), KDR, vascular endothelial
(VE)-cadherin, CD31, CD34, tyrosine
kinase with immunoglobulin (Ig) and epidermal growth factor
homology domains-2 (Tie-2) and von Willebrand factor
(vWF) in ECFCs, cDNA was amplified using Premix Ex Taq
(Shanghai Sangon Biological Engineering Co., Ltd., Shanghai, China)
and sequence-specific primers (Table
I) that were previously described (1–3,13). GAPDH was used as a control.
The PCR program was as follows: 95°C for 3 min, followed by 35
cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C for 45 sec, and
a final step of 72°C for 5 min. The amplified samples were run on a
1% agarose gel and images were captured using the Q550CW image
acquisition and analysis system (Leica Microsystems, Inc., Buffalo
Grove, IL, USA). Four replicates were performed.
 | Table I.Primers for human endothelial
colony-forming cells. |
Table I.
Primers for human endothelial
colony-forming cells.
| Gene | Primer
direction | Primer
sequence | Amplicon, bp | (Refs.) |
|---|
| CD31 | Forward |
5′-GCTGTTGGTGGAAGGAGTGC-3′ | 645 | (1) |
|
| Reverse |
5′-GAAGTTGGCTGGAGGTGCTC-3′ |
|
|
| CD34 | Forward |
5′-CTAGCCTTGCAACATCTCCC-3′ | 409 | (3) |
|
| Reverse |
5′-GAATAGCTCTGGTGGCTTGC-3′ |
|
|
| KDR | Forward |
5′-CAACAAAGTCGGGAGAGGAG-3′ | 819 | (1) |
|
| Reverse |
5′-ATGACGATGGACAAGTAGCC-3′ |
|
|
| Flt-1 | Forward |
5′-AGCAAGTGGGAGTTTGC-3′ | 617 | (2) |
|
| Reverse |
5′-AGGTCCCGATGAATGC-3′ |
|
|
| VE-cadherin | Forward |
5′-AAGACATCAATGACAACTTCC-3′ | 594 | (2) |
|
| Reverse |
5′-CCTCCACAGTCAGGTTATACC-3′ |
|
|
| vWF | Forward |
5′-GAGGCTGAGTTTGAAGTGC-3′ | 477 | (2) |
|
| Reverse |
5′-CTGCTCCAGCTCATCCAC-3′ |
|
|
| Tie-2 | Forward |
5′-TGTTCCTGTGCCACAGGCTG-3′ | 312 | (13) |
|
| Reverse |
5′-CACTGTCCCATCCGGCTTCA-3′ |
|
|
| ec-NOS | Forward |
5′-AAGACATTTTCGGGCTCACGCTGCGCACCC-3′ | 548 | (1) |
|
| Reverse |
5′-TGGGGTAGGCACTTTAGTAGTTCTCCTAAC-3′ |
|
|
| GAPDH | Forward |
5′-TGAAGGTCGGAGTCAACGGATTTG-3′ | 983 | (1) |
|
| Reverse |
5′-CATGTGGGCCATGAGGTCCACCAC-3′ |
|
|
Growth kinetics assay of human
UC-ECFCs
Growth of ECFCs during passage three was plotted.
Adherent cells were harvested at 80% confluence by digestion with
0.25% trypsin/EDTA (Thermo Fisher Scientific, Inc.). Subsequently,
mononuclear cells were seeded in 24-well tissue culture plates at
2×104 cells/cm2 in 0.5 ml complete EGM-2
supplemented with 10% FBS, which was replaced every 4 days. The
experiment continued for 7 days, and the number of living cells in
three wells was evaluated by 0.4% trypan blue staining
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) after
trypsinization at 4°C for 24 h, followed by observation under BX61
light microscope (Olympus Corp., Tokyo, Japan). Subsequently, the
mean cell number was calculated and growth curves were generated.
Four replicates were performed.
Cell cycle analysis
Human UC-ECFCs in the logarithmic phase were fixed
with precooled 75% ethanol at 4°C for 1 h after trypsinization and
digested with RNase A (10 µg/ml; Fuzhou Maixin Biotechnology
Development Co., Ltd., Fuzhou, China) at 37°C for 30 min. Treated
cells were incubated with propidium iodide (50 µg/ml) for 5 min at
4°C in the dark. Cells were analyzed by flow cytometry using a
FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes, NJ,
USA). The procedure was performed as previously described (20). Four replicates were performed.
Flow cytometry
All monoclonal antibodies used in the present
experiment were purchased from BD Pharmingen (San Diego, CA, USA).
