Introduction
Diabetes mellitus is an increasing global health
problem. Estimates for 2013 by the International Diabetes
Federation (IDF) indicate that a total of 382 million have diabetes
in 2013, and the number is expected to rise to 592 million by 2035
(1–3). Diabetic nephropathy (DN) is one of the
most common complications of diabetes. An estimate of 30–50% of
patients with diabetes develop renal manifestations (4–6). DN
often leads to chronic kidney disease (CKD). Nearly 44% of
end-stage renal disease (ESRD) patients that require hemodialysis
are diabetic nephropathy patients (7). Inflammation and oxidative stress are
associated with the pathogenesis of DN (8). Transcription factors such as nuclear
factor κ of activated B cells (NF-κB) (9), pro-inflammatory cytokines such as tumor
necrosis factor-α (TNF-α) and interleukin 1 (IL-1) (10) are associated with inflammatory
pathways in DN. Thus, it is important to treat DN using
anti-inflammatory drugs.
Research using experimental models of type 2
diabetes with nephropathy may provide an enhanced understanding of
this complication in this multifactorial disease (11). Qi et al (12) reported that KK/HlJ mice are more
prone to DN, whereas the most widely used C57BL/6J mice are
relatively resistant to developing DN. Rhein lysinate (RHL) is the
lysine salt of rhein, which is one of the active components of
rhubarb root (Rheum palmatum Linn or Rheum tanguticum
Maxim) (13). A previous study
by our group found that RHL reduced the levels of TNF-α, IL-6 and
NF-κB, decreased the incidence of glomerulonephritis and prolonged
the median survival time of senescence-prone inbred strain 10 mice
(14). However, to the best of our
knowledge, the effect of RHL on DN has not been previously
reported. The present study investigated the effect of RHL on DN in
KK/HlJ mice.
Materials and methods
Chemicals and reagents
Rhein lysinate (RHL) was synthesized at the Oncology
Department of the Institute of Medicinal Biotechnology, Chinese
Academy of Medical Sciences and Peking Union Medical College
(Beijing, China; patent no. 2008100890258). The structural formula
of this compound is presented in our previous study (13). Streptozotocin (STZ) was obtained from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Malondialdehyde
(MDA), superoxide dismutase (SOD) and glutathione peroxidase
(GSH-px) kits were obtained from Nanjing Jiancheng Co. (Nanjing,
China). Antibodies targeting TNF-α (3707s), IL-6 (12912s), NF-κB
(8242s) and phosphorylated (p)-NF-κB (3033s) were purchased from
Cell Signaling Technology, Inc. (Danvers, MA, USA). Antibody
targeting β-actin (sc-8432) was provided by Santa Cruz
Biotechnology (Dallas, TX, USA). The appropriate anti-mouse (7076s)
and anti-rabbit (7074s) horseradish peroxidase-conjugated secondary
antibodies were obtained from Cell Signaling Technology, Inc.
Prestained protein marker p7708V was provided by New England
Biolabs, Ltd. (Ipswitch, MA, USA). Immobilon™ western kit and
polyvinylidene difluoride (PVDF) membranes were obtained from
Millipore (Billerica, MA, USA).
Animals and induction of DN in inbred
mice
Induction of DN was performed according to a
previous protocol (12). The inbred
male C57BL/J mice (n=12) and KK/HlJ mice (n=36) (weight, 18–24 g;
age, 8 weeks) and mouse food used in the present study were
purchased from Beijing HFK Bioscience Co. (Beijing, China). The
diet contained water (≤8%), crude protein (≥18%), crude fat (≥6%),
crude fiber (≥5%), crude ash (≤7%), as well as minerals and trace
elements. All protocols were approved by the institutional animal
care and use committee of Beijing Hospital (Beijing, China). Mice
were housed in an environmentally-controlled facility maintained on
an automatic 12-h light/dark cycle. Food and water were provided
ad libitum throughout the study. The study used the
following 4 groups: C57BL/J control mice (n=12), the KK/HlJ model
mice (n=12), the 25 mg/kg/day RHL-treated KK/HlJ mice (n=12) and
the 50 mg/kg/day RHL-treated KK/HlJ mice (n=12). At 10 weeks of
age, STZ was administered to KK/HlJ mice by intraperitoneal
injection (50 mg/kg/day, made freshly in 0.1 mol/l citrate buffer,
pH 4.5) for 5 consecutive days with normal diet and mice received
the diabetic diet for the remaining treatment time. After all of
STZ injections, 25 or 50 mg/kg/day RHL was respectively
administered to the animals by gavage in the 25 or 50 mg/kg/day
RHL-treated KK/HlJ group for 15 weeks.
Analysis of urinary albumin
Urinary albumin was assessed by determining the
albumin-creatinine ratio (ACR) in morning urine. Urine was
collected monthly by a home-made mouse urine collection device
which used a 96-well plate as the floor. Mice were able to move
freely on this 96-well plate until they naturally urinated. Urine
was collected by a pipette without contamination by feces. Urinary
albumin and creatinine concentrations were determined using a
microalbumin/creatinine reagent kit (SIEM-6011A) supplied by
Siemens medical solutions diagnostics (Tarrytown, NY, USA).
Measurement of laboratory parameters
in serum
Serum was collected at the end of the experiment.
Serum creatinine, urea nitrogen and blood glucose were measured in
the clinical laboratory using standard protocols.
Analysis of antioxidant activity
At the end of the experiment, mice were anesthetized
by intraperitoneal injection of 10% chloral hydrate and sacrificed;
the kidneys were quickly removed and placed in cold PBS. Renal
tissue (50 mg) was dissected and homogenized in a glass Teflon
homogenizer containing 450 ml precooled PBS and the homogenate was
then centrifuged at 3,000 × g for 15 min at 4°C. The activities of
SOD and GSH-px and the MDA content in the obtained supernatant were
measured using test kits, referring to the supplier's manual.
