Introduction
Diabetes mellitus (DM) is a major leading cause of
end-stage renal failure, characterized by kidney inflammation and
glomerular dysfunction, worldwide (1). Novel therapeutic targets for end-stage
renal disease in diabetic nephropathy (DN) are urgently required
(2). Recent studies have
demonstrated that DN is characterized by an inflammatory response,
which is associated with renal glomerular and tubular epithelial,
endothelial, and interstitial cell apoptosis (3–6), which
may be closely associated with inflammatory cell damage (7).
Nicotinamide phosphoribosyltransferase (Nampt) binds
to and activates the insulin receptor and induces an insulin-like
effect, both in vitro and in vivo (8). Notably, Nampt is a key enzyme in the
nicotinamide adenine dinucleotide (NAD)+ biosynthetic
pathway, where it acts as an adipokine and improves glucose
tolerance via an insulin mimetic action (6,8). Recent
results have indicated that nuclear factor (NF)-κB signaling has a
major role in innate immunity defense whereas sirtulin (SIRT)1
regulates the oxidative respiration and cellular survival (9). Additionally, NF-κB p65 signaling may
stimulate glycolytic energy flux during acute inflammation, whereas
SIRT1 activation inhibits NF-κB p65 signaling and enhances
oxidative metabolism and reduces the expression of inflammatory and
fibrotic factors, including tumor necrosis factor-α and
transforming growth factor-β) (10).
Previous findings have indicated that SIRT1 inhibits NF-κB p65
signaling directly by deacetylating the p65 subunit of NF-κB
complex (10). Furthermore, SIRT1
stimulates oxidative energy production via the activation of
AMP-activated protein kinase, peroxisome proliferator-activated
receptor-α and peroxisome proliferator-activated receptor gamma
coactivator 1α, and simultaneously, these factors inhibit NF-κB p65
signaling and suppress inflammation (11). However, NF-κB p65 signaling has also
been indicated to downregulate SIRT1 activity through the
expression of microRNA-34a, interferon-γ and reactive oxygen
species (12).
Exogenous extracellular Nampt has been reported to
increase the synthesis of pro-fibrotic molecules in renal cell
types (13). Although high glucose
(HG) levels have been reported to increase endogenous Nampt
expression in rat mesangial cells, murine podocytes, and tubular
cells (14), the role of endogenous
Nampt enzymatic activity in diabetic renal cells, particularly
those associated with inflammation and fibrosis, is unknown beyond
the regulation of glucose uptake.
The present study examined the molecular mechanisms
of the antagonistic signaling between NF-κB p65 and SIRT1 and
described how this crosstalk may control inflammatory process and
how endogenous Nampt upregulation is associated in signaling
crosstalk through promoting chronic inflammation in DN
diseases.
Materials and methods
Animal models and groupings
Sprague-Dawley (SD) rats (male; aged 6–8 weeks old;
weighing 200–250 g) were obtained from the Experimental Animal
Center of Guilin Medical University (Guilin, China). Rats were
housed in standard polypropylene cages and maintained under
conditions of controlled room temperature (23±1°C) and humidity
(50–60%) with a 12-h light/dark cycle. Rats received standard rat
chow ad libitum throughout this study. A total of 80 rats
were acclimatized for 1 week prior to random allocation into the
control group (n=30), which were fed a normal diet (regular chow
comprising 5% fat, 63% carbohydrate and 23% protein, with total
calorific value of 25 kJ/kg) or the experimental group (n=50) which
were fed a high-fat diet (high-fat chow comprising 32% fat, 48%
carbohydrate and 20% protein, with total calorific value of 54.3
kJ/kg) and both groups animal received distilled water ad
libitum. Once the rats reached a mean body weight of 300±20 g,
those in the experimental group were administered with 50 mg/kg
streptozocin (STZ; cat. no. S0130; Sigma-Aldrich; Merck KGaA;
Darmstadt, Germany) dissolved in 0.01 mmol/l chilled citrate buffer
(pH 4.5) (cat. no. ZLI-9065; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd., Beijing, China) via intraperitoneal
injection over a 30-min period for 2 months. Control group rats
were administered with same volume citrate buffer alone. Blood
samples were subsequently collected from tail veins and used to
determine glucose levels, as detailed below. A rat was considered a
diabetic model once the fasting blood glucose concentration reached
16.7 mmol/l (Table I). A total of 20
rats in the experimental group did not develop diabetes; the 10
diabetic rats remainedwith the same high-sugar and high-fat diet
for an additional week and were subsequently sacrificed. The liver,
pancreas, kidney, and muscle tissue were carefully dissected from
each ratfor immunohistochemical analysis.
 | Table I.Levels of blood glucose and insulin
in diabetic rats. |
Table I.
Levels of blood glucose and insulin
in diabetic rats.
| Group | n | Blood glucose
(mmol/l) | Insulin
(mU/ml) |
|---|
| Control | 10 |
5.74±2.22 |
24.91±1.23 |
| Diabetes | 35 |
17.68±2.58a |
45.691±2.17b |
For the in vivo nicotinamide mononucleotide
(NMN) experiment, the diabetes induced rats (indicated above) were
randomly divided into two groups (10 rats in each group): One group
was administered 120 mg/kg NMN (cat. no. N3501; Sigma-Aldrich, Merk
KGaA; Darmstadt, Germany) in sterile PBS (200 µl) subcutaneously in
the loose skin around the neck and shoulder area every other day
for 20 days, and another group (control group) was administered
with 200 µl sterile PBS via subcutaneous injection continuously
every other day for 20 days. Following sacrifice, the animal kidney
tissues were harvested to extract total RNA for reverse
transcription-quantitative polymerase chain reaction.
A total of 5 insulin gene-mutant male C57/LB6 mice
(10–11 weeks old; Michigan Diabetes Research and Training Center of
the University of Michigan; Ann Arbor, MI, USA) (15) were spontaneously induced diabetic
mice that served as a model for a subset of human insulin gene
mutations that have been discovered to cause the autosomal-dominant
syndrome, mutant insulin gene-induced diabetes, in younger persons
(16). Wild-type C57/LB6 mice were
used as a control group (n=5), which were purchased from the
Michigan Diabetes Research and Training Center of the University of
Michigan. Both groups of mice were maintained in an air-conditioned
colony room (temperature, 21±1°C; humidity, 50–60%) on a natural
light-dark cycle and received a normal diet with standard mice chow
and water ad libitum. Housing conditions and experiments
were approved by the Ethics Committee of Animal Experimentation of
Guilin Medical University (Guilin, China). All mice were
anesthetized using diethy ether inhalation and beheaded.
Subsequently, kidney tissue was harvested for immunofluorescence
and immune-focused detection for pathological tissue analysis. When
all blood glucose levels in mice reached >27.6 mmol/l, this
indicated the mice suffered from severe diabetes, and subsequently,
the mice were sacrificed.
Cell culture experiments
The rat glomerular mesangial HBZY-1cell line was
purchased from the Basic Medical Institute of the Chinese Academy
of Medical Sciences (Beijing, China). Cells were maintained in
Dulbecco's modified Eagle's medium (DMEM; Hyclone, Logan, UT, USA)
containing 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) at 37°C. For in vitro assays, cells
were cultured under normal glucose conditions (5.6 mmol/l) at 37°C
for 24 h, and subsequently subjected to high glucose (HG; 200
mmol/l) as oxidative stress milieu (5% CO2, 37°C) for
1–6 days, in which the medium was changed every other day.
Furthermore, cells were grouped by incubation time (24-96-h
groups), and were removed to detect the changes of oxidative stress
and apoptosis in HG milieu. To determine the intervention of NMN at
different time-points and doses, the cells were divided into four
groups after 3 days of incubation at 37°C in HG milieu, and treated
with 0, 0.5, 1 or 2 mmol/l NMN, respectively for a further 24 h.
Subsequently, the protein expression levels of Nampt, NF-κB p65,
Sirt1, FoxO1 and Bax were measured using western blot analysis.
In the same experimental conditions, the protein
expression of Nampt, Sirt1, vimentin and fibronectin of the HBZY-1
cells were also detected by immunofluorescence techniques.
To further evaluate the effect of NMN, the following
the experiment was measured and the cells were grouped into 7
groups after 3 days treated in HG milieu: i) normal glucose
cultured (control group); ii) normal glucose + 10 µmol/l FK866
(Sigma-Aldrich; Merck KGaA); iii) normal glucose + 1 mmol/l NMN;
iv) HG cultured (200 mmol/l HG; HG control group); v) HG +10 µmol/l
FK866; vi) HG + 1 mmol/l NMN; and vii) HG + 10 µmol/l FK866 + 1
mmol/l NMN. FK866 and NMN were added into the cells at 24 h prior
to cell harvesting. The protein expression of Nampt, vimentin and
fibronectin were detected by western blot analysis. Once the cells
were cultured with HG for 4 days, 10 µmol/l FK866 and 1 mmol/l NMN
were added (in accordance with the previous NMN dose-effect) and
the protein expression levels of Nampt, Sirt1 and NF-κB p65 were
also analyzed using western blot analysis.
Blood glucose and insulin
detection
Collected from rat tails in regular intervalsonce
every 3 days for one week, blood glucose levels were measured using
commercially available colorimetric diagnostic kits (GT-1640; Omron
Corp., Kyoto, Japan) according to the manufacturer's instructions.
The blood samples intended for serum insulin testing were collected
as described in in the instructions of Iodine
[125I]Insulin Radio immuno assay k*-it (Beijing Research
Institute of Biotechnology of the North, Beijing, China). Serum
insulin levels were measured using a FJ-2008PS Gamma Immune Counter
(Beijing North Biotechnology Research Institute, Beijing,
China).
Immunohistochemistry and
immunofluorescence assay
The HBZY-1 A cells cultured on the slide were fixed
with 4% paraformaldehyde at room temperature for 20 min,
permeabilized with 0.5% Triton X-100 for 15 min, and blocked with
2% bovine serum albumin (Gibco; Thermo Fisher Scientific, Inc.) for
1 h at room temperature. Nampt, vimentin, fibronectin, nuclear
factor (NF)-κB p65 and sirtuin 1 (Sirt1) proteins were detected via
incubation for 1 h at room temperature with antibodies of Nampt
(1:100 dilution; sc-166866), vimentin (1:100 dilution; sc-32322),
fibronectin (1:100 dilution; sc-52331; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), NF-κB p65 (1:500 dilution; ab16502) and
Sirt1 (1:1,000 dilution; ab7343) (Abcam, Cambridge, UK) were usedat
4°C overnight. Subsequently, blots were incubated at room
temperature for 1 h and in the dark with Alexa Fluor 488-conjugated
(1:500 dilution; A32723) or Alexa Fluor 594-conjugated (1:1,000
dilution; A-11007) second antibody (Molecular Probes; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and diluted in 2% normal goat
serum, 1% bovine serum albumin and 0.1% Triton X-100 in PBS. The
cells nuclei stained with 5-µmol/l DAPI nuclear stain (Thermo
Fisher Scientific, Inc.) for 1 h at room temperature. Stained
slides were mounted and imaged using a Fluoview 500 confocal
microscope (Olympus Corporation, Tokyo, Japan).
