Introduction
Carcinogenesis is a process where potential
alterations in oncogenes, proto-oncogenes, conductive genes and
loss in tumor suppressor genes accumulate over time (1). Nevertheless, recent evidence has shown
that changes in the metabolism of cancer cells might be an
important factor responsible for neoplastic development. In the
Hanahan and Weinberg (2) model, the
authors propose that changes in metabolism and in cellular
microenvironment are important modulators of a tumor's sustainment
and progression. Changes in the tissue microenvironment including
lack of nutrients, hypoxia, and acidosis activate intracellular
signaling pathways leading to a selective cell physiologic program
to sustain adaptation to a new microenvironment (3). Also, recent reports have been
identified that acidity modulates significantly the generation of
metastasis (4). However, there is
still a lack of knowledge regarding the potential role of hypoxia
and acidity on intracellular transcriptomic phenotype.
During tumor development, the Hydrogen potential
(pH) plays a potential role in the cellular division, the normal
physiological pH it is estimated at a range between of 7.1–7.2
(1). In this context, when tumor
development occurs, this pH decreases; acidifying the
microenvironment intra or extracellularly, an acid microenvironment
is favored, promoting a metastatic cell phenotype (5,6).
The normal atmospheric level of oxygen is 21%,
(normoxia) (7), hypoxia occurs when
there is a decrease in oxygen tension below the critical level (1
to 2%) (8). Cellular hypoxia can
occur in both physiological conditions and in different
pathological conditions (9), and
cells adapt immediately by inducing the hypoxia-inducible factor 1α
(HIF1α) (10). The
heterodimer HIF1α is composed of alpha and beta subunits. In
normoxia conditions, HIF1α is constitutively expressed and
degraded in a short period of time (11–14).
Under a hypoxic scenario HIF1α regulates cellular
homeostasis and systemic response by positive or negatively
modulating the transcriptional activation of selective genes,
including those involved in energetic metabolism [glucose
transporter 1 (GLUT1)], angiogenesis [vascular endothelial
growth factor (VEGF)], survival [erythropoietin
(EPO)], pH maintaining [carbonic anhydrase 9 (CA9)],
cell death (CASP3 and CASP7), and others that
facilitate oxygen supply or metabolic adaptation to hypoxia
(11).
To address the effect of hypoxia on cell physiology,
two classical cellular micro-environmental models have been carried
out. The first model uses a hypoxia sealed chamber containing a
mixture of gases where oxygen is at 1 to 2%. The second one uses a
chemical agent, which is highly reactive with oxygen, for example,
iron (deferoxamine); flavonoids, for example, quercetin; an
inhibitor dependent of 2-oxoglutarate oxygenase; and transition
metals, like cobalt (15). In fact,
chemical induction with cobalt chloride (CoCl2) is the
chemical model most frequently used (16). The gas chamber and chemical models
have been used interchangeably to evaluate the effect of hypoxia in
different cellular environments. In general, we sought to identify
networks of selective metabolic genes activated by both hypoxic
models, cobalt chloride and chamber sealed by hypoxia, using
microarray analysis. We hypothesized that chemical or cellular
hypoxia could regulate time-dependent genes involved in cell
viability, apoptosis, and cell proliferation.
Materials and methods
Cell culture and maintenance
We used the cell line Caco-2 [adenocarcinoma
colorectal; American Type Culture Collection (ATCC), Rockville, MD,
USA] in all assays. Caco-2 cell line was cultivated in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum,
1% penicillin-streptomycin, and was maintained at 37°C, with 5%
CO2.
Gas chamber with induced hypoxia
A total of 150,000 Caco-2 cells were seeded in each
well, incubated for 24 h to allow their adhesion. The cultures were
then placed into the gas chamber (Stemcell Technologies, Inc.,
Vancouver, BC, Canada). The chamber was closed and the gas mixture
(1% O2, 5% CO2 and 94% N2) was
injected for 5 min (at a controlled pressure of 2 psi) to ensure
equilibrium of the oxygen concentration in the cell medium, after
which the chamber was sealed and finally placed in the incubator at
37°C. Hypoxic treatment was maintained for 0, 3, 6, 24, 48 and 72
h.
Chemical hypoxia induction with cobalt
chloride (CoCl2)
For chemical hypoxia 150,000 Caco-2 cells were
seeded in each well in the 6-well plates, incubated for 24 h to
allow their adhesion and 100 µM CoCl2 (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) were added to the medium and were
incubated for 0, 3, 6, 24, 48 and 72 h.
Acidosis assay
To simulate acidosis in our hypoxic model, the pH of
the medium was determined by adding 25 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) or 25 mM
3-(N-morpholino) propanesulfonic acid (MOPS). Subsequently,
the pH was measured using a SevenMulti standard potentiometer
(Mettler Toledo, Greifensee, Switzerland), initially calibrated
with reference buffer solutions (Hanna Instruments, Woonsocket, RI,
USA), and was finally adjusted to 6.5, 6.7 or 6.9 with 1 M HCl.
Cell viability assay
Once pH and hypoxia conditions were selected, cell
viability in the Caco-2 cell line was evaluated, with the
chemiluminescent assay using the Cell Titer Glo Luminescent kit
(Promega Corporation, Madison, WI, USA). Caco-2 cells were seeded
in 96-well plates in a density of 15,000 cells/well and were
cultivated for 24 h at 37°C in 5% CO2 to allow their
adhesion. They were then exposed to pH and hypoxia protocols during
0, 24, 48 and 72 h and the cell viability was measured. This method
is based on the quantification of ATP in the culture, and the
levels are directly proportional to the presence of metabolically
active cells.
Apoptosis chemiluminescent assay
Apoptosis was determined using the
Caspase-GloR 3/7 Assay Protocol kit (Promega
Corporation). This assay measured caspase 3/7 activity and the
luminescent signal that are directly proportional to the apoptosis
level in the evaluated cells.