Histological and immunohistochemical
analysis
For histological examination, kidney tissues fixed
with 4% buffered paraformaldehyde were dehydrated in ethanol and
embedded in paraffin wax. The embedded kidney tissues were serially
sectioned into 3-µm slices and stained with hematoxylin (staining 5
min)-eosin (staining 2 min) (Beijing Solarbio Science &
Technology Co., Ltd., Beijing China) at room temperature. To better
characterize the inflammation of kidney tissues,
immunohistochemistry detection of TNF-α and IL-6 was performed.
After dewaxing with xylene, samples were incubated with 3%
H2O2 to block endogenous peroxidase. All
slices were incubated with 5% bovine serum albumin (Sigma Aldrich;
Meck KGaA, Darmstadt, Germany) at 37°C for 1 h to block
non-specific binding. Subsequently, the slices were incubated with
anti-TNF-α (1:1,000) or anti-IL-6 (1:1,000) antibodies at 4°C
overnight according to the manufacturer's protocol. As a negative
control, the primary antibody was replaced with 5% bovine serum
albumin at 4°C overnight. Positive staining was identified by
visual observation of a yellow/brown pigmentation under the light
microscope. Images were captured with an Olympus IX81 inverted
microscope (Olympus, Tokyo, Japan).
Western blot analysis
The expression of TNF-α, IL-6 and NF-κB, as well as
the phosphorylation of NF-κB in the kidneys were determined by
western blot analysis using a procedure identical to that used in a
previous study by our group (14).
Statistical analysis
Values are expressed as the mean ± standard
deviation. Statistical analysis was performed using SPSS 11.0 for
Windows (SPSS, Inc., Chicago, IL, USA). Differences between groups
of values were compared using a one-way analysis of variance and
the Student-Newman-Keuls test. P<0.05 was considered to indicate
a statistically significant difference.
Results
RHL improves the kidney function of
mice with DN
The ACR in mice in each group was determined at 5
weeks. Compared with that in the C57BL/6J control group, the ACR
was significantly increased in the KK/HlJ model group (P<0.05;