Immunohistochemical analysis was performed on rat
tissues that were fixed in 4% paraformaldehyde (Sigma-Aldrich;
Merck KGaA) at room temperature for 5 min, dehydrated in ascending
grades of alcohols, cleared in xylene, embedded in paraffin and cut
into 4 µm-thick on the slide. Sections were dewaxed with
dimethylbenzene, rehydrated with gradient alcohol and permeablized
with 1X citric acid solutionfor 2–3 min. Sections were incubated
overnight with a primary antibody against Nampt (1:300 dilution;
sc-373717; Santa Cruz Biotechnology, Inc.) in a humidified chamber
at 4°C. A horseradish peroxidase-labeled sheep anti-rabbit
secondary at room temperature for 1 h (31460; 1:200 dilution;
Thermo Fisher Scientific, Inc.) was subsequently added to the
sections at room temperature for 20 min. For coloration, slides
were incubated with a solution of 0.05% 3,3′-diaminobenzidine at
37°C for 10 minand 0.01% H2O2, using a DAB
chromogenic reagent kit (3PW017; Sangon Biotech Co., Ltd.,
Shanghai, China). Subsequently, brown staining, which indicated
immune response strength of Nampt in the tissue sections, was
analyzed using an Image-Pro Plus Version 6.0 color image analysis
system (Media Cybernetics, Inc., Rockville, MD, USA). The degree of
positive staining was determined from the mean value of 10
different fields per slide and proportions of positive cells were
scored as follows: 0, no positive cells; 1, <10%; 2, 11–50%; 3,
51–75%; and 4, >75%. The degree of positivity was scored as
follows: 0, no staining (i.e., consistent with the background
color); 1, light yellow (slightly higher than the background); 2,
tan (significantly higher than the background); and 3, brown. The
values were multiplied to yield total scores, which were classified
as follows: 0–2, negative (−); 3–4, weakly positive (+); 5–8,
moderately positive (++); and 9–12 strongly positive (+++) as
described previously (17).
For immunofluorescence, mice kidney tissues were cut
into 3-µm thick paraffin-embedded sections on glass slides were
prepared, dewaxed, and rehydrated. The sections were permeabilized
in 0.1% Triton X-100 at room temperature for 20 min. Subsequently,
the slides were blocked in 5% donkey serum 37°C for 10 min (3AS10
1563; Agrisera, Los Angeles, USA) in PBS and then incubated at 4°C
overnight with anti-mouse primary antibodies Nampt (1:50; BS6044),
vimentin (1:100; BS1491) and fibronectin (1:100; BS4646; all
Bioworld Technology, Inc., St. Louis Park, MN, USA). Samples were
then incubated with Alexa Fluor 594-conjugated (1:1,000; R37121)
and Alexa Fluor488-conjugated (1:250; A-21121; both Thermo Fisher
Scientific, Inc.) secondary antibodies at room temperature for 1 h
and maintained in a dark place. Cell nuclei were stained with
5-µmol/l 4′,6-diamidino-2-phenylindole (D1306; Thermo Fisher
Scientific, Inc.) at room temperature for 5 min. Stained slides
were mounted and imaged (magnification, ×64) using a Fluoview 500
confocal microscope (Olympus Corporation). Immunofluorescence
signals were quantified using Image J version 2 software (National
Institutes of Health, Bethesda, MD, USA), as described by Yu et
al (17), and the integrated
densities of immune fluorescence positive signals were
measured.
NAD/NADH assay
For NAD/NADH assays of rat tissues, 30 mg frozen
muscle, pancreas, liver, kidney or an erythrocyte cell pellet (30
µl packed erythrocytes/ml Earle's balanced salt solution (Thermo
Fisher Scientific, Inc.) subjected to 4-h incubation at 37°C in 10
µmol/l N1-methylnicotinamide (AG-D-34551; Angene International
Ltd., Nanjing, China) and subsequent centrifugation at 1,500 × g
and 4°C for 10 min) were each homogenized in 400 µl NAD/NADH
Extraction Buffer (BioVision, Inc., Milpitas, CA, USA), followed
lysates incubated at room temperature for 5 min and subjected to
ultrafiltration through 10-kD cut-off filters (BioVision, Inc.).
The lysates were further subjected to analysis using NAD/NADH
quantification kits (K337-100; BioVision, Inc.) according to the
manufacturer's instructions.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the kidney tissues
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The sequences of the
primers used were as follows: Nampt (109 bp), forward
5′-ATAGGGGCATCTGCTCATTT-3′ and reverse, 5′-ACTGTGCTCTGCCGCTGGAA-3′;
NF-κB p65 (221 bp) forward, 5′-GACCTGGCATCTGTGGACAAC-3′ and
reverse, 5′-TCCGCAATGGAGGAGAAGTCT-3′; Sirt1 (506 bp), forward,
5′-GGATCCTTCATTTATCAGAGTTGCCACC-3′ and reverse,
5′-CTTCGAGGTTCTTCTAAACTTGGACTCTGG-3′; vimentin (50 bp) forward,
5′-ATGAAAGTGTGGCTGCCAAGAAC-3′ and reverse,
5′-GTGACTGCACCTGTCTCCGGTA-3′; and β-actin (30 bp) forward,
5′-GATGGTGGGTATGGGTCAGAAGGAC-3′ and reverse,
5′-GCTCATTGCCGATAGTGATGACCT-3′. All primers were synthesized by
Shanghai Sangon Biological Engineering Technology & Services
Co., Ltd., (Shanghai, China). For the reverse transcription
template, in a 0.5-ml tube, 1 µl Thermo Script RT, 1 µl Oligo
(dT)20, 2 µl of 50 ng/µl random primer, 4 µl dNTP mix, 1
µl 40 U RNaseOUT, 10 µl of 0.1 M DTT, 2 µl E. coli RNase and
20 µl 5X cDNase Synthesis Buffer were added and adjusted to a final
volume volume 50 µl with DEPC-treated water. RT reaction included
was performed at 56°C for 1 h before the addition of 1 µl RNase H.
The reaction was incubated at 37°C for 20 min and 2 µl aliquot was
removed for PCR. qPCR was performed with 1 µl cDNA, using an RT-PCR
kit (K1691; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The reaction conditions included
denaturation at 94°C for 2 min, renaturation at 58°C for 50 sec and
extension at 72°C for 30 sec, with 30 cycles in total. The
collected amplification products were separated via 2% agarose gel
electrophoresis and visualized by ethidium bromide staining. Band
intensity analysis was performed with Gene Tools software (UVP,
Inc., Upland, CA, USA). All RT-PCR procedures were performed using
a Thermo Script RT-PCR System (Thermo Fisher Scientific, Inc.).
Western blot analysis
Total proteins from HBZY-1 cells and from rat
tissues (liver, pancreas, muscle and kidney) were extracted using a
Protein Extraction Kit (Kangchen Biotechnology, Shanghai, China)
according to the manufacturer's protocol. Protein quantification
was performed using a BCA protein concentration determination kit
(23235; Leagene Biotechnology Ltd., Beijing, China) according to
the manufacturer's instructions. The protein (30 µg) was loaded and
separated using 10% SDS-PAGE, and subsequently transferred onto a
polyvinylide fluoride sheets membranes (Thermo Fisher Scientific,
Inc.). The membranes were blocked in 5% skimmed milk dissolved in
tris-buffered saline with Tween-20 (TBST) at room temperature for 1
h. The membranes were incubated with rabbit polyclonal antibodies
against Nampt (1:200 dilution; sc-166866), Poly(ADP-ribose)
polymerase-1 (PARP-1; 1:500 dilution; sc-136208), Sirt1 (1:500
dilution, sc-74465), vimentin (1:200 dilution; sc-166866),
fibronectin (1:200 dilution; sc-69681), forkhead box protein O1
(FoxO1; 1:500 dilution; sc-374427), NF-κB p65 (1:200 dilution;
ab-86299) and B-cell lymphoma 2-like protein 4 (Bax; 1:400
dilution; sc-20067; Santa Cruz Biotechnology, Inc.) and a rabbit
monoclonal antibody against β-actin (1:200 dilution; TA310155;
OriGene Technologies, Inc., Beijing, China) overnight at 4°C.
Subsequently, following incubation with horseradish
peroxidase-conjugated goat anti-rabbit (1:2,000 dilution; 170-6515;
Bio-Rad, Hercules, CA, USA) and goat anti-rat IgG H&L (1:2,000
dilution; ab97057; Abcam, Cambridge, UK) secondary antibodies at
37°C for 1 h, blots were developed using an enhanced
chemiluminescence substrate according to the manufacturer's
protocol (GE Healthcare, Chicago, IL, USA). Gray-scale (total raw
density) values of blots were measured using the VisionWorksLS
analysis software provided with the UVP EC3 (600) Imaging System
(Quantity One; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
Statistical analyses were performed using SPSS 16.0
package (SPSS, Inc., Chicago, IL, USA). A one-way analysis of
variance was used for the analyses, and data are presented as the
mean ± standard error of the mean. P<0.05 was considered to
indicate a statistically significant difference.
Results
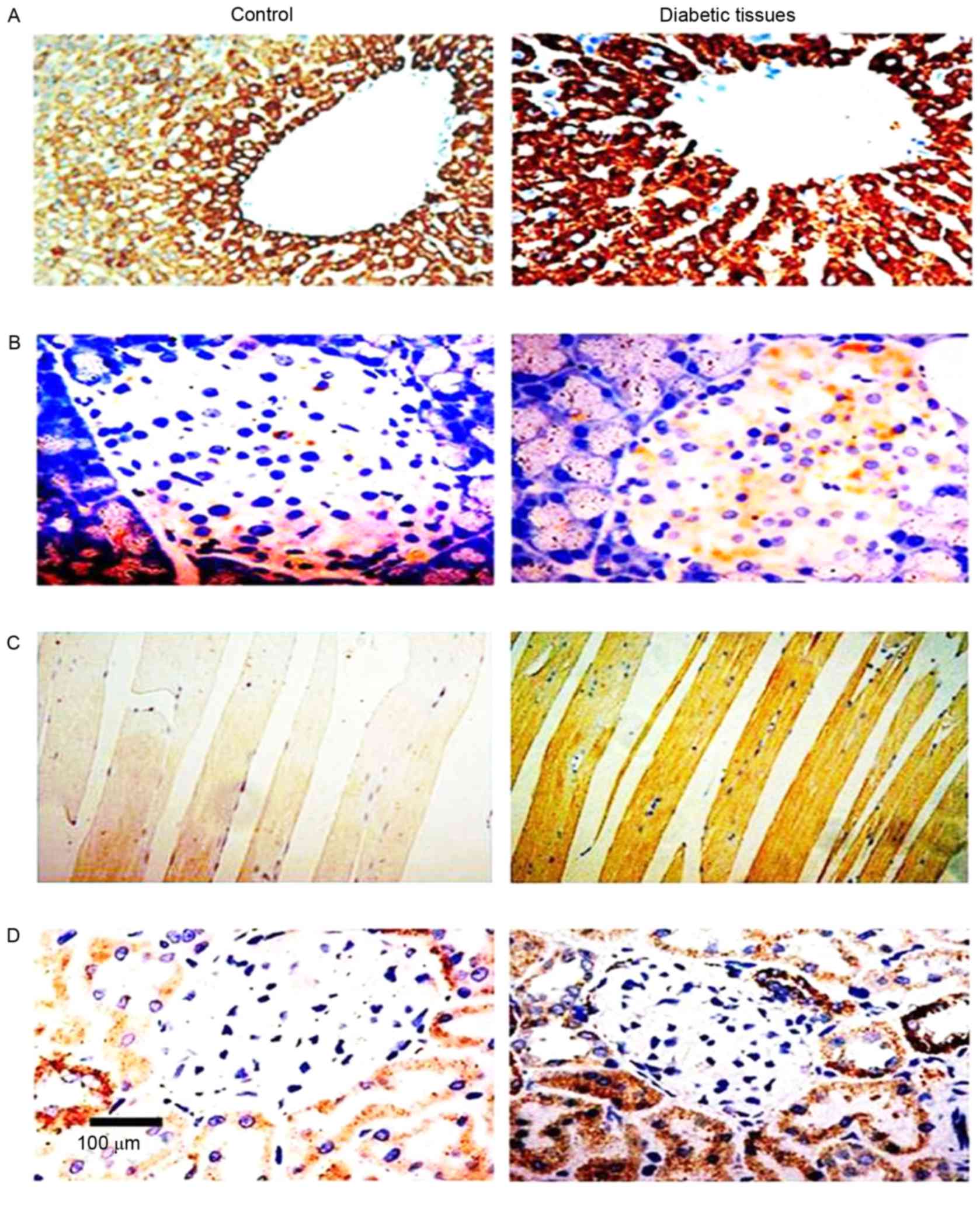
Upregulation of endogenous Nampt and
PARP1 in STZ-induced diabetic mice
Endogenous Nampt expression in rat tissues were
presented (Figs. 1 and 2; Table II)
via immunohistochemistry and western blot techniques. Expression
levels of endogenous Nampt in the liver, pancreas, muscles, and
kidney in diabetic rats were all higher than those of the control
group; respective increases of 3.22-, 2.54-, 2.73-, and 2.36-fold
were observed (Fig. 2A and B).
