Introduction
Alcoholic liver disease (ALD) refers to the liver
injury caused by excessive drinking (1), the early clinical symptoms mainly
include alcoholic fatty liver, alcoholic hepatitis and alcoholic
liver fibrosis, and it can ultimately develop into the end-stage
liver disease. With the improvement of people's living standards
and the sharp increase in alcohol consumption, its morbidity and
mortality rates are increasingly higher in the world (2). The specific pathogenesis of ALD is more
complex, and a large number of studies have shown that the
oxidative stress, inflammation and lipid metabolism disorder
(3) caused by alcohol metabolism
play key roles in the occurrence and development of ALD. The
relevant mechanism research has shown that alcohol metabolism can
increase the synthesis of triglycerides (TG) in the body. The
increased TG and the accumulation of fatty acids in hepatocytes
result in the fatty degeneration of liver (4), and excessive fat accumulation can also
improve the oxidative stress and inflammatory response, thereby
aggravating ALD and leading to lipid hepatitis (5). Some relevant studies have also
confirmed that the overexpression of very-low density lipoprotein
receptor (VLDLR) in hepatocytes is an important mechanism leading
to alcoholic fatty liver (6). In the
lipid hepatitis caused by high-fat diet, the overexpression of
VLDLR in hepatocytes induced by endoplasmic reticulum stress plays
an important role (7). The
expression of VLDLR is regulated by nuclear factor 2 (Nrf2), and
oxidative stress can activate Nrf2. Therefore, the upregulation of
Nrf2 caused by oxidative stress in ALD can positively regulate the
protein expression of VLDLR (6).
Ethyl pyruvate (EP) has strong anti-bacterial,
anti-virus, anti-inflammatory, anti-allergy and vasodilator effects
(8). Many experimental studies have
also confirmed its strong anti-oxidant and anti-apoptotic effects
(9). Therefore, it is boldly
speculated that EP can alleviate the liver damage caused by ALD.
This study aimed to investigate the protective effect of EP on ALD
and its possible relevant mechanism using the ALD mouse model
(10,11).
Materials and methods
Animals and treatments
A total of 30 male C57/BL6 mice of clean grade aged
6–8 weeks weighing 21±2 g were randomly divided to three groups:
normal control group (Control group, n=10), alcoholic liver disease
group (ALD group, n=10) and ethyl pyruvate group (EP group, n=10).
EP group was pretreated with gavage using EP (100 mg/kg) for 15
consecutive days. Control group and ALD group were treated with the
same volume of normal saline. After the last gavage, EP and ALD
group were treated with the intraperitoneal injection of 50%
alcoholic solution (10 ml/kg). This study was approved by the
Animal Ethics Committee of Institute of Materia Medica of Shandong
Academy of Medical Sciences (Jinan, China). After that, ALD and EP
group were treated with alcohol (4 g/kg·day, alcohol content of
52%); alcohol weight (g) = liquor volume (ml) × alcohol content in
liquor (v/v) × alcohol density (0.8 g/ml). Mice were fed for 4
weeks and mice in Control group were treated with the gavage using
the same volume of normal saline. After the gavage, blood was drawn
from the retrobulbar venous plexus of mice in each experimental
group. Instantly, one part of it was used to measure fasting blood
glucose (FBG) and to count platelets, and for statistical analysis;
another part was centrifuged for 10 min at 2,500 × g to separate
the serum. Then a small piece of liver tissue was taken from the
same part of the right lobe of liver in the thoracotomy. Liver
homogenate (10%) was prepared using the normal saline at 4°C, and
centrifuged for 10 min at 2,500 × g; and the supernatant was taken.
Superoxide dismutase (SOD) and malondialdehyde (MDA) in liver
tissues in each group were measured. The samples of left lobe of
liver were soaked in 4% formaldehyde solution, and the remaining
liver tissues were cut into small pieces and stored at −80°C for
detection later.
Serum aspartate aminotransferase
(AST), Alanine aminotransferase (ALT), triglyceride (TG), free
fatty acid (FFA) assays
Liver enzyme and blood lipid in the serum samples
collected above were detected, including ALT, AST, TG and FFA.
Experimental animals (Keygen, Nanjing, China) were used in the
detection using the fully automatic biochemical analyzer.
Detection of morphological changes in
liver tissues and apoptosis
After fixation using 4% formaldehyde solution, the
liver tissues collected from each experimental group were prepared
into paraffin sections according to the typical method (12). The pathological changes in liver
tissues were observed under an optical microscope (×400).
RNA isolation and real-time PCR
Total RNA was isolated from frozen liver tissues and
serum and real-time reverse transcription-polymerase chain reaction
(RT-PCR) was performed as previously described (13). The primers used in the present study
(Table I) were selected from the
PubMed database. The real-time PCR data were analyzed using the
relative gene expression (i.e., ∆∆Cq) method (University
of Leicester, London, UK).