Table I). Compared with those in the
KK/HlJ model group, RHL (25 and 50 mg/kg/day) significantly
decreased the ACR (P<0.05; Table
I). The ACR was 40.8±8.6, 486.5±82.9, 424.7±78.6, 385.1±52.4
µg/mg in the C57BL/6J control group, KK/HlJ model group, 25 and 50
mg/kg/day RHL treatment groups, respectively at 5 weeks (Table I).
 | Table I.Effect of RHL on ACR, blood glucose,
creatinine and urea. |
Table I.
Effect of RHL on ACR, blood glucose,
creatinine and urea.
|
|
| KK/HlJ |
|---|
|
|
|
|
|---|
| Groups | C57BL/6J control | Model | RHL 25 mg/kg/day | RHL 50 mg/kg/day |
|---|
| ACR (µg/mg) 5
weeks |
40.8±8.6 |
486.5±82.9a |
424.7±78.6a,b |
385.1±52.4a,b |
| ACR (µg/mg) 15
weeks |
45.12±10.5 |
553.6±82.1a |
442.3±61.5a,b |
310.5±49.7a,b |
| Blood glucose
(mmol/l) |
6.53±0.51 |
12.36±3.0a |
9.53±2.81a,b |
8.50±1.90a,b |
| Creatinine
(mmol/l) |
65.5±24.8 |
130.0±28.8a |
92.5±26.7a,b |
70.0±25.1b |
| Urea (mmol/l) |
7.0±2.1 |
12.5±3.2a |
11.7±1.8a |
9.6±2.2a,b |
RHL protects kidney damage in diabetic
KK/HlJ mice
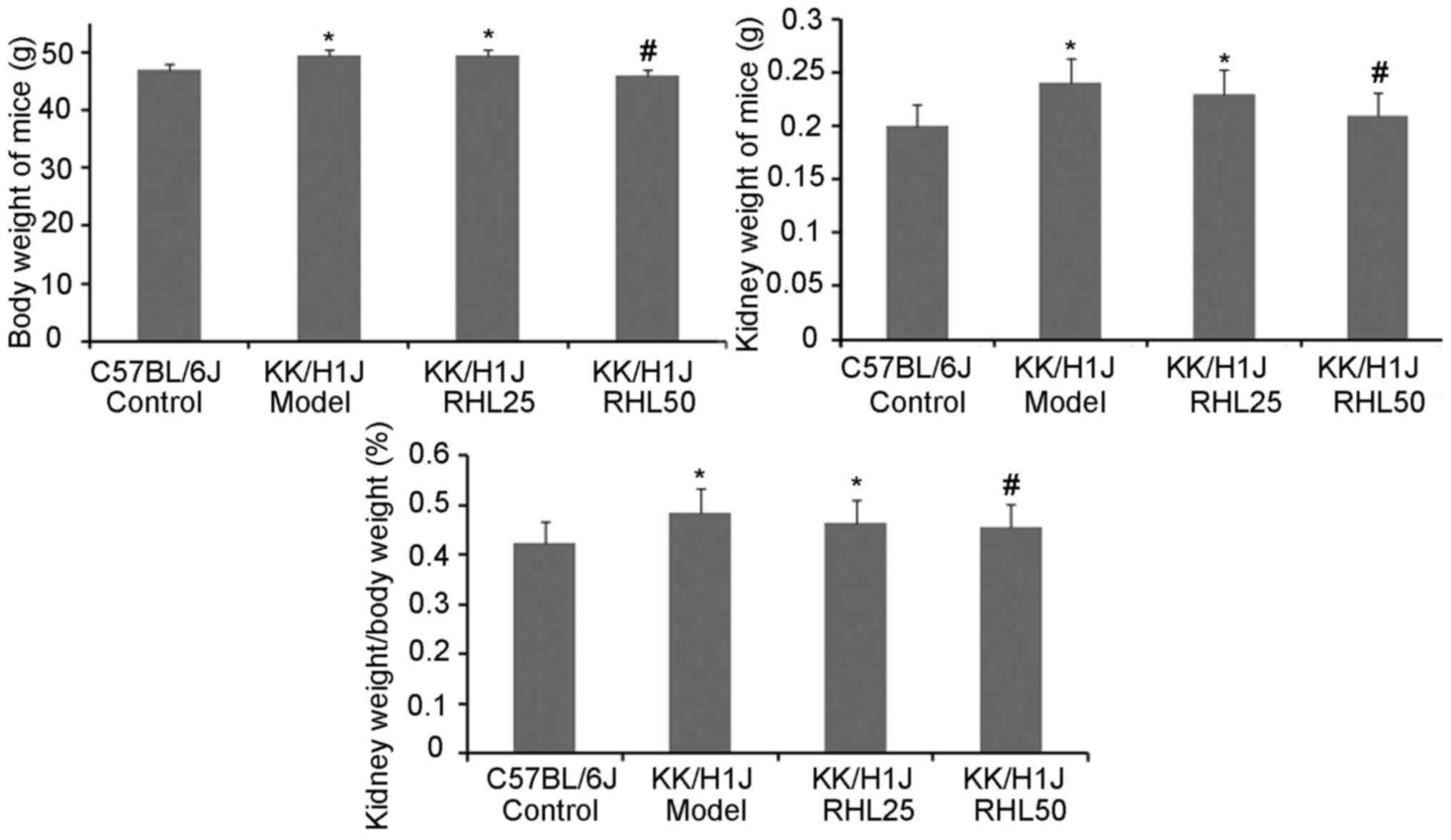
To assess the effects of RHL on DN in KK/HlJ mice,
the effect of RHL on body and kidney weight was investigated
(Fig. 1). Compared with those in the
C57BL/6J control group, the body and kidney weights in the KK/HlJ
model group and the 25 mg/kg/day RHL-treated group were increased.
Compared with those in the KK/HlJ model group, the body and kidney
weights in the 50 mg/kg/day RHL-treated group were decreased. The
kidney weight-to-body weight ratio in KK/HlJ and C57BL/6J mice was
also determined. As presented in Fig.
1, a significant increase in the kidney weight-to-body weight
ratio was observed in all KK/HlJ mice, compared with that in
C57BL/6J mice. The kidney weight-to-body weight ratio in the RHL 50
mg/kg/day group was decreased compared with that in the KK/HlJ
model group. In addition, renal hypertrophy and hydronephrosis were
observed in the KK/HlJ groups, including the RHL treatment
groups.
RHL decreases the ACR, as well as
blood glucose, creatinine and urea in mice with DN