ECFCs (1×105 cells/well) were separately labeled with
eight antibodies, including fluorescein isothiocyanate
(FITC)-conjugated mouse anti-human CD90 (cat no. 555595),
FITC-mouse anti-human human leukocyte antigen-antigen D related
protein (HLA-DR; cat no. 555560), phycoerythrin (PE)-conjugated
mouse anti-human CD31 (cat no. 560983), PE-mouse anti-human CD34
(cat no. 555822), PE-mouse anti-human CD73 (cat no. 550257),
PE-mouse anti-human CD105 (cat no. 560839), PE-mouse anti-human
VEGRF2 (cat no. 560872) and a peridinin-chlorophyll-protein
complex-conjugated mouse anti-human CD45 (cat no. 564106). Each
antibody used at a dilution of 1:10. Incubation with antibodies was
performed for 30 min at room temperature in the dark. After washing
2–3 times (5 min each time) with PBS, cells were analyzed by flow
cytometry using a FACSCalibur flow cytometer. Data were quantified
using CELLQuest software (version no. 5.1; BD Biosciences). The
procedure was performed as previously described (21). Four replicates were performed.
Colony-forming assay
Colony-forming assays were performed in triplicate
as previously described (22). Cells
(1×103 cells/plate) were incubated in 6-cm cell culture
dishes with 2 ml DF-12 culture medium (Lonza) for 14 days at 37°C
and 5% CO2. After fixing in methanol at 37°C for 5 min,
cells were stained with 0.5% crystal violet (Sigma-Aldrich; Merck
KGaA) and observed under a BX61 microscope. A cell cluster that
contained >50 cells was considered a colony. Four replicates
were performed.
Matrigel assay
In vitro tube formation was assayed with BD
Matrigel basement membrane matrix (BD Biosciences), according to
the manufacturer's instructions. Matrigel basement membrane matrix
(300 µl/well) was added to a precooled 24-well plate and then
incubated at 37°C for 30 min. Subsequently, 300 µl UC-ECFCs from a
monolayer (at 70–80% confluence, as recommended) was added to each
well. After 24 h of culture at 37°C, capillary tube formation was
assessed by BX61 microscope.
1,1′-dioctadecyl-3,3,3′,3′-tetra-methyl-indocarbocyanine
perchlorate-labeled acetylated low-density lipoprotein (DiL-ac-LDL)
uptake and FITC-Ulex europaeus (UEA)-1 binding assays
To assess the uptake of DiL-ac-LDL (Molecular
Probes; Thermo Fisher Scientific, Inc.) by EPCs, the cells were
seeded in 24-well tissue culture plates at 1×105
cells/cm2 in 1 ml of DF-12 culture medium containing
DiL-ac-LDL (10 µg/ml) at 37°C for 24 h. Following analysis by
fluorescence microscopy, cells were fixed with 4% paraformaldehyde
at 4°C for 20 min, washed twice (2 min each time) with PBS and
incubated in PBS containing 10 µg/ml plant lectin from U.
europaeus conjugated to FITC (FITC-UEA-1; Sigma-Aldrich; Merck
KGaA) at room temperature for 1 h. Four replicates were
performed.
Assessment of cell-surface vWF
expression
Immunofluorescence was used to detect cell-surface
expression of vWF, which is an endothelial cell-specific gene.
Cells (1×103 cells/well) were seeded on coverslips for 3
days, and then fixed in 1% paraformaldehyde at 4°C for at least 10
min. After being washed twice (2 min each time) with PBS, cells
were permeabilized with 0.1% Triton X-100 (Sigma-Aldrich; Merck
KGaA) four times, 5 min each time. Membranes were blocked with goat
serum (cat no. SL038; dilution 1:50; Beijing Solarbio Science &
Technology Co., Ltd., Beijing, China) at 37°C for 10 min.
Subsequently, cells were incubated with 100 µl mouse anti-human vWF
antibody (cat no. sc-516102; dilution 1:50; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) for 1 h at 37°C. After
washing three times (2 min each time) with PBS, the cells were
incubated with the secondary antibody, FITC-conjugated AffiniPure
goat anti-mouse IgG (H+L) antibody (cat no. 127-095-099; dilution
1:50; Jackson Laboratory, Ben Harbor, ME, USA), at 37°C for 1 h.
Nuclear counterstaining was performed with 1.5 µg/ml DAPI (Biotium,
Inc., Hayward, CA, USA) for 30 min at 4°C. Immunofluorescence was
observed under a fluorescence microscope. Four replicates were
performed.
Statistical analysis
All experiments were repeated at least three times.
Data were presented as the mean ± standard deviation. SPSS 13.0
software (SPSS, Inc., Chicago, IL, USA) was used for all
statistical analyses. The significance of differences between mean
values was assessed using Student's t-tests. P<0.05 was
considered to indicate a statistically significant difference.