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Sequence |
|---|
| IL-6 | F:
5′-CCCTGCGTTTCTCTGCAAAC-3′ |
|
| R:
3′-TTTCAGGGTGGAAGGCAGAC-5′ |
| TNF-α | F:
5′-CATCCGTTCTCTACCCAGCC-3′ |
|
| R:
3′-AATTCTGAGCCCGGAGTTGG-5′ |
| IL-10 | F:
5′-CCTCTGGATACAGCTGCGAC-3′ |
|
| R:
3′-GTAGATGCCGGGTGGTTCAA-5′ |
| TGF-β1 | F:
5′-AGGGCTACCATGCCAACTTC-3′ |
|
| R:
3′-CCACGTAGTAGACGATGGGC-5′ |
| PPAR-α | F:
5′-GGGTACCACTACGGAGTTCACG-3′ |
|
| R:
5′-CAGACAGGCACTTGTGAAAACG-3′ |
| CYP2E1 | F:
5′-CTTGCTTGTCTGGATCGCCA-3′ |
|
| R:
5′-GGGGCAGGTTCCAACTTCTA-3′ |
| Nrf2 | F:
5′-GGGTACCACTACGTTCACAG-3′ |
|
| R:
5′-CAGACAGGCACTTGTGACG-3′ |
| VLDLR | F:
5′-CTCTTGTCTGGATCAAGCCA-3′ |
|
| R:
5′-GGCAGGTTCCAACAATTCTA-3′ |
SOD and MDA concentration assays
The oxidative stress indexes of the serum samples
and liver tissue homogenate collected above were detected,
including MDA and SOD. MDA content was detected using the
thiobarbituric acid (TBA) method; and SOD activity was determined
using the nitrite method (14).
Western blot analysis
Liver tissues were homogenized and proteins were
detected by western blot using specific antibodies as previously
described (13). Anti-CYP2E1
anti-PPAR-α, anti-Nrf2, anti-VLDLR and anti-GAPDH were obtained
from Abcam (Cambridge, UK).
Statistical analysis
SPSS 18.0 (version X; IBM, Armonk, NY, USA) software
was used for the statistical analysis of relevant experimental data
in this study. All data are presented as mean ± standard deviation.
The differences between the two groups were compared using t-test.
P<0.05 suggested that the difference was statistically
significant.
Results