To assess the effect of RHL on the kidney function
of KK/HlJ mice with DN, the ACR, as well as blood glucose,
creatinine and urea were detected at 15 weeks (Table I). Compared with those in the
C57BL/6J control group, the ACR, as well as blood glucose,
creatinine and urea were increased in the KK/HlJ model group, and
certain parameters were also increased in the 25 and 50 mg/kg/day
RHL-treated groups. Compared with those in the KK/HlJ model group,
RHL (25 and 50 mg/kg/day) decreased the ACR, as well as blood
glucose and creatinine, and RHL at 50 mg/kg/day also decreased the
levels of urea (Table I).
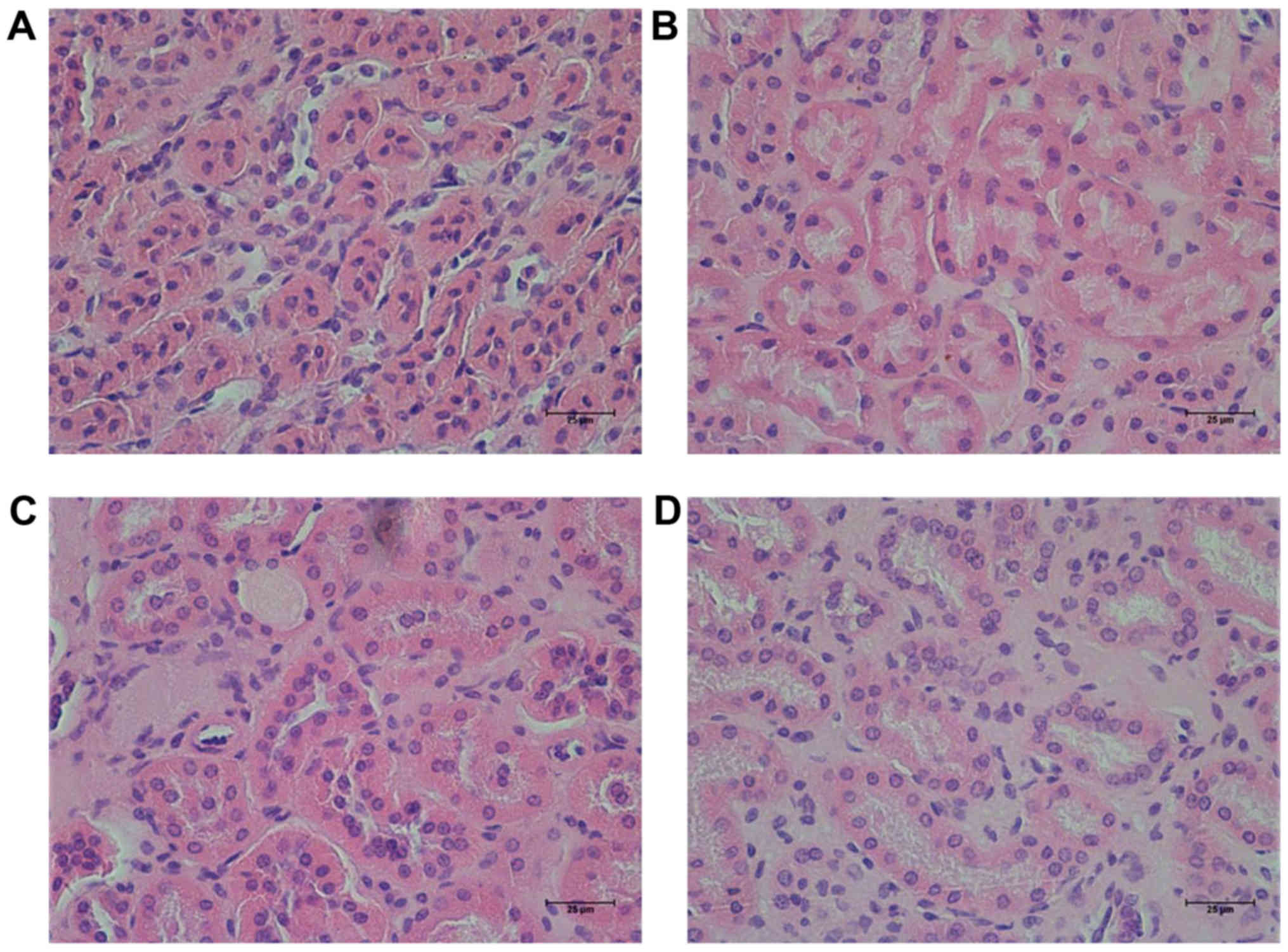
RHL improves kidney function in mice
with DN
In the present study, it was observed that the size
and weight (0.05 g) of one of the kidneys from one mouse was
decreased in the KK/HlJ model group. However, this was not observed
in the other groups. Hematoxylin-eosin staining revealed that the
characteristic changes of the kidney in the KK/HlJ mouse models of
DN were renal tubular epithelial cell edema (Fig. 2), which demonstrated that renal
tubular epithelial cell edema was shallow. No other structural
changes were observed in the DN model group. In comparison,
administration of RHL (25 and 50 mg/kg/day) improved renal tubular
epithelial cell edema (Fig. 2).
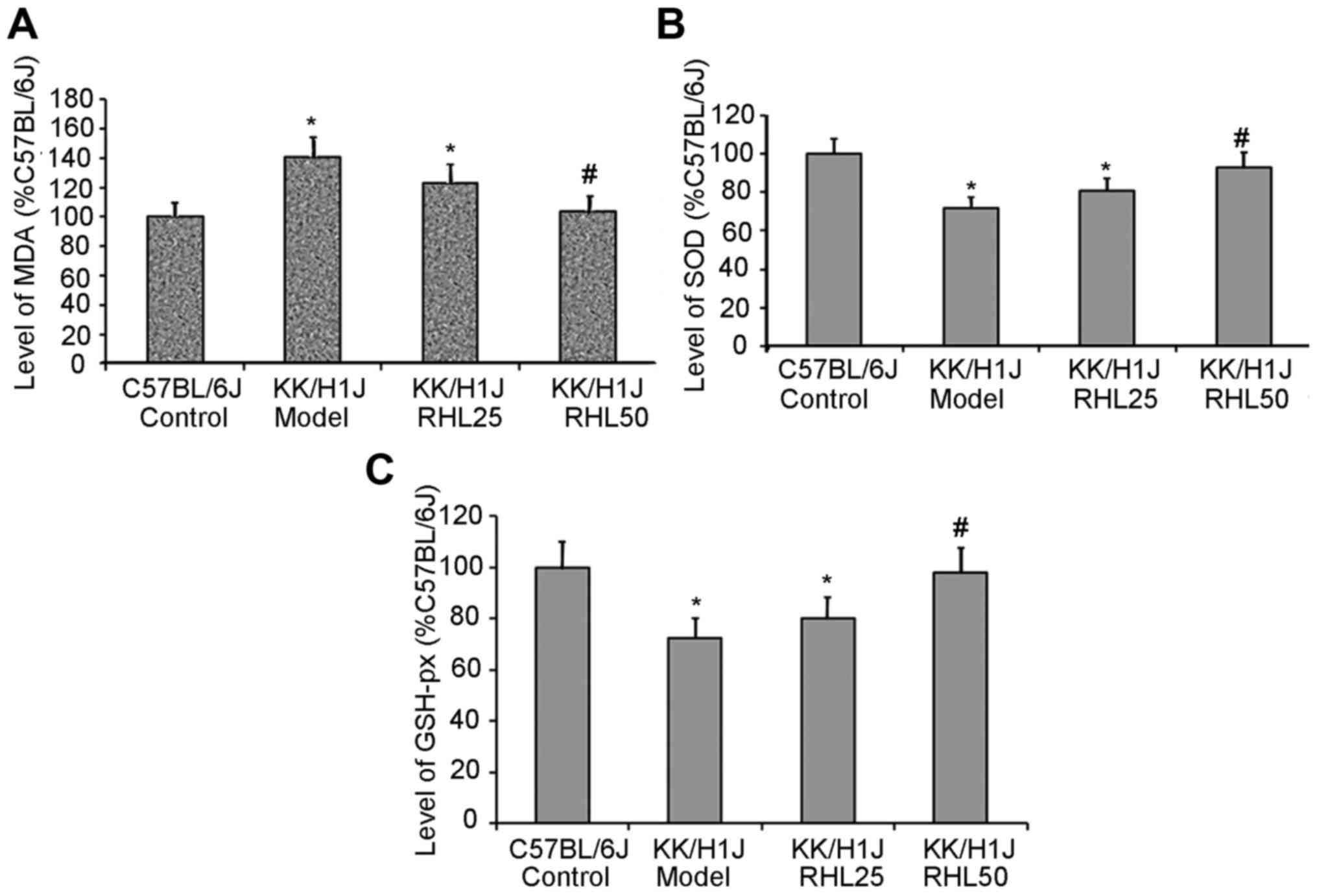
RHL increases the activity of SOD and
GSH-px and decreases the levels of MDA in kidney tissue
In the KK/HlJ model group, the activities of SOD and
GSH-px were 28 and 27% lower, respectively, than those in the
C57BL/6J control group; however, the MDA levels were 40% higher. In
the KK/HlJ group treated with 25 mg/kg/day RHL, the activities of
SOD and GSH-px were 12 and 10% higher, respectively, than those in
the KK/HlJ model group; however, the MDA levels were 13% lower. In
the KK/HlJ group treated with 50 mg/kg/day RHL, the activities of
SOD and GSH-px were 30 and 35% higher, respectively, than those in
the KK/HlJ model group; however, the MDA levels were 26% lower
(Fig. 3).
