Introduction
Postmenopausal osteoporosis is characterized by a
decrease in bone mass and a deterioration in bone architecture. The
lifetime risk for women to have an osteoporotic fracture is 30–40%
worldwide (1), which has a notable
social, physical and economic impact (2–5).
Experimental animal models have contributed
tremendously to knowledge of the pathophysiology and treatment
targets of postmenopausal osteoporosis. The ovariectomy (OVX) rat
is the most commonly used animal model for evaluating the
mechanisms underlying postmenopausal osteoporosis and therapeutic
strategies for treating this disease (6–9).
However, compared with rats, mice may be a more effective model as
they have a more easily manipulated genome and lower drug doses are
required for treatment. Several strains of mice have been used in
postmenopausal osteoporosis research (10–13).
However, there are controversies regarding the use of C57BL/6J mice
as an animal model for postmenopausal osteoporosis (14–16). The
starting age of C57BL/6J mice subjected to ovariectomy varies from
4 to 30 weeks old (10,15–18).
Certain combinations of age, skeletal site and time post-surgery
may lead to varying levels of bone alterations in response to
estrogen deficiency in female rats (6). It is reasonable to hypothesize that the
starting age may result in varying levels of bone deterioration
upon estrogen deficiency in C57BL/6J mice. Therefore, to improve
the understanding of disease pathogenesis and to develop novel
therapies, further characterization of C57BL/6J mice as an animal
model for postmenopausal osteoporosis is required.
The current study investigated whether C57BL/6J mice
were a valid model for postmenopausal osteoporosis. Mice were used
in three different age groups (8, 12 and 16 weeks old) to evaluate
the extent of bone loss in response to OVX by micro-computed
tomography (µCT). To further evaluate bone mass changes of C57BL/6J
mice in response to OVX and estrogen replacement, the inbred strain
(BALB/c) and outbred strains (ICR and Kunming) of mice were used in
this study, and OVX was performed in those mice at 8 weeks of
age.
Materials and methods
Animals
Female C57BL/6J mice (8, 12 and 16 weeks old). The
total number of 8-, 12- and 16-week-old mice was 26, 21 and 22,
respectively. The range of body weight of 8-, 12- and 16-week-old
mice was 16.3–20.3, 15.9–24.7 and 20.7–25.6 g, respectively), and
female BALB/c, ICR and Kunming mice (all 8 weeks old). The total
number of BALB/c, ICR and Kunming was 24, 28 and 28, respectively.
The range of body weight of BALB/c, ICR and Kunming mice was
20.9–27.5, 26.5–32 and 32.4–41.8 g, respectively) were purchased
from Cavens Biological Technology Co., Ltd., Nanjing, China,
https://www.biomart.cn/56079/index.htm. All mice were
maintained under a 12-h light/dark cycle at room temperature
(22±2°C) with a humidity of 45% and allowed ad libitum
access to water and standard rodent chow. All animal procedures
were approved by the Animal Care and Use Committee of the Model
Animal Research Center of Nanjing University (Nanjing, China).
Ovariectomy and estrogen
supplement
Ovariectomy was conducted in 8-, 12- and 16-week-old
female C57BL/6J mice and 8-week-old female BALB/c, ICR and Kunming
mice. Mice were assigned to the following groups (Table I): Baseline, sacrificed at the
corresponding age; SHAM, were subjected to sham operation; OVX,
received bilateral ovariectomy; and OVX + 17β-estradiol (E2), at 1
week following surgery administered E2 (30 µg/kg; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany; cat no. E2758) subcutaneously five
times per week for 7 weeks. The dose of E2 used was similar to the
levels used in a previous study where 20 µg/kg/day resulted in a
serum E2 concentration that was similar to the levels in
SHAM-operated mice (19). C57BL/6J
mice were divided into 8-, 12- and 16-week-old groups
(OVX8, OVX12 and OVX16,
respectively) from the beginning of OVX. BALB/c, ICR and Kunming
mice were subject to surgery at the age of 8 weeks old
(OVX8). Surgery was conducted at the Model Animal
Research Center of Nanjing University by two operators (SZ and
GHW), who were skilled in OVX. In brief, animals were anesthetized
(100 mg/kg ketamine, purchased from Fujian Gutian Pharmaceutical
Co., Ltd., Fujian, China, http://www.fjgtyy.com/; and 5 mg/kg xylazine,
purchased from Jilin Huamu Animal Health Product Co., Ltd., Jilin,
China) for ~1 h. For the OVX group, anterior uterine horns were cut
to remove the ovaries; for the SHAM group dorsoventral incisions
were made through the skin, muscles and periosteum without removal
of the ovaries. At 8 weeks post-surgery, animals were sacrificed by
inhalation of CO2 and left femurs were harvested for µaT
analysis. The uteri were excised and weighed.
 | Table I.Number of mice in each group. |
Table I.
Number of mice in each group.