RNA extraction, reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
To evaluate the effect of cellular stress by hypoxia
and acidity on cell metabolism, we carried out a Real-Time mRNA
expression analysis of a panel of genes involved in
hypoxia-metabolism: HIF1α, EPO, GLUT1, VEGF, LDH and
CA9 by qPCR and SYBR-Green using GAPDH as gene
normalizer. We previously performed an assay for the selection of
the endogenous gene (data not shown) was made by the comparison of
GAPDH, GPI and EMC7 using a gene stability assay
(17). Our results showed that
compared to the GPI and EMC7 genes, the GAPDH
gene was the most stable gene. Cultures were maintained under the
hypoxia and acid protocols and total RNA was isolated using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), following
instructions from the fabricators, followed by a treatment with
DNAse (Qiagen, Inc., Valencia, CA, USA). RNA integrity and
concentration was evaluated by spectrophotometry with Nanodrop 8000
(Thermo Fisher Scientific, Inc.).
cDNA was synthesized from 1 µg of total RNA using
High-Capacity cDNA Reverse Transcription kit (Applied Biosystems;
Thermo Fisher Scientific, Inc.), following the fabricator's
instructions. The primers for the metabolic genes and the
constitutive gene were designed in accordance with the sequences
from the database Gen Bank (Table
I). The real-time PCR thermal cycler LightCycler 480 (Roche
Diagnostics, Indianapolis, IN, USA) and the reagent SYBR-Green I
Master (Roche Diagnostics) were used. All the qPCR reactions were
triplicated and three independent assays were carried out. The
expression analysis was carried out by the relative
2−ΔΔCq quantification method (18).
 | Table I.Primer sequence used in the
quantification of the expression of genes sensible to stress. |
Table I.
Primer sequence used in the
quantification of the expression of genes sensible to stress.
| A, Hypoxia
sensitive genes |
|---|
|
|---|
| Gene | Sequence |
|---|
| HIF1α |
|
| F |
ATCCATGTGACCATGAGGAAATG |
| R |
TCGGCTAGTTAGGGTACACTTC |
| EPO |
|
| F |
GGAGGCCGAGAATATCACGAC |
| R |
CCCTGCCAGACTTCTACGG |
| CA9 |
|
| F |
GGATCTACCTACTGTTGAGGCT |
| R |
CATAGCGCCAATGACTCTGGT |
| VEGF |
|
| F |
AGGGCAGAATCATCACGAAGT |
| R |
AGGGTCTCGATTGGATGGCA |
| GLUT1 |
|
| F |
TCTGGCATCAACGCTGTCTTC |
| R |
CGATACCGGAGCCAATGGT |
|
| B, Endogenous
gene |
|
| Gene | Sequence |
|
| GAPDH |
|
| F |
ACAACTTTGGTATCGTGGAAGG |
| R |
GCCATCACGCCACAGTTTC |
Microarrays analysis (expression
profiles)
The RNA, sample processing, microarray
hybridization, and gene expression analysis were conducted using
the GeneChip 3′ IVT Express kit (Affymetrix; Thermo Fisher
Scientific, Inc.). The hybridization mixture was prepared and
applied to the GeneChip Human Genome U133 Plus 2.0 Array
(Affymetrix; Thermo Fisher Scientific, Inc.), measuring >43,000
transcripts representing >20,000 human genes. Washing and
scanning processes were realized in the Fluidics Station 400 and
GeneChip Scanner 3000 7G, respectively, and preliminary data
analysis was completed using Microarray Suite software version
5.0.0.032. Normalization was performed using robust microarray
analysis and quantile normalization.
Statistical analysis
Data were compared using a two-way ANOVA (treatment
and time) and Dunnet's test was performed to compare viability and
apoptosis in all trials. The variables were previously normalized
(Kolmogorov-Smirnov). The analysis was performed using SPSS
software v20.0 (SPSS, Inc., Chicago, IL, USA). All graphs were done
with GraphPad Prism v6.0 (GraphPad Software, Inc., La Jolla, CA,
USA). All assays were performed in triplicate. Results are
expressed as the mean ± standard deviation.