Furthermore, PARP-1 expression levels in the liver, pancreas,
muscles, and kidney of diabetic rats were higher than those of the
control group; respective increases of 2.47-, 2.57-, 1.60- and
3.63-fold were observed (Fig. 2C and
D). Significant increases of endogenous Nampt and PARP-1
expression were observed in tissues from diabetic rats, compared
with controls (P<0.01; Fig. 2B and
D).
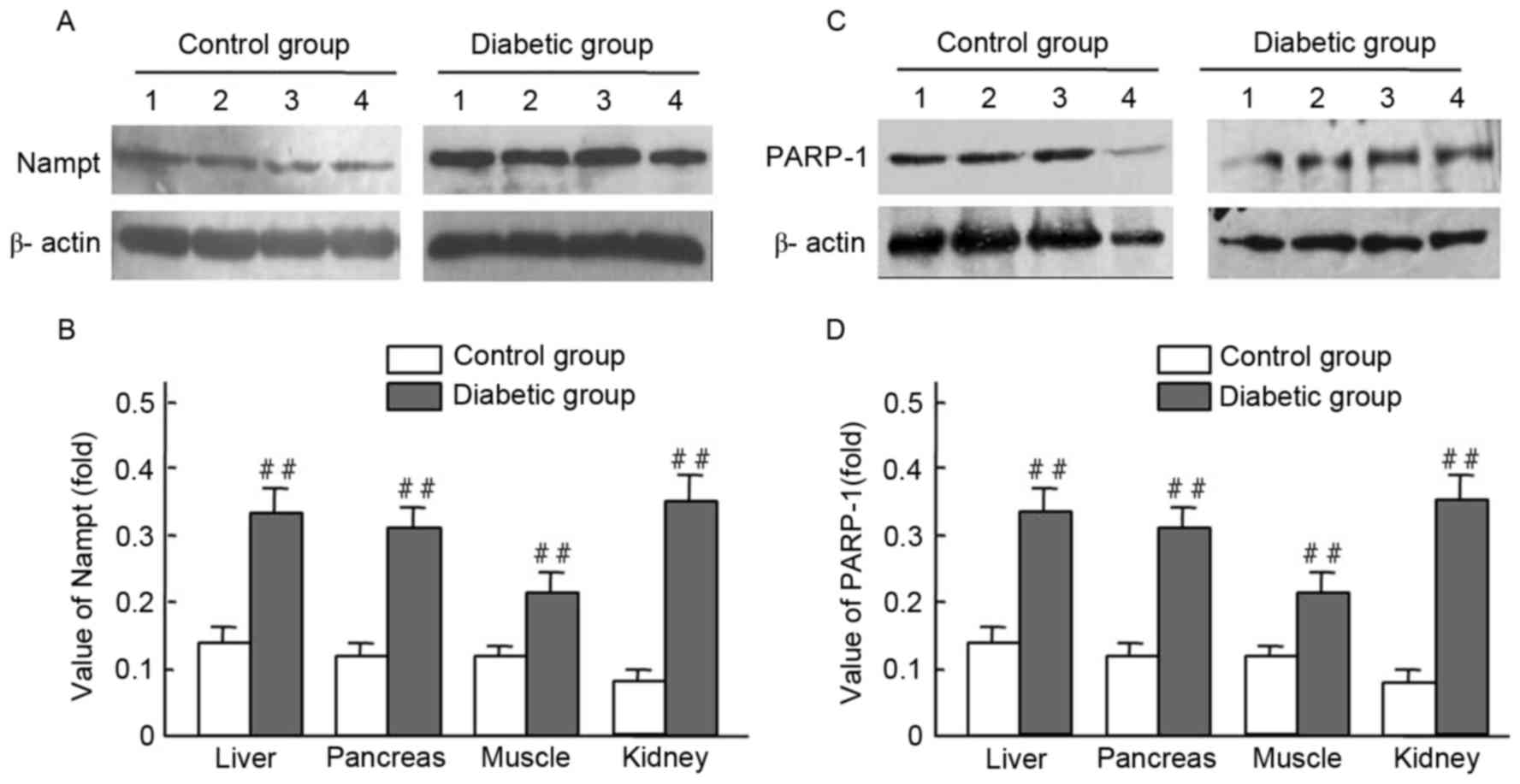 | Figure 2.Nampt and PARP-1 expression levels in
tissues from streptozocin-induced diabetic and normal rats. Rat
tissue lysates were subjected to western blot analysis with
antibodies against Nampt, PARP-1 and β-actin (control); experiments
were performed at least in duplicate. Representative images of
bands are shown. Data are presented as means ± standard deviations
for 3 pancreatic sections from 3 rats per group. (A-D) The
expression levels of Nampt and PARP-1 were obiviously increased in
the diabetic liver, pancreas, musle and kidney tissues compared
with those of normal rats, ##P<0.01 vs. control.
Nampt, nicotinamide phosphoribosyltransferase; PARP-1,
Poly(ADP-ribose) polymerase-1; band 1, liver; band 2, pancreas;
band 3, muscle; band 4, kidney. |
| Table II.Endogenous Nampt expression in
diabetic rat tissue samples. |
Table II.
Endogenous Nampt expression in
diabetic rat tissue samples.
| Group | n | Liver | Pancreas | Muscle | Kidney |
|---|
| Control | 10 | ++ | + | + | ++ |
| Diabetes | 35 | +++ | ++ | +++ | +++ |
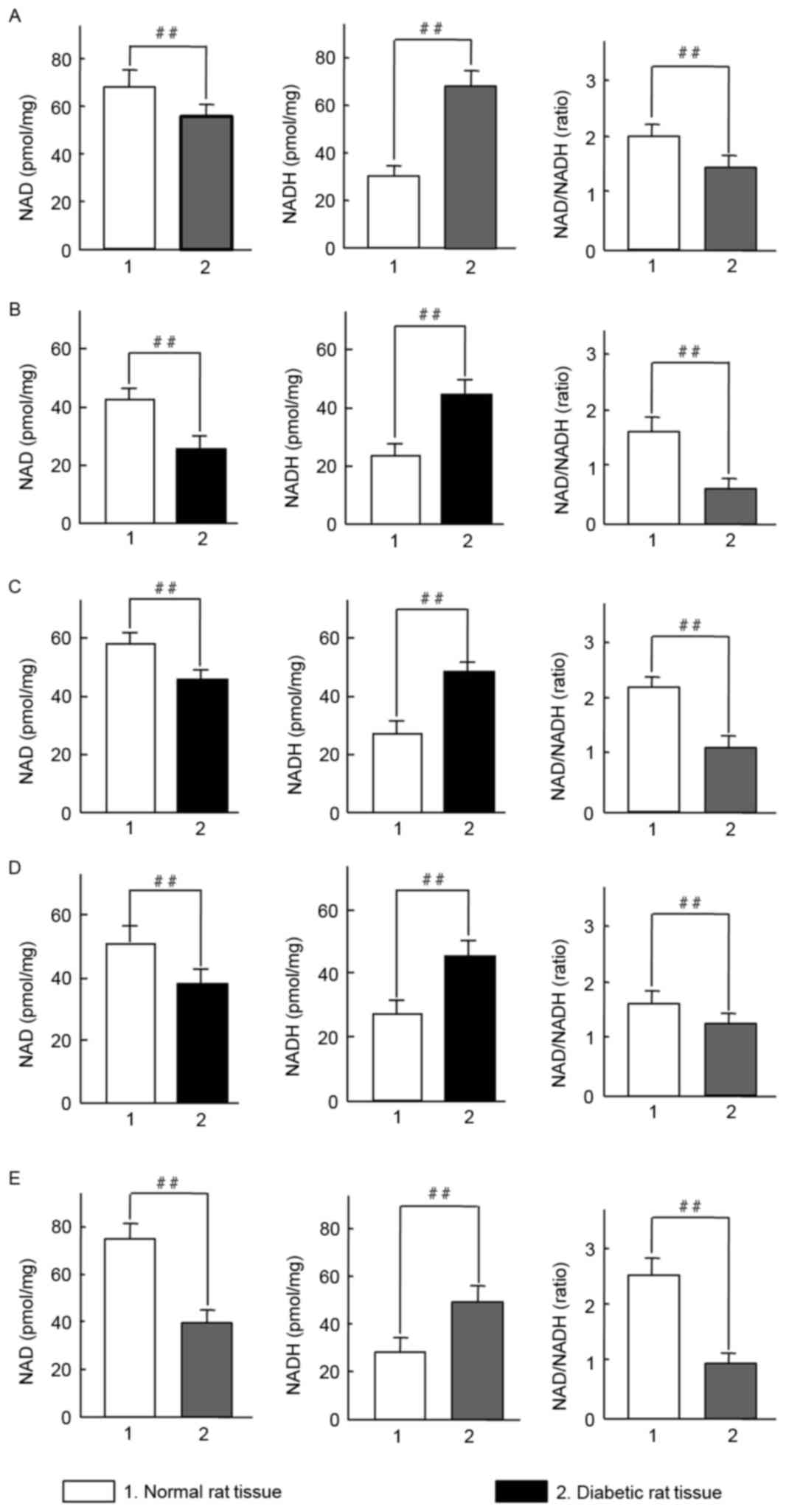
Dysregulated NAD and NADH levels in
STZ-induced diabetic rats
Diabetes is known to be associated with oxidative
stress (18). It was therefore
examined whether oxidative stress was an etiological factor in
diabetic rats. Initially, changes of NAD, NADH and the NAD/NADH
ratio in rat tissues were evaluated. Notably, a significant
increase in NADH levels were observed in the erythrocytes, muscle,
pancreas, liver and kidney of diabetic rats, whereas NAD levels
were significantly decreased compared with the control group
(P<0.01; Fig. 3). All values of
NAD/NADH ratio were significantly reduced in the tissue from
diabetic rats compared with controls (P<0.01; Fig. 3A-E). The results indicate that
STZ-induced diabetes in SD rats may induce an imbalance in the
NAD/NADH ratio under oxidative stress.
Increased Nampt and NF-κB p65
expression and decreased Sirt1 expression in response to HG
conditions over time
The expression levels of endogenous Nampt, NF-κB
p65, and Sirt1 were measured in HBZY-1 cells treated continuously
in an HG milieu for 24, 48, 72, 96, 120, and 144 h (Fig. 4). The western blot results indicated
endogenous Nampt and NF-κB p65 expression levels were increased in
a time-dependent manner (Fig. 4A),
and 72-144-h incubation resulted in a significant increase in
expression compared with 24-h incubation (P<0.05 and P<0.01;
Fig. 4B and D). Furthermore, Sirt1
expression was significantly decreased when the cells were cultured
in HG milieu for 72–120 h compared with 24-h incubation (P<0.05
and P<0.01; Fig. 4C). These
results suggest that Nampt upregulation may affect the expression
of NF-κB p65 and Sirt1 following exposure of cells to HG
milieu.
