Introduction
Liver cancer, also known as hepatic cancer, is a
common malignant tumor that represents a severe threat to human
health. According to the latest epidemiological study from the
American Cancer Society, liver cancer is the fifth leading cause of
cancer-associated mortality in men and the eighth leading cause in
women. Furthermore, the death rate due to liver cancer is
increasing (1). In China, liver
cancer has the second highest mortality rate of all cancers in men
and the fourth highest in women. Between 2000 and 2011, an
increasing trend in the incidence and death rates of liver cancers
has been observed (2). Therefore, it
is an urgent issue to investigate the underlying molecular
mechanisms to develop novel drugs and identify more sensitive tumor
markers for liver cancer. However, knowledge of the molecular
mechanisms associated with liver cancer remains insufficient and
further elucidation thereof may improve the early diagnosis and
prediction of liver cancer patient prognosis as an urgent issue.
Numerous studies have assessed tumor markers of liver cancer. At
present, α-fetoprotein (AFP) is the most widely used tumor marker
for the early detection of liver cancer (3). Besides AFP, AFP-L3 and squamous cell
carcinoma antigen are used for the early diagnosis of liver cancer
and heat shock protein 70, TGF-β and microRNA-500/29/112 are used
as prognostic markers for liver cancer (4). However, the specificity and sensitivity
of these tumor markers are not satisfactory. Therefore, the present
study assessed whether GDF11 may be a novel tumor marker in liver
cancer.
Growth differentiation factor 11 (GDF11), also known
as bone morphogenetic protein 11 (BMP11), was first cloned and
characterized as a member of the BMP/transforming growth factor β
(TGF-β) superfamily (5). The GDF11
gene was mapped to human chromosome 12q13.2 by aligning the GDF11
sequence (GenBank ID AF100907) with the genomic sequence (GRCh38).
It encodes a 407-amino acid protein with a signaling sequence for
secretion, an RXXR proteolytic processing site and a region at the
C-terminal end containing a highly conserved pattern of cysteine
residues (6). GDF11 protein, cleaved
by pro-protein convertase subtilisin/kexin type 5 (PCSK5), forms a
non-covalent latent complex, which contains an inhibitory
pro-domain at the N-terminus and two disulfide-linked active
domains at the C-terminus (7,8). For the
GDF11-induced signal pathway, GDF11 binds to activin receptor type
IIB (ACTRIIB) and subsequently, the complex recruits the activin
receptor-like kinases (ALKs) ALK4, ALK5 and ALK7 to activate the
receptor Smad (R-Smad) pathway involving Smad2/3, which associate
with Smad4. The Smad protein complex then translocates into the
nucleus to positively or negatively control gene expression
(9,10).
GDF11 not only contributes to embryonic development
and histogenesis, but also has a role in metabolic disorders,
cardiovascular diseases and ageing (11–14).
GDF11 is expressed in the liver and has an inhibitory role in liver
development. It was reported that overexpression of GDF11 in
embryos by mRNA microinjection produced a small liver phenotype.
The mechanism possibly comprises the suppression of hepatocyte
proliferation (15). The observation
that GDF11 suppresses liver development raises the question whether
GDF11 has a similar function in liver cancer. In the field of
cancer research, various studies have investigated the role of
GDF11, indicating that it is probably involved in colorectal
cancer, breast cancer and leiomyoma uteri (11,16–18).
However, the role of GDF11 in liver cancer has been rarely
reported, which was therefore investigated in the present
study.
In the present study, the levels of GDF11 in liver
cancer patients and liver cancer cell lines compared with those in
matched normal liver tissue or cells were assessed in order to
identify whether the level of GDF11 changes with the development of
liver cancer. Due to the high homology between the active domain of
GDF11 and myostatin, the expression was assessed at the mRNA level
(19). As GDF11 belongs to the
TGF-β/BMP superfamily, the canonical signaling associated with the
latter should be the basis of the mechanism of action of GDF11.
Therefore, the effects of GDF11 on Smad signaling in the human
hepatoma cancer cell line HepG2 were identified and the effect of
GDF11 on the viability of HepG2 cells was then studied. The present
study was the first to investigate the expression and role of GDF11
in liver cancer patients.
Materials and methods
Reagents and cell lines
The complementary (c)DNA cohort of 10 pairs of human
malignant liver cancer tissues and their corresponding adjacent
non-cancerous liver tissues was purchased from Shanghai Outdo
Biotech Co., Ltd (Shanghai, China; cat. no. cDNA-HLivH30PG01). The
samples were derived from Chinese patients with hepatocellular
carcinoma and were characterized by a pathological grade of II–III,
an age range from 51 to 72 years, no distant metastasis, no
perineural invasion and an American Joint Committee on Cancer
clinical stage of 1–2 (Table I).
Fetal bovine serum (FBS) was purchased from Natocor-Industria
Biológica (Córdoba, Argentina). Recombinant human/mouse/rat GDF-11
was obtained from PeproTech (Rocky Hill, NJ, USA; cat. no. 120–11).
Anti-phosphorylated (p)-Smad3 (Ser423/425), anti-Smad3,
anti-p-Smad2 (Ser465/467), anti-Smad2 and anti-Smad2/3 were
obtained from Cell Signaling Technology (Danvers, MA, USA; cat. no.