Results
Viability assays in chemical and
sealed gas chamber hypoxia models
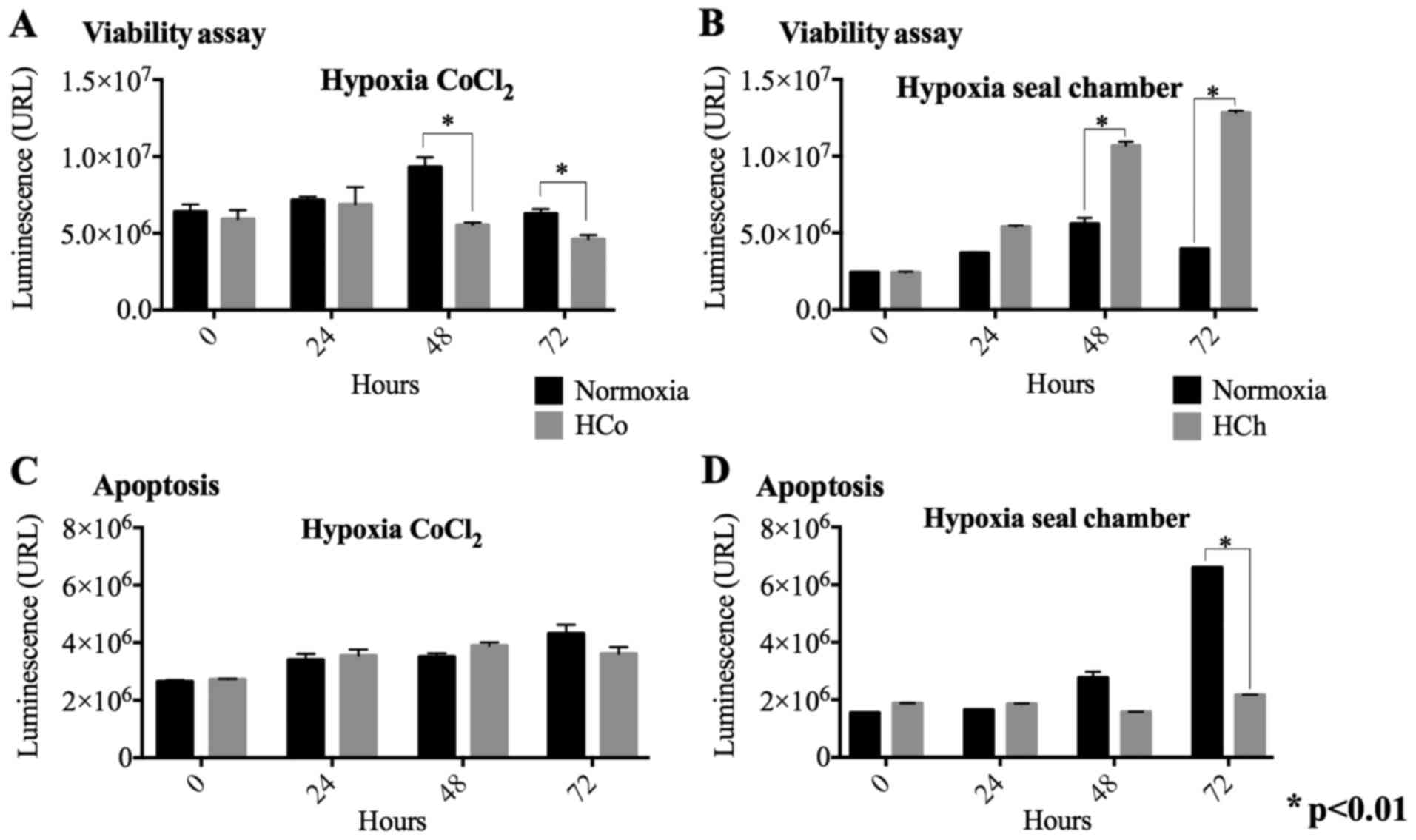
To determine the effect of hypoxia on cell
viability, cells treated with CoCl2 or maintained in the
gas chamber were quantified as described. When comparing viability
based on time and treatment, a highly significant difference was
found in time (F=15.396, P<0.01) and for the treatment
(F=17.288, P<0.01). We found a significant decrease in cell
viability in cells treated with CoCl2 at 48 h and 72 h
compared to the control without chemical hypoxia (Fig. 1A). In contrast, hypoxia-induced in
sealed gas chamber shows a substantial increase in cell viability
at a similar time frame to chemical hypoxia, 48 and 72 h (Fig. 1B).
Apoptosis evaluation in chemical and
sealed gas chamber hypoxia models
The caspase 3/7 activity ratio was measured to
evaluate apoptosis in both hypoxia models. The results of apoptosis
displayed significant changes between the control at time 72 h
(F=964.923, P<0.01) and for the treatment of hypoxia chamber
(F=1,154.081, P<0.01). It was observed that chemically induced
hypoxia does not promote apoptotic activation when compared to the
normoxia control in all the times (Fig.
1C). We did find that hypoxia induced by the gas chamber
decreased apoptosis significantly at 72 h compared to the normal
control (Fig. 1D). This data
suggests that a hypoxic environment induced by gas chamber
incubation may have a severe effect on oxygen depletion leading to
cell death and does not favor proliferation.
Evaluation of the effect of pH and pH+
hypoxia on viability and apoptosis in both models of hypoxia
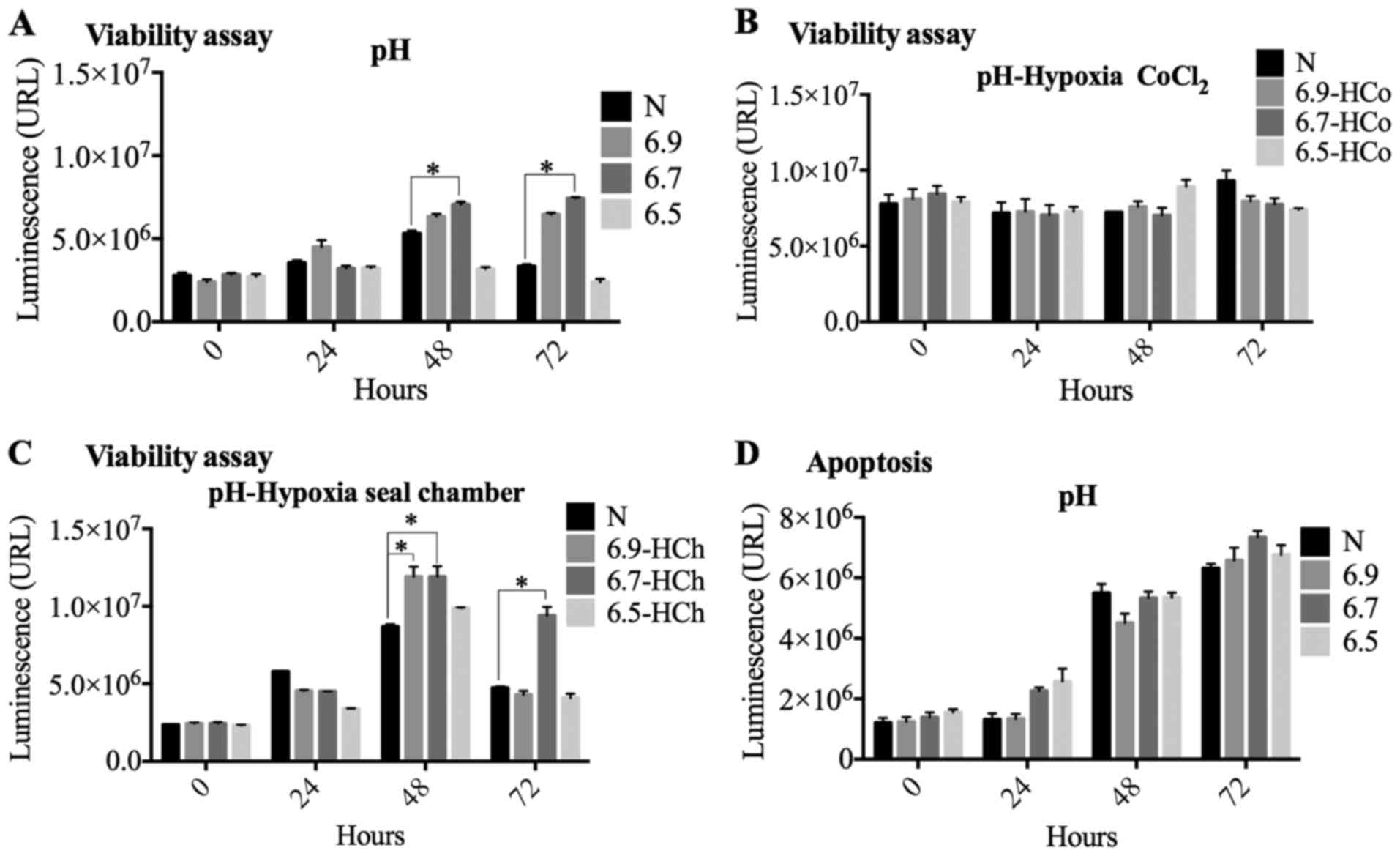