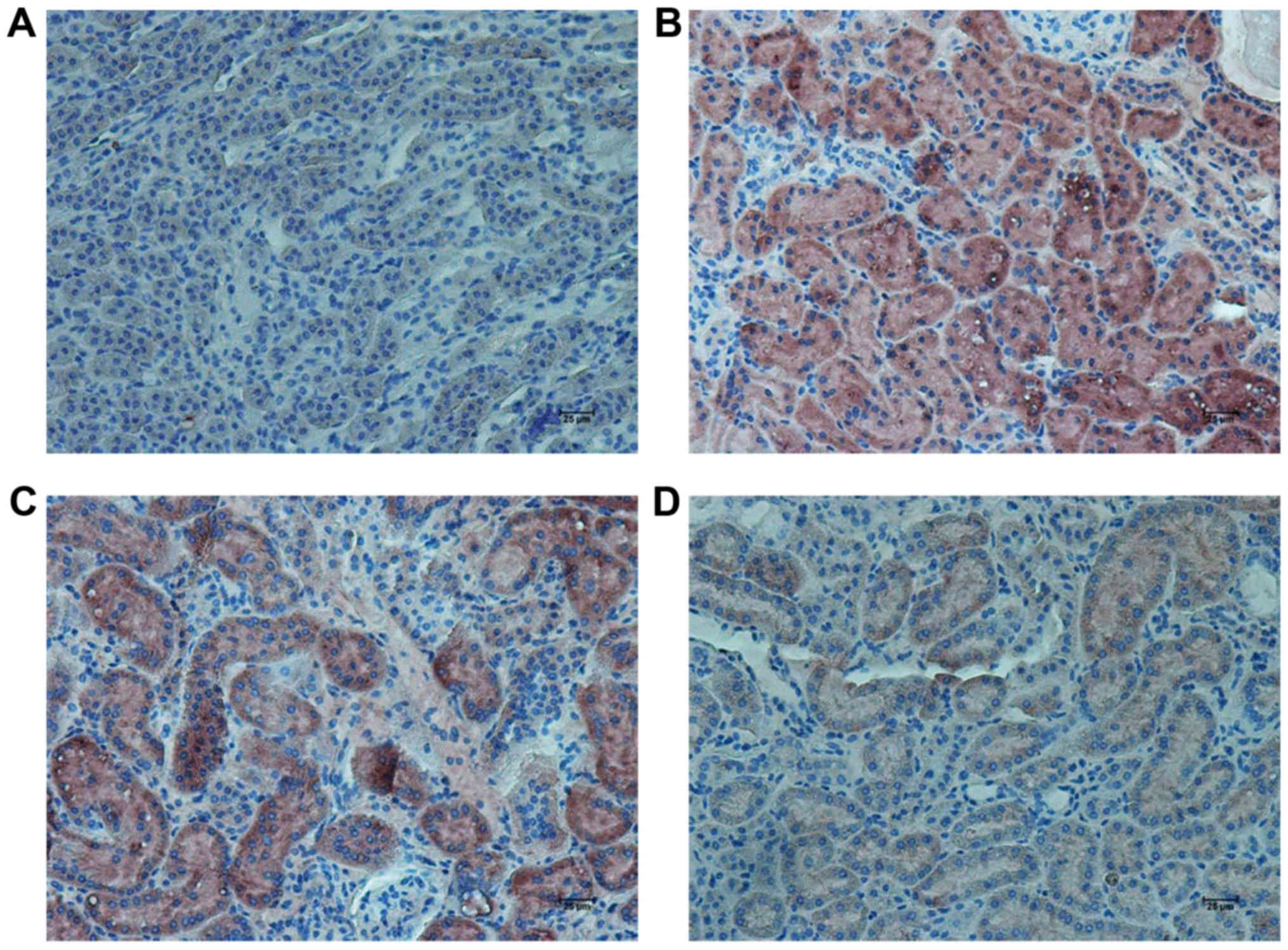
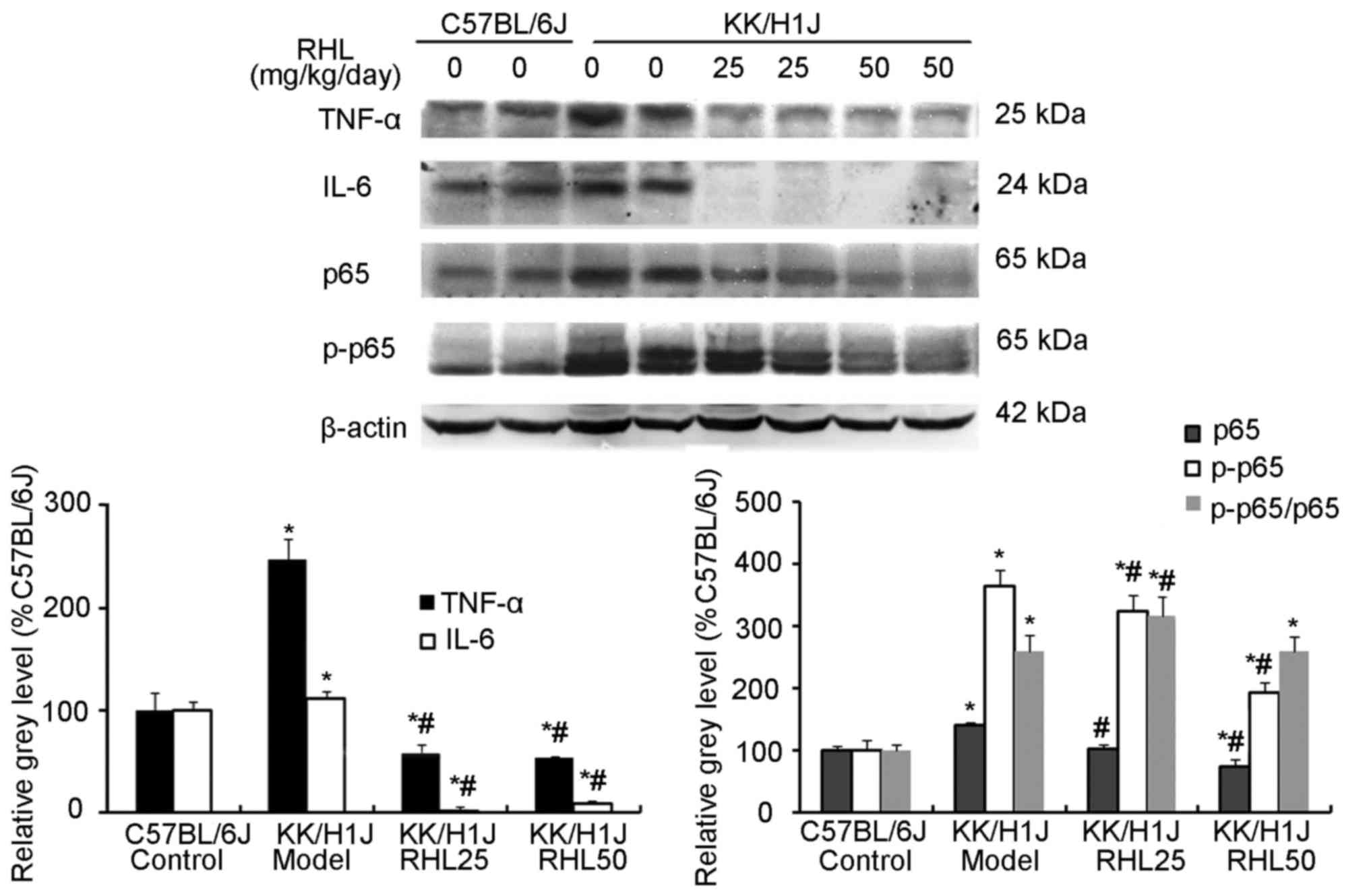
RHL suppresses the expression of
inflammatory factors and associated proteins in the kidney of mice
with DN
In the KK/HlJ model group, the TNF-α expression
levels were higher than those in the C57BL/6J control group. RHL
treatment (25 and 50 mg/kg/day) decreased the expression of TNF-α
and IL-6, as indicated by immunohistochemical and western blot
analysis (Figs. 4–6). In addition, compared with the C57BL/6J
control group, the KK/HlJ model group exhibited increased
expression and phosphorylation of NF-κB. Compared with the KK/HlJ
model group, the RHL groups (25 and 50 mg/kg/day) exhibited
decreased phosphorylation and expression of NF-κB. Compared with
the C57BL/6J control group, the KK/HlJ model and RHL groups (25 and
50 mg/kg/day) exhibited increased p-NF-κB/NF-κB ratios; however,
compared with the KK/HlJ model group, 25 mg/kg/day RHL treatment
exhibited increased p-NF-κB/NF-κB ratios and 50 mg/kg/day RHL
treatment exhibited no significant effect on the p-NF-κB/NF-κB
ratios (Fig. 6).
Discussion
The role of rhein and its analogues in the
management of chronic kidney disease (CKD) has been addressed in
several previous studies. Rhein improved the symptoms of
nephropathy through decreasing the production of proinflammatory
cytokines, including IL-1β, prostaglandin E2 and TNF-α and
inhibiting the expression of transforming growth factor-β1
(15). A previous study by our group
also observed that RHL protects the kidney from impairment in a