12747). Anti-GDF11 antibody was purchased from Abcam (Cambridge,
UK; cat. no. ab71347). Anti-β-actin was obtained from Zhongshan
Goldenbridge Bio (Beijing, China; cat. no. TA-09). The SYBR-Green
RT-PCR kit was obtained from Takara Bio, Inc. (Otsu, Japan).
Primers were designed and synthesized by Shanghai Sangon Biotech.
Co., Ltd. (Shanghai, China). Smad3 inhibitor SIS3 was purchased
from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
 | Table I.Clinicopathological variables of 10
liver cancer patients. |
Table I.
Clinicopathological variables of 10
liver cancer patients.
| Variable | N/value |
|---|
| Age (years ±
SEM) | 57.0±7.77 |
| Gender |
|
| Male | 9 |
|
Female | 1 |
|
Pathological grade | II |
| Tumor site |
|
| Right
liver | 7 |
| Left
liver | 3 |
| Nerve invasion |
|
|
Absent | 6 |
|
Unknown | 4 |
| Vascular
invasion |
|
|
Absent | 4 |
|
Present | 3 |
|
Unknown | 3 |
| AJCC stage |
|
| 1 | 4 |
| 2 | 3 |
|
Unknown | 3 |
Cell culture
The L-02, HepG2 and SMMC-7721 cell lines were
provided by the Chongqing Engineering Research Center of Antitumor
Natural Drugs (Chongqing, China) and purchased from Shanghai Zhong
Qiao Xin Zhou Biotechnology Co., Ltd. (Shanghai, China). HepG2 is
known to be misidentified as a hepatoblastoma cell line (20). Cells were cultured in RPMI 1640
medium (Hyclone; GE Healthcare, Little Chalfont, UK) supplemented
with 10% FBS in a humidified incubator containing 5%
CO2. Recombinant GDF11 pure protein used for treating
the cells was purchased from PeproTech Inc. and for the preparation
of a stock solution, 20 µg GDF11 was dissolved in 40 µl sterile
ultra-pure water according to the manufacturer's protocol.
Recombinant GDF11 pure protein was utilized according to methods
described in previous studies (13,14).
Cells in the control group were treated with the same volume of
sterile ultra-pure water.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
The cDNA microarray is a novel type of cDNA cohort
to detect the expression of a target gene through RT-qPCR. cDNA
samples were derived from 10 pairs of human malignant liver cancer
tissues and their corresponding adjacent non-cancerous liver
tissues with complete clinical information. For the cell lines,
total RNA was extracted from the normal liver cell line L-02 and
liver cancer cell lines (HepG2 and SMMC-7721) with TRIzol reagent
(Thermo Fisher Scientific, Inc., Waltham, MA, USA). The total RNA
was then converted into cDNA with the RevertAid First Strand cDNA
Synthesis Kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Real-time qPCR was performed in a
Roche LightCycler 480II (Roche Diagnostics, Basel, Switzerland)
with the SYBR Green PCR master mix (Takara Bio Inc.). β-actin was
used as an internal reference. The specific primers used to detect
GDF11 were as follows: Forward, 5′-GCCATCAACACCACTCACATT-3′ and
reverse, 5′-CCAATCCCTACTCTGCCAAG-3′. The specific primers used for
β-actin were as follows: Forward, 5′-GAAGAGCTACGAGCTGCCTGA-3′ and
reverse, 5′-CAGACAGCACTGTGTTGGCG-3′. The thermocycling conditions
were as follows: Initial denaturation at 95°C for 30 sec, followed
by 40 cycles of denaturation at 95°C for 5 sec and an
annealing/elongation step at 60°C for 30 sec. Experiments were
performed in triplicate and the values were normalized to β-actin.
Relative expression was analyzed via the 2−∆∆Cq method
(21).
Western blot analysis
Western blot analysis was performed as described in
previous studies by our group (14,22).
Cells were lysed with radioimmunoprecipitation assay buffer
(Beyotime Institute of Biotechnology, Haimen, China) and
centrifuged at 12,000 × g for 15 min at 4°C. The supernatants were
collected and the protein concentrations were determined with a
bicinchoninic acid assay kit (Beyotime Institute of Biotechnology).
The protein (50 µg/lane) was subjected to 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes (Beyotime
Institute of Biotechnology). After blocking with 5% nonfat milk at
4°C for 2 h, the membranes were probed with primary antibodies
against the analyte proteins stated above (1:500 dilution) and
β-actin (1:1,000 dilution) at 4°C overnight. Following three washes
with PBS, the membrane was incubated with mouse anti-rabbit
immunoglobulin G conjugated to horseradish peroxidase (1:1,000
dilution; Cell Signaling Technology, Inc.; cat. no. 7074) at room
temperature for 1 h. A chemiluminescence kit (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) was then used to perform chemiluminescent
detection, according to the manufacturer's instructions. Western
blot bands were quantified by using the ChemiDoc™ Touch Imaging
System (Bio-Rad Laboratories, Inc.) and Image Lab™ Touch Software
(version 1.2; Bio-Rad Laboratories, Inc.).