| Group | Baseline mice
(n) | SHAM mice (n) | OVX mice (n) | OVX + E2 mice
(n) |
|---|
| 8-week old
C57BL/6 | 6 | 6 | 7 | 7 |
| 12-week old
C57BL/6 | 8 | 6 | 7 | – |
| 16-week old
C57BL/6 | 8 | 6 | 8 | – |
| 8-week old
BALB/c | 6 | 6 | 6 | 6 |
| 8-week old ICR | 7 | 8 | 7 | 6 |
| 8-week old
Kunming | 7 | 7 | 7 | 7 |
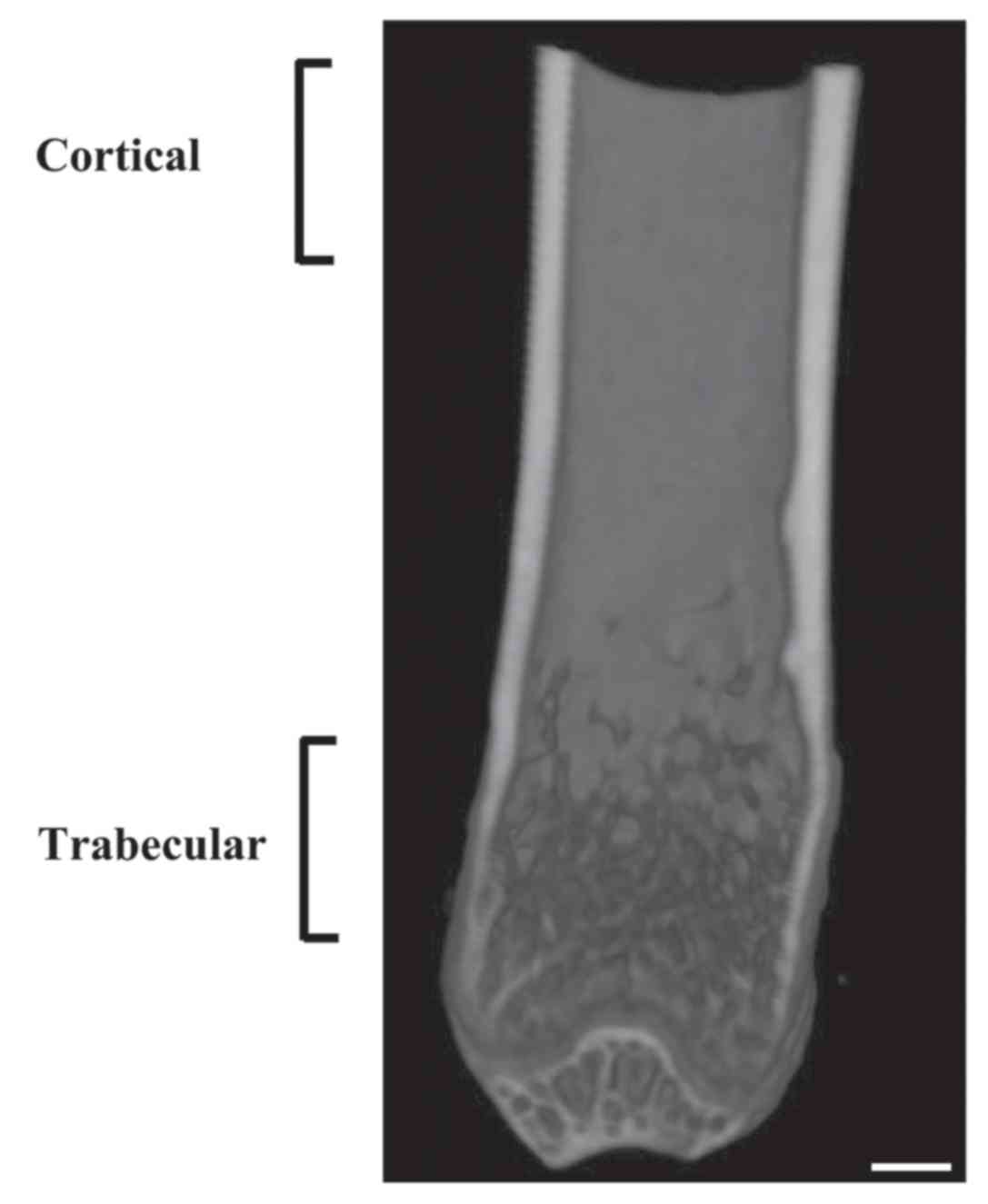
µCT analysis
Isolated left femurs were fixed with 4%
paraformaldehyde for 24 h at 4°C. Then, femurs were placed in
plastic tubes and stored within the animal bed inside the
SkyScan1176 (Bruker Corporation, Billerica, MA, USA). Femurs were
scanned using an 18 µ. resolution protocol: (45 kV, 556 µA, 0.1-mm
Cu filter, and 0.2° rotation step, 7-min scan). Volumetric
reconstruction software NRecon version 1.5 (Bruker Corporation) was
used to reconstruct CT images. Quantification of bone mineral
density and trabecular morphometric parameters was performed in a
hand-picked cancellous bone area within the primary ossification
center. The analyzed area was 0–2 mm above the distal growth plate
(Fig. 1). The analysis was performed
using scanner software (CTAn, Version 1.13, Bruker Corporation,
Billerica, MA, USA). Trabecular parameters were as follows:
Trabecular volumetric bone mineral density [vBMD; mg hydroxyapatite
(HA)/cm3], bone volume fraction (BV/TV; %), trabecular
thickness (Tb.Th; mm), trabecular number (Tb.N; 1/mm), trabecular
separation (Tb.Sp; mm), structure model index (SMI) and
connectivity density (Conn.D; 1/mm3). The present study
evaluated an area 1 mm above and below the midline of the femur to
measure the bone mineral density (cortical BMD; mg
HA/cm3) and cross-sectional thickness (Cs.Th; mm) of
cortical bone (Fig. 1).
Statistical analysis
The effect of age on the body weight, trabecular and
cortical bone properties in C57BL/6J were identified using one-way
analysis of variance and followed by Fisher's least significant
difference (LSD) if the variance was equal, otherwise Dunnett's
post hoc test was performed. To ascertain whether bone loss in
C57BL/6J following OVX was influenced by age, a two-way analysis of
variance followed by Bonferroni's post hoc test was used with
treatment (SHAM vs. OVX) and age. An unpaired Student's t-test was
used to compare body weight and bone morphology between SHAM and
OVX to detect OVX effects in three different age C57BL/6J
groups.
To compare the body mass and skeletal parameter
differences among the inbred and outbred mice strains, the values
of baseline were compared using a one-way analysis of variance
followed by an LSD post hoc test if the variance was equal,
otherwise Dunnett's post hoc test was used. To evaluate whether
bone loss following OVX or bone gain following subsequent E2
treatment was influenced by genetic factors, a two-way analysis of
variance followed by Bonferroni's post hoc test was used with
treatment (SHAM vs. OVX, or OVX vs. OVX+ E2) and strain. An
unpaired Student's t-test was used to compare body weight and bone
morphology between SHAM and OVX, or OVX and OVX+E2 to detect OVX or
estrogen supplement effects among inbred and outbred mice.
All statistical tests were performed using SPSS
version 17.0 software (SPSS, Inc., Chicago, IL, USA) and P<0.05
was considered to indicate a statistically significant difference.
Data are presented as the mean ± standard error.