senescence-prone inbred strain 10 mice (14). However, the effect of RHL, the lysin
salt of rhein, on DN has remained elusive. In the present study, a
KK/HlJ mouse model of DN was induced by STZ injection and a
specific diet. The ACR was detected at 5 weeks; compared with that
in the C57BL/6J control group, the ACR was increased in the KK/HlJ
model group and in the KK/HlJ RHL treatment groups. It was
demonstrated that the kidney function in the KK/HlJ model group and
the KK/HlJ RHL treatment groups was impaired. The levels of ACR
were also detected at 15 weeks; overall, it was demonstrated that
RHL improved kidney function impairment of KK/HlJ mice at 5 and 15
weeks. After 16 weeks of treatment, the mice were sacrificed, and
kidney weights and the kidney weight-to-body weight ratio in the
KK/HlJ model group were revealed to be higher than those in the
C57BL/6J control group, and to be reduced by treatment with RHL at
50 mg/kg/day. A previous study by our group reported that RHL
decreased the body weight and blood glucose in high-fat diet and
STZ-induced diabetic mice (16). In
the present study, it was also observed that RHL treatment at 25
and 50 mg/kg/day decreased blood glucose, and RHL at 50 mg/kg/day
also decreased the body weight, compared with that in the KK/HlJ
model group. Blood biochemistry analysis indicated that creatinine
and urea, the biomarkers of kidney function, were increased in the
KK/HlJ model group, compared with those in the C57BL/6J control
group, which was decreased by RHL treatment.