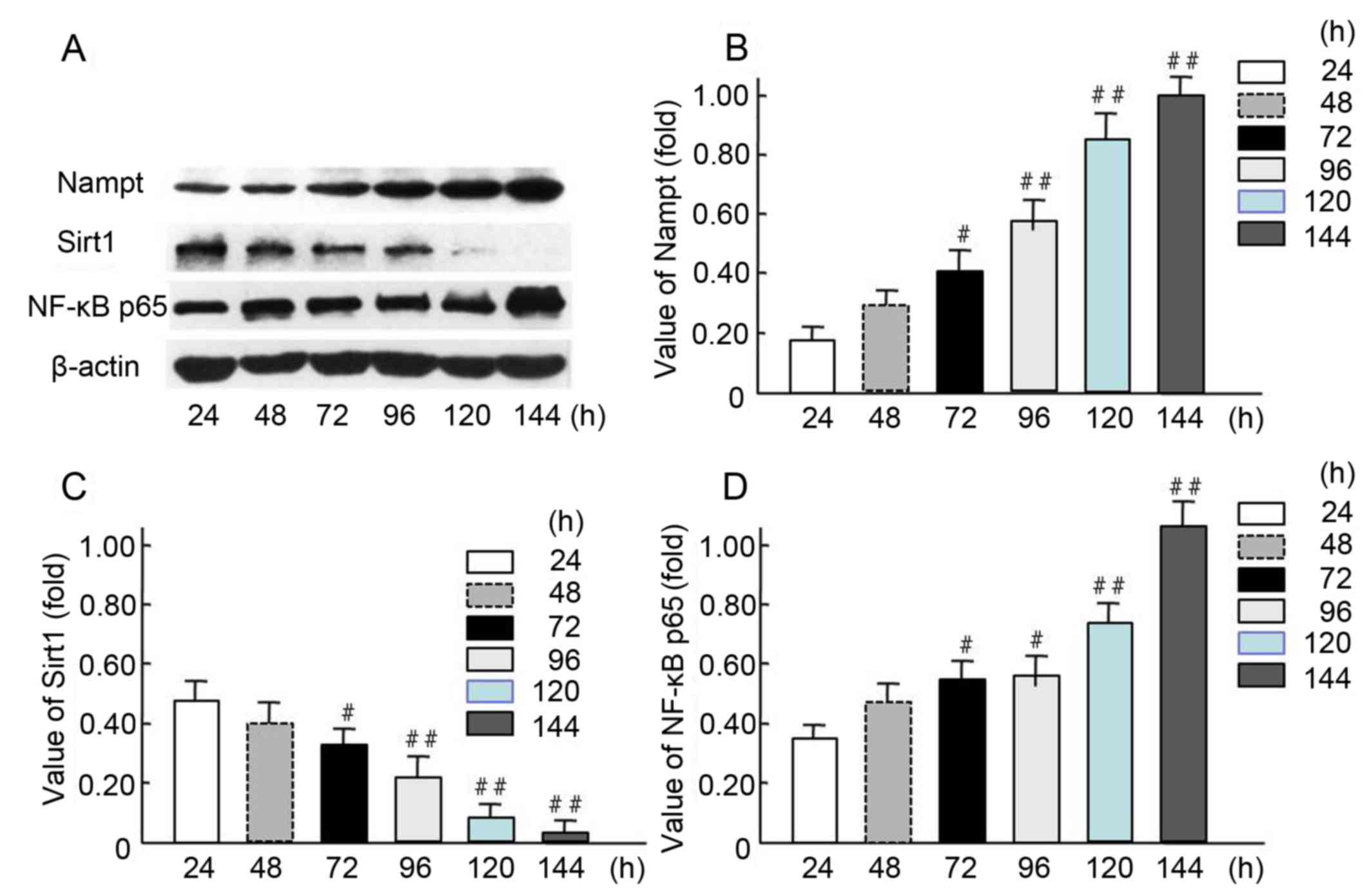 | Figure 4.Increased endogenous Nampt and NF-κB
p65 expression and decreased Sirt1 expression in oxidative
stress-impaired HBZY-1 cells. (A) HBZY-1 cells were incubated in a
high glucose media (200 mmol/l as oxidative stress milieu) for 24,
48, 72, 96, 120, or 144 h, then the expression levels of endogenous
Nampt, NF-κB p65, Sirt1, and β-actin proteins were measured via
western blot analysis. Quantification of (B) Nampt, (C) NF-κB p65
and (D) Sirt1 levels are presented. Each independent experiment was
repeated three times per group. Data are presented as means ±
standard deviations of three experiments. #P<0.05,
##P<0.01 vs. 24-h treated group. Nampt, nicotinamide
phosphoribosyltransferase; Sirt1, sirtuin 1; NF, nuclear
factor. |
To further investigate this, HBZY-1 cells were
exposed to HG for a further 120 h prior to immunofluorescence
analysis. Nampt upregulation was indicated under fluorescence
microscopy as increased green flurorescence was observed in the HG
milieu group compared with the control. Further staining revealed a
notable increase in NF-κB p65 expression in HG cells and
translocation from the cytosolinto the nuclei, compared with
controls (Fig. 5A). Quantification
of the images revealed that the increase in NF-κB p65 was
statistically significant (P<0.01; Fig. 5B). Similarly, Nampt expression was
significantly increased (P<0.01; Fig.
5B). Conversely, in HG cells, Sirt1 expression was decreased
compared with the 24-h control group; however, Nampt expression was
increased (Fig. 5C). Quantification
of these results indicated these differences were statistically
significant (P<0.01; Fig.
5D).
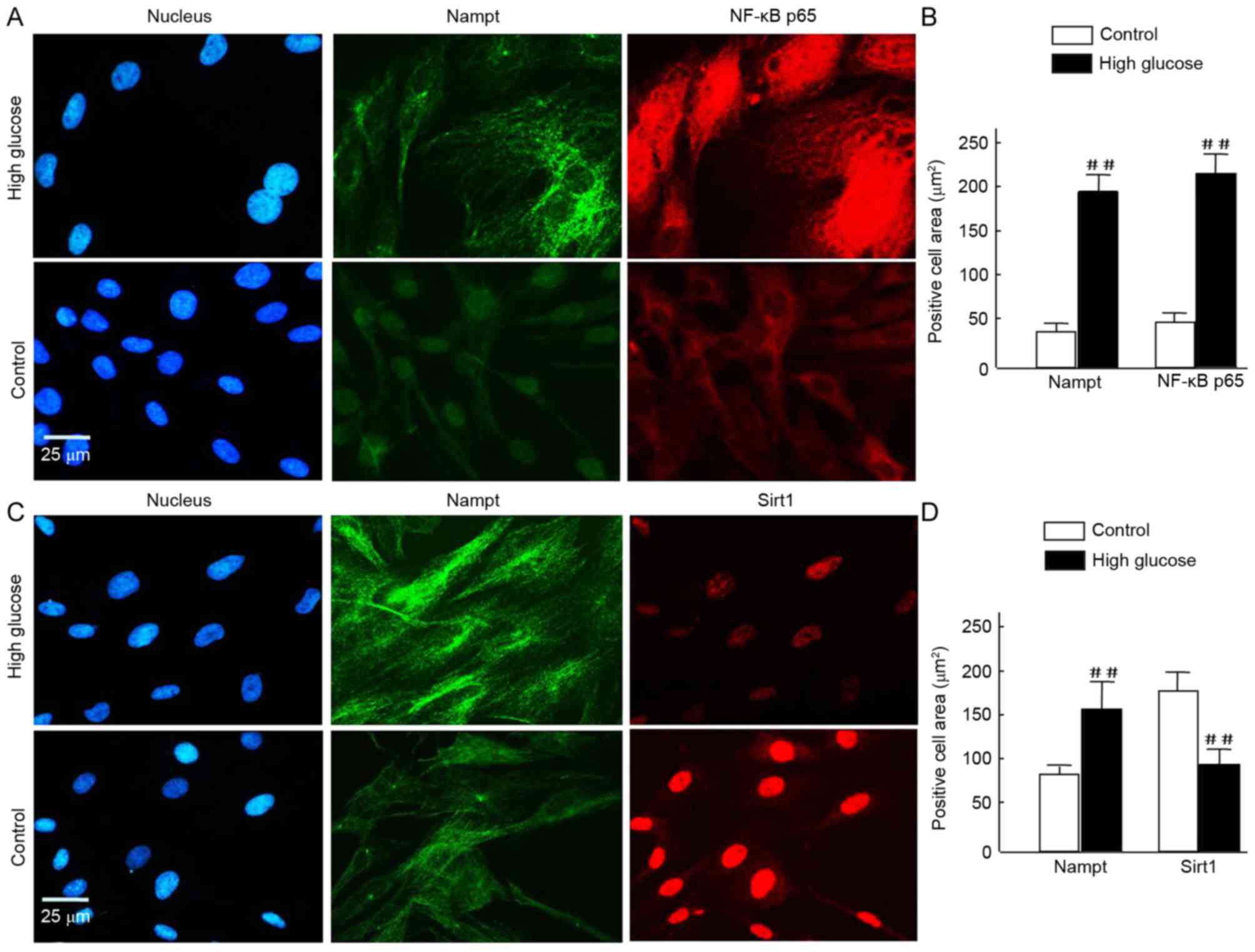 | Figure 5.(A-D) To assess cell morphology using
fluorescence microscopy, HBZY-1 cells were exposed tohigh glucose
for 120 h and subsequently fixed and stained with Alexa Fluor 488-
and Alexa Fluor 594-conjugated rabbit antibodies against Nampt,
NF-κB p65, and Sirt1, and DAPI (blue) for nuclear visualization.
Similar results were obtained in 3 independent experiments. (A and
C) In the images, Nampt staining is indicated in green, and NF-κB
p65 or Sirt1 staining is indicated in red. Merged images reveal
that the majority of Nampt staining co-localizes with both NF-κB
p65 and Sirt1. (B and D) Positive signals were normalized to the
cell area (integrated density/mm2) and quantitatively
analyzed using Image J software. In each nephritic section, ≥10
areas containing ~100 cells were analyzed. Data are expressed as
means ± standard deviations of four independent experiments (~50
cells per experiment). Magnification, ×40, Scale bar=25 µm.
##P<0.01 vs. control. Nampt, nicotinamide
phosphoribosyltransferase; NF, nuclear factor; Sirt1, sirtuin
1. |
Induction of cell apoptosis by
HG-mediated oxidative stress
To further confirm the upregulation of endogenous
Nampt following exposure to oxidative stress milieu, cells were
incubated in HG DMEM medium for 24–96 h. Subsequently, relative
protein expression levels of Nampt, NF-κB p65, Sirt1, FoxO1 and Bax
at different time points were measured by western blotting
(Fig. 6). A representative image of
the western blot results is indicated in Fig. 6A. Endogenous Nampt expression
significantly increased by 96 h to 3.47 times that at 24 h
(Fig. 6B). Furthermore, NF-κB p65,
FoxO1 and Bax expression subsequently increased to varying degrees
following Nampt upregulation. Protein expression levels of Nampt,
FoxO1 and Bax peaked at 96 h, whereasNF-κB p65 peaked at 72 h
(Fig. 6C, D and F). As indicated in
Fig. 6E, Sirt1 expression was
significantly decreased following 96 h exposure to a HG milieu
(P<0.01).