Results
Age-associated alterations in body
weight, distal femur trabecular and femoral mid-shaft cortical bone
density and architecture in C57BL/6J female mice by µCT
Age-associated alterations of body weight and distal
femur metaphyseal trabecular architecture in C57BL/6J female mice
are presented in Table II. The body
weight of C57BL/6J mice increased steadily with age from 8–24 weeks
old by 43.9% (P<0.05). Trabecular vBMD plateaued from 8–16 weeks
and declined thereafter. Cancellous BV/TV decreased continuously
from 12–24 weeks of age. A similar trend was found in Tb.Th, Tb.N
and Conn.D (a measure of trabecular connectedness).
Correspondingly, Tb.Sp followed an opposite pattern of alterations.
SMI remained relatively constant from 8–16 weeks of age, but were
increased significantly at 20 (P<0.05 vs. 8, 12 and 16 weeks)
and 24 weeks (P<0.05 vs. 12 and 16 weeks), indicative of a shift
to a more rod-like architecture.
| Table II.Body weight, bone mass and
microarchitecture in 8-, 12-, 16-, 20- and 24-week old female
C57BL/6 mice. |
Table II.
Body weight, bone mass and
microarchitecture in 8-, 12-, 16-, 20- and 24-week old female
C57BL/6 mice.
| Parameter | 8-week old | 12-week old | 16-week old | 20-week old | 24-week old |
|---|
| Body weight
(g) |
17.49±0.30c,e |
20.55±1.00e |
23.26±0.59a | 22.92±1.19 |
25.16±0.902a,b |
| Distal femur:
Trabecular |
|
|
|
|
|
| vBMD
(mgHA/cm3) | 0.137±0.004 | 0.137±0.006 |
0.142±0.002e | 0.129±0.004 |
0.123±0.005c |
| BV/TV
(%) | 9.342±0.634 |
14.361±2.151e |
12.983±0.912e | 8.118±1.402 |
6.467±0.813b,c |
| Tb.Th
(mm) | 0.074±0.002 |
0.080±0.003d |
0.077±0.001d |
0.070±0.002b,c | 0.073±0.002 |
| Tb.N
(mm−1) |
1.253±0.074b,c |
1.826±0.202a,d,e |
1.681±0.099a,d,e |
1.137±0.182a–c |
0.875±0.096b,c |
| Tb.Sp
(mm) | 0.359±0.029 |
0.259±0.013e |
0.260±0.006e | 0.303±0.028 |
0.329±0.014b,c |
|
SMI |
2.559±0.033e |
2.547±0.088d,e |
2.537±0.057d,e |
2.751±0.076b,c |
2.779±0.067a–c |
| Conn.D
(mm−3) |
41.546±4.465e |
69.722±1.015e |
55.104±0.508e | 41.531±0.781 |
19.872±2.994a–c |
| Femoral midshaft:
Cortical |
|
|
|
|
|
| BMD
(mgHA/cm3) |
0.848±0.027b–e |
1.064±0.008a,c–e |
1.107±0.008a,b,d |
1.163±0.009a–c |
1.136±0.009a,b |
| Cs.Th
(mm) |
0.138±0.002b–e |
0.170±0.003a,c–e |
0.183±0.004a,b |
0.182±0.006a,b |
0.190±0.003a,b |
Age-related changes in the mid-femur diaphysis are
also presented in Table II.
Cortical BMD and Cs.Th increased continuously with age; the largest
increase occurred between 8 and 12 weeks, and measurements
plateaued thereafter.
Influence of age on the sensitivity to
estrogen deprivation in C57BL/6J female mice
When mice were sacrificed, the abdominal cavity was
explored. The ovaries could not be located and the uteruses were
very small, which indicated that OVX mice had their ovaries
removed. As expected, OVX8, OVX12 and
OVX16 were also heavier than mice in the SHAM group,
although there was not a significant difference between
OVX16 and SHAM groups (P<0.01, P<0.05 and P=0.057,
respectively; Table III).
| Table III.Effect of estrogen deprivation on
body weight, bone mass and microarchitecture in OVX8,
OVX12 and OVX16 female C57BL/6J mice. |
Table III.
Effect of estrogen deprivation on
body weight, bone mass and microarchitecture in OVX8,
OVX12 and OVX16 female C57BL/6J mice.
|
| 8-week-old | 12-week-old | 16-week-old |
|
|---|
|
|
|
|
|
|
|---|
| Parameter | SHAM | OVX | Difference between
SHAM and OVX (%) | SHAM | OVX | Difference between
SHAM and OVX (%) | SHAM | OVX | Difference between
SHAM and OVX (%) |
P-valuetreatment*age |
|---|
| Body weight
(g) | 22.00±0.80 | 26.81±0.46 | 21.9b | 22.92±1.19 | 27.51±1.34 | 20.0a | 25.16±0.90 | 28.10±1.05 | 11.7 | NS |
| Distal femur:
trabecular |
|
|
|
|
|
|
|
|
|
|
| vBMD
(mgHA/cm3) | 0.143±0.006 | 0.092±0.003 | −35.1c | 0.129±0.004 | 0.100±0.005 | −22.6b | 0.123±0.005 | 0.104±0.005 | −15.8a | 0.016 |
| BV/TV
(%) | 8.005±1.513 | 2.862±0.318 | −64.2a | 8.118±1.402 | 4.292±1.068 | −47.1a | 6.467±1.154 | 4.452±1.068 | −31.2 | NS |
| Tb.Th
(mm) | 0.069±0.003 | 0.068±0.002 | −1.4 | 0.070±0.002 | 0.072±0.005 | 2.9 | 0.073±0.002 | 0.075±0.004 | 2.7 | NS |
| Tb.N
(mm−1) | 1.119±0.160 | 0.415±0.039 | −62.9a | 1.137±0.182 | 0.568±0.110 | −50.0a | 0.875±0.096 | 0.571±0.121 | −34.7 | NS |
| Tb.Sp
(mm) | 0.307±0.013 | 0.630±0.025 | 100.0c | 0.303±0.028 | 0.416±0.015 | 37.3b | 0.329±0.014 | 0.402±0.024 | 22.2a | <0.001 |
|
SMI | 2.684±0.071 | 2.725±0.060 | 1.5 | 2.751±0.076 | 2.956±0.153 | 7.5 | 2.779±0.067 | 3.005±0.084 | 8.1 | NS |
| Conn.D
(mm−3) | 33.369±0.418 | 12.722±1.129 | −61.9c | 41.531±0.781 | 18.015±2.523 | −56.6a | 19.872±2.994 | 16.511±3.189 | −16.9 | 0.035 |
| Femoral midshaft:
Cortical |
|
|
|
|
|
|
|
|
|
|
| BMD
(mgHA/cm3) | 1.136±0.004 | 1.132±0.008 | −0.4 | 1.163±0.009 | 1.123±0.006 | −3.5b | 1.136±0.009 | 1.102±0.008 | −2.9a | NS |
| Cs.Th
(mm) | 0.177±0.003 | 0.165±0.002 | −6.8b | 0.182±0.006 | 0.179±0.003 | −1.6 | 0.190±0.003 | 0.184±0.005 | −3.2 | NS |
The patterns of microarchitectural deterioration in
response to OVX varied among the three age groups (Table III). In general, microarchitectural
deterioration was severe in OVX8, intermediate in
OVX12 and lowest in OVX16. This was
highlighted by significant differences between OVX and SHAM across
the parameters examined in OVX8, with the exception of
Tb.Th and SMI. For trabecular vBMD, Tb.Sp and Conn.D in the distal
femur, the response to OVX depended on the age, as determined by
two-way analysis of variance analyses between treatment and age
(Ptreatment*age=0.016,
Ptreatment*age<0.001, P
treatment*age=0.035, respectively).