EP reduces ALT/AST levels and liver
morphological changes caused by ALD, inhibites TG deposition and
reduces FFA level
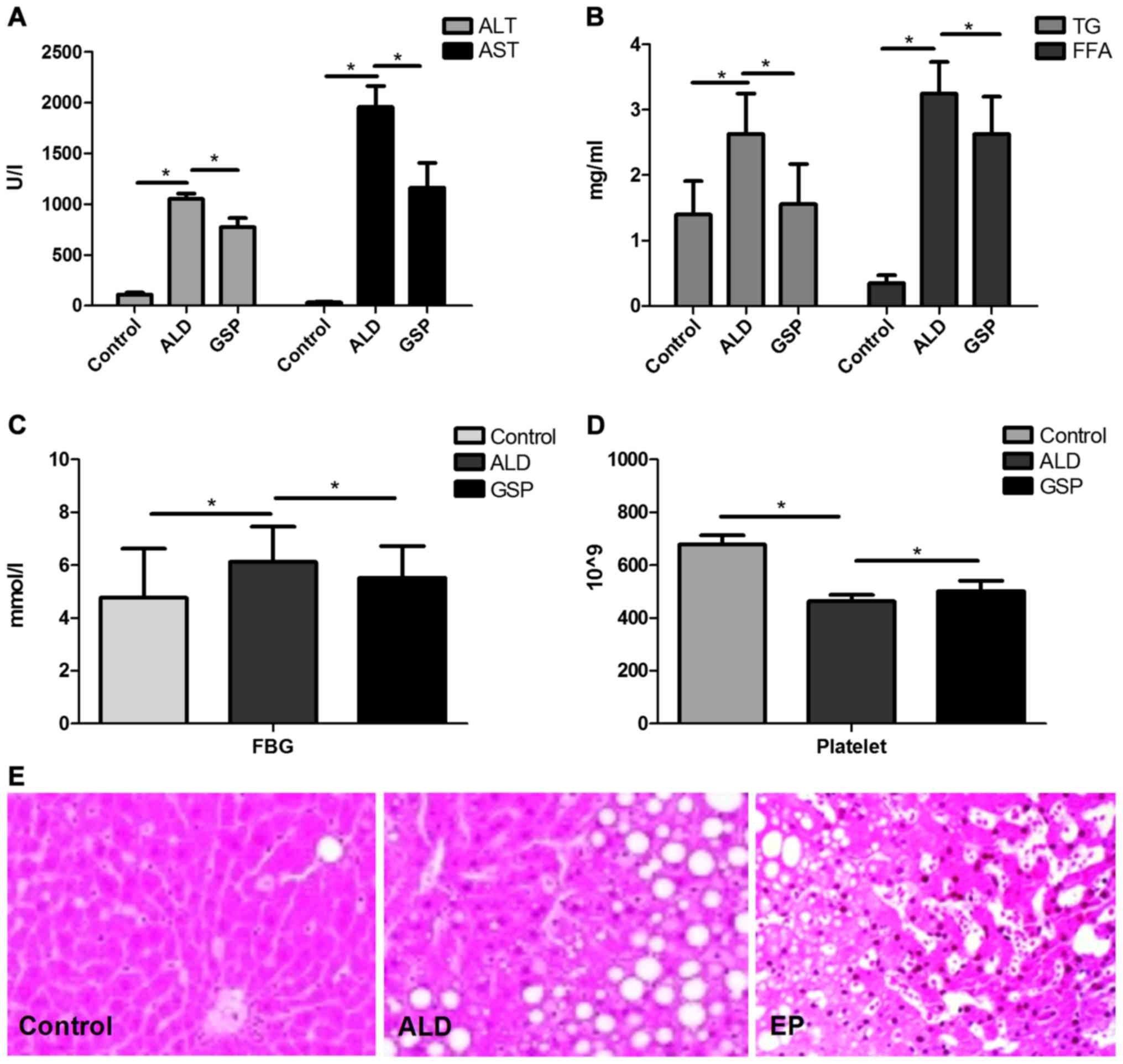
As shown in Fig. 1,
both the significantly increased ALT/AST (Fig. 1A) levels and heavier fatty changes
(Fig. 1E) in ALD group compared with
those in Control group, proved that intraperitoneal injection of
alcohol can cause acute alcoholic liver injury. The significant
increases in TG/FFA (Fig. 1B) and
FBG (Fig. 1C) confirmed that acute
alcoholic liver injury can lead to ALD; compared to the ALD group,
the significantly decrease of ALT/AST, TG/FFA, FBG in EP group
coupled with the significant increase of platelet in EP (Fig. 1D), proved that EP can alleviate liver
injury caused by alcohol and prevent ALD.
EP decreases the expression levels of
pro-inflammatory factors and increased the expression levels of
anti-inflammatory factors in ALD
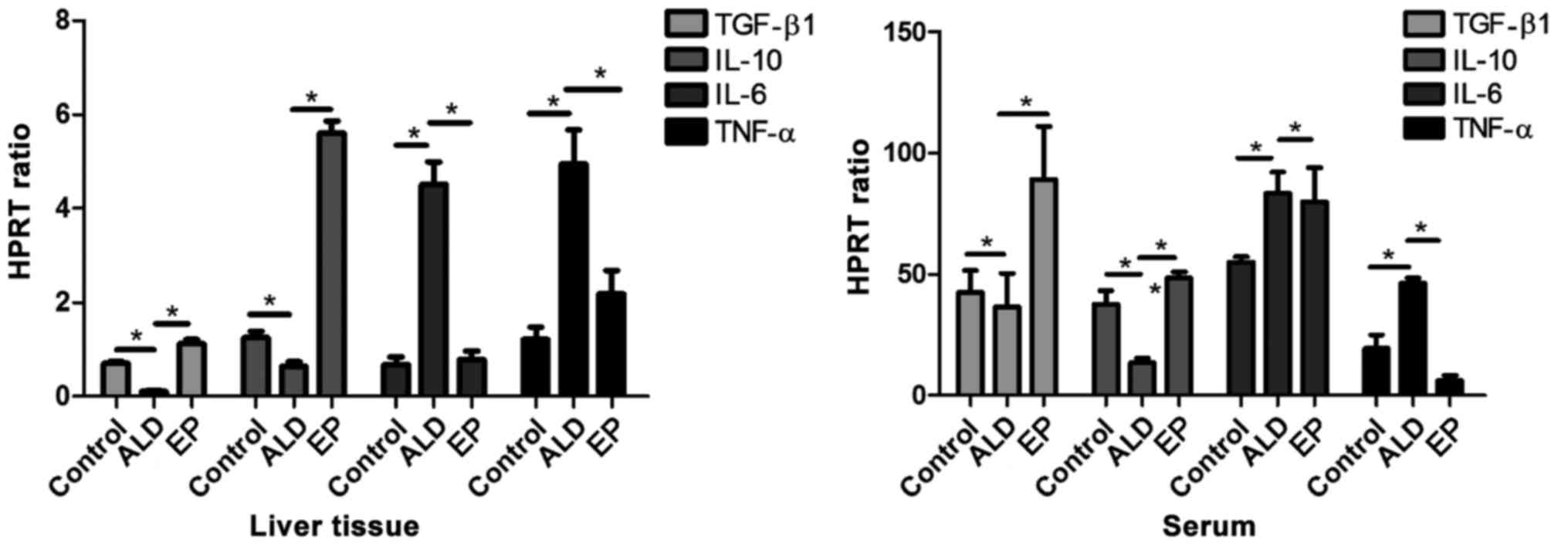
As shown in Fig. 2,
the mRNA expression levels of anti-inflammatory factors
[transforming growth factor-β (TGF-β)/interleukin-10 (IL-10)] in
liver tissues and serum in EP group were significantly increased
compared with those in ALD group, and the differences were
statistically significant. The mRNA expression levels of
pro-inflammatory factors (IL-6/TNF-α) in EP group were
significantly decreased compared with those in ALD group, and the
differences were statistically significant (P<0.05), proving
that EP can alleviate the damage caused by ALD through increasing
the expression levels of anti-inflammatory factors and decreasing
the expression levels of pro-inflammatory factors.