Results
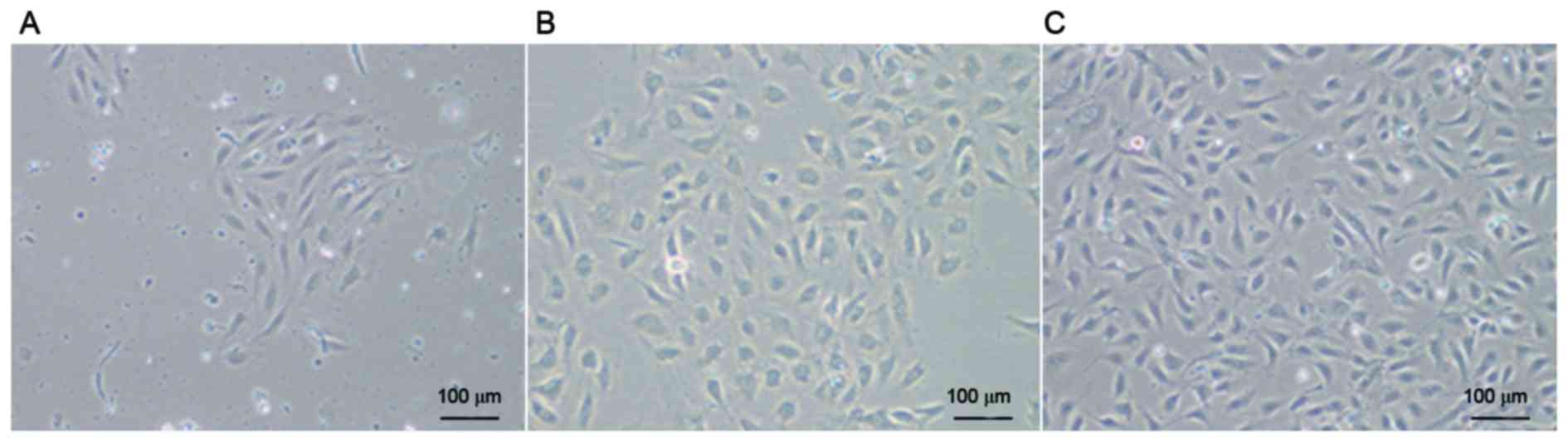
Morphology of human UC-ECFCs
After culturing for 3–7 days, some cells started to
adhere to the wall (Fig. 1A). After
continuous culture for 7–10 days, cells became confluent and formed
colonies (Fig. 1B), with a
cobblestone appearance, which is typical of ECFCs. There were no
marked changes in cell morphology during passage for 10 generations
(Fig. 1C).
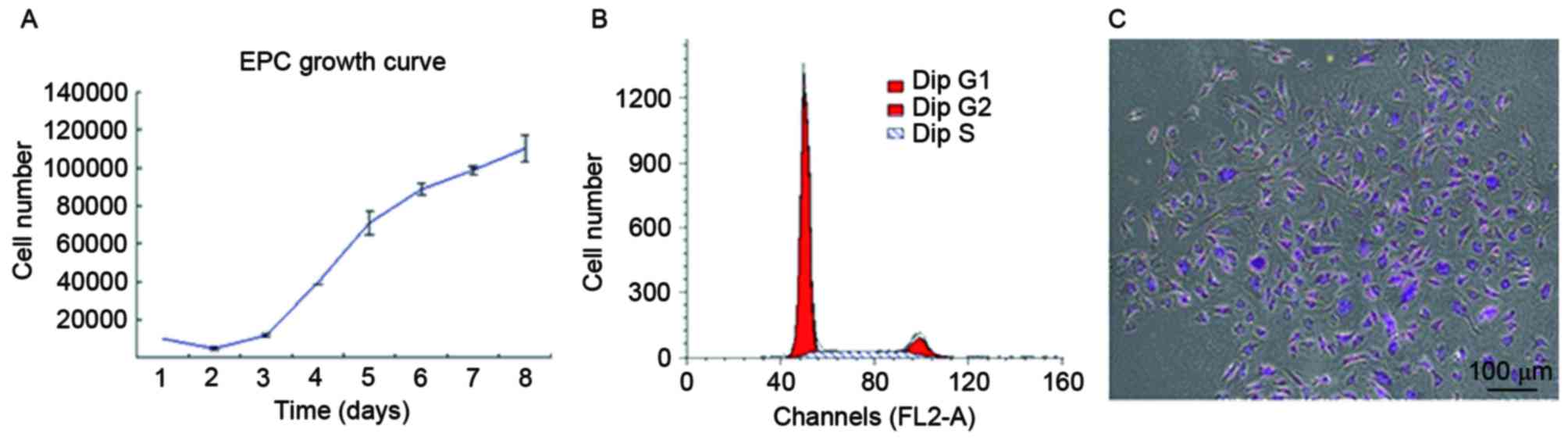
Human UC-ECFC proliferation
To quantitatively assess proliferative potential,
the growth of UC-derived ECFCs was plotted (Fig. 2A). The results demonstrated a basic
cell growth curve, strikingly divided into an incubation period,
logarithmic phase and plateau phase, and all cells maintained their
normal growth state. For UC-ECFCs, the incubation period was on
days 1–2, which was followed by a logarithmic phase on days 3–7.