We evaluated the acidity effect in the chemical and
sealed gas chamber hypoxia models by setting three pH values: 6.5,
6.7 and 6.9. An increase in cell viability was observed at 48 and
72 h when the cells were exposed to the most acidic pH (Ph 6.7 and
6.9), compared against normoxia conditions (Fig. 2A and B). We also found that viability
and apoptosis showed time-dependent significant changes (F=18.759,
F=55.840, P<0.01), whereas at pH 6.5 there was a decrease in
cell proliferation when compared to normoxia. This indicates that
pH 6.5 significantly affects cell viability (Fig. 2A). We also found that changes in pH
do not affect the number of apoptotic cells at 24, 48 and 72 h when
compared to normoxia (Fig. 2B).
Next, we determined cell viability in the
pH+chemical hypoxia model. We found no changes at any of the three
pH values analyzed (Fig. 2C).
Finally, we observed that the pH+sealed chamber hypoxia model
promotes an increase in viability at pH 6.9 and 6.7 when compared
to normoxia at 48 h (Fig. 2D). After
72 h, we still found an increase in cell viability at pH 6.7 and no
changes at pH 6.9 and 6.5 (Fig. 2D).
These results reproduce, at least in part; those observed in the
independent measurements of hypoxia or acidity and support the
effect of pH on cell viability during hypoxia.
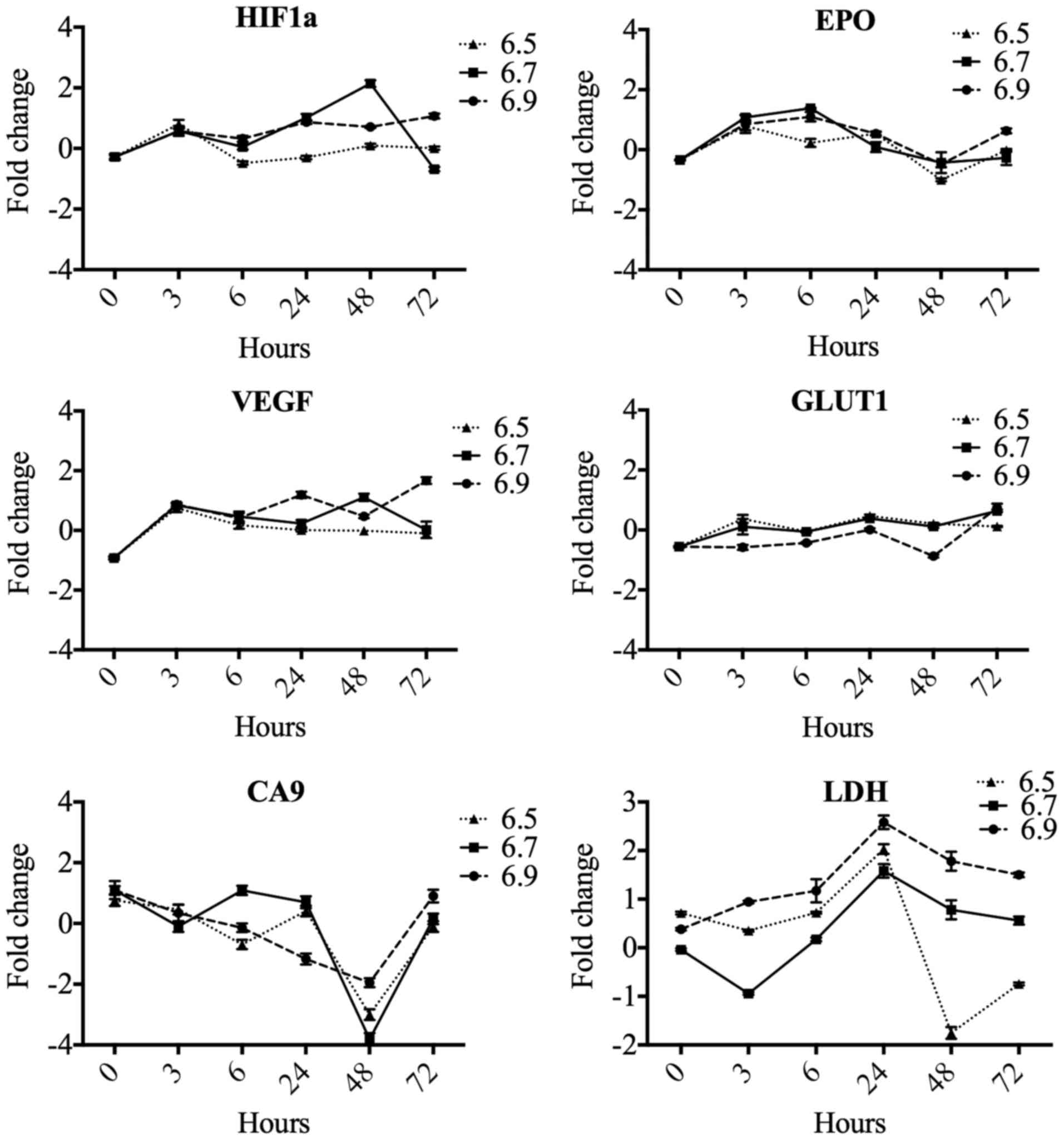
Effect of pH on the expression of
hypoxia response genes
To implement the standardization of the hypoxia
model, we initially evaluated the effect of pH on six genes related
to hypoxia response: HIF1α, EPO, VEGF, GLUT1, CA9, and
LDH. These results suggest that pH changes are not involved
as genetic modulators of HIF1α, EPO, VEGF, GLUT1, CA9, and
LDH (Fig. 3).