MTT assay
Cell viability was assessed using an MTT assay
(Biosharp, Shanghai, China), which measures mitochondrial succinate
dehydrogenase activity in living cells. The assay is dependent on
the ability of viable cells to metabolize a water-soluble
tetrazolium salt into a water-insoluble formazan product. Cells
were trypsinized and seeded in a 96-well plate. After adherence,
complete medium was replaced with basal medium for 12 h. Following
an incubation of 24, 48 and 72 h at 37°C with the respective
reagents, a total of 100 µl RPMI-1640 medium (Hyclone; GE
Healthcare) containing 0.5 g/l MTT was added to each well, followed
by 4 h of incubation. Subsequently, the medium was removed by
aspiration, followed by addition of 50 µl dimethyl sulfoxide.
Following incubation at 37°C for 10 min, the absorbance of each
well at 490 nm was measured using a microplate reader.
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. Significance was determined by using Student's t-test or
one-way analysis of variance, followed by the Holm-Sidak test.
Statistical analysis of the data was performed using GraphPad Prism
5 software (GraphPad Software, Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
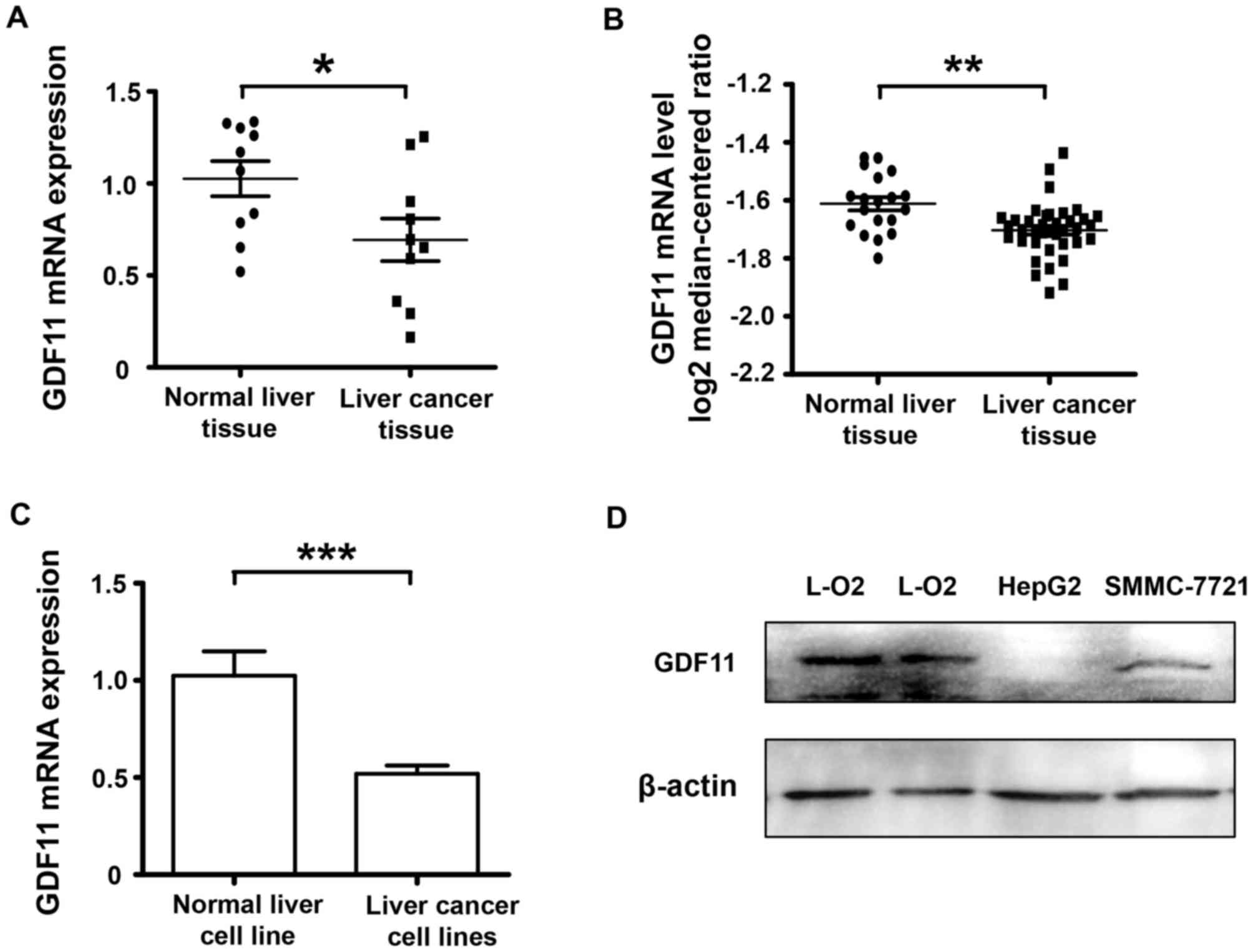
GDF11 is downregulated in liver cancer
tissues and cell lines
A cDNA microarray of 10 paired cancerous and normal
tissue samples from liver cancer patients was used to detect the
mRNA levels of GDF11. The results indicated that the expression of
GDF11 in human malignant liver cancer tissues at the mRNA level was
decreased compared with that in the corresponding adjacent
non-cancerous liver tissues (P<0.05; Fig. 1A). To verify the results, the
database Oncomine was consulted (www.oncomine.com) and the search indicated that GDF11
expression was also declined according the database information
(Fig. 1B). As GDF11 mRNA levels were
lower in the clinical liver cancer tissue samples compared with
those in the normal adjacent tissues, GDF11 levels were also
assessed in liver cancer cell lines and a normal liver cell line to
confirm this trend. In vitro, the level of GDF11 in the
liver cancer cell lines SMMC-7721 and HepG2 was compared with that
in the normal liver cancer cell line L-O2 at the mRNA and protein
level. According to Fig. 1C and D,
GDF11 mRNA and protein levels were significantly decreased in liver
cancer cell lines. These results indicate that GDF11 may serve as a
tumor suppressor in liver cancer. Subsequently, the mechanisms of
GDF11 in liver cancer were assessed in in vitro
experiment.