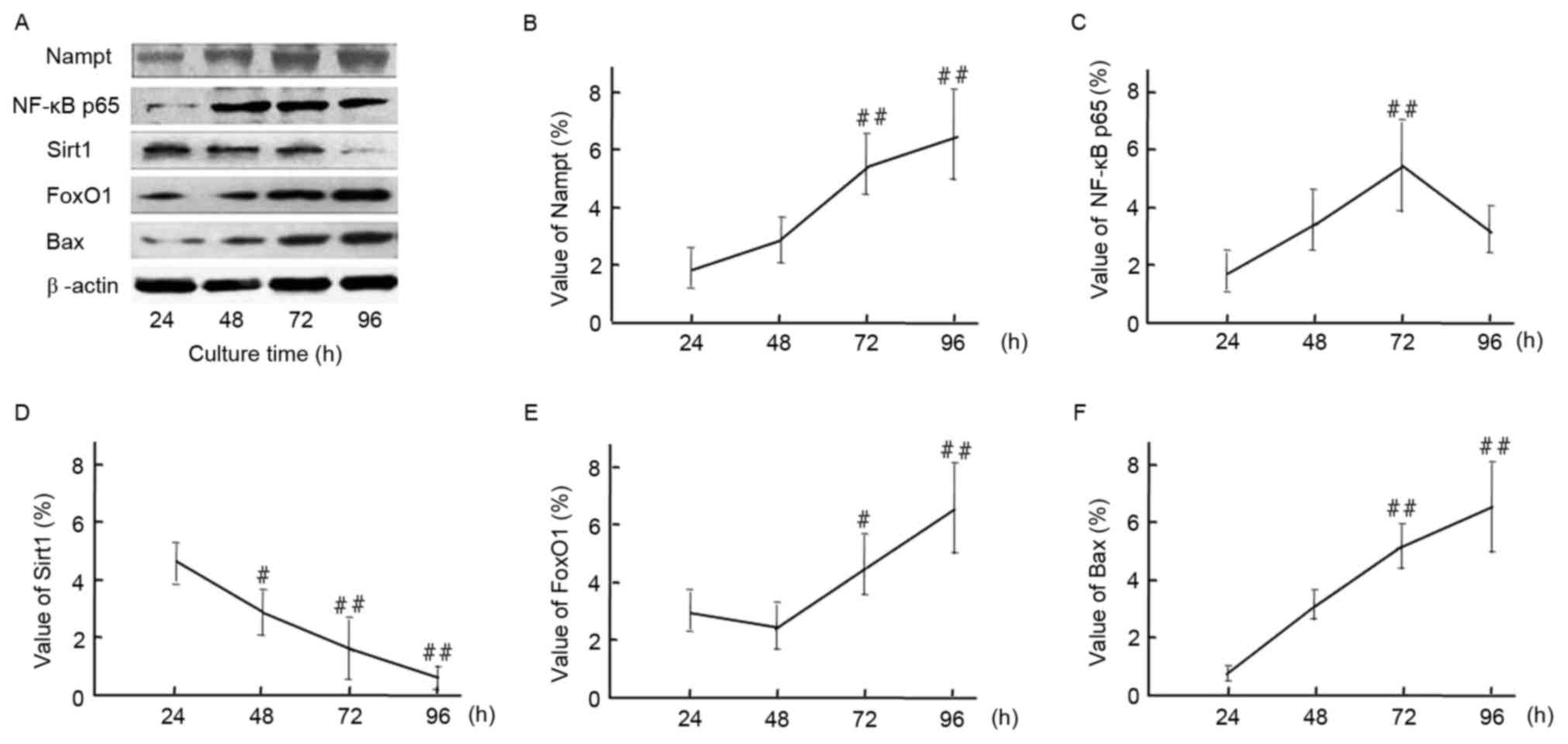 | Figure 6.(A) Time course of endogenous Nampt,
NF-κB p65, Sirt1, FoxO1, and Bax protein expression in HBZY-1 cells
exposed to a high-glucose milieu for 24, 48, 72, or 96 h.
Quantification of western blot analysis indicated the protein
expression levels of (B) Nampt, (C) NF-κB p65, (D) Sirt1, (E) FoxO1
and (F) Bax, and β-actin were indicated. Data are presented as
means ± standard deviations of three experiments.
#P<0.05, ##P<0.01 vs. 24 h (control).
Nampt, nicotinamide phosphoribosyltransferase; NF, nuclear factor;
Sirt1, sirtuin 1; FoxO1, forkhead box protein O1; Bax, B-cell
lymphoma 2-like protein 4. |
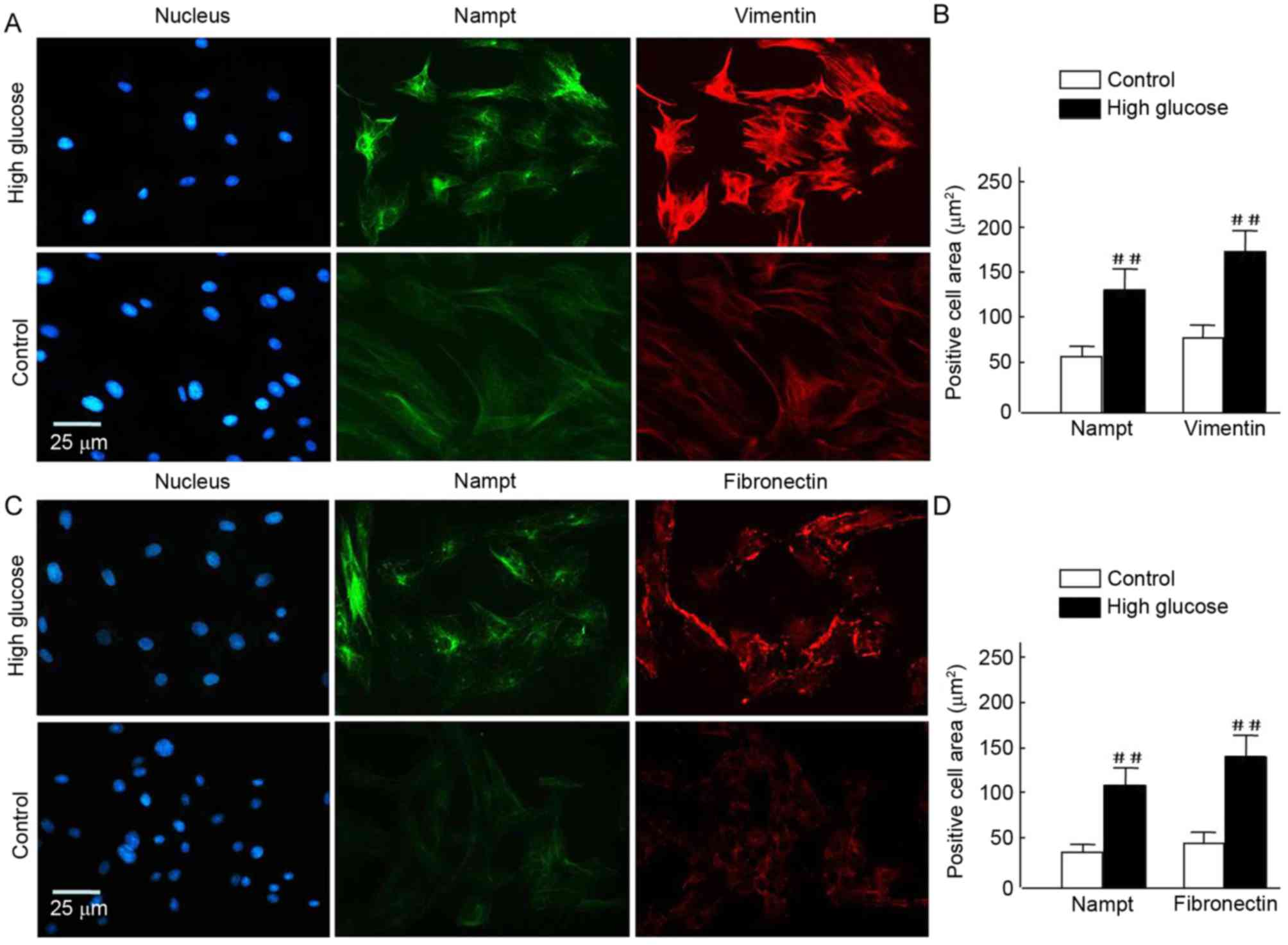
Upregulation of Nampt is closely
associated with renal fibrosis in diabetic mice
It was investigated whether Nampt upregulation was
able to induce the expression of renal cell fibrotic factors, such
as vimentin and fibronectin, in an insulin gene-mutant diabetic
C57/LB6 mouse model. With double immunofluorescence staining,
glomerularmorphology and distribution, and localization of target
proteins in kidney sections were detected viaconfocal microscopy
(Fig. 7). The findings
revealedsevere glomerular atrophy and size reductions in sections
of diabetic mice kidney. Following Nampt upregulation in renal
tubular and glomerular tissue, increased vimentin expression was
observed specifically in glomerular cells but not in renal tubular
cells (Fig. 7A and B). Notably,
Nampt and vimentin expression levels were significantly higher in
diabetic sections compared with control sections (P<0.01;
Fig. 7E). Fibronectin expression was
increased in diabetic glomerular cells compared with controls
(P<0.01; Fig. 7C, D and F).
Similar results were also observed in HBZY-1 cells when subjected
to HG-mediated oxidative stress (P<0.01; Fig. 8). These results suggested that there
may be an association between Nampt and fibrotic factors, such as
vimentin and fibronectin.
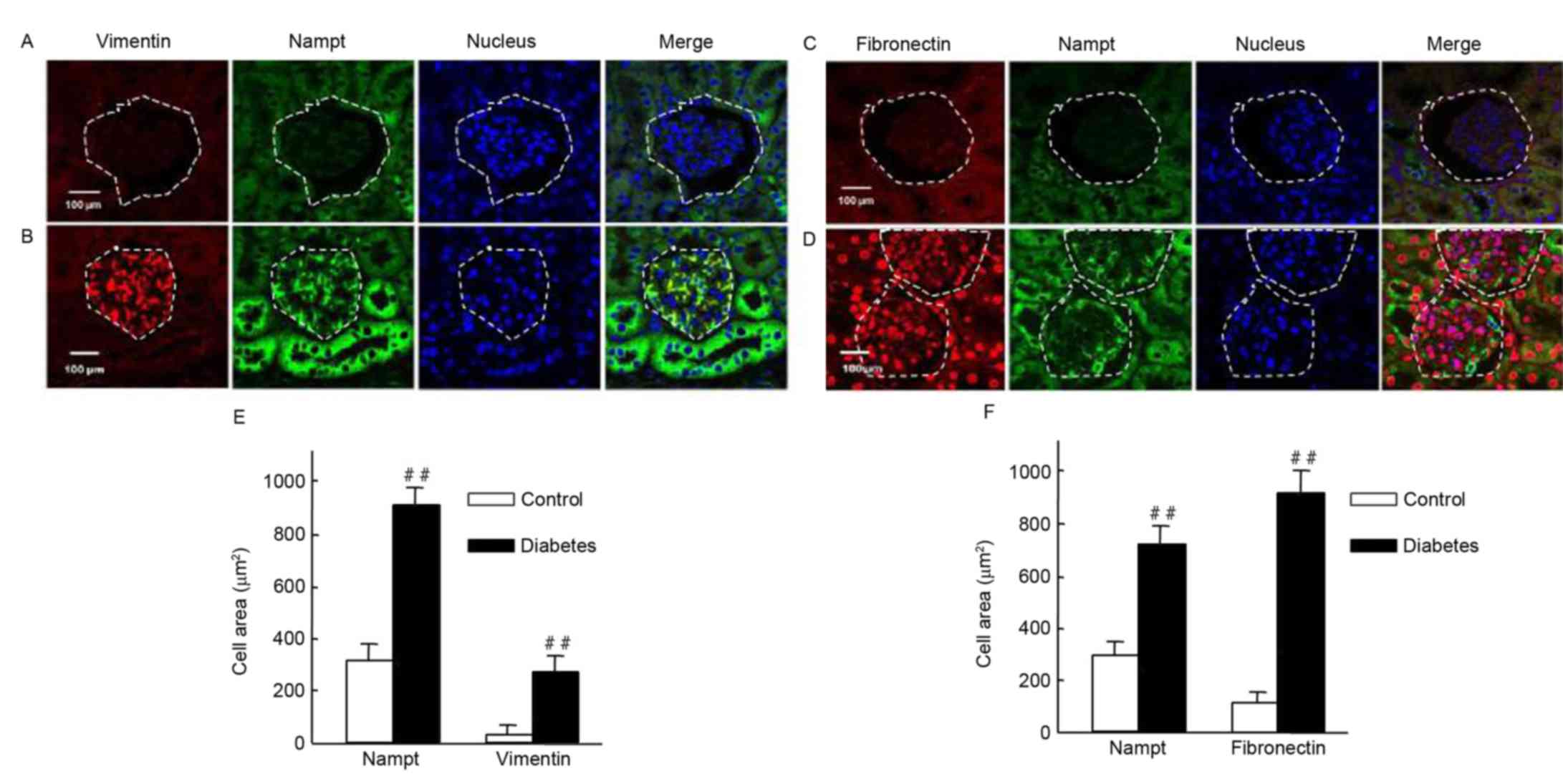 | Figure 7.Endogenous Nampt, vimentin, and
fibronectin expression in nephritic tissues of diabetic mice. The
inner circle of the broken line indicated the glomerulus cells,
which were surrounded by renal tubular cells. (A-D) Representative
images of double immunestaining for Nampt (green) and fibronectin
or vimentin (red). Nuclei were stained with DAPI (blue).