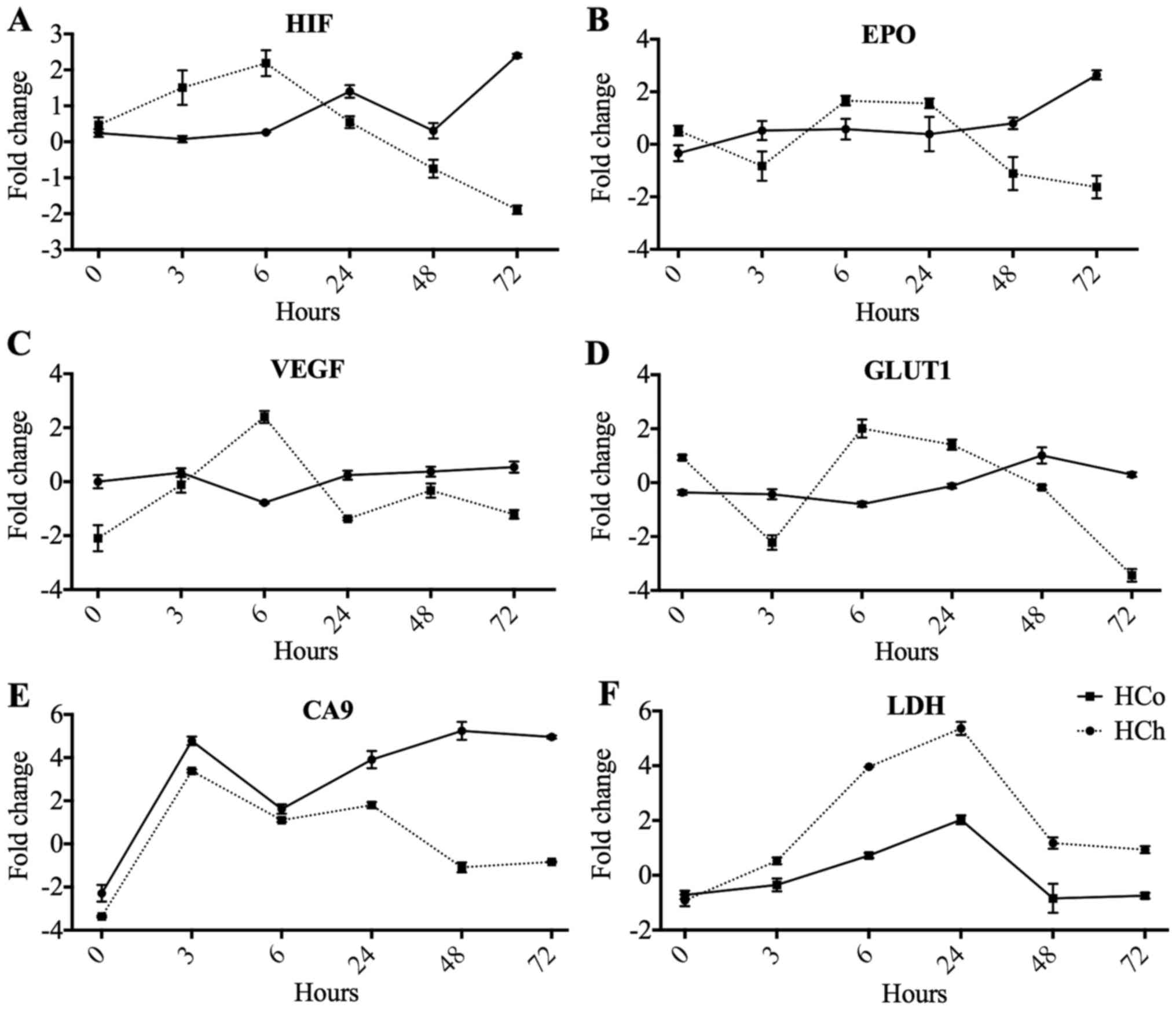
Gene expression comparison in the two
models of hypoxia
We evaluated the expression of the same six genes in
two hypoxia models. When we compared the two models we observed
that the hypoxia induced by CoCl2 promotes an acute time
specific increase in gene expression at 6 h and later showed a
decrease to control values (Fig. 4).