EP increased the mRNA expression level
of PPAR-α and decreased the mRNA expression levels of CYP2E1, Nrf2
and VLDLR
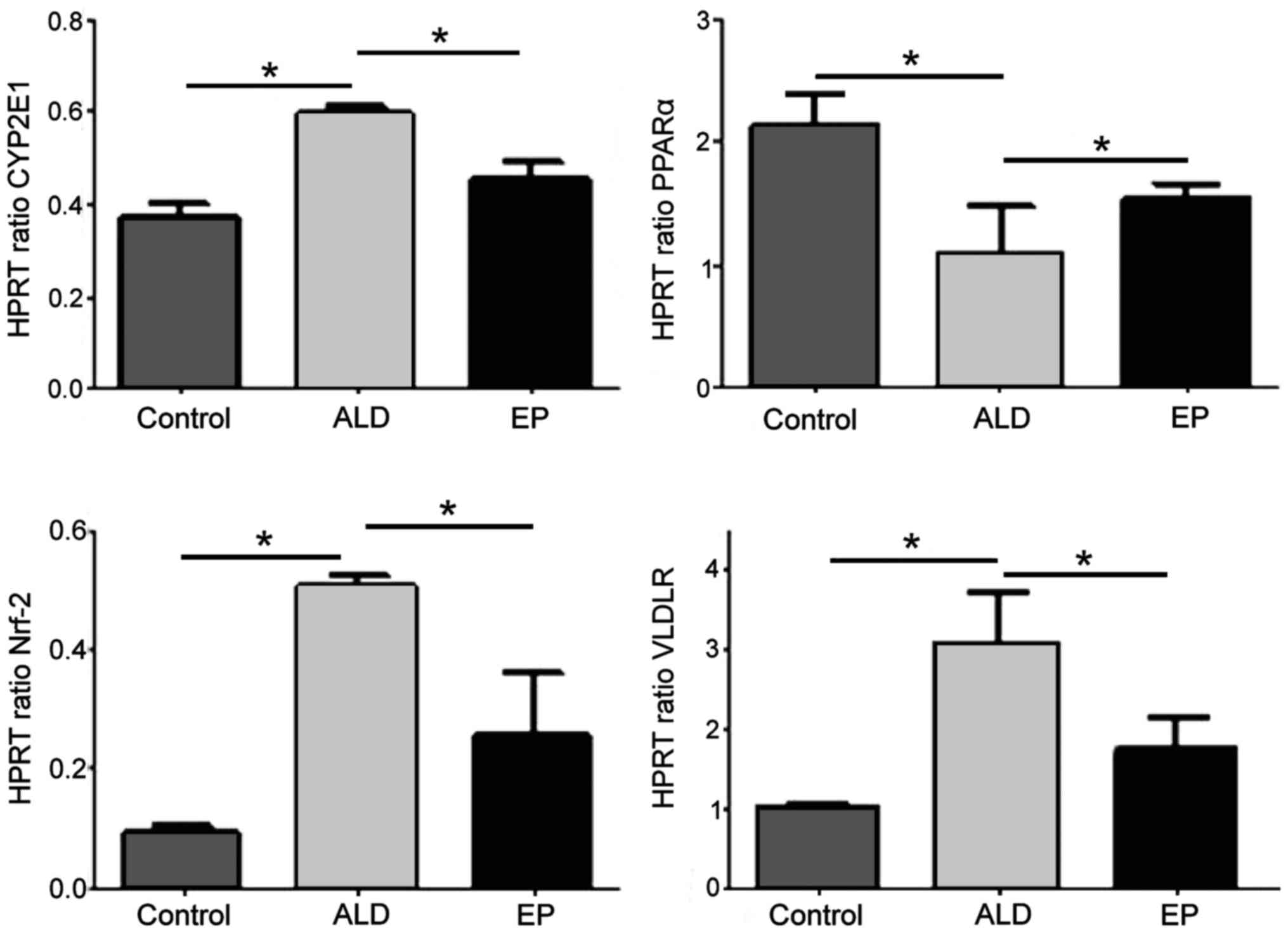
As shown in Fig. 3,
the mRNA expression levels of CYP2E1, Nrf2 and VLDLR in ALD group
were significantly increased compared with those in EP group, and
the differences were statistically significant (P<0.05).
However, the mRNA expression level of PPAR-α in ALD group was
significantly decreased compared with that in EP group, and the
difference was statistically significant (P<0.05), indicating
that EP can inhibit the expression levels of CYP2E1, Nrf2 and VLDLR
and increase the expression of PPAR-α in ALD.