Upon ovariectomy, trabecular vBMD in all OVX groups
was significantly reduced relative to their SHAM counterparts with
OVX8 demonstrating the greatest reduction (−35.1%).
BV/TV declined significantly in OVX8 (−64.2%, P=0.0078
vs. SHAM) and OVX12 (−47.1%, P=0.0496 vs. SHAM) and
declined non-significantly in OVX16 (−31.2%, P=0.1834
vs. SHAM). For Tb.Th, there were no marked alterations in all age
groups. All age groups exhibited a decline in Tb.N, ranging from
−34.7 to −62.9%, but this decline was only statistically
significant in OVX8 (−62.9%, P=0.0017 vs. SHAM) and
OVX12 (−37.3%, P=0.0184 vs. SHAM). No significant
alterations in SMI existed among the three groups. Conn.D decreased
significantly in OVX8 (−61.9%, P=0.0008 vs. SHAM) and
OVX12 (−56.6%, P=0.0108 vs. SHAM), but OVX16
exhibited a non-significant decrease.
The effect of OVX on mid-femoral cortical bone did
not appear to depend on age (Table
III). Cortical BMD exhibited a statistically significant
decline in OVX12 (−3.5%, P=0.028 vs. SHAM) and
OVX16 (−2.9%, P=0.02 vs. SHAM), except for
OVX8. By contrast, Cs.Th only declined significantly in
OVX8 (−6.8%, P=0.0114).
Differences in body mass and bone
morphology among inbred and outbred strains of mice
To further explore the bone mass alterations in
C57BL/6J mice in response to OVX and estrogen replacement, the
above studies were extended by evaluating the inbred and outbred
strains, and using an estrogen supplement.
At 8 weeks (baseline), body weight was highest in
Kunming, intermediate in ICR and BALB/C, and lowest in C57BL/6J
mice (Table IV). Femur trabecular
bone in C57BL/6J mice was characterized by the lowest trabecular
vBMD, BV/TV, Tb.Th, Tb.N and Conn.D, whereas these mice also
exhibited the highest Tb.Sp and SMI. Kunming mice had the most
trabeculae and consequently the least Tb.Sp. In trabecular regions,
Tb.Th was significantly higher in BALB/c compared with ICR mice
(P<0.05).
| Table IV.Body weight, bone mass and
microarchitecture in four inbred and outbred strains. |
Table IV.
Body weight, bone mass and
microarchitecture in four inbred and outbred strains.
| Parameter | C57BL/6J (n=6) | BALB/c (n=6) | ICR (n=7) | Kunming (n=6) |
|---|
| Body weight
(g) |
17.49±0.30b–d |
23.16±0.85a |
29.20±0.43a |
36.29±0.82a |
| Distal femur:
Trabecular |
|
|
|
|
| vBMD
(mgHA/cm3) |
0.137±0.004b–d |
0.239±0.012a |
0.230±0.008a |
0.267±0.018a |
| BV/TV
(%) |
9.342±0.634b–d |
26.236±1.920a |
20.161±1.844a |
29.749±3.233a |
| Tb.Th
(mm) |
0.074±0.002b–d |
0.104±0.003a,c |
0.091±0.003a,b,d |
0.102±0.003a,c |
| Tb.N
(mm−1) |
1.253±0.074b–d |
2.535±0.175a |
2.182±0.144a |
2.881±0.236a |
| Tb.Sp
(mm) |
0.359±0.286b,d |
0.270±0.017a |
0.301±0.025d |
0.218±0.014a,c |
|
SMI |
2.559±0.033b–d |
1.646±0.108a |
1.984±0.063a |
1.725±0.133a |
| Conn.D
(mm−3) |
41.546±4.465b–d |
81.648±5.149a |
73.165±4.898a,d |
92.052±9.051a,c |
| Femoral midshaft:
cortical |
|
|
|
|
| BMD
(mgHA/cm3) |
0.848±0.027b–d |
1.119±0.011a |
1.151±0.006a |
1.129±0.009a |
| Cs.Th
(mm) |
0.138±0.002b–d |
0.182±0.005a,c,d |
0.215±0.006a,b |
0.210±0.009a,b |
In the femoral cortical bone, BMD was significantly
lower in C57BL/6J than the other three strains of mice, whereas
Cs.Th was highest in ICR and Kunming, intermediate in BALB/c and
lowest in C57BL/6J (Table IV).
Influence of strain on the sensitivity
to estrogen deprivation and supplement
C57BL/6J and BALB/c mice subjected to OVX exhibited
significant increases 21.9% (P=0.0001) and 6.5% (P=0.0353) of their
body mass compared with SHAM counterparts (Tables III and V). Only C57BL/6J and Kunming mice treated
with estrogen exhibited significantly decreased body masses (−8.3%,
P=0.0066; −4.8%, P=0.0465; respectively; Table VI).
| Table V.Effect of estrogen deprivation on
body weight, bone mass and microarchitecture in three inbred and
outbred mice. |
Table V.