Observations of the hypoxia-induced sealed gas chamber show a
stable effect on gene expression displaying a lower, gradual and
maintained elevation. At 72 h, the effect is maintained and
HIF1α, EPO, and CA9 are still overexpressed. Finally,
the gene GLUT1 stands out in that it only maintains basal
levels of expression in both models (Fig. 4). At this stage, our results suggest
that chemical hypoxia modulates gene expression profile at 6 h
whereas hypoxia-dependent chamber leads its major effects at 72
h.
Finally, we found an early increase in HIF1α
expression that peaked at 6 h following a decrease later on
(Fig. 4A). At 6 h, we also
identified an increase of EPO, VEGF, GLUT1 and LDH
genes (Fig. 4B-D and F). No
significant changes were found in CA9 (Fig. 4E).
Global gene expression profile in the
two hypoxia models using microarrays
We used microarray analysis to determine the effect
of global gene expression in the two hypoxia models: Chemical (6 h)
and sealed chamber (72 h) induced hypoxia. We identified a gene
expression profile for each model (Fig.
5A and B). The chemical hypoxia model promotes downregulation
of 3 genes (CYP1A1, CYP1B1, and CYCS) and the
overexpression of 27 genes: TXNIP, ALDH1L2, FAM169A, ZBTB20,
RORA, CREBRF, FBXL20, LOX, ZNF846, YPEL2, PPM1A, FAM225A, NGFR,
VLDLR-AS1, MT1M, ANKRD37, SPAG4, BNIP3L, ATP8B2, AKR1C1, SLC7A11,
AQP10, AKR1C1, KLHL24, DDIT4, PCK1, JMY. For the sealed gas
chamber model, we found the subexpression of 2 genes: UTS2
and MMP1 and the overexpression of 28 genes: LINC01293,
CASP5, RORA, SLC6A8, ZG16, GNRH2, CREBRF, FAM129A, ANKRD37, CLIC3,
ENO2, PCAT6, BHLHE40, BNIP3L, TMEM45A, TMEM27, DDIT4, FRMD3, CXCR4,
NRN1, ATP8B2, SPAG4, LGALS14, AQP10, LOX, BNIP3, PDK1, NDRG1.
We also observed that nine genes were shared between both models
(SPAG4, LOX, BNIP3L, ATP8B2, AQP10, RORA, DDIT4, CREBRF, and
ANKRD37) (Fig. 5C).
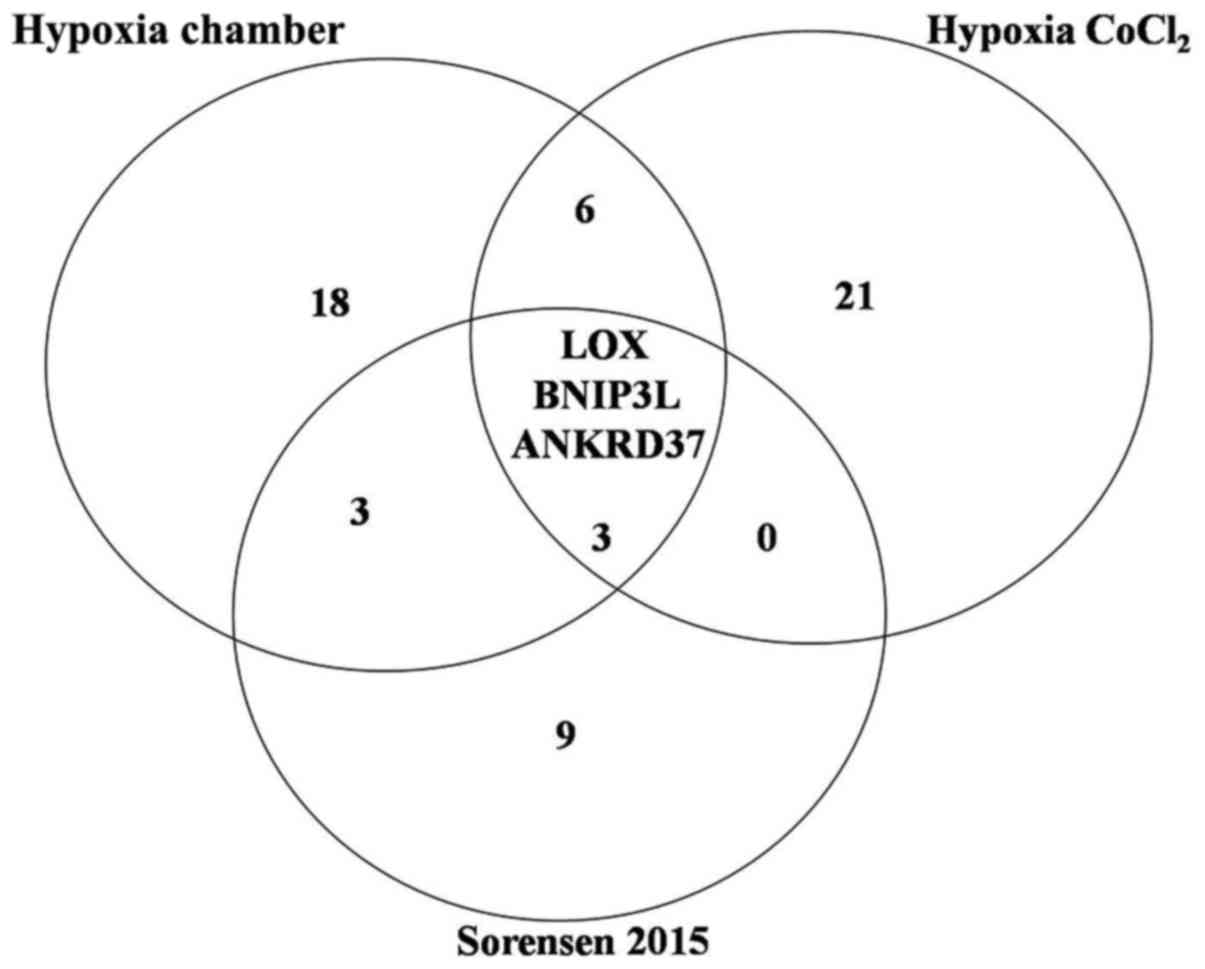
On the other hand, we compared the gene signatures
obtained from both models with the report of Sørensen et al
(19) that reported a signature of
15 genes sensitive to hypoxia, finding 3 common genes in all the
genetic signatures, suggesting that our data correlates with a
universal genetic signature for the hypoxia profile (Fig. 6).