After the logarithmic phase, the cells entered a plateau phase. The
cell doubling time in the logarithmic growth phase was 43.53±5.38
h. This indicated that, in vitro, ECFCs demonstrated
relatively stable and rapid proliferation.
Flow cytometric analysis demonstrated that 75.58% of
cells were in G0/G1 phase, while 24.42% were in the S and G2/M
phases (Fig. 2B). The results of the
cell cycle analysis were consistent with the stemness potency of
the UC-ECFCs (3).
The colony-forming ability of these cells was also
measured. In the colony-forming assay, cell colonies appeared as
well-circumscribed monolayers under the microscope (Fig. 2C). The colonies were counted in
triplicate, and there were ~36 colonies per 103 single
cells seeded in a cell culture dish.
mRNA expression of EPC-specific genes
in human UC-ECFCs
By RT-PCR, the mRNA expression of Flt-1,
KDR, VE-cadherin, CD31, CD34,
Tie-2, vWF and ec-NOS was assessed (Fig. 3). All of these EPC-specific genes
were expressed in the UC-ECFCs.
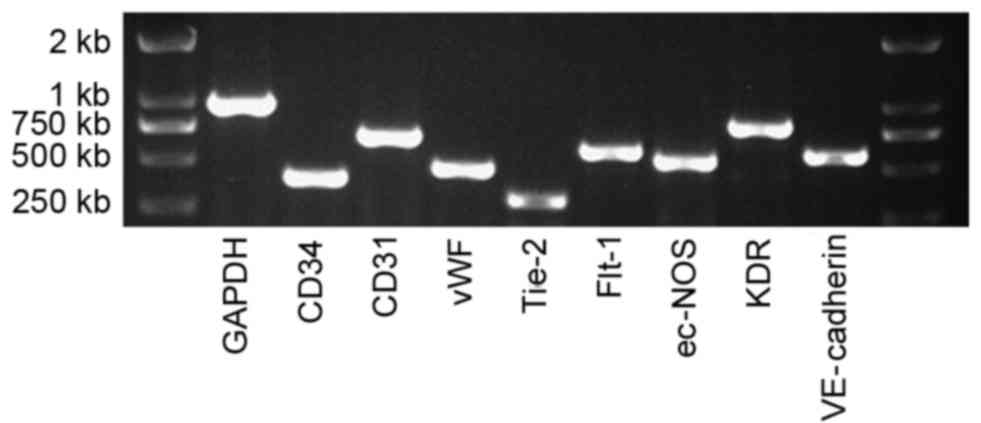 | Figure 3.mRNA expression of human UC-ECFCs at
passage three. The endothelial cell genes, CD34, CD31, vWF,
Tie-2, Flt-1, ec-NOS, KDR and VE-cadherin were all
expressed by these cells, suggesting that these cells were EPCs.
GAPDH was used as a positive control. UC-ECFCs, umbilical
cord-derived endothelial colony-forming cells; EPC, endothelial
progenitor cells; CD, cluster of differentiation; vWF, von
Willebrand factor; Tie-2, epidermal growth factor homology
domains-2; Flt-1, Fms related tyrosine kinase 1; ec-NOS,
endothelial-nitric oxide synthase; KDR, kinase insert domain
receptor; VE, vascular endothelial. |
Cell-surface marker expression on
UC-ECFCs
Flow cytometric analysis demonstrated that adherent
cells in the human UC-ECFC cultures were positive for CD31
(99.73%), CD34 (20.62%), CD73 (99.53%), CD105 (99.71%) and VEGFR2
(79.93%). The adherent UC-ECFCs were negative for the hematopoietic
marker CD45 (0.15%), mesenchymal marker CD90 (0.43%) and HLA-DR (a
major histocompatibility complex-II molecule; 0.11%). These results
indicated that the cultures were not contaminated with mesenchymal
cells (Fig. 4A). These cells weakly
expressed the hematopoietic lineage marker CD34, which decreased
with passaging (data not published).