Effect of estrogen deprivation on
body weight, bone mass and microarchitecture in three inbred and
outbred mice.
|
| BALB/c | ICR | Kunming |
|
|---|
|
|
|
|
|
|
|---|
| Parameter | SHAM | OVX | Difference between
SHAM and OVX (%) | SHAM | OVX | Difference between
SHAM and OVX (%) | SHAM | OVX | Difference between
SHAM and OVX (%) |
P-valuetreatment*age |
|---|
| Body weight
(g) | 24.30±0.31 | 25.88±0.66 | 6.5a | 36.65±0.59 | 33.46±0.91 | −0.6 | 44.80±1.55 | 48.09±1.93 | 7.5 | NS |
| Distal femur:
trabecular |
|
|
|
|
|
|
|
|
|
|
| vBMD
(mg HA/cm3) | 0.276±0.006 | 0.198±0.010 | −28.4c,d | 0.231±0.016 | 0.159±0.014 | −31.2b,e | 0.234±0.020 | 0.183±0.010 | −22.0a,d | NS |
| BV/TV
(%) | 21.590±0.561 | 9.740±1.842 | −54.9c | 19.827±2.447 | 11.052±1.858 | −44.3b | 21.719±3.144 | 12.025±1.358 | −44.6 | NS |
| Tb.Th
(mm) | 0.091±0.001 | 0.077±0.002 | −15.4c | 0.097±0.005 | 0.082±0.004 | −15.7a | 0.104±0.002 | 0.091±0.002 | −12.7c | NS |
| Tb.N
(mm−1) | 2.384±0.053 | 1.249±0.211 | −47.6c | 2.013±0.165 | 1.313±0.185 | −34.8a | 2.067±0.279 | 1.322±0.139 | −36.0a | NS |
| Tb.Sp
(mm) | 0.262±0.004 | 0.438±0.043 | 67.2a | 0.300±0.025 | 0.417±0.045 | 39.0a | 0.288±0.025 | 0.423±0.033 | 47.2a | 0.007 |
|
SMI | 1.798±0.043 | 2.274±0.070 | 26.5c | 1.997±0.087 | 2.299±0.085 | 15.1a | 2.013±0.184 | 2.253±0.081 | 11.9 | NS |
| Conn.D
(mm−3) | 61.169±1.637 | 40.762±7.984 | −33.4 | 62.382±0.657 | 42.713±5.481 | −31.5a | 45.978±0.814 | 29.597±0.430 | −35.6 | 0.035 |
| Femoral midshaft:
Cortical |
|
|
|
|
|
|
|
|
|
|
| BMD (mg
HA/cm3) | 1.218±0.009 | 1.213±0.008 | −0.4 | 1.258±0.008 | 1.253±0.008 | −0.4 | 1.209±0.007 | 1.220±0.009 | 0.9 | NS |
| Cs.Th
(mm) | 0.222±0.002 | 0.209±0.007 | −5.9 | 0.255±0.006 | 0.255±0.007 | 0.4 | 0.212±0.004 | 0.207±0.011 | −2.6 | NS |
| Table VI.Effect of estrogen replacement on
body weight, bone mass and microarchitecture in four inbred and
outbred mice. |
Table VI.
Effect of estrogen replacement on
body weight, bone mass and microarchitecture in four inbred and
outbred mice.
|
| C57BL/6J | BALB/c | ICR | Kunming |
|
|---|
|
|
|
|
|
|
|
|---|
| Parameter | OVX+E2 | Difference between
OVX and OVX+E2 (%) | OVX+E2 | Difference between
OVX and OVX+E2 (%) | OVX+E2 | Difference between
OVX and OVX+E2 (%) | OVX+E2 | Difference between
OVX and OVX+E2 (%) |
P-valuetreatment*strain |
|---|
| Body weight
(g) | 24.60±0.491 | −8.3a | 25.59±0.42 | −1.1 | 31.167±0.83 | −6.8 | 43.257±1.008 | −4.8a | NS |
| Distal femur:
trabecular |
|
|
|
|
|
|
|
|
|
| vBMD
(mgHA/cm3) | 0.118±0.005 | 26.8b | 0.218±0.009 | 10.2 | 0.179±0.009 | 12.5 | 0.176±0.013 | −3.4 | NS |
| BV/TV
(%) | 4.394±0.451 | 53.5 | 11.570±1.009 | 18.8 | 12.421±1.772 | 12.4 | 11.433±1.284 | −4.9 | NS |
| Tb.Th
(mm) | 0.078±0.003 | 14.2a | 0.087±0.002 | 13.7b | 0.088±0.002 | 7.2 | 0.104±0.002 | 14.9c | NS |
| Tb.N
(mm−1) | 0.559±0.049 | 34.6a | 1.323±0.102 | 5.9 | 1.420±0.198 | 8.2 | 1.096±0.115 | −17.1 | NS |
| Tb.Sp
(mm) | 0.429±0.015 | −31.9c | 0.360±0.015 | −17.7d | 0.357±0.031 | −14.4d | 0.457±0.054 | 8.0e | 0.02 |
|
SMI | 2.947±0.036 | 8.1a | 2.350±0.074 | 3.3 | 2.358±0.083 | 2.6 | 2.481±0.034 | 10.2 | NS |
| Conn.D
(mm−3) | 15.932±2.532 | 25.2 | 30.063±3.984 | −33.4 | 44.729±7.660 | 4.7 | 21.349±3.498 | −27.9 | NS |
| Femoral midshaft:
Cortical |
|
|
|
|
|
|
|
|
|
| BMD
(mgHA/cm3) | 1.154±0.005 | 2.0a | 1.224±0.009 | 0.8 | 1.262±0.007 | 0.7 | 1.229±0.008 | 0.8 | NS |
| Cs.Th
(mm) | 0.181±0.002 | 9.5c | 0.217±0.002 | 4.0 | 0.266±0.007 | 4.1 | 0.213±0.002 | 3.4 | NS |
All four strains of mice lost cancellous bone in the
femur following OVX, with no treatment-strain interaction observed,
except in Tb.Sp and Conn.D. (Ptreatment*age =0.007;
Ptreatment*age =0.035, respectively; Tables III and V). Femur trabecular vBMD was lower in OVX
than the SHAM group in all four strains of mice (Tables III and V). The effect of OVX on trabecular vBMD in
C57BL/6J (−35.1%, P<0.001 vs. SHAM) differed significantly from
the effect in BALB/c (−28.4%, P<0.001 vs. SHAM; P<0.001 vs.