EP negatively regulated PPAR-α-CYP2E1
signaling pathway and ROS-Nrf2 signaling pathway to reduce the
VLDLR protein expression level
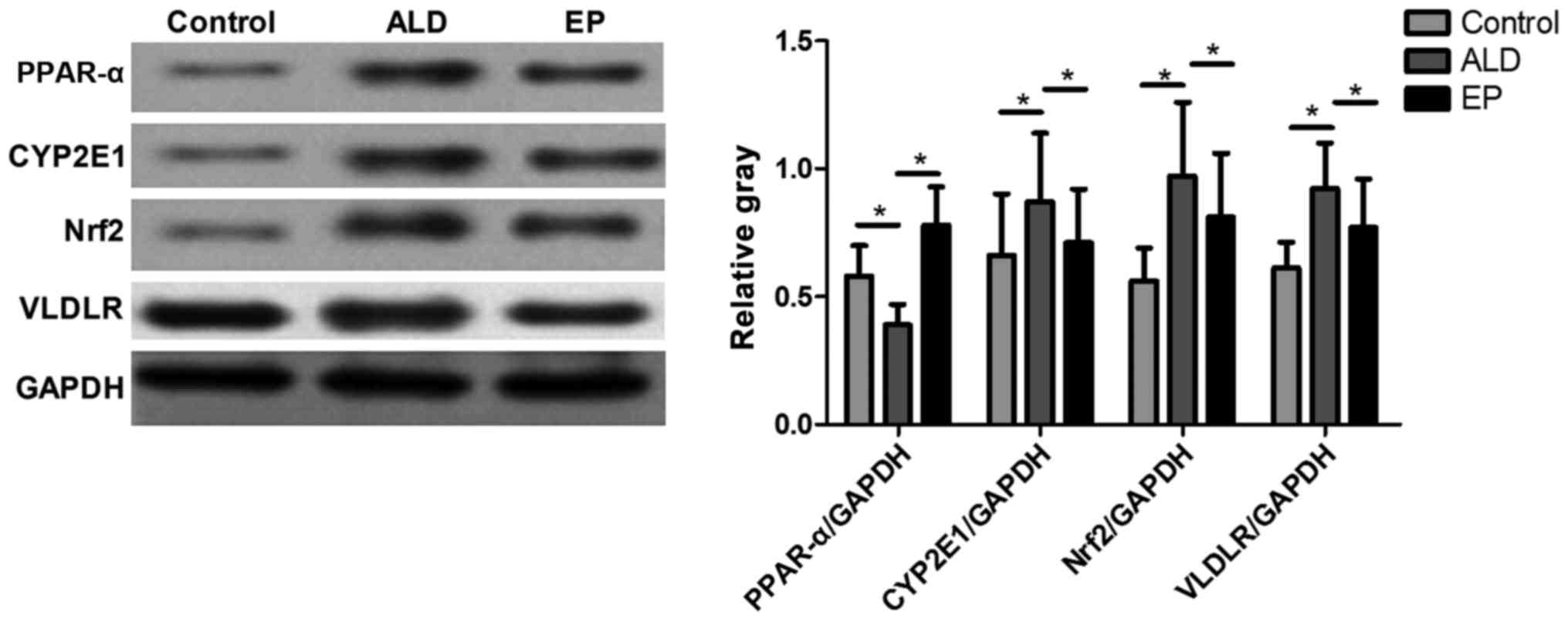
As shown in Fig. 4,
the protein expression levels of CYP2E1, Nrf2 and VLDLR in ALD
group were significantly increased compared with those in EP group,
but the protein expression level of PPAR-α in ALD group was
significantly decreased compared with that in EP group and the
difference was statistically significant (P<0.05), indicating
that EP can inhibit the protein expression levels of CYP2E1, Nrf2
and VLDLR and increase the protein expression of PPAR-α in ALD. As
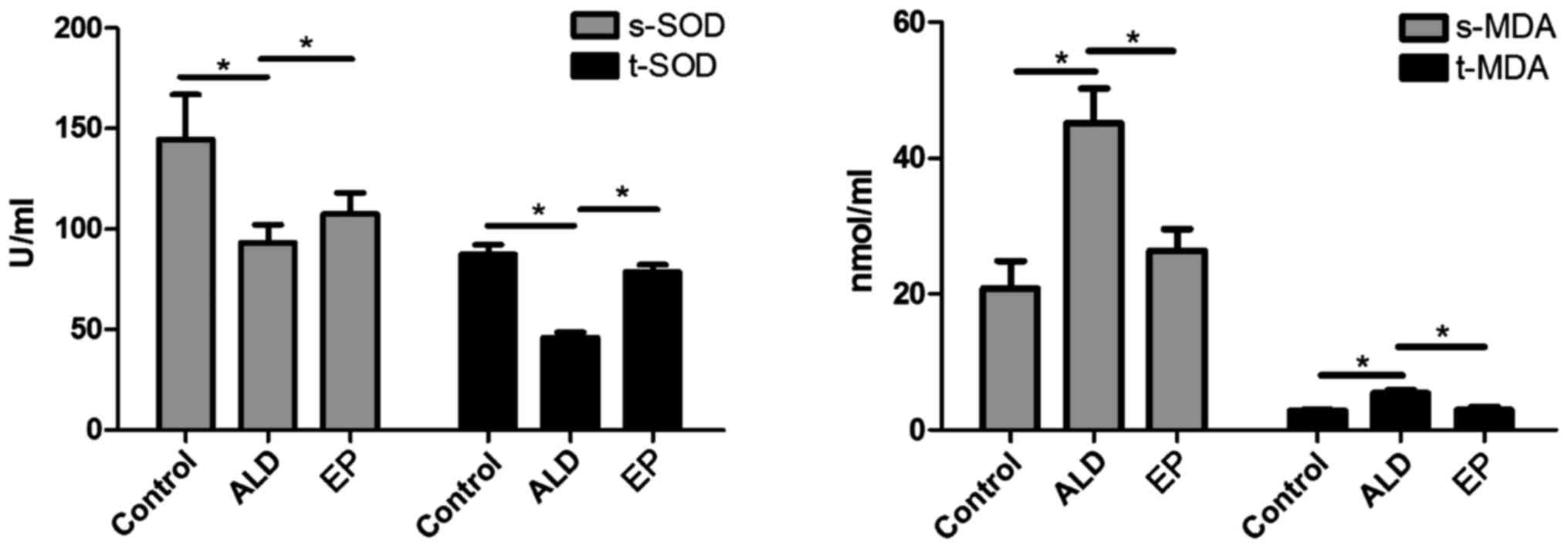
shown in Fig. 5, the SOD content in
plasma and liver tissues in EP group was significantly increased
compared with that in ALD group, and the difference was
statistically significant (P<0.05); but the MDA content in EP
group was significantly decreased compared with that in ALD group
(P<0.05), indicating that EP can alleviate the oxidative stress
injury caused by ALD.
As shown in Figs.
3–5, in EP group, CYP2E1
expression and MDA content were decreased, while PPAR-α expression
and SOD content were increased, indicating that EP can negatively
regulate the CYP2E1-PPAR-α-ROS signaling pathway to alleviate the
oxidative stress injury. However, in EP group, the expression
levels of Nrf2 and VLDLR were decreased, indicating that EP can
negatively regulate the Nrf2-VLDLR signaling pathway and reduce the
VLDLR expression to alleviate the alcoholic fatty liver.
Discussion
The results of this study confirmed that EP can
reduce the serum ALT/AST levels, TG and FFA concentrations and
VLDLR protein expression in ALD mice, and alleviate the
degeneration degree of alcoholic fatty liver, which has a good
preventive and protective effect on ALD. Studies have confirmed
that the oxidative stress, inflammation and lipid metabolism
disorder caused by alcohol metabolism are important causes of ALD
(1), so we investigated the
preventive and protective effect of EP on ALD from the
inflammation, oxidative stress and lipid metabolism.