Discussion
Experimental evidence has shown the effect of
hypoxia induction in vitro using the technical strategies
implemented in our study (20–23). In
general terms, one of the best candidates to identify the hypoxia
model is the evaluation of HIF1α expression. However, additional
potential candidates are emerging to validate sensor genes in
hypoxia and its metabolic effect, but they are not yet validated
(13,24,25). In
this study, we evaluated five target genes of HIF1α along with
parameters of viability and apoptosis in two models of hypoxia,
widely cited in the literature and with the potential to be
interchangeable. We found that both models show similarities in the
final induction of hypoxia and show no significant differences.
However, we identified selective time-dependent changes
particularly related to the induction of HIF1α as a response to
hypoxia. In addition, when the expression of the rest of the genes
is analyzed, a variable expression is observed in the different
times evaluated. For example, at 6 h in the CoCl2 model,
we found the highest expression of all the genes evaluated,
followed by a low expression and tendency to reach normal values in
later times, such as 48 and 72 h. In contrast, the gas chamber
model showed increased and gradual gene expression, which was
maintained for up to 72 h. This indicates that in the chemical
model, the cells normalize the system rapidly at 3 to 6 h and in
the hypoxia CoCl2 model the effect is rapidly reversed.
On the other hand, in the gas chamber model, the cells slowly
adapted to the change in a time-dependent manner and the effect was
observed and maintained in response to hypoxia.
We propose that these observations must be
considered when choosing one of this hypoxia models in order to
define if the effect that is desired to quantify is analyzed from 3
to 6 h or even a more prolonged time, as was proposed recently
(26).
One of the major achievements of our study is that
we show, for the first time, a comparison in both chemical and
chamber hypoxia models in different scenarios and identify a
selective expression of the genes induced by HIF1α. Our
results also show that the effect of hypoxia induction is time
sensitive and selective for each model. For example, a study
reported by Danli Wu in 2011 using both hypoxia models quantifies
HIF1α as the only hypoxia marker in this scenario (16).
Potentials pathways mainly affected by hypoxia are
glycolysis and oxidative phosphorylation (27). In this scenario cell viability assays
may have technical limitations when the main goal is to quantify
cell viability (28–30). For instance, no changes in oxidative
phosphorylation and cellular ATP levels have been reported in in
vitro trials trying to simulate conditions of lack of nutrients
and hypoxia models (28–30). In our project we determined cell
viability by using ATP quantification as a reliable marker of cell
viability. However, we also agree that ATP levels can be altered by
hypoxic conditions and this measurement might not be an indicator
of cell viability so metabolic activity marker in cells.
Despite that our two hypoxia models show a selective
time leading to HIF1α upregulation, we found 9 common
hypoxia-inducible genes between both models, as observed in the
Venn diagram (Fig. 5C). Our data
correlate with previous results showing that hypoxia regulates
similar metabolic pathways such as: Glycolysis, pyruvate
metabolism, ATP synthesis, regulation of processes (catalytic
activity, hydrolases, oxidoreductases) and other cellular processes
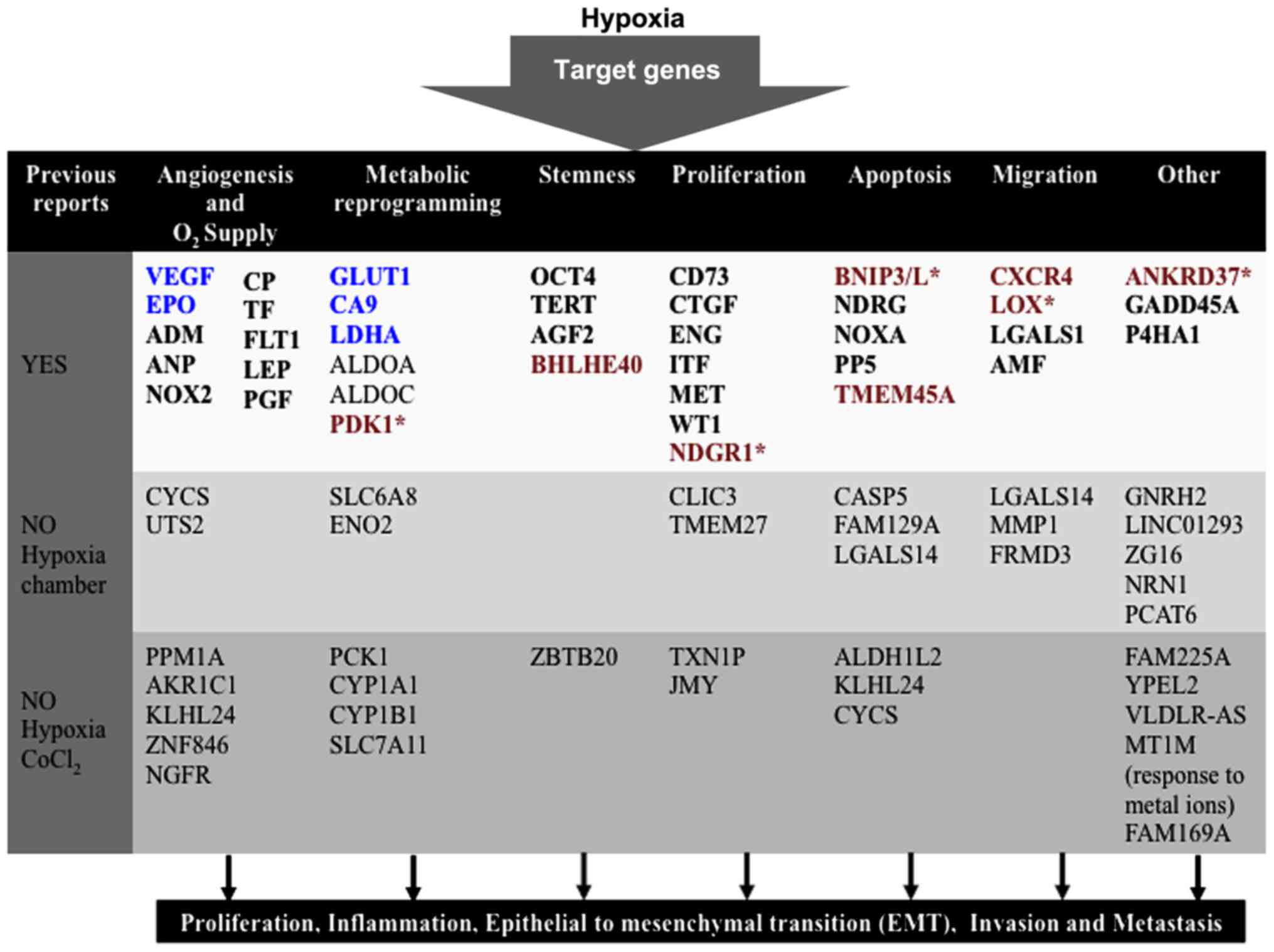
like adhesion or transport (Fig. 7)
(23,24). In particular, Sørensen et al
(19) in 2015 reported a 15
hypoxia-regulated genes signature as a universal hypoxia profile in