C57BL/6J), ICR (−31.2%, P=0.002 vs. SHAM; P=0.001 vs. C57BL/6J) and
Kunming (−22.0%, P=0.024 vs. SHAM; P<0.001 vs. C57BL/6J).
Similar to femur trabecular vBMD, C57BL/6J exhibited the greatest
decline in BV/TV (−64.2%, P=0.001 vs. SHAM; Tables III and V). The other strains also exhibited a
decline in femur BV/TV, ranging from −44.3 to −54.9%, but this
decline was statistically significant only in BALB/c (P<0.001
vs. SHAM) and ICR (P=0.008 vs. SHAM). A significant difference in
Tb.Th between SHAM and OVX group was not detected in the C57BL/6J
strain, whereas BALB/c exhibited a −15.4% decrease (P<0.001 vs.
SHAM), ICR −15.7% (P=0.016 vs. SHAM) and Kunming −12.7% (P<0.001
vs. SHAM). Tb.N decreased in C57BL/6J (−62.9%, P=0.016 vs. SHAM),
BALB/c (−47.6%, P<0.001 vs. SHAM), ICR (−34.8%, P=0.011 vs.
SHAM) and Kunming (−36.0%, P=0.013 vs. SHAM). Tb.Sp increased in
C57BL/6J (100%, P<0.001 vs. SHAM), BALB/c (67.2%, P=0.025 vs.
SHAM), ICR (39.0%, P=0.023 vs. SHAM) and Kunming (47.2%, P=0.026
vs. SHAM). SMI increased significantly in BALB/c (26.5%, P<0.001
vs. SHAM) and ICR (15.1%, P=0.020 vs. SHAM) whereas Conn.D declined
in C57BL/6J (−61.9%, P<0.001 vs. SHAM) and ICR (−31.5%, P=0.042
vs. SHAM). With regard to cortical bone indices, significant
alterations were only found in C57BL/6J (Tables III and V). The difference in Cs.Th between SHAM and
OVX mice was −6.8% in C57BL/6J mice (P=0.002; Tables III and V).
For Tb.Sp, the response to estrogen supplement was
dependent on the strain (Ptreatment*strain=0.02;
Table VI). The alteration in this
index following estrogen supplement differed significantly between
C57BL/6J and BALB/c (P=0.002), ICR (P=0.001) and Kunming (P=0.027).
Notably, C57BL/6J mice were the only strain markedly responsive to
estrogen supplement, demonstrating significant alterations in all
parameters except for BV/TV and Conn.D. Whereas estrogen decreased
Tb.Sp by 31.9% in the metaphyseal trabecular bone of C57BL/6J mice
(P<0.001 vs. OVX), it increased trabecular vBMD by 26.8%
(P=0.003 vs. OVX; Table VI), BV/TV
by 53.5% (P=0.059 vs. OVX; Table
VI), Tb.Th by 14.2% (P=0.045 vs. OVX) and SMI by 8.1% (P=0.014
vs. OVX). In the mid-femoral cortical bone of C57BL/6J mice,
estrogen increased cortical BMD by 2.0% (P=0.023 vs. OVX; Table VI) and Cs.Th by 9.5% (P<0.001 vs.
OVX). In contrast with C57BL/6J, estrogen treatment failed to
significantly alter any above indices in BALB/c and Kunming mice
except Tb.Th (13.7%, P=0.001 vs. OVX; 14.9%, P<0.001;
respectively). ICR was the least sensitive to estrogen treatment
showing no significant alterations across all the parameters
examined.
Discussion
OVX model mice have been widely used in the study of
postmenopausal osteoporosis. Multiple lines of evidence have
revealed that genetic factors serve a profound role in regulating
the balance of bone metabolism in mice (14,20,21). In
the present study, it was demonstrated that the peak of cancellous
mass in C57BL/6J female mice was present at 8 weeks old. Mice
subjected to OVX at 8 weeks old had marked cancellous bone loss,
and OVX-induced cancellous bone loss was restored by estrogen
treatment. The data also suggested that OVX-induced loss of
cancellous bone may gradually decline with age in C57BL/6J female
mice.
The BV/TV at distal femur in C57BL/6J mice has been
shown to be greatest at 6-weeks old and sequentially decline with
age (22). The present study
demonstrated an age-associated reduction in cancellous bone in
C57BL/6J female mice, as trabecular vBMD and BV/TV were greatest at
8 weeks old. Notably, it was revealed that OVX-induced loss of
cancellous bone was associated with age, and the data indicated
that the rate of trabecular bone loss in mice subjected to OVX at 8
weeks old was higher than that at 12 weeks old. In addition, there
were no significant differences in cancellous bone alterations in
mice subjected to OVX at 16 weeks old when compared with the SHAM
group. These findings indicated that the effect of estrogen on
regulating turnover of cancellous bone in C57BL/6J female mice may
gradually decline with age. Further studies are necessary to
explore cellular mechanisms underlying the reduced response to
estrogen with age.
In contrast with cancellous bone, the results showed
that an age-associated increase of cortical bone persisted
throughout 8–16 weeks of age in C57BL/6J female mice. However, only
a slight reduction in cortical bone, in an age-independent manner,
was observed in those mice in response to OVX, suggesting that
cortical bone was not sensitive to estrogen deprivation in C57BL/6J
female mice.