Magnification, ×64, Scale bar=100 µm. (E) Endogenous Nampt and
vimentin expression levels were significantly higher in nephritic
tissue cells from diabetic micethan in cells from control mice. (F)
Similarly, fibronectin levels were significantly higher in tissues
from diabetic mice than from control mice. Images were analyzed
using Image J software. Data are presented as means ± standard
deviations for 3 mice per group and 3 nephritic sections per mouse.
In each nephritic section, ≥10 areas containing ~100 cells were
analyzed. ##P<0.01 vs. controls. Magnification, ×64,
Scale bar=100 µm. Nampt, nicotinamide
phosphoribosyltransferase. |
NMN regulates kidney cell fibrosis by
Nampt-NF-κB p65 and Sirt1 signaling pathway in vivo and in
vitro
The mRNA expression of Nampt, NF-κB p65 and vimentin
were significantly decreased in the NMN treated group compared with
the untreated group (P<0.01), whereas expression of Sirt1 was
significantly increased in the same group (P<0.01). These
results suggest that the Nampt-NF-κB p65 signal pathway was
specifically inhibited by NMN via a negative feedback loop
(Fig. 9). To determine whether the
Nampt-NF-κB p65 and Sirt1 signaling pathway has a role in vimentin
expression, the expression levels of Sirt1 were measured in time-
and dose-mediated manners in HG-cultured HBZY-1 cells by
intervention of exogenous NMN (Fig.
10). The results indicated that NMN was able to activate Sirt1
by inhibiting the Nampt pathway. Following NMN treatment, the
expression of Sirt1 was significantly increased compared with the
NMN-untreated group. (P<0.01; Fig.
10A), which may induce the downregulation of vimentin
expression via interacting withNF-κB p65. Furthermore, results
indicated that Sirt1 expression was significantly increased in all
NMN-treated groups compared with NMN-untreated group (P<0.01;
Fig. 10B). The result suggested
that exogenous NMN may protect cells from fibrosis by inhibiting
Nampt upregulation and promoting Sirt1 expression, but restraining
NF-κB p65.
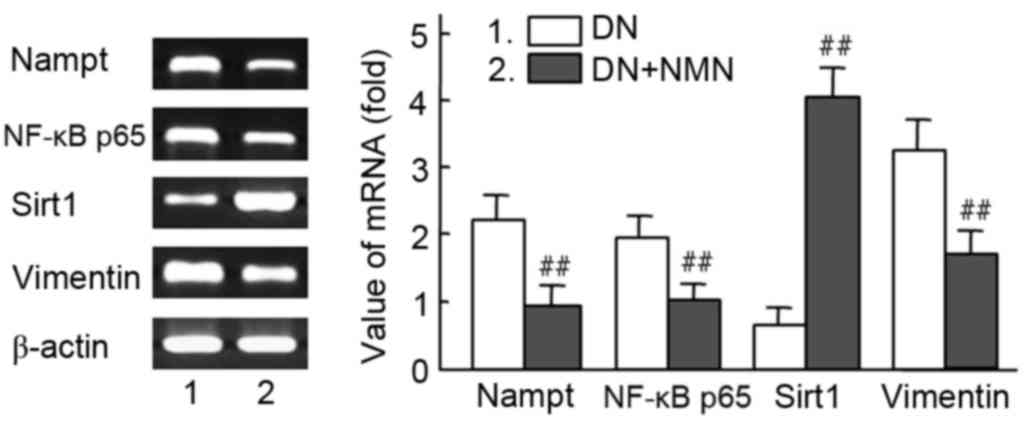 | Figure 9.Effect of NMN on RNA expression of
Nampt, NF-κB p65 and vimentin in diabetic kidney tissues. The
Nampt-NF-κB p65 and Sirt1 signaling pathway were regulated with
NMN, which participated in intervened vimentin expression in
vivo. Diabetic rats were treated with NMN for 20 days. The mRNA
expression analysis of Nampt, NF-κB, Sirt1 and vimentin in diabetic
rats is presented. β-actin was used as a loading control. Relative
density value of PCR products=target gene grey value/β-actin grey
value. Experiments were performed in triplicate, and means ±
standard deviations of 3 experiments are presented.
##P<0.01 vs. DN. Lane 1, DN rat; lane 2, DN rat+NMN.
NMN, nicotinamide mononucleotide; Nampt, nicotinamide
phosphoribosyltransferase; NF, nuclear factor; Sirt1, sirtuin 1;
DN, diabetic nephropathy. |
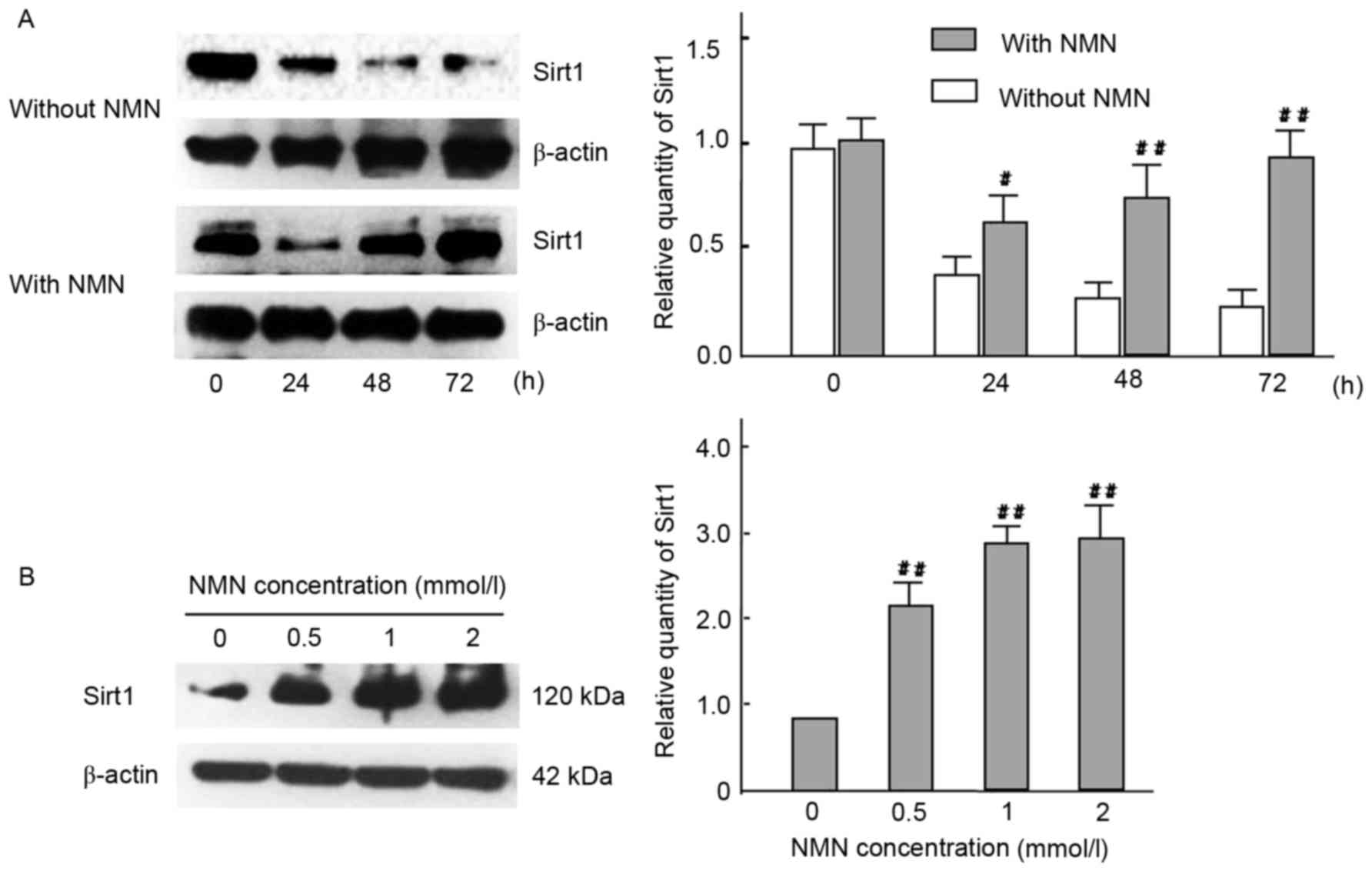 | Figure 10.Effect of time- and dose-course in
expression of Sirt1 with HG cultured HBZY-1 cells. (A) Cells in HG
medium were treated with or without 1 mmol/l NMN for 0, 24, 48 or
72 h, and Sirt1 protein expression were analyzed by western blot
analysis. β-actin was used as a loading control.
#P<0.05, ##P<0.01 vs. without NMN. (B)
Cells in HG medium were treated with 0, 0.5, 1 or 2 mmol/l NMN for
24 h, and Sirt1 protein expression were analyzed by western
blotting. β-actin was used as a loading control.
##P<0.01 vs. 0 mmol/l. The experiments were performed
three times, and means ± standard deviations of 3 experiments are
presented. Sirt1, sirtuin 1; HG, high glucose; NMN, nicotinamide
mononucleotide. |
Vimentin and Fibronectin expression
levels were downregulated in response to FK866 and NMN
treatment
Western blot analysis was used to investigate the
mechanisms underlying Vimentin and Fibronectin over expression,
following the hypothesis that endogenous Nampt may affect NF-κBp65
pathway activation (Fig. 11). A
representative western blot is presented in Fig. 11A. Notably, Vimentin and Fibronectin
were significantly decreased in NMN-treated cells compared with HG
control groups (P<0.01; Fig.
11A-D) following the upregulation of endogenous Nampt (Fig. 11B). In HBZY-1 cells pretreated with
10 µmol/l FK866 under HG conditions (Fig. 11E), Nampt upregulation was
significantly blocked by NMN and FK866 treatment in group 3, 4 and
5 compared with group 2 (P<0.01; Fig. 11E and G), respectively. The
expression of NF-κB p65 was significantly decreased in groups 3, 4
and 5, in which expression was inhibited by both FK866 and NMN
compared with group 2 (P<0.05 and P<0.01, respectively;
Fig. 11E and H). The expression of
SIRT1 was notably increased in groups 3, 4 and 5 compared with
group 2 (P<0.05 and P<0.01, respectively), but it's
expression was not notably changed with FK866 compared with the 5D
+ control group (Fig. 11E and F).