Studies have confirmed that TNF-α can stimulate the
decomposition of adipose tissues and increase the FFA content in
the body (15), but the excessive
FFA will be transported to the liver and accumulated in hepatocytes
(16), eventually leading to the
fatty degeneration or even necrosis of hepatocytes (17). Studies have also shown that the
action of excessive FFA on hepatocytes can increase the oxygen
consumption of liver tissues and aggravate the lipid peroxidation,
further aggravating the liver injury (18). Therefore, it is speculated that EP
can alleviate the alcoholic liver injury due to its strong
anti-inflammatory effect, which can inhibit the expression levels
of pro-inflammatory factors and increase the expressions of
anti-inflammatory factors. In this experiment, compared with those
in ALD group (Fig. 2), the mRNA
expression levels of pro-inflammatory factors (IL-6/TNF-α) in liver
tissues and plasma in EP group were significantly decreased, while
the mRNA expression levels of anti-inflammatory factors
(TGF-β/IL-10) were significantly increased, which were consistent
with our speculation.
In addition, it is reported in the literature
(19) that SOD is one of the
important antioxidant enzymes for the body to scavenge free
radicals, and the intake of alcohol can inhibit its activity and
accelerate liver injury (20). It is
also reported in the literature that oxidative stress can induce
the hepatocyte mitochondrial dysfunction, leading to hepatocyte
degeneration or even necrosis (21).
Therefore, it is speculated that the strong antioxidant effect of
EP is one of the reasons to alleviate alcoholic liver injury, which
is manifested in that it can increase the SOD activity and decrease
the MDA production. In this study, the SOD activities in serum and
liver tissues in EP group (Fig. 5)
were significantly increased compared with those in ALD group, but
the MDA content was decreased, and the study proved that MDA is one
of the important products of oxidative stress (22). The results of this experiment are
also consistent with our speculation.