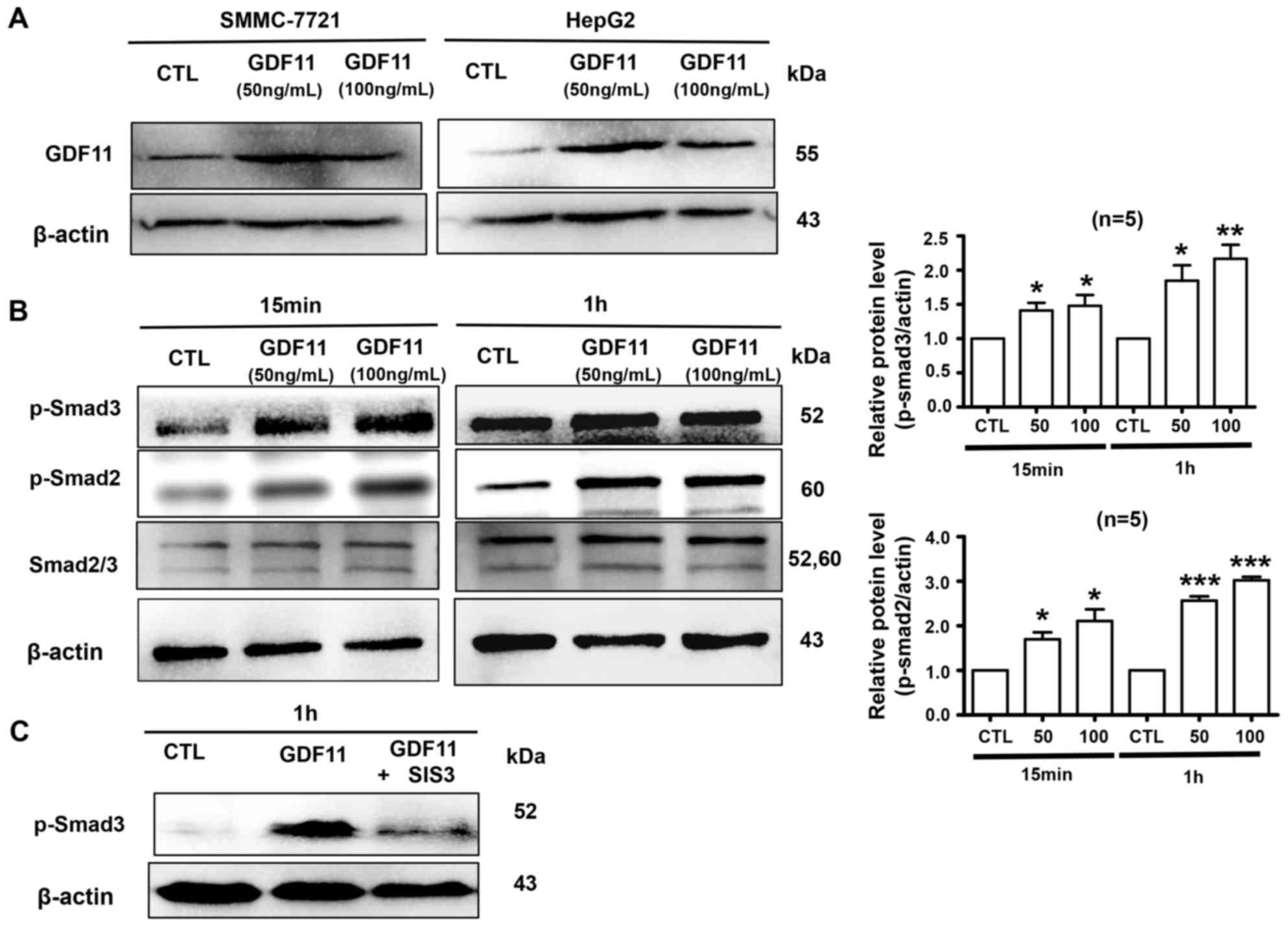
GDF11 activates Smad2/3 signaling in
HepG2 cells and decreases the viability of HepG2 cells
Firstly, we verified GDF11 expression was indeed
increased following the treatment of cells with recombinant GDF11.