C57BL/6J mice are typically used to produce gene
knockout and transgenic animals. A number of previous studies have
identified bone loss when the ovariectomy timing was 2 months old
in this strain (23,24). However, C57BL/6J mice have been
questioned as an ideal model for the study of postmenopausal
osteoporosis as they have been found to be resistant to OVX-induced
loss of bone. Bouxsein et al (14) previously discovered that 4-month-old
C57BL/6J mice were resistant to OVX-induced trabecular bone
deterioration in the proximal tibia. Iwaniec et al (15) demonstrated that no significant
difference was observed between C57BL/6J OVX and SHAM groups in the
distal femur cancellous BV/TV 3 months post-surgery. Klinck and
Boyd (16) demonstrated that the
majority of morphology parameters in the C57BL/6J femur and tibia
were not statistically significant at 5 weeks post-ovariectomy.
This data from previous research were obtained from mice subjected
to OVX at the age of 12 or 16 weeks, which is consistent with the
findings of the present study. The present study also revealed that
alterations of cancellous bone in C57BL/6J mice at the age of 8
weeks were greatest.
Previous studies have demonstrated that the skeletal
response to OVX varied among inbred mice (14–16).
This study extended the above by adding one inbred (BALB/c) and two
outbred strains (ICR and Kunming), and all strains of mice were
subjected to OVX at 8 weeks old. It was demonstrated that C57BL/6J
mice were most sensitive to OVX-induced loss of bone among the four
strains, exhibiting greater alterations in body weight, trabecular
vBMD, BV/TV, Tb.N, Conn.D and Cs.Th. In contrast with previous
results (14–16), the present study revealed that
C57BL/6J mice were the most sensitive to estrogen deficiency among
the four strains. It should be noted that Cs.Th, reflecting the
cortical bone, decreased only in C57BL/6J mice. Contradicting
results may be due to the age of the animals at OVX (previous
studies using no less than 3-month-old C57BL/6J mice) and the
duration of time elapsed following OVX (8 weeks). The various
skeletal alterations during the 2–4 months old period caused by
differing post-pubertal architectural development patterns among
different strains may account for discrepancies arising from the
different starting ages (25). It
would be interesting to determine the underlying cellular
mechanisms that determine the different strain-associated responses
to estrogen deficiency by analysis of histomorphometry or
biochemical markers.
E2 has been documented to improve bone quantity and
miscrostructure both in mice and rats following OVX (19,26).
With respect to humans, estrogen therapy prevents postmenopausal
bone loss (27). In the present
study, it was revealed that skeletal responses to estrogen were
strain-dependent where C57BL/6J improved most in regard to
trabecular vBMD, BV/TV, Tb.Th, cortical BMD and Cs.Th. As for
Conn.D, estrogen failed to significantly increase this as this
index was thought to be irreversible when bone loss has occurred
(28), as it was unclear whether the
anabolic treatment parathyroid hormone restored any connectivity
(29,30).
The findings of the present study raise notable
issues for selecting an animal model to study the mechanism of
certain genes in postmenopausal osteopenia and evaluate the
effectiveness of a novel candidate agent for treatment. Following
OVX at 4 months old, C57BL/6J prostaglandin E2 receptor (one of the
four prostanoid receptors) knockout mice exhibited protection
against bone loss in femur and L4 vertebrae (31). However, when OVX was performed on
30-week-old female Col.1-PPARγ mice, ovariectomy-induced bone loss
was accelerated (10). As the
present results revealed that the factor age of C57BL/6J influenced
the sensitivity of bone to OVX, the conclusions of a specific gene
on osteoporosis must be identified with care. In addition, BALB/c,
ICR and Kunming mice have been exploited in searching for
anti-osteoporotic medicine, and drug testing results may be
negative as the present study revealed that C57BL/6J was the most
sensitive to the anti-osteoporotic drug estrogen.
There are certain limitations associated with this
study. First, only one bone (femur) was assessed to evaluate the
extent of OVX-induced bone loss and response to estrogen. Second,
only a single time point (8 weeks) after OVX was assessed and
therefore the rate of bone loss was not assessed. Third, no
histomorphometry or biochemical markers were performed, and thus it
is impossible to observe the underlying cellular mechanisms that
determine the different strain-associated responses to estrogen
deficiency. In spite of these shortcomings, conclusions were made
by ex vivo analysis of femoral bone microarchitecture by
µCT, as the bone (femur), the postsurgery time (8 weeks) and method
(µCT) were the most commonly used in previous literature.
In conclusion, the results revealed that OVX-induced
bone loss may be age- and strain-specific, which emphasizes the
importance of appropriate selection of mouse strains and their age
in postmenopausal osteoporosis research. It was demonstrated that
C57BL/6J female mice subjected to OVX at 8 weeks old resulted in
pronounced loss of bone that was sensitive to estrogen. Therefore,
the present data may give novel insights into the age- and
strain-associated effect of OVX in regulating turnover of bone in
female mice, and this must be considered when using C57BL/6J mice
as an animal model of postmenopausal osteoporosis.
Acknowledgements
The authors would like to thank Jason Kang for
editing the manuscript. The present study was supported by the
National Science Foundation for Distinguished Young Scholars of
China (grant no. 81125013) and National Natural Science Foundation
of China (grant no. 81472116).
References
|
1
|
Jagtap VR, Ganu JV and Nagane NS: BMD and
serum intact osteocalcin in postmenopausal osteoporosis women.
Indian J Clin Biochem. 26:70–73. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Johnell O and Kanis JA: An estimate of the
worldwide prevalence and disability associated with osteoporotic
fractures. Osteoporos Int. 17:1726–1733. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cauley JA, Thompson DE, Ensrud KC, Scott
JC and Black D: Risk of mortality following clinical fractures.