The pathophysiological characteristics of DN include
renal hypertrophy, decrease of renal function, glomerular and
tubular basement membranes thickening, mesangial matrix expansion,
ultimately causing glomerulosclerosis and interstitial fibrosis
(17–19). In the present study, a renal function
decrease, renal hypertrophy and renal tubular edema were observed,
while glomerulosclerosis and interstitial fibrosis were not seen.
It may be speculated that DN in KK/HlJ mice induced in the present
study was at its early stage. Thus, early DN may be cured by RHL;
however, if glomerulosclerosis and interstitial fibrosis had been
present, DN would have hardly been cured. Oxidative stress has been
reported to be responsible for the development of peripheral
diabetic neuropathy (20,21). MDA, SOD and GSH-px are indicators of
the oxidative stress status (22,23). In
the present study, the activities of SOD and GSH-px in the KK/HlJ
model group were lower than those in the C57BL/6J control group;
however, the levels of MDA in the KK/HlJ model group were higher
than those in the C57BL/6J control group. These results indicated
that RHL increased the levels of SOD and GSH-px and decreased the
levels of MDA in kidney tissues of KK/HlJ mice. Thus, it may be
deduced that RHL protects the kidney by reducing the levels of
reactive oxygen species. These results were similar to those of a
previous study by our group (14).
Inflammatory factors reportedly have a role in
diabetes and in the progression of DN (24–26).
Furthermore, diabetes is associated with increased levels of
inflammatory biomarkers, including IL-6 and TNF-α (27,28). The
TNF-α-308G/A polymorphism was reported to be associated with the
expression levels of TNF-α and may be as a genetic susceptibility
factor for DN (29). However, the
effect of RHL on inflammatory factors associated with DN has
remained elusive. The present results demonstrated that the levels
of TNF-α and IL-6 in the KK/HlJ model group were higher than those
in the C57BL/6J control group, and the expression levels of TNF-α
and IL-6 were markedly suppressed in RHL-treated KK/HlJ mice.
Therefore, it was speculated that inflammatory factors (TNF-α and
IL-6) take part in the progression of DN in KK/HlJ mice and that
RHL treatment decreases the production of inflammatory factors to
thereby protect renal function.
TNF-α is one of the key proinflammatory cytokines
and is involved in a number of inflammatory processes. TNF-α
activates the NF-κB transcription factor. NF-κB has two functions:
First, in malignant cells, it promotes cell survival and
proliferation. Furthermore, NF-κB activates the immune response,
particularly the production of proinflammatory cytokines (30), and inflammation mediated by NF-κB has
a critical role in the pathogenesis of DN (31). The present study revealed that TNF-α
was involved in renal tubular edema via the TNF-α/NF-κB biochemical
pathway. RHL decreased the expression of TNF-α and NF-κB, and
inhibited the phosphorylation of NF-κB downstream of TNF-α.
Therefore, RHL had the ability to inhibit the immune response by
directly or indirectly blocking the TNF-α/NF-κB biochemical
pathway.
In conclusion, oxygen free radicals and inflammatory
factors took part in the progression of DN in KK/HlJ mice. RHL (25
and 50 mg/kg/day) significantly decreased kidney inflammation by
reducing the levels of oxygen free radicals, blocking the
TNF-α/NF-κB biochemical pathway and reducing renal function
impairment. This finding is expected to inspire future
investigations on the efficacy of RHL in DN.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81671391),
Beijing Hospital Nova Project (grant no. BJ-2016-033) and the
General Program of Natural Science Foundation of Hebei Province of
China (grant no. H2012401030).
References
|
1
|
Forouhi NG and Wareham NJ: Epidemiology of
diabetes. Medicine (Abingdon). 42:698–702. 2014.PubMed/NCBI
|
|
2
|
Harjutsalo V and Groop PH: Epidemiology
and risk factors for diabetic kidney disease. Adv Chronic Kidney
Dis. 21:260–266. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Noubiap JJ, Naidoo J and Kengne AP:
Diabetic nephropathy in Africa: A systematic review. World J
Diabetes. 6:759–773. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Assogba GF, Couchoud C, Roudier C, Pornet
C, Fosse S, Romon I, Druet C, Stengel B and Fagot-Campagna A:
Prevalence, screening and treatment of chronic kidney disease in
people with type 2 diabetes in France: The ENTRED surveysnd (2001
and 2007). Diabetes Metab. 38:558–566. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bakris GL: Recognition, pathogenesis, and
treatment of different stages of nephropathy in patients with type
2 diabetes mellitus. Mayo Clin Proc. 86:444–456. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thomas MC, Weekes AJ, Broadley OJ, Cooper
ME and Mathew TH: The burden of chronic kidney disease in
Australian patients with type 2 diabetes (the NEFRON study). Med J
Aust. 185:140–144. 2006.PubMed/NCBI
|
|
7
|
Chokhandre MK, Mahmoud MI, Hakami T, Jafer
M and Inamdar AS: Vitamin D & its analogues in type 2 diabetic
nephropathy: A systematic review. J Diabetes Metab Disord.