HepG2 and SMMC-7721 cells were treated with GDF11 (50 or 100
ng/ml), which significantly increased the levels of GDF11 (Fig. 2A). Activation of Smad2/3 is a typical
effect of TGF-β members (23);
therefore, it was then detected whether GDF11 activates Smad2/3
signaling in HepG2 cells. The hepatoma cell line HepG2 was treated
with GDF11 (50 and 100 ng/ml) for 15 min or 1 h. The results
indicated that GDF11 significantly increased the levels of p-Smad2
and p-Smad3 compared with those in the control group (Fig. 2B). Furthermore, Smad3 activation by
GDF11 was inhibited by Smad3 inhibitor SIS3 (5 µM; Fig. 2C). Smad2 and Smad3 are known to
participate in tumor-associated processes, Smad2 mutations have
been detected in numerous cancer types and Smad2, as a tumor
suppressor, mediates the TGF-β-induced tumor-suppressive function
(24–26). Similarly, Smad3 also has an
inhibitory function in tumors by suppressing cell proliferation and
promoting apoptosis (27).
Therefore, GDF11 may suppress processes associated with liver
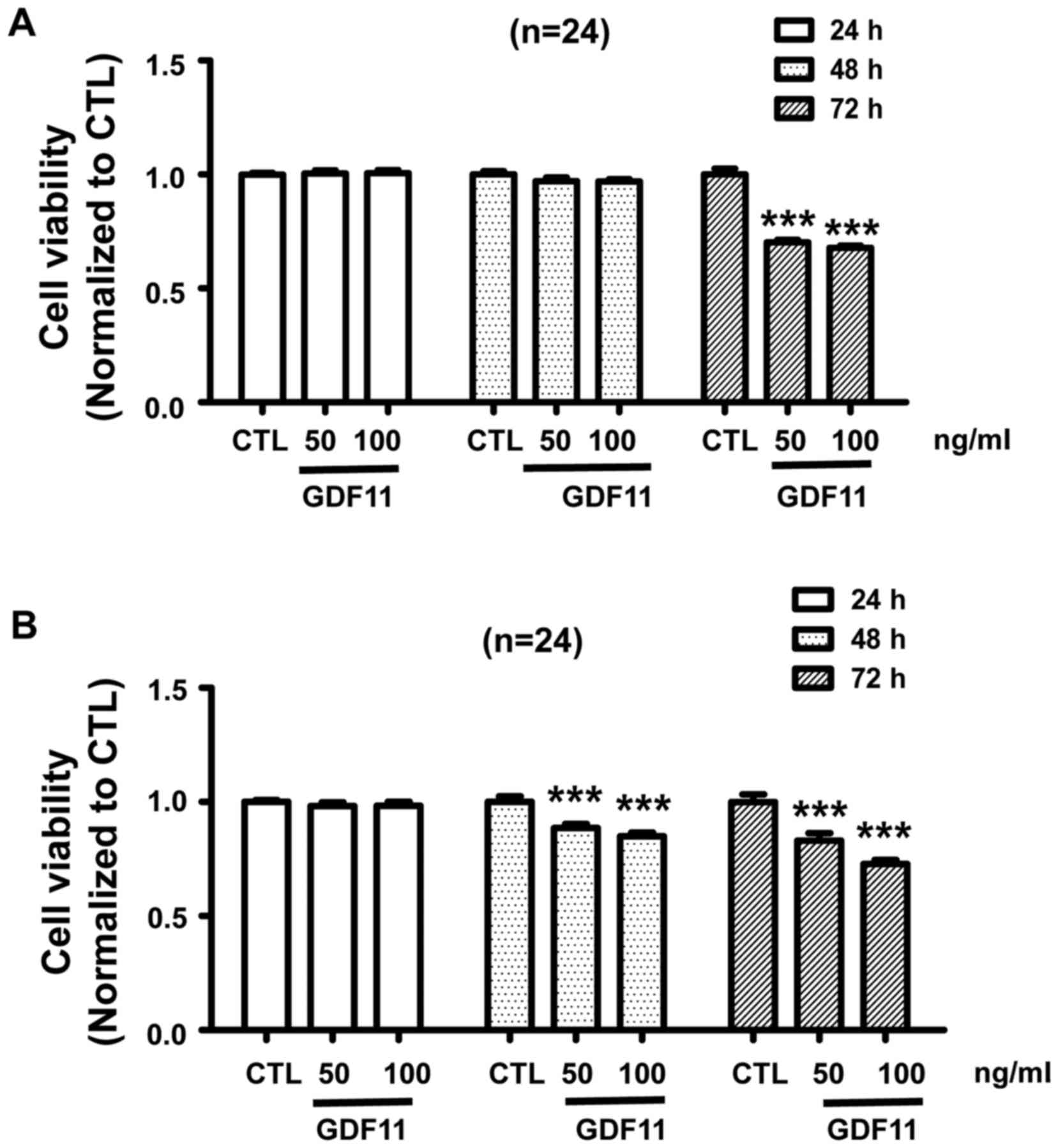
cancer by activating Smad2/3 signaling. In addition, In order to
verify whether GDF11 acts as a tumor suppressor in vitro,
HepG2 and SMMC-7721 cells were treated with GDF11 for different
duration (24, 48 and 72 h) at the concentration of 50 or 100 ng/ml.
According to the results, GDF11 did not affect the cell viability
for 24 and 48 h, but the viability of HepG2 cells was significantly
decreased after treatment for 72 h (Fig.