A further study (23)
indicated that EP regulation of inflammation and oxidative stress
is closely related to PPAR-α. Studies (24) have confirmed that the phosphorylated
protein PPAR-α can enhance the body's sensitivity to insulin,
inhibit the production of inflammatory factors and inflammation and
inhibit the oxidative stress response; and its decreased expression
can aggravate the inflammation and oxidative stress injury
(25). It has been reported in
literature (26) that CYP2E1 can
inhibit the expression of PPAR-α. Therefore, it is speculated that
EP can negatively regulate the CYP2E1-PPAR-α signaling pathway,
thereby alleviating the oxidative stress and inflammatory injury.
According to the experimental results (Figs. 3 and 4), the mRNA transcriptional level and
protein expression level of CYP2E1 in EP group were significantly
decreased compared with those in ALD group, but the mRNA
transcriptional level and protein expression level of PPAR-α were
significantly increased, which are consistent with our
speculation.
It has been reported that oxidative stress can
activate Nrf2, and the activation of Nrf2 can induce the VLDLR
overexpression in hepatocytes, leading to alcoholic liver injury
(6). Nrf2 gene silencing can inhibit
the upregulation of VLDLR in hepatocytes caused by oxidative stress
(27). Therefore, it is speculated
that one of the mechanisms by which EP protects ALD is that EP can
negatively regulate the Nrf2-VLDLR signaling pathway. The
experimental results (Figs. 3 and
4) showed that EP could downregulate
the mRNA and protein expression levels of Nrf2 and VLDLR in EP
group, which is consistent with our speculation. In conclusion, we
consider that the mechanism by which EP alleviates ALD involves its
anti-inflammatory and anti-oxidative effects; EP can negatively
regulate the CYP2E1-PPAR-α signaling pathway to inhibit the
oxidative stress response, thus negatively regulating the
Nrf2-VLDLR signaling pathway to alleviate the severity of ALD.
In conclusion, the experimental results showed that
EP can increase the levels of anti-inflammatory factors and
decrease the levels of pro-inflammatory factors, enhance the SOD
activity and decrease the FFA and TG contents through
anti-inflammatory and anti-oxidative effects; moreover, it can
upregulate the PPAR-α expression by negative regulation of
CYP2E1-PPAR-α signaling pathway and downregulate the Nrf2
expression by negative regulation of Nrf2-VLDLR signaling pathway,
thus alleviating ALD. This may be the new mechanism of the many
plant ingredients alleviating ALD, which needs further experimental
study. This finding, with important clinical significance, provides
a new direction for the treatment and prevention of ALD.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FS designed the study, ZW was involved in the animal
model establishment, WL and YL performed data analysis. FS was a
major contributor in writing the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Animal Ethics
Committee of Institute of Materia Medica of Shandong Academy of
Medical Sciences (Jinan, China).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yin HQ, Choi YJ, Kim YC, Sohn DH, Ryu SY
and Lee BH: Salvia miltiorrhiza Bunge and its active
component cryptotanshinone protects primary cultured rat
hepatocytes from acute ethanol-induced cytotoxicity and fatty
infiltration. Food Chem Toxicol. 47:98–103. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cioboată R, Găman A, Traşcă D, Ungureanu
A, Docea AO, Tomescu P, Gherghina F, Arsene AL, Badiu C, Tsatsakis
AM, et al: Pharmacological management of non-alcoholic fatty liver
disease: Atorvastatin versus pentoxifylline. Exp Ther Med.
13:2375–2381. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tilg H, Moschen AR and Kaneider NC:
Pathways of liver injury in alcoholic liver disease. J Hepatol.
55:1159–1161. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhong W, Zhao Y, Tang Y, Wei X, Shi X, Sun
W, Sun X, Yin X, Sun X, Kim S, et al: Chronic alcohol exposure
stimulates adipose tissue lipolysis in mice: Role of reverse
triglyceride transport in the pathogenesis of alcoholic steatosis.
Am J Pathol. 180:998–1007. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gurel H, Genc H, Celebi G, Sertoglu E,
Cicek AF, Kayadibi H, Ercin CN and Dogru T: Plasma pentraxin-3 is
associated with endothelial dysfunction in non-alcoholic fatty
liver disease. Eur Rev Med Pharmacol Sci. 20:4305–4312.
2016.PubMed/NCBI
|
|
6
|
Wang Z, Dou X, Li S, Zhang X, Sun X, Zhou
Z and Song Z: Nuclear factor (erythroid-derived 2)-like 2
activation-induced hepatic very-low-density lipoprotein receptor
overexpression in response to oxidative stress contributes to
alcoholic liver disease in mice. Hepatology. 59:1381–1392. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jo H, Choe SS, Shin KC, Jang H, Lee JH,
Seong JK, Back SH and Kim JB: Endoplasmic reticulum stress induces
hepatic steatosis via increased expression of the hepatic very
low-density lipoprotein receptor. Hepatology. 57:1366–1377. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fawcett TW, Martindale JL, Guyton KZ, Hai
T and Holbrook NJ: Complexes containing activating transcription
factor (ATF)/cAMP-responsive-element-binding protein (CREB)
interact with the CCAAT/enhancer-binding protein (C/EBP)-ATF
composite site to regulate Gadd153 expression during the stress
response. Biochem J. 339:135–141. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Harding HP, Zhang Y, Bertolotti A, Zeng H
and Ron D: Perk is essential for translational regulation and cell
survival during the unfolded protein response. Mol Cell. 5:897–904.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lieber CS and DeCarli LM: Effects of
mineral and vitamin supplementation on the alcohol-induced fatty
liver and microsomal induction. Alcohol Clin Exp Res. 13:142–143.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang X and Cederbaum AI: Acute ethanol
pretreatment increases FAS-mediated liver injury in mice: Role of
oxidative stress and CYP2E1-dependent and -independent pathways.