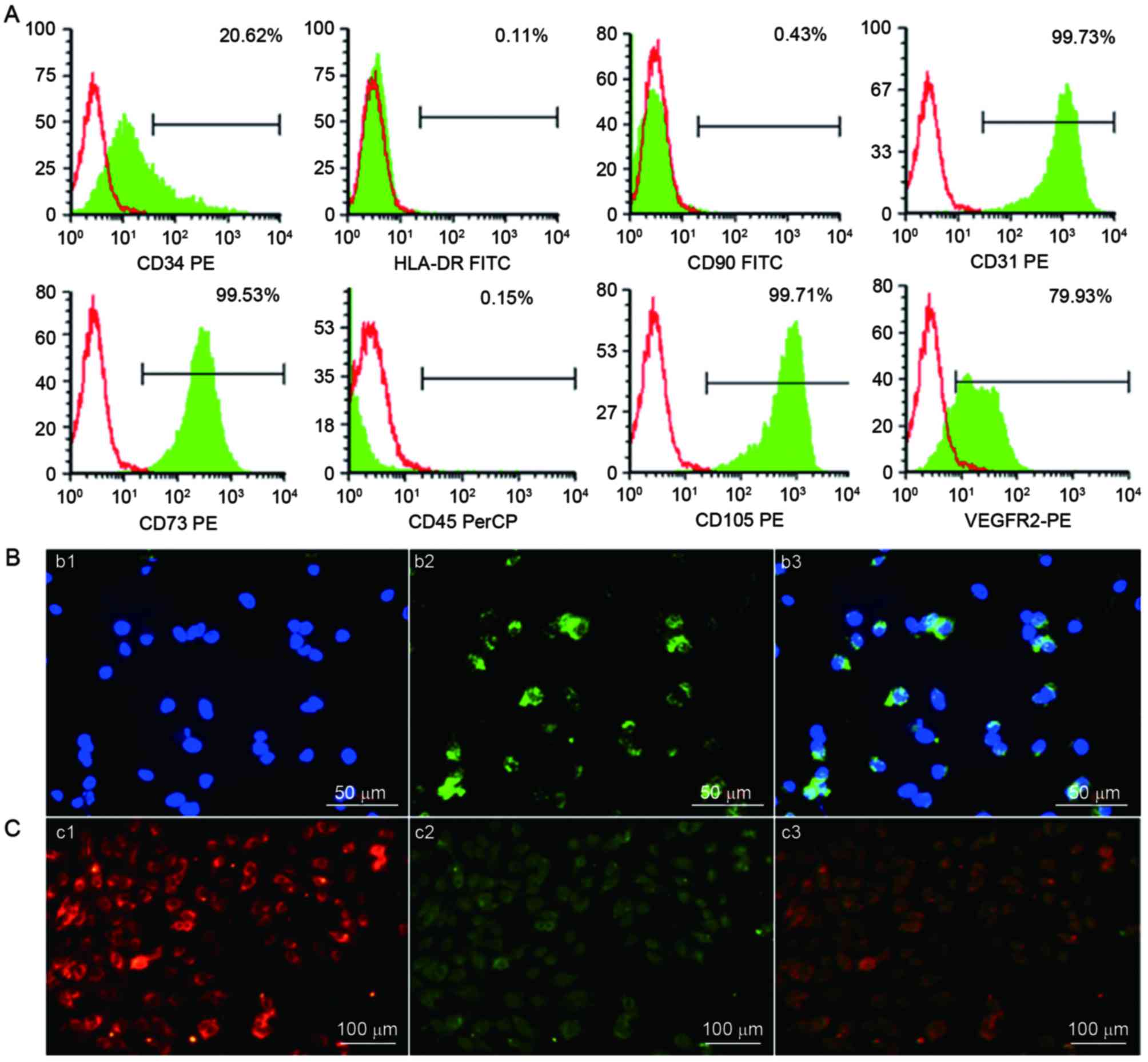 | Figure 4.Identification of human UC-ECFCs. (A)
Immunophenotype of human UC-ECFCs by FACSCalibur flow cytometry.
Human UC-ECFCs were positive for CD31 (99.73%), CD34 (20.62%), CD73
(99.53%), VEGFR-2 (79.93%), CD105 (99.71%) and negative for CD45
(0.15%), CD90 (0.43%) and HLA-DR (0.11%). Filled white histograms
represent antigen staining with negative isotype, and the green
part represents positive isotype. (B) Human UC-ECFCs expressed the
endothelial cell-surface marker vWF. The immunofluorescence
detection results demonstrated that the cells could be strained by
(b1) DAPI (blue) and (b2) mouse anti-human vWF antibody (green).
(b3) Double stained cells (merged b1 and b2) Scale bar, 50 µm. (C)
Following 24 h of culture, the adherent cells were identified by
the uptake of (c1)
1,1′-dioctadecyl-3,3,3′,3′-tetra-methyl-indocarbocyanine
perchlorate-labeled acetylated low-density lipoprotein (red
fluorescence) and binding of (c2) FITC-Ulex europaeus
lectin-l (green fluorescence). (c3) Double-positive stained cells
showed in yellow. Scale bar, 100 µm. UC-ECFCs, umbilical
cord-derived endothelial colony-forming cells; CD, cluster of
differentiation; HLA-DR, human leukocyte antigen-antigen D related
protein; VEGFR, vascular endothelial growth factor receptor; PE,
phycoerythrin; FITC, fluorescein isothiocyanate; PerCP,
peridin-chlorophyll-protein. |
The results of indirect immunofluorescent
stainingwas performed to examine the expression of endothelial
colony (EC) markers. The results demonstrated that the isolated
ECFCs expressed vWF in a punctate pattern within the cytoplasm,
which is characteristic of ECs (23,24)
(Fig. 4B).