14:582015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu J, Wang C, Liu F, Lu Y and Cheng J:
Metabonomics revealed xanthine oxidase-induced oxidative stress and
inflammation in the pathogenesis of diabetic nephropathy. Anal
Bioanal Chem. 407:2569–2579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mezzano S, Aros C, Droguett A, Burgos ME,
Ardiles L, Flores C, Schneider H, Ruiz-Ortega M and Egido J:
NF-kappaB activation and overexpression of regulated genes in human
diabetic nephropathy. Nephrol Dial Transplant. 19:2505–2512. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ayepola OR, Cerf ME, Brooks NL and
Oguntibeju OO: Kolaviron, a biflavonoid complex of Garcinia
kola seeds modulates apoptosis by suppressing oxidative stress
and inflammation in diabetes-induced nephrotoxic rats.
Phytomedicine. 21:1785–1793. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Soler MJ, Riera M and Batlle D: New
experimental models of diabetic nephropathy in mice models of type
2 diabetes: Efforts to replicate human nephropathy. Exp Diabetes
Res. 2012:6163132012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qi Z, Fujita H, Jin J, Davis LS, Wang Y,
Fogo AB and Breyer MD: Characterization of susceptibility of inbred
mouse strains to diabetic nephropathy. Diabetes. 54:2628–2637.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin YJ and Zhen YS: Rhein lysinate
suppresses the growth of breast cancer cells and potentiates the
inhibitory effect of taxol in athymic mice. Anticancer Drugs.
20:65–72. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu G, Liu J, Zhen YZ, Xu R, Qiao Y, Wei J,
Tu P and Lin YJ: Rhein lysinate increases the median survival time
of SAMP10 mice: Protective role in the kidney. Acta Pharmacol Sin.
34:515–521. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meng Z, Yan Y, Tang Z, Guo C, Li N, Huang
W, Ding G, Wang Z, Xiao W and Yang Z: Anti-hyperuricemic and
nephroprotective effects of rhein in hyperuricemic mice. Planta
Med. 81:279–285. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin YJ, Hu G, Li KJ, Zhao YF, Wei J and
Zhen YZ: The protection of Rhein lysinate to liver in diabetic mice
induced by high-fat diet and streptozotocin. Arch Pharm Res.
38:885–892. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cao Z and Cooper ME: Pathogenesis of
diabetic nephropathy. J Diabetes Investig. 2:243–247. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dronavalli S, Duka I and Bakris GL: The
pathogenesis of diabetic nephropathy. Nat Clin Pract Endocrinol
Metab. 4:444–452. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Furukawa M, Gohda T, Tanimoto M and Tomino
Y: Pathogenesis and novel treatment from the mouse model of type 2
diabetic nephropathy. ScientificWorldJournal. 2013:9281972013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dobretsov M, Romanovsky D and Stimers JR:
Early diabetic neuropathy: Triggers and mechanisms. World J
Gastroenterol. 13:175–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wattanathorn J, Thiraphatthanavong P,
Muchimapura S, Thukhammee W, Lertrat K and Suriharn B: The combined
extract of Zingiber officinale and Zea mays (purple
color) improves neuropathy, oxidative stress, and axon density in
streptozotocin induced diabetic rats. Evid Based Complement
Alternat Med. 2015:3010292015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Meng H, Zhang D and Yang H: Effects of
amyloid precursor protein 17 peptide on the protection of diabetic
encephalopathy and improvement of glycol metabolism in the diabetic
rat. J Diabetes Res. 2013:6898412013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang XW, Liu FQ, Guo JJ, Yao WJ, Li QQ,
Liu TH and Xu LP: Antioxidation and anti-inflammatory activity of
Tang Bi Kang in rats with diabetic peripheral neuropathy. BMC
Complement Altern Med. 15:662015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Al-Rejaie SS, Aleisa AM, Abuohashish HM,
Parmar MY, Ola MS, Al-Hosaini AA and Ahmed MM: Naringenin
neutralises oxidative stress and nerve growth factor discrepancy in
experimental diabetic neuropathy. Neurol Res. 37:924–933. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Semeraro F, Cancarini A, Dell'Omo R,
Rezzola S, Romano MR and Costagliola C: Diabetic retinopathy:
Vascular and inflammatory disease. J Diabetes Res. 2015:5820602015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu JS, Liu Y, Shi R, Lu X, Ma YM and Cheng
NN: Effects of combinations of Xiexin decoction constituents on
diabetic nephropathy in rats. J Ethnopharmacol. 157:126–133. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Akram KA, Tofighiyan T and Rakhshani MH:
Effects of synbiotics on inflammatory markers in patients with type
2 diabetes mellitus. Glob J Health Sci. 7:(7 Spec No). 1–5.
2015.
|
|
28
|
Nolasco EL, Zanoni FL, Nunes FP, Ferreira
SS, Freitas LA, Silva MC and Martins JO: Insulin modulates liver
function in a type I diabetes rat model. Cell Physiol Biochem.
36:1467–1479. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peng Y and Li LJ: TNF-α-308G/A
polymorphism associated with TNF-α protein expression in patients
with diabetic nephropathy. Int J Clin Exp Pathol. 8:3127–3131.
2015.PubMed/NCBI
|
|
30
|
Wang SS, Purdue MP, Cerhan JR, Zheng T,
Menashe I, Armstrong BK, Lan Q, Hartge P, Kricker A, Zhang Y, et
al: Common gene variants in the tumor necrosis factor (TNF) and TNF
receptor superfamilies and NF-kB transcription factors and
non-Hodgkin lymphoma risk. PLoS One. 4:e53602009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xie X, Chang X, Chen L, Huang K, Huang J,
Wang S, Shen X, Liu P and Huang H: Berberine ameliorates
experimental diabetes-induced renal inflammation and fibronectin by
inhibiting the activation of RhoA/ROCK signaling. Mol Cell
Endocrinol. 381:56–65. 2013. View Article : Google Scholar : PubMed/NCBI
|