Free Radic Biol Med. 42:971–984. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kozutsumi Y, Segal M, Normington K,
Gething MJ and Sambrook J: The presence of malfolded proteins in
the endoplasmic reticulum signals the induction of
glucose-regulated proteins. Nature. 332:462–464. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang X, Wang Z, Li J, Gu D, Li S, Shen C
and Song Z: Increased 4-hydroxynonenal formation contributes to
obesity-related lipolytic activation in adipocytes. PLoS One.
8:e706632013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tsung A, Kaizu T, Nakao A, Shao L, Bucher
B, Fink MP, Murase N and Geller DA: Ethyl pyruvate ameliorates
liver ischemia-reperfusion injury by decreasing hepatic necrosis
and apoptosis. Transplantation. 79:196–204. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cabezas Castro M, Erkelens DW and van Dijk
H: Free fatty acids: Mediators of insulin resistance and
atherosclerosis. Ned Tijdschr Geneeskd. 146:103–109. 2002.(In
Dutch). PubMed/NCBI
|
|
16
|
Yellen P and Foster DA: Inhibition of
fatty acid synthase induces pro-survival Akt and ERK signaling in
K-Ras-driven cancer cells. Cancer Lett. 353:258–263. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Boden G, She P, Mozzoli M, Cheung P,
Gumireddy K, Reddy P, Xiang X, Luo Z and Ruderman N: Free fatty
acids produce insulin resistance and activate the proinflammatory
nuclear factor-kappaB pathway in rat liver. Diabetes. 54:3458–3465.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chang BH, Li L, Paul A, Taniguchi S,
Nannegari V, Heird WC and Chan L: Protection against fatty liver
but normal adipogenesis in mice lacking adipose
differentiation-related protein. Mol Cell Biol. 26:1063–1076. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Samarghandian S, Azimi-Nezhad M, Borji A,
Samini M and Farkhondeh T: Protective effects of carnosol against
oxidative stress induced brain damage by chronic stress in rats.
BMC Complement Altern Med. 17:2492017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Collins AR: Oxidative DNA damage,
antioxidants, and cancer. BioEssays. 21:238–246. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiong Y, Ye T, Wang M, Xia Y, Wang N, Song
X, Wang F, Liu L, Zhu Y, Yang F, et al: A novel cinnamide YLT26
induces breast cancer cells apoptosis via ROS-mitochondrial
apoptotic pathway in vitro and inhibits lung metastasis in vivo.
Cell Physiol Biochem. 34:1863–1876. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang R, Han X, Delude RL and Fink MP:
Ethyl pyruvate ameliorates acute alcohol-induced liver injury and
inflammation in mice. J Lab Clin Med. 142:322–331. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Naour S, Espinoza BM, Aedo JE, Zuloaga R,
Maldonado J, Bastias-Molina M, Silva H, Meneses C,
Gallardo-Escarate C, Molina A, et al: Transcriptomic analysis of
the hepatic response to stress in the red cusk-eel (Genypterus
chilensis): Insights into lipid metabolism, oxidative stress
and liver steatosis. PLoS One. 12:e01764472017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ramadori P, Drescher H, Erschfeld S,
Schumacher F, Berger C, Fragoulis A, Schenkel J, Kensler TW, Wruck
CJ, Trautwein C, et al: Hepatocyte-specific Keap1 deletion reduces
liver steatosis but not inflammation during non-alcoholic
steatohepatitis development. Free Radic Biol Med. 91:114–126. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun X, Yamasaki M, Katsube T and Shiwaku
K: Effects of quercetin derivatives from mulberry leaves: Improved
gene expression related hepatic lipid and glucose metabolism in
short-term high-fat fed mice. Nutr Res Pract. 9:137–143. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hamed AM, El-Kharashi OA, Boctor SS and
Abd-Elaziz LF: Potential involvement of PPAR α activation in
diminishing the hepatoprotective effect of fenofibrate in NAFLD:
Accuracy of non-invasive panel in determining the stage of liver
fibrosis in rats. Biomed Pharmacother. 85:68–78. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qiu P, Dong Y, Li B, Kang XJ, Gu C, Zhu T,
Luo YY, Pang MX, Du WF and Ge WH: Dihydromyricetin modulates p62
and autophagy crosstalk with the Keap-1/Nrf2 pathway to alleviate
ethanol-induced hepatic injury. Toxicol Lett. 274:31–41. 2017.
View Article : Google Scholar : PubMed/NCBI
|