UC-ECFC DiL-ac-LDL uptake and
FITC-UEA-l binding
Cellular immunostaining demonstrated that all
adherent cells were capable of taking up DiL-ac-LDL and binding to
FITC-UEA-l (Fig. 4C). This was
characteristic of EPCs (23,24).
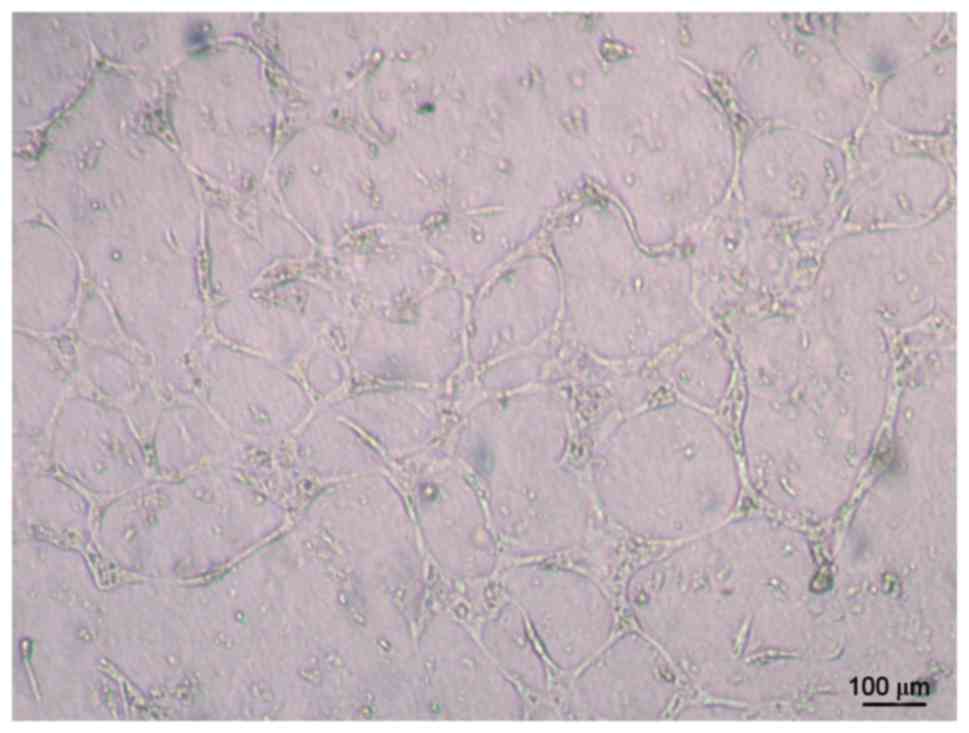
Tube formation capability of
UC-ECFCs
The results demonstrated that human UC-ECFCs had
true endothelial cell potential. Cells seeded on Matrigel and
incubated for 24 h assembled into vascular tube-like structures
(Fig. 5).
Discussion
A series of studies have indicated that ECFCs, as
EPCs, demonstrated a proliferative hierarchy of clones and the
capability to form blood vessels in vivo (25). In the present study, human UCs were
selected as a source of ECFCs as newborn UCs may be easily
obtained. To isolate human UC-ECFCs, a single enzyme approach was
applied, as a previously reported approach using magnetic activated
cell sorting was expensive and complicated, and the cells obtained
using this method demonstrated low functional activity (24,26). In
the present method, collagenase type II was injected into the
umbilical veins to liberate the cells. The isolation procedure was
simple and economical, and sufficient amounts of ECFCs were
successfully obtained through sequential flushing, digesting,
flushing and collecting. Furthermore, compared with mechanical
dissection methods, this single enzyme approach may better prevent
cell damage and contamination.
The results of the present study demonstrated that
all human UC-ECFCs obtained from the 10 UCs were stable. Compared
to the limited expansion ability of EPCs isolated from other
sources (24–26), UC-ECFCs exhibited a markedly higher
growth rate when cultured in vitro. The cell doubling time
was calculated as 43.53±5.38 h; the majority of cells were
quiescent (in G0/G1 phase); and a few cells (24.42%) were in the M
and S phase. The results of the cell cycle analysis were consistent
with the reported stemness potencies of EPCs (23,24).