3A). For the SMMC-7721 cell line, a similar result was
obtained, namely that GDF11 decreased cell viability after
treatment of 48 and 72 h (Fig. 3B).
These results demonstrate the tumor suppressive effect of GDF11 in
liver cancer.
Discussion
Exploring novel tumor markers is important for
improving the diagnosis and treatment of liver cancer. GDF11 was
reported to be expressed in the liver and to have a regulatory
function in liver development (13).
Overexpression of GDF11 by mRNA microinjection in an embryonic
animal model led to a small liver phenotype in a dose-dependent
manner, indicating that GDF11 suppressed the growth/expansion phase
of liver development, possibly by acting as an inhibitor of cell
proliferation. Based on this result, it may be hypothesized that
GDF11 has a similar role in liver cancer, namely that of a tumor
suppressor. Several studies have suggested that GDF11 is involved
in cancer (11,17,18). A
study by Yokoe et al (11)
indicated that GDF11 was positively associated with processes of
colorectal cancer. They identified that the expression of GDF11
mRNA was increased in colorectal cancer tissues and that patients
with high GDF11 expression had a poorer prognosis. However, histone
deacetylases (HDACs), key transcriptional regulators that inhibit
GDF11 gene expression, were proved to promote tumor growth in
animals (28,29). Several HDAC inhibitors, including
vorinostat (SAHA), romidepsin, belinostat and panobinostat, have
been approved by the USA food and drug administration to treat
cancer (29). The mechanisms of
action of HDAC inhibitors may include inhibition of abnormal cell
growth by inactivation of HDAC3 and activation of GDF11 expression
(14). The function of GDF11 in
various cancer types remains controversial. Therefore, the present
study investigated the possible role of GDF11 as a tumor promoter
or inhibitor in liver cancer. First, the mRNA expression of GDF11
in cancerous liver tissues was compared with that in normal liver
tissue. The mRNA and protein levels of GDF11 in a normal liver cell
line and liver cancer cell lines were also assessed. In a
subsequent in vitro experiment, the effect of GDF11 on the
viability of the liver cancer cell lines HepG2 and SMMC-7721 was
observed. The results indicated that GDF11 expression was decreased
in liver cancer tissues compared with that in normal liver tissues,
which was in accordance with the result retrieved from the Oncomine
database. These results were consistent with those obtained with
the cell lines results, namely that GDF11 was downregulated in
liver cancer cell lines. In vitro, treatment with
recombinant GDF11 for up to 72 h led to a reduction of cell
viability in HepG2 and SMMC-7721 cells, possibly by activating
Smad2/3 signaling. It was previously demonstrated that GDF11
treatment leads to an upregulation of p-Smad3 in multiple cell
types, including pluripotent stem cell-derived cardiomyocytes,
human skeletal muscle-derived cells and human umbilical vein
endothelial cells, to regulate cell proliferation and muscle
regeneration (10,13,14,30),
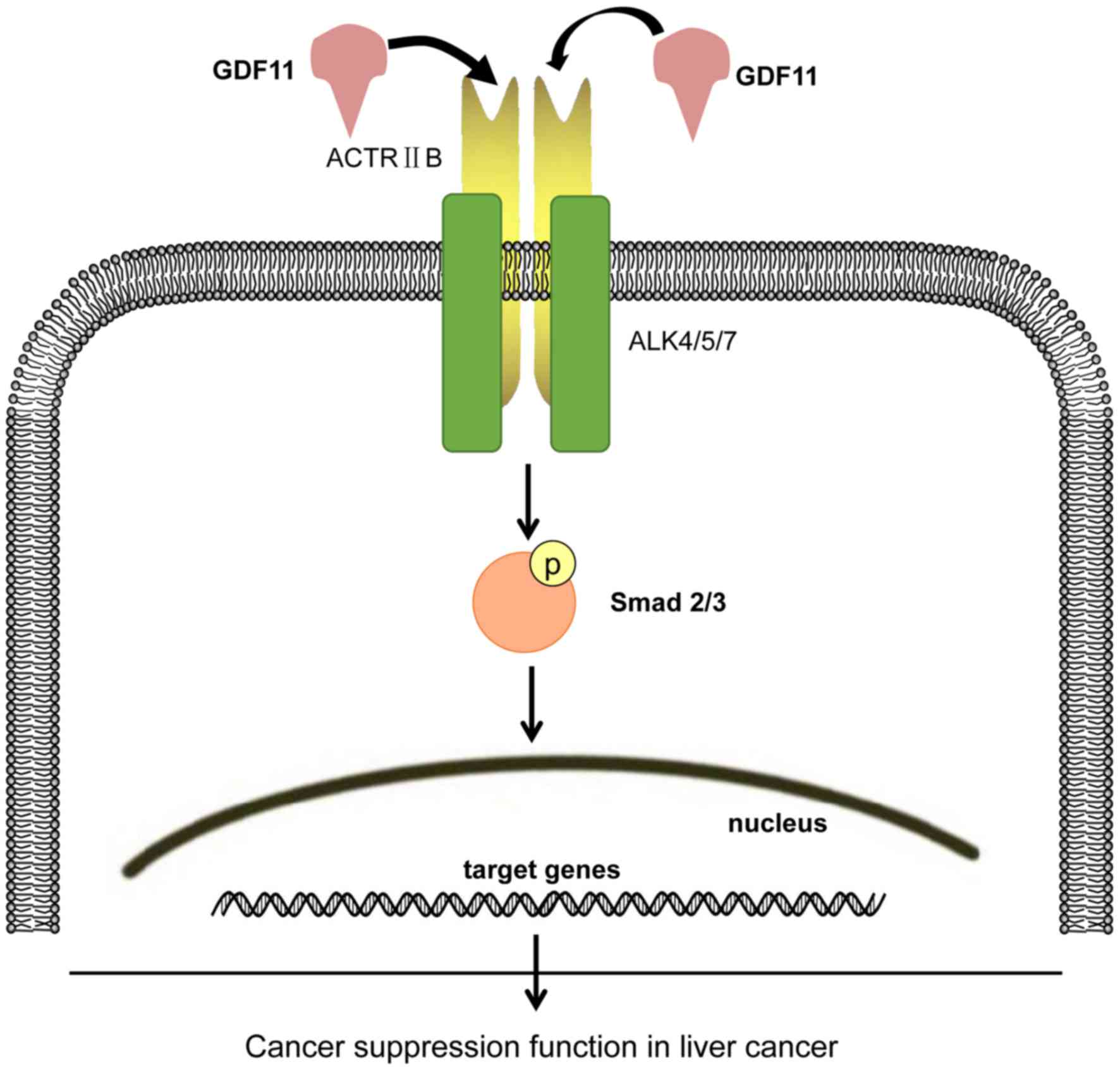
which is consistent with the present results. Following activation
of Smad2/3 by phosphorylation through GDF11, p-Smad2/3 forms a
complex with Smad4, which then translocates to the nucleus to
regulate cell function by affecting the expression of associated
genes. The probable signaling pathway is summarized in Fig. 4.