Osteoporos Int. 11:556–561. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Frost SA, Nguyen ND, Center JR, Eisman JA
and Nguyen TV: Excess mortality attributable to hip-fracture: A
relative survival analysis. Bone. 56:23–29. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reginster JY and Burlet N: Osteoporosis: A
still increasing prevalence. Bone. 38 2 Suppl 1:S4–S9. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Francisco JI, Yu Y, Oliver RA and Walsh
WR: Relationship between age, skeletal site and time
post-ovariectomy on bone mineral and trabecular microarchitecture
in rats. J Orthop Res. 29:189–196. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu XL, Li CL, Lu WW, Cai WX and Zheng LW:
Skeletal site-specific response to ovariectomy in a rat model:
Change in bone density and microarchitecture. Clin Oral Implants
Res. 26:392–398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Esteves CM, Moraes RM, Gomes FC, Marcondes
MS, Lima GM and Anbinder AL: Ovariectomy-associated changes in
interradicular septum and in tibia metaphysis in different
observation periods in rats. Pathol Res Pract. 211:125–129. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Anderson JJ, Ambrose WW and Garner SC:
Biphasic effects of genistein on bone tissue in the ovariectomized,
lactating rat model. Proc Soc Exp Biol Med. 217:pp. 345–350. 1998;
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cho SW, Yang JY, Her SJ, Choi HJ, Jung JY,
Sun HJ, An JH, Cho HY, Kim SW, Park KS, et al: Osteoblast-targeted
overexpression of PPARγ inhibited bone mass gain in male mice and
accelerated ovariectomy-induced bone loss in female mice. J Bone
Miner Res. 26:1939–1952. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hsu YH, Chen WY, Chan CH, Wu CH, Sun ZJ
and Chang MS: Anti-IL-20 monoclonal antibody inhibits the
differentiation of osteoclasts and protects against osteoporotic
bone loss. J Exp Med. 208:1849–1861. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jun AY, Kim HJ, Park KK, Son KH, Lee DH,
Woo MH, Kim YS, Lee SK and Chung WY: Extract of Magnoliae Flos
inhibits ovariectomy-induced osteoporosis by blocking
osteoclastogenesis and reducing osteoclast-mediated bone
resorption. Fitoterapia. 83:1523–1531. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Y, Wei L, Miron RJ, Shi B and Bian
Z: Anabolic bone formation via a site-specific bone-targeting
delivery system by interfering with semaphorin 4d expression. J
Bone Miner Res. 30:286–296. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bouxsein ML, Myers KS, Shultz KL, Donahue
LR, Rosen CJ and Beamer WG: Ovariectomy-induced bone loss varies
among inbred strains of mice. J Bone Miner Res. 20:1085–1092. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Iwaniec UT, Yuan D, Power RA and Wronski
TJ: Strain-dependent variations in the response of cancellous bone
to ovariectomy in mice. J Bone Miner Res. 21:1068–1074. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Klinck J and Boyd SK: The magnitude and
rate of bone loss in ovariectomized mice differs among inbred
strains as determined by longitudinal in vivo micro-computed
tomography. Calcif Tissue Int. 83:70–79. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sabsovich I, Clark JD, Liao G, Peltz G,
Lindsey DP, Jacobs CR, Yao W, Guo TZ and Kingery WS: Bone
microstructure and its associated genetic variability in 12 inbred
mouse strains: microCT study and in silico genome scan. Bone.
42:439–451. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kara FM, Doty SB, Boskey A, Goldring S,
Zaidi M, Fredholm BB and Cronstein BN: Adenosine A(1) receptors
regulate bone resorption in mice: Adenosine A(1) receptor blockade
or deletion increases bone density and prevents ovariectomy-induced
bone loss in adenosine A(1) receptor-knockout mice. Arthritis Rheu.
62:534–541. 2010. View Article : Google Scholar
|
|
19
|
Modder UI, Riggs BL, Spelsberg TC, Fraser
DG, Atkinson EJ, Arnold R and Khosla S: Dose-response of estrogen
on bone versus the uterus in ovariectomized mice. Eur J Endocrinol.
151:503–510. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Beamer WG, Donahue LR, Rosen CJ and
Baylink DJ: Genetic variability in adult bone density among inbred
strains of mice. Bone. 18:397–403. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li CY, Schaffler MB, Wolde-Semait HT,
Hernandez CJ and Jepsen KJ: Genetic background influences cortical
bone response to ovariectomy. J Bone Miner Res. 20:2150–2158. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Glatt V, Canalis E, Stadmeyer L and
Bouxsein ML: Age-related changes in trabecular architecture differ
in female and male C57BL/6J mice. J Bone Miner Res. 22:1197–1207.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Duque G, Huang DC, Dion N, Macoritto M,
Rivas D, Li W, Yang XF, Li J, Lian J, Marino FT, et al:
Interferon-γ plays a role in bone formation in vivo and rescues
osteoporosis in ovariectomized mice. J Bone Miner Res.
26:1472–1483. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang F, Wang PX, Wu XL, Dang SY, Chen Y,
Ni YY, Gao LH, Lu SY, Kuang Y, Huang L, et al: Deficiency of
adiponectin protects against ovariectomy-induced osteoporosis in
mice. PLoS One. 8:e684972013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Buie HR, Moore CP and Boyd SK:
Postpubertal architectural developmental patterns differ between
the L3 vertebra and proximal tibia in three inbred strains of mice.
J Bone Miner Res. 23:2048–2059. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thompson DD, Simmons HA, Pirie CM and Ke
HZ: FDA Guidelines and animal models for osteoporosis. Bone.
17:125S–133S. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bansal N, Katz R, de Boer IH, Kestenbaum
B, Siscovick DS, Hoofnagle AN, Tracy R, Laughlin GA, Criqui MH,
Budoff MJ, et al: Influence of estrogen therapy on calcium,
phosphorus and other regulatory hormones in postmenopausal women:
The MESA study. J Clin Endocrinol Metab. 98:4890–4898. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Campbell GM, Buie HR and Boyd SK: Signs of
irreversible architectural changes occur early in the development
of experimental osteoporosis as assessed by in vivo micro-CT.
Osteoporos Int. 19:1409–1419. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sato M, Zeng GQ and Turner CH:
Biosynthetic human parathyroid hormone (1–34) effects on bone
quality in aged ovariectomized rats. Endocrinology. 138:4330–4337.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sato M, Westmore M, Ma YL, Schmidt A, Zeng
QQ, Glass EV, Vahle J, Brommage R, Jerome CP and Turner CH:
Teriparatide [PTH(1–34)] strengthens the proximal femur of
ovariectomized nonhuman primates despite increasing porosity. J
Bone Miner Res. 19:623–629. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang M, Feigenson M, Sheu TJ, Awad HA,
Schwarz EM, Jonason JH, Loiselle AE and O'Keefe RJ: Loss of the
PGE2 receptor EP1 enhances bone acquisition, which protects against
age and ovariectomy-induced impairments in bone strength. Bone.
72:92–100. 2015. View Article : Google Scholar : PubMed/NCBI
|