The results of the present study suggested that
human UC-ECFCs may be readily expanded for clinical-scale
production in a short culture period. The results of gene
expression analysis demonstrated that ECFCs expressed markers
identical to those in EPCs from bone marrow and cord blood, such as
Flt-1, KDR, VE-cadherin, CD31,
CD34, Tie-2, vWF and ec-NOS (23). In addition, the flow cytometry
results demonstrated the presence of CD73, CD31, CD105 and VEGFR-2,
and the absence of CD45, CD90 and HLA-DR, consistent with
previously reported data (27). The
present flow cytometry assay results met the aim of the present
experiments.
While complex antigen phenotypes may be more
specific, they are difficult to reproduce, and the complexity of
the antigenic combination detected does not necessarily improve the
performance of EPCs as clinical biomarkers. Thus, research should
be aimed at making their isolation, identification and
quantification more simple and reproducible. Furthermore, tube
formation, uptake of DiL-ac-LDL and binding of FITC-UEA-l by the
isolated human UC-ECFCs was observed in the present study. It was
demonstrated that cultured ECFCs had all the characteristics of
EPCs, and the capacity of human UC-ECFCs to form tubes was greater
than that of EPCs isolated from cord blood (data not published).
The above findings suggested that the cells obtained in the present
study were true EPCs. In addition, one generation of cells from one
10-cm cord may be passaged for 7–15 days, and it was possible to
obtain ~1×107 cells by culturing for another 2–3 days. A
clinical trial demonstrated that the amount of EPCs required for
one autologous EPC transplantation is 2.5 million cells per kg
(19,27). Thus, using the UC-ECFC isolation
method from the present study, it may be possible to obtain
sufficient EPCs for clinical applications in a short time. However,
further studies are required to confirm the characteristics of
UC-derived ECFCs and their potential clinical applications.
In conclusion, the present study described a simple
and rapid method for isolating ECFCs from human UCs. The ability of
these cells to proliferate, take up DiL-ac-LDL, bind FITC-UEA-1,
express endothelial/progenitor cell-specific antigens and form
tubules in vitro was also evaluated. These results suggest
that human UCs may be a feasible and efficient source of ECFCs for
vascular injury.
Acknowledgements
The present study was supported by the ‘863
Projects’ of Ministry of Science and Technology of China (grant no.
2011AA020114), Military Clinical High-Tech Key Program (grant no.
2010gxjs100), Clinical Feature and Application Research of Capital
(grant no. Z111107058811107), Science and Technology Development
Projects of Shandong Province (grant no. 2012YD18066), Shandong
Province Commission for Population and Family Planning (grant no.
201309) and Jining Science and Technology Bureau (grant no.
2012jnnk03).
References
|
1
|
Khakoo AY and Finkel T: Endothelial
progenitor cells. Annu Rev Med. 56:79–101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zwaginga JJ and Doevendans P: Stem
cell-derived angiogenic/vasculogenic cells: Possible therapies for
tissue repair and tissue engineering. Clin Exp Pharmacol Physiol.
30:900–908. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Asahara T, Murohara T, Sullivan A, Silver
M, van der Zee R, Li T, Witzenbichler B, Schatteman G and Isner JM:
Isolation of putative progenitor endothelial cells for
angiogenesis. Science. 275:964–967. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ingram DA, Mead LE, Moore DB, Woodard W,
Fenoglio A and Yoder MC: Vessel wall-derived endothelial cells
rapidly proliferate because they contain a complete hierarchy of
endothelial progenitor cells. Blood. 105:2783–2786. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Asahara T, Masuda H, Takahashi T, Kalka C,
Pastore C, Silver M, Kearne M, Magner M and Isner JM: Bone marrow
origin of endothelial progenitor cells responsible for postnatal
vasculogenesis in physiological and pathological
neovascularization. Circ Res. 85:221–228. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin Y, Weisdorf DJ, Solovey A and Hebbel
RP: Origins of circulating endothelial cells and endothelial
outgrowth from blood. J Clin Invest. 105:71–77. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ahrens I, Domeij H, Topcic D, Haviv I,
Merivirta RM, Agrotis A, Leitner E, Jowett JB, Bode C, Lappas M and
Peter K: Successful in vitro expansion and differentiation of cord
blood derived CD34+ cells into early endothelial
progenitor cells reveals highly differential gene expression. PLoS
One. 6:e232102011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu CH, Li ZM, DU ZM, Zhang AX, Yang DY and
Wu GF: Human umbilical cord-derived endothelial progenitor cells
promote growth cytokines-mediated neorevascularization in rat
myocardial infarction. Chin Med J (Engl). 122:548–555.
2009.PubMed/NCBI
|
|
9
|
Rapp BM, Saadatzedeh MR, Ofstein RH,
Bhavsar JR, Tempel ZS, Moreno O, Morone P, Booth DA, Traktuev DO,
Dalsing MC, et al: Resident endothelial progenitor cells from human
placenta have greater vasculogenic potential than circulating
endothelial progenitor cells from umbilical cord blood. Cell Med.