In the present study, 10 pairs of human malignant
liver cancer tissues and their corresponding adjacent non-cancerous
liver tissues were assessed in a cDNA microarray. The samples are
characterized by pathological grading II–III, age ranging from 51
to 72 years, no distant metastasis, no perineural invasion and
American Joint Committee on Cancer clinical stage 1–2, as presented
in Table I. GDF11 mRNA expression
was declined in these liver cancer tissues compared with that in
adjacent non-cancerous liver tissues. However, Yokoe et al
(11) reported that the expression
of GDF11 mRNA was increased in colorectal cancer tissues and that
patients with high GDF11 expression in their tumors had a higher
frequency of lymph node metastasis and more cancer-associated
mortalities. This divergent result may be due to the dual role of
TGF-β members. In early carcinomas, TGF-β signaling pathways exert
tumor suppressor effects and as tumors develop and progress, the
role of TGF-β signaling switches to promote cancer metastasis
(31). Therefore, further
investigation is required to explore the exact functional role of
GDF11 regarding the regulation of cell viability, death and
proliferation of liver cancer cells and the underlying molecular
mechanisms, in addition to the study of GDF11 expressional changes
in late-stage liver carcinomas.
In conclusion, the present study indicated that
GDF11 reduced the viability of liver cancer cells, probably and at
least in part via activation of Smad2/3 signaling. GDF11 mRNA
expression was downregulated in liver cancer tissues compared with
that in the corresponding normal tissues as indicated by a cDNA
microarray and a search of the database Oncomine, which provided
similar results to those obtained with liver cancer cell lines.
These results indicate that GDF11 is a novel candidate for a tumor
marker in patients with liver cancer.
Acknowledgements
This study was supported by the Science and
Technology Research Projects of Chongqing Education Commission
(grant nos. KJ1725391 and KJ1710238), the Natural Science Research
Projects of Chongqing Three Gorges Medical College (grant no.
2016xmpxz02) and the National Natural Science Foundation of China
(grant no. 61705025).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baig JA, Alam JM, Mahmood SR, Baig M,
Shaheen R, Sultana I and Waheed A: Hepatocellular carcinoma (HCC)
and diagnostic significance of A-fetoprotein (AFP). J Ayub Med Coll
Abbottabad. 21:72–75. 2009.PubMed/NCBI
|
|
4
|
Zhao YJ, Ju Q and Li GC: Tumor markers for
hepatocellular carcinoma. Mol Clin Oncol. 1:593–598. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nakashima M, Toyono T, Akamine A and
Joyner A: Expression of growth/differentiation factor 11, a new
member of the BMP/TGFbeta superfamily during mouse embryogenesis.
Mech Dev. 80:185–189. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gamer LW, Wolfman NM, Celeste AJ,
Hattersley G, Hewick R and Rosen V: A novel BMP expressed in
developing mouse limb, spinal cord, and tail bud is a potent
mesoderm inducer in Xenopus embryos. Dev Biol. 208:222–232. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Essalmani R, Zaid A, Marcinkiewicz J,
Chamberland A, Pasquato A, Seidah NG and Prat A: In vivo functions
of the proprotein convertase PC5/6 during mouse development: Gdf11
is a likely substrate. Proc Natl Acad Sci USA. 105:pp. 5750–5755.