These data suggest that NMN may protect HBZY-1 cells from
inflammatory fibrosis in response to an HG milieu by inhibiting
overexpression of endogenous Nampt and NF-κB p65, but inducing
Sirt1 activation.
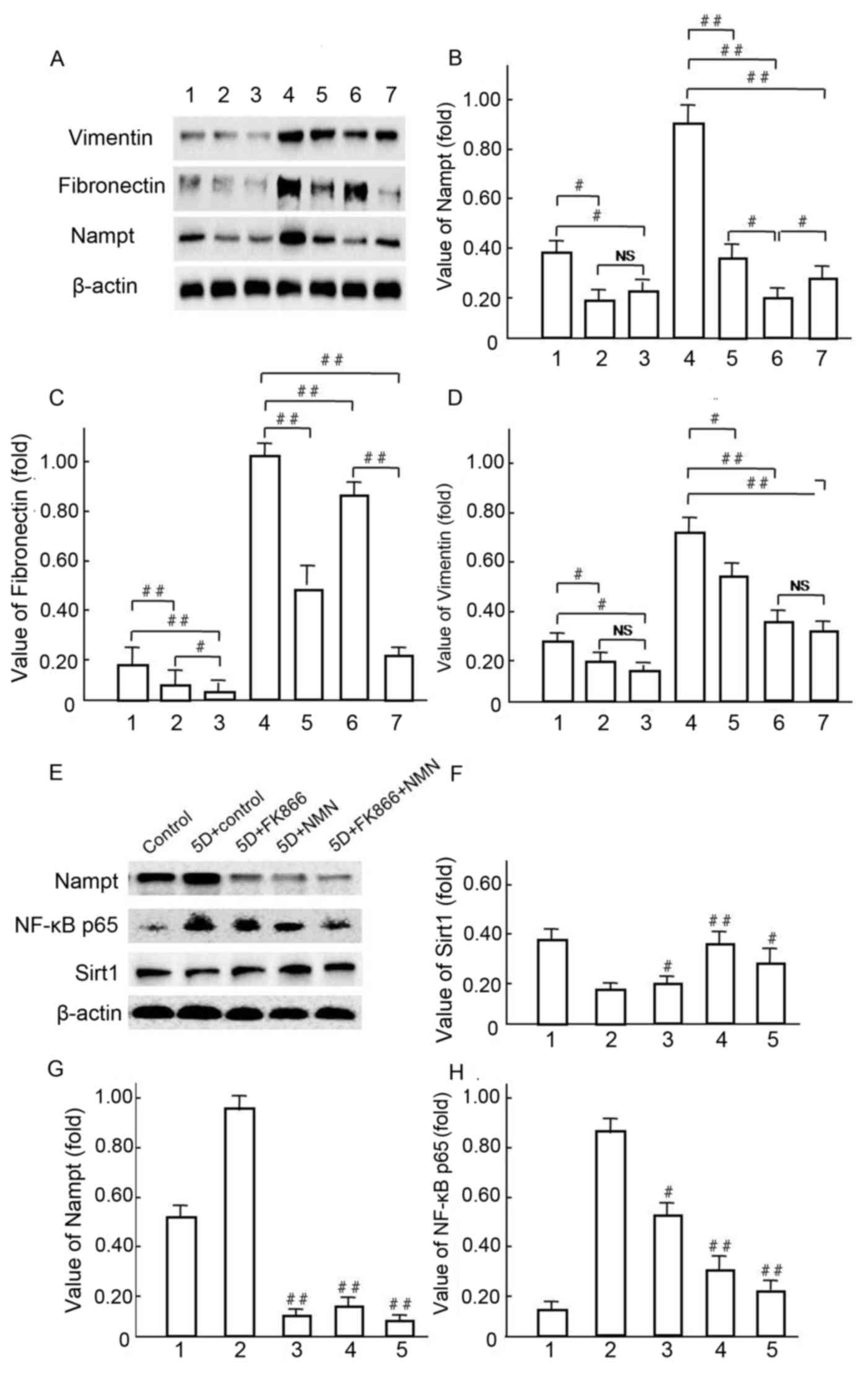 | Figure 11.Vimentin and fibronectin expression
levels were regulated by endogenous Nampt. (A-D) Following 5-day
incubation in HG medium, HBZY-1 cells were treated with 10 µmol/l
FK866 or 1 mmol/l NMN for a further 24 h. Subsequently, endogenous
Nampt, vimentin, fibronectin and β-actin protein expression levels
were measured via western blot analysis. (A-D): 1, low glucose
control group; 2, low glucose+FK866; 3, low glucose+NMN; 4, HG; 5,
HG+FK866; 6, HG+NMN; 7, HG+FK866+NMN. #P<0.05 and
##P<0.01 as indicated. (E-H) Following 4 days of
incubation in HG milieu, HBZY-1 cells were treated respectively
with 10 µmol/l FK866 and 1 mmo/l NMN for 24 h, in which the
endogenous Nampt, NF-κB p65, Sirt1 and β-actin protein expression
levels were measured via western blot analysis. (E-H): 1, NG
control group; 2, HG; 3, HG+FK866; 4, HG+NMN; 5, HG+FK866+NMN.
#P<0.05 and ##P<0.01 vs. 2. Data are
presented as the mean ± standard deviations of three experiments.
Each western blot experiment was performed in triplicate. NS, not
significant; Nampt, nicotinamide phosphoribosyltransferase; NMN,
nicotinamide mononucleotide; HG, high glucose; NF, nuclear factor;
Sirt1, sirtuin 1. |
Discussion
Nampt has been described as a regulating factor in
the cellular biosynthesis of NAD, which involves the cellular
energy metabolism and oxidation-reduction systems (19). However, as an extracellular
pro-inflammatory cytokine, Nampt/PBEF directly induces Toll-like
receptor 4 (TLR4)-mediated NF-κBp65 activation, independently of
lymphocyte antigen 96-TLR4 binding and in the absence of additional
chaperones or cofactors such as lipopolysaccharide (20). The present study demonstrated that
both endogenous Nampt and PARP-1 expression increased significantly
in diabetic rat tissues. Notably, Yu et al (17) reported that cell damage may induce
PARP-1 activation, which initiates PAR polymer formation and
apoptosis-inducing factors that result in programmed necrotic cell
death. This process may be a result of ATP and NAD reduction, as
well as increased levels of AMP and NADH in the impaired cells.
The lack of an increase in NAD in response to the
increased expression of endogenous Nampt may be attributed to the
fact that an oxidative stress reaction induced by an HG
milieugenerates NADH, and thus reverses the NAD/NADH ratio. Revollo
et al (21) demonstrated the
essential role of endogenous Nampt in the NAD biosynthetic pathway
and implicated NAD in the regulation of glucose-stimulated insulin
secretion (GSIS) in pancreatic β-cells. Accordingly, a low
pancreatic level of NAD may lead to an insulin secretion defect.
Benito-Martin et al (13)
also reported that endogenous Nampt enzymatic activity has a key
role in protecting renal cells from the adverse consequences of an
inflammatory environment, such as cell death or a prolonged
inflammatory response. The present study examined whether the
upregulation of endogenous Nampt was able to induce glomerular cell
inflammation and fibrosis in an insulin gene-mutant diabetic
C57/LB6 mouse model and observed the overexpression of pro-fibrotic
factors in glomerular cells. The findings suggested that severe
glomerular atrophy and size reductions were observed concomitantly
with endogenous Nampt upregulation when the mice suffered from DN,
while fibrosis factors, including vimentin and fibronectin were
over expressed following Nampt upregulation in glomerular cells of
themouse kidney.
Similar results were observed in HBZY-1 cells
subjected to in HG-mediated oxidative stress. The present study
indicated that Nampt is an inflammatory factor and its excessive
endogenous upregulation induced excessive expression of vimentin
and fibronectin in glomerular cells, while Sirt1 expression reduced
and NF-κB p65 increased expression under oxidative stress with HG
milieu. The results suggest that there is an association between
endogenous Nampt upregulation and the fibrosis factors. The present
data demonstrated these findings in vivo and in vitro
using cells cultured in HG milieu and the diabetic C57/LB6 mice.
The undesirable consequence may be caused by the imbalance
NAD+/NADH redox in oxidative stress and the cells
inflammatory response of NF-κB p65 over expression and activation,
which promotes inflammation and fibrosis in normal cells or
tissues. Previous studies have suggested that Sirt1 inhibition
disrupted oxidative energy metabolism and stimulated NF-κB p65 to
induce the inflammatory response characteristic of various chronic
metabolic and age-associated diseases (12,22,23).
Notably, the NF-κB p65 signaling pathway is able to stimulate
glycolytic energy flux in the context of chronic metabolic
diseases, including diabetes, where as Sirt1 activation inhibits
NF-κB p65 signaling to enhance oxidative metabolism and resolve
inflammation (24). Conversely,
NF-κB 65 signaling has been indicated to also downregulate Sirt1
activity by promoting the expression of microRNA-34a, interferon γ
and reactive oxygen species (12).
In the present study, increased expression of NF-κB
p65, FoxO1 and Bax, and decreased expression of Sirt1 were observed
in response to endogenous Nampt upregulation after cells were
subjected to oxidative stress in HG milieu. Endogenous Nampt
expression was increased significantly in a time-dependent manner
and NF-κB p65 expression peaked at 72 h, but subsequently decreased
at 96 h. Previous findings have indicated that Sirt1 has a pivotal
role in cellular energy metabolism and exhibits renal protective
functions through the deacetylation to regulate various factors,
including the transcription factors p53, NF-κB p65 subunit, signal
transducer and activator of transcription, FoxO1, and FoxO3, which
are associated with apoptosis, cellular aging, and inflammation
(25). In addition, Sirt1 also
exhibits histone deacetylation activity and exerts its renal
protective effects through the epigenetic regulation of gene
expression (26).
Based on the renal immunohistochemistry analysis
results of diabetic rats we speculate that the glomerular
inflammation and fibrosis are likely induced by endogenous Nampt
overexpression. To explore this hypothesis, the cells were exposed
to oxidative stress in high concentrations and were treated with
FK866, a specific inhibitor of Nampt (27), and NMN, which is the product of the
enzyme (28). These results also
indicated that NMN activated Sirt1 by increased NAD, due to
inhibiting the Nampt upregulation feedback loop. This likely led to
the downregulation of vimentin and fibronectin expression by
interacting withNF-κB p65. Previous research has demonstrated that
vimentin is associated with renal fibrosis (13). Coincidentally, the 5′ sequence
terminal of the vimentin gene includes NF-κB p65 binding sites, in
which active NF-κB p65 just increases vimentin expression (29). Fibronectin is key protein associated
with the extracellular matrix (ECM), which has a vital role in the
pathogenesis of inflammatory fibrosis of DN (30). Multiple growth factor (GF) binding
sites may be selected to enhance fibronectin activity to expand the
collaborative ECM-GF paradigm (31).
Furthermore, fibronectin involvement has been demonstrated in the
reactive oxygen species/c-Jun N-terminal kinase signaling pathway,
which is associated with the NF-κB p65 signaling pathways (32). Abnormal upregulation of endogenous
Nampt may lead to an imbalance between Sirt1 and NF-κB p65, and
further aggravate cells that have been seriously damaged by
oxidative stress, eventually leading to cellular dysfunction and
apoptosis (33). These findings
suggest that suitable horizontal gene expression of endogenous
Nampt may be of the utmost importance for the physical state of the
cell, whereas excess expression may promote pathological
changes.