2:85–96. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Matsumura G, Miyagawa-Tomita S, Shin'oka
T, Ikada Y and Kurosawa H: First evidence that bone marrow cells
contribute to the construction of tissue-engineered vascular
autografts in vivo. Circulation. 108:1729–1734. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kaushal S, Amiel GE, Guleserian KJ,
Shapira OM, Perry T, Sutherland FW, Rabkin E, Moran AM, Schoen FJ,
Atala A, et al: Functional small diameter neovessels created using
endothelial progenitor cells expanded ex vivo. Nat Med.
7:1035–1040. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun XT, Yuan XW, Zhu HT, Deng ZM, Yu DC,
Zhou X and Ding YT: Endothelial precursor cells promote
angiogenesis in hepatocellular carcinoma. World J Gastroenterol.
18:4925–4933. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scheubel RJ, Zorn H, Silber RE, Kuss O,
Morawietz H, Holtz J and Simm A: Age-dependent depression in
circulating endothelial progenitor cells in patients undergoing
coronary artery bypass grafting. J Am Coll Cardiol. 42:2073–2080.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mandraffino G, Sardo MA, Riggio S,
D'Ascola A, Alibrandi A, Saitta C, Versace A, Castaldo M, Mormina
E, Imbalzano E, et al: Circulating progenitor cells and the
elderly: A seven-year observational study. Exp Gerontol.
47:394–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
O'Brien TA, Tiedemann K and Vowels MR: No
longer a biological waste product: Umbilical cord blood. Med J Aus.
184:407–410. 2006.
|
|
16
|
Tse W and Laughlin MJ: Umbilical cord
blood transplantation: A new alternative option. Hematology Am Soc
Hematol Educ Program. 377–383. 2005.PubMed/NCBI
|
|
17
|
Yoder MC: Endothelial progenitor cell: A
blood cell by many other names may serve similar functions. J Mol
Med (Berl). 91:285–295. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aragona CO, Imbalzano E, Mamone F, Cairo
V, Lo Gullo A, D'Ascola A, Sardo MA, Scuruchi M, Basile G, Saitta A
and Mandraffino G: Endothelial progenitor cells for diagnosis and
prognosis in cardiovascular disease. Stem Cells Int.
2016:80437922016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sidney LE, Branch MJ, Dunphy SE, Dua HS
and Hopkinson A: Concise review: Evidence for CD34 as a common
marker for diverse progenitors. Stem Cells. 32:1380–1389. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Reinisch A and Strunk D: Isolation and
animal serum free expansion of human umbilical cord derived
mesenchymal stromal cells (MSCs) and endothelial colony forming
progenitor cells (ECFCs). J Vis Exp. pii: 1525. 2009. View Article : Google Scholar
|
|
21
|
Zeisberger SM, Zoller S, Riegel M, Chen S,
Krenning G, Harmsen MC, Sachinidis A and Zisch AH: Optimization of
the culturing conditions of human umbilical cord blood-derived
endothelial colony-forming cells under xeno-free conditions
applying a transcriptomic approach. Genes Cells. 15:671–687. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shintani S, Murohara T, Ikeda H, Ueno T,
Sasaki K, Duan J and Imaizumi T: Augmentation of postnatal
neovascularization with autologous bone marrow transplantation.
Circulation. 103:897–903. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dome B, Timar J, Dobos J, Meszaros L, Raso
E, Paku S, Kenessey I, Ostoros G, Magyar M, Ladanyi A, et al:
Identification and clinical significance of circulating endothelial
progenitor cells in human non-small cell lung cancer. Cancer Res.
66:7341–7347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Resch T, Pircher A, Kähler CM, Pratschke J
and Hilbe W: Endothelial progenitor cells: Current issues on
characterization and challenging clinical applications. Stem Cell
Rev. 8:926–939. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang H, Zhang B, Tao Y, Cheng M, Hu J, Xu
M and Chen H: Isolation and characterization of mesenchymal stem
cells from whole human umbilical cord applying a single enzyme
approach. Cell Biochem Funct. 30:643–649. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yoder MC, Mead LE, Prater D, Krier TR,
Mroueh KN, Li F, Krasich R, Temm CJ, Prchal JT and Ingram DA:
Redefining endothelial progenitor cells via clonal analysis and
hematopoietic stem/progenitor cell principals. Blood.
109:1801–1809. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao YH, Yuan B, Chen J, Feng DH, Zhao B,
Qin C and Chen YF: Endothelial progenitor cells: Therapeutic
perspective for ischemic stroke. CNS Neurosci Ther. 19:67–75. 2013.
View Article : Google Scholar : PubMed/NCBI
|