various cancer cell types. Our data correlate with the LOX,
ANKRD37, BNIP3L genes (Fig. 6),
previously reported by Sørensen et al (19). This indicates that these three genes
are highly regulated by the effect of hypoxia regardless of the
hypoxia model induced with. At this stage, we propose that LOX,
ANKRD37, BNIP3L genes might potentially be added as universal
hypoxia genes along with HIF1α, EPO, VEGF, GLUT1, CA9
and LDH previously characterized.
In summary, in this study, we show that the effect
of hypoxia is time-dependent regarding the hypoxia model used.
Chemical induction simulates an acute hypoxia whereas chamber of
gases maintains the effect of hypoxia in extended periods. These
two hypoxia models share 9 susceptible genes and when comparing the
two models of hypoxia with that reported by Sørensen et al
(19), we found 3 genes (LOX,
ANKRD37, BNIP3L) that could be considered as universal hypoxia
genes along with HIF1α, EPO, VEGF, GLUT1, CA9, and
LDH.
Our study opens a new research area showing the
significant value of identifying potential oxygen sensors and key
genes involved in metabolism and stress conditions during hypoxia.
Finally, it might be required to study the expression and
methylation of these genes to amplify the knowledge in this area
and to better evaluate the effect of hypoxia in the mechanisms of
cancer progression and metabolism of tumor cells.
References
|
1
|
Gatenby RA, Smallbone K, Maini PK, Rose F,
Averill J, Nagle RB, Worrall L and Gillies RJ: Cellular adaptations
to hypoxia and acidosis during somatic evolution of breast cancer.
Br J Cancer. 97:646–653. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Joyce JA and Pollard JW:
Microenvironmental regulation of metastasis. Nat Rev Cancer.
9:239–252. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Martínez-Zaguilán R, Seftor EA, Seftor RE,
Chu YW, Gillies RJ and Hendrix MJ: Acidic pH enhances the invasive
behavior of human melanoma cells. Clin Exp Metastasis. 14:176–186.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Griffiths JR: Are cancer cells acidic? Br
J Cancer. 64:425–427. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chiche J, Brahimi-Horn MC and Pouysségur
J: Tumour hypoxia induces a metabolic shift causing acidosis: A
common feature in cancer. J Cell Mol Med. 14:771–794. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Abaci HE, Truitt R, Luong E, Drazer G and
Gerecht S: Adaptation to oxygen deprivation in cultures of human
pluripotent stem cells, endothelial progenitor cells and umbilical
vein endothelial cells. Am J Physiol Cell Physiol. 298:C1527–C1537.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Solaini G, Baracca A, Lenaz G and Sgarbi
G: Hypoxia and mitochondrial oxidative metabolism. Biochim Biophys
Acta. 1797:1171–1177. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dayan F, Mazure NM, Brahimi-Horn MC and
Pouysségur J: A dialogue between the hypoxia-inducible factor and
the tumor microenvironment. Cancer Microenviron. 1:53–68. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ke Q and Costa M: Hypoxia-inducible
factor-1 (HIF-1). Mol Pharmacol. 70:1469–1480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weidemann A and Johnson RS: Biology of
HIF-1alpha. Cell Death Differ. 15:621–627. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jewell UR, Kvietikova I, Scheid A, Bauer
C, Wenger RH and Gassmann M: Induction of HIF-1alpha in response to
hypoxia is instantaneous. FASEB J. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Messineo S1, Laria AE1, Arcidiacono B1,
Chiefari E1, Huertas Luque RM, Foti DP and Brunetti A: Cooperation
between HMGA1 and HIF-1 contributes to hypoxia-induced VEGF and
visfatin gene expression in 3T3-L1 adipocytes. Front Endocrinol
(Lausanne). 7