Further analysis in the present study revealed a
decrease in vimentin expression in response to NMN, which may be
attributed to theregulation of endogenous Nampt expression by NMN
through a negative feedback pathway, as NMN is a down stream
product of Nampt enzymatic activity (33). Consistent with previous findings,
reduced NF-κB p65 expression was observed subsequent to a decrease
in endogenous Nampt expression following FK866 treatment; however,
Sirt1 expression was significantly increased under such conditions,
which was likely due to FK866 promoting cell apoptosis directly
(21). Therefore, it was speculated
that in mesangial cells, synthetic endogenous Nampt may serve a
pivotal role in the regulation of protein expression. However, the
specific underlying mechanism remains to be elucidated. For
example, the protein structure of Nampt and its regulatory
mechanism are not yet understood. Furthermore, a defined amount of
either endogenous or exogenous Nampt expression may affect enzyme
activity in the body, but either endogenous or exogenous Nampt may
lead cell to apoptosis or induce an inflammatory reaction (33,34). The
present results demonstrate that Nampt upregulation may lead an
imbalance of Sirt1 and NF-κB p65 that may induce serious
pathological processes, such as glomerular inflammatory fibrosis
during DN.
To explore the effect of these results in the whole
animal, severely diabetic rats were treated with NMN in the present
study, and mRNA was isolated and measured from kidney tissues of
the rats. The results indicated that the mRNA expression levels of
Nampt, NF-κB p65 and vimentin were significantly decreased in the
NMN-treated tissue, whereas Sirt1 expression was significantly
increased compared with the untreated groups, respectively. The
results further indicated that the Nampt-NF-κB p65 signaling
pathway is likely inhibited via NMN-induced inhibition of Nampt in
a negative feedback loop. This suggests that endogenous Nampt
overexpression may be closely involved in the pathogenesis of
glomeruli fibrosis of DN.
Together, the results of the present study provide
the first evidence, to the best of our knowledge, that excess
endogenous Nampt upregulation may have an important role in the
pathogenesis of diabetes mellitus via the NF-κB p65 and Sirt1
signaling pathway. It was observed that NMN may inhibit Nampt
upregulation via negative feedback regulation to indirectly inhibit
NF-κB p65-dependent inflammatory responses in HBZY-1 cells. The
effect was also reflected in enhanced Sirt1 expression, which is
expected to reduce inflammatory cytokine expression. In conclusion,
the present study demonstrated that NMN, a dominating product of
Nampt, may act as a compensatory, protective mechanism against
apoptosis and attenuate renal inflammatory fibrosis in the context
of DN.
Acknowledgements
The authors wish to thank Professor Liu Ming of the
Division of Metabolism, Endocrinology and Diabetes, Department of
Internal Medicine, University of Michigan Medical School (Ann
Arbor, MI, USA) for assisting with the preparation of histological
samples, and Dr Tan Ning and Dr Xianqiong Zhou of the Experiment
Center of Science, Guilin Medical University (Guilin, China) for
assistance with confocal immunofluorescence analysis.
The present study was supported by grants from the
Science and Technology Research Projects of Guangxi Universities
(grant no. YB2014266) and the National Natural Science Foundation
of China (grant nos. 31060161 and 81460164).
References
|
1
|
Aldukhayel A: Prevalence of diabetic
nephropathy among Type 2 diabetic patients in some of the Arab
countries. Int J Health Sci (Qassim). 11:1–4. 2017.PubMed/NCBI
|
|
2
|
Pofi R, Di Mario F, Gigante A, Rosato E,
Isidori AM, Amoroso A, Cianci R and Barbano B: Diabetic
nephropathy: Focus on current and future therapeutic strategies.
Curr Drug Metab. 17:497–502. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Donate-Correa J, Martin-Nunez E,
Muros-de-Fuentes M, Mora-Fernandez C and Navarro-Gonzalez JF:
Inflammatory cytokines in diabetic nephropathy. J Diabetes Res.
2015:9484172015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barutta F, Bruno G, Grimaldi S and Gruden
G: Inflammation in diabetic nephropathy: Moving toward clinical
biomarkers and targets for treatment. Endocrine. 48:730–742. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kumar Anil P, Welsh GI, Saleem MA and
Menon RK: Molecular and cellular events mediating glomerular
podocyte dysfunction and depletion in diabetes mellitus. Front
Endocrinol (Lausanne). 5:1512014.PubMed/NCBI
|
|
6
|
Garten A, Schuster S, Penke M, Gorski T,
de Giorgis T and Kiess W: Physiological and pathophysiological
roles of nampt and nad metabolism. Nat Rev Endocrinol. 11:535–546.
2015.PubMed/NCBI
|
|
7
|
Vanden Berghe T, Kaiser WJ, Bertrand MJ
and Vandenabeele P: Molecular crosstalk between apoptosis,
necroptosis, and survival signaling. Mol Cell Oncol. 2:e9750932015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fukuhara A, Matsuda M, Nishizawa M, Segawa
K, Tanaka M, Kishimoto K, Matsuki Y, Murakami M, Ichisaka T,
Murakami H, et al: Visfatin: A protein secreted by visceral fat
that mimics the effects of insulin. Science. 307:426–430. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Busch F, Mobasheri A, Shayan P, Stahlmann
R and Shakibaei M: Sirt-1 is required for the inhibition of
apoptosis and inflammatory responses in human tenocytes. J Biol
Chem. 287:25770–25781. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin Q, Geng Y, Zhao M, Lin S, Zhu Q and
Tian Z: MiR-21 regulates TNF-α-Induced CD40 expression via the
SIRT1-NF-κB pathway in renal inner medullary collecting duct cells.
Cell Physiol Biochem. 41:124–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rubattu S, Bianchi F, Busceti CL, Cotugno
M, Stanzione R, Marchitti S, Di Castro S, Madonna M, Nicoletti F
and Volpe M: Differential modulation of AMPK/PPARα/UCP2 axis in
relation to hypertension and aging in the brain, kidneys and heart
of two closely related spontaneously hypertensive rat strains.
Oncotarget. 6:18800–18818. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kauppinen A, Suuronen T, Ojala J,
Kaarniranta K and Salminen A: Antagonistic crosstalk between NF-κB
and SIRT1 in the regulation of inflammation and metabolic
disorders. Cell Signal. 25:1939–1948. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Benito-Martin A, Ucero AC, Izquierdo MC,
Santamaria B, Picatoste B, Carrasco S, Lorenzo O, Ruiz-Ortega M,
Egido J and Ortiz A: Endogenous NAMPT dampens chemokine expression
and apoptotic responses in stressed tubular cells. Biochim Biophys
Acta. 1842:293–303. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song HK, Lee MH, Kim BK, Park YG, Ko GJ,
Kang YS, Han JY, Han SY, Han KH, Kim HK, et al: Visfatin: A new
player in mesangial cell physiology and diabetic nephropathy. Am J
Physiol Renal Physiol. 295:F1485–F1494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Støy J, Edghill EL, Flanagan SE, Ye H, Paz
VP, Pluzhnikov A, Below JE, Hayes MG, Cox NJ, Lipkind GM, et al:
Insulin gene mutations as a cause of permanent neonatal diabetes.
Proc Natl Acad Sci USA. 104:15040–15044. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Konno R, Yamakawa H, Utsunomiya H, Ito K,
Sato S and Yajima A: Expression of surviving and Bcl-2 in the
normal human endometrium. Mol Hum Reprod. 6:529–534. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu L, Su Y, Paueksakon P, Cheng H, Chen X,
Wang H, Harris RC, Zent R and Pozzi A: Integrin a1/Akita
double-knockout mice on a Balb/c background develop advanced
features of human diabetic nephropathy. Kidney Int. 81:1086–1097.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim BH, Lee ES, Choi R, Nawaboot J, Lee
MY, Lee EY, Kim HS and Chung CH: Protective effects of curcumin on
renaloxidative stress and lipid metabolism in a rat model of type 2
diabetic nephropathy. Yonsei Med J. 57:664–673. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kover K, Tong PY, Watkins D, Clements M,
Stehno-Bittel L, Novikova L, Bittel D, Kibiryeva N, Stuhlsatz J,
Yan Y, et al: Expression and regulation of Nampt in human islets.
PLoS One. 8:e587672013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Camp SM, Ceco E, Evenoski CL, Danilov SM,
Zhou T, Chiang ET, Moreno-Vinasco L, Mapes B, Zhao J, Gursoy G, et
al: Unique toll-like receptor 4 activation by NAMPT/PBEF induces
NFκB signaling and inflammatory lung injury. Sci Rep. 5:131352015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Revollo JR, Körner A, Mills KF, Satoh A,
Wang T, Garten A, Dasgupta B, Sasaki Y, Wolberger C, Townsend RR,
et al: Nampt/pbef/visfatin regulates insulin secretion in beta
cells as a systemic NAD biosynthetic enzyme. Cell Metab. 6:363–375.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Caito S, Rajendrasozhan S, Cook S, Chung
S, Yao H, Friedman AE, Brookes PS and Rahman I: SIRT1 is a
redox-sensitive deacetylase that is post-translationally modified
by oxidants and carbonyl stress. FASEB J. 24:3145–3159. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lim JH, Lee YM, Chun YS, Chen J, Kim JE
and Park JW: Sirtuin 1 modulates cellular responses to hypoxia by
deacetylating hypoxia-inducible factor 1alpha. Mol Cell.
38:864–878. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Salminen A, Huuskonen J, Ojala J,
Kauppinen A, Kaarniranta K and Suuronen T: Activation of innate
immunity system during aging: NF-kB signaling is the molecular
culprit of inflamm-aging. Ageing Res Rev. 7:83–105. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hori YS, Kuno A, Hosoda R and Horio Y:
Regulation of FOXOs and p53 by SIRT1 modulators under oxidative
stress. PLoS One. 8:e738752013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wakino S, Hasegawa K and Itoh H: Sirtuin
and metabolic kidney disease. Kidney Int. 88:691–698. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nakahata Y, Sahar S, Astarita G, Kaluzova
M and Sassone-Corsi P: Circadian control of the NAD+ salvage
pathway by CLOCK-SIRT1. Science. 324:654–657. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Spinnler R, Gorski T, Stolz K, Schuster S,
Garten A, Beck-Sickinger AG, Engelse MA, de Koning EJ, Körner A,
Kiess W, et al: The adipocytokine Nampt and its product NMN have no
effect on beta-cell survival but potentiate glucose stimulated
insulin secretion. PLoS One. 8:e541062013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lilienbaum A and Paulin D: Activation of
the human vimentin gene by the Tax human T-cell leukemia virus. I.
Mechanisms of regulation by the NF-kappa B transcription factor. J
Biol Chem. 268:2180–2188. 1993.PubMed/NCBI
|
|
30
|
El Mesallamy HO, Ahmed HH, Bassyouni AA
and Ahmed AS: Clinical significance of inflammatory and fibrogenic
cytokines in diabetic nephropathy. Clin Biochem. 45:646–650. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu J and Clark RA: Fibronectin at select
sites binds multiple growth factors (GF) and enhances their
activity: Expansion of the collaborative ECM-GF paradigm. J Invest
Dermatol. 134:895–901. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tong YF, Liu Y, Hu ZX, Li ZC and Agula A:
Protocatechuic aldehyde inhibits TNF-α-induced fibronectin
expression in human umbilical vein endothelial cells via a c-Jun
N-terminal kinase dependent pathway. Exp Ther Med. 11:277–282.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alaee M, Khaghani S, Behroozfar K, Hesari
Z, Ghorbanhosseini SS and Nourbakhsh M: Inhibition of nicotinamide
phosphoribosyltransferase induces apoptosis in estrogen
receptor-positive MCF-7 breast cancer cells. J Breast Cancer.
20:20–26. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dahl TB, Holm S, Aukrust P and Halvorsen
B: Visfatin/NAMPT: A multifaceted molecule with diverse roles in
physiology and pathophysiology. Annu Rev Nutr. 32:229–243. 2012.
View Article : Google Scholar : PubMed/NCBI
|