2008; View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsuda T, Iwai N, Deguchi E, Kimura O, Ono
S, Furukawa T, Sasaki Y, Fumino S and Kubota Y: PCSK5 and GDF11
expression in the hindgut region of mouse embryos with anorectal
malformations. Eur J Pediatr Surg. 21:238–241. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rochette L, Zeller M, Cottin Y and Vergely
C: Growth and differentiation factor 11 (GDF11): Functions in the
regulation of erythropoiesis and cardiac regeneration. Pharmacol
Ther. 156:26–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Camici GG, Savarese G, Akhmedov A and
Lüscher TF: Molecular mechanism of endothelial and vascular aging:
Implications for cardiovascular disease. Eur Heart J. 36:3392–3403.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yokoe T, Ohmachi T, Inoue H, Mimori K,
Tanaka F, Kusunoki M and Mori M: Clinical significance of growth
differentiation factor 11 in colorectal cancer. Int J Oncol.
31:1097–1101. 2007.PubMed/NCBI
|
|
12
|
Smith SC, Zhang X, Zhang X, Gross P,
Starosta T, Mohsin S, Franti M, Gupta P, Hayes D, Myzithras M, et
al: GDF11 does not rescue aging-related pathological hypertrophy.
Circ Res. 117:926–932. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Loffredo FS, Steinhauser ML, Jay SM,
Gannon J, Pancoast JR, Yalamanchi P, Sinha M, Dall'Osso C, Khong D,
Shadrach JL, et al: Growth differentiation factor 11 is a
circulating factor that reverses age-related cardiac hypertrophy.
Cell. 153:828–839. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang YH, Cheng F, Du XT, Gao JL, Xiao XL,
Li N, Li SL and Dong de L: GDF11/BMP11 activates both Smad1/5/8 and
Smad2/3 signals but shows no significant effect on proliferation
and migration of human umbilical vein endothelial cells.
Oncotarget. 7:11063–11074. 2016.
|
|
15
|
Farooq M, SuLochana KN, Pan X, To J, Sheng
D, Gong Z and Ge R: Histone deacetylase3(HDAC3) is specifically
required for liver development in zebrafish. Dev Biol. 317:336–353.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bajikar SS, Wang CC, Borten MA, Pereira
EJ, Atkins KA and Janes KA: Tumor-suppressor inactivation of GDF11
occurs by precursor sequestration in triple-negative breast cancer.
Dev Cell. 43:418–435. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Alvarez C, Aravena A, Tapia T, Rozenblum
E, Solís L, Corvalán A, Camus M, Alvarez M, Munroe D, Maass A and
Carvallo P: Different Array CGH profiles within hereditary breast
cancer tumors associated to BRCA1 expression and overall survival.
BMC Cancer. 16:2192016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Auguściak-Duma A and Sieroń AL: Molecular
characteristics of leiomyoma uteri based on selected compounds of
the extracellular matrix. Postepy Hig Med Dosw (Online).
62:148–165. 2008.(In Polish). PubMed/NCBI
|
|
19
|
McNally EM: Questions and answers about
myostatin, GDF11, and the aging heart. Circ Res. 118:6–8. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
López-Terrada D, Cheung SW, Finegold MJ
and Knowles BB: Hep G2 is a hepatoblastoma-derived cell line. Hum
Pathol. 40:1512–1515. 2009. View Article : Google Scholar
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sheng Y, Sun B, Guo WT, Liu X, Wang YC,
Xie X, Xiao XL, Li N and Dong DL:
[4-(6-(4-isopropoxyphenyl)pyrazolo(1,5-a)pyrimidin-3-yl)quinoline]
is a novel inhibitor of autophagy. Br J Pharmacol. 171:4970–4980.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dwivedi SK, McMeekin SD, Slaughter K and
Bhattacharya R: Role of TGF-β signaling in uterine carcinosarcoma.
Oncotarget. 6:14646–14655. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Levy L and Hill CS: Alterations in
components of the TGF-beta superfamily signaling pathways in human
cancer. Cytokine Growth Factor Rev. 17:41–58. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Prunier C, Ferrand N, Frottier B, Pessah M
and Atfi A: Mechanism for mutational inactivation of the tumor
suppressor Smad2. Mol Cell Biol. 21:3302–3313. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang J, Wahdan-Alaswad R and Danielpour D:
Critical role of Smad2 in tumor suppression and transforming growth
factor-beta-induced apoptosis of prostate epithelial cells. Cancer
Res. 69:2185–2190. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Millet C and Zhang YE: Roles of Smad3 in
TGF-beta signaling during carcinogenesis. Crit Rev Eukaryot Gene
Expr. 17:281–293. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ropero S and Esteller M: The role of
histone deacetylases (HDACs) in human cancer. Mol Oncol. 1:19–25.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Y and Seto E: HDACs and HDAC inhibitors
in cancer development and therapy. Cold Spring Harb Perspect Med.
6(pii): a0268312016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Egerman MA, Cadena SM, Gilbert JA, Meyer
A, Nelson HN, Swalley SE, Mallozzi C, Jacobi C, Jennings LL, Clay
I, et al: GDF11 increases with age and inhibits skeletal muscle
regeneration. Cell Metab. 22:164–174. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lebrun JJ: The dual role of TGFβ in human
cancer: From tumor suppression to cancer metastasis. ISRN Mol Biol.
2012:3814282012. View Article : Google Scholar : PubMed/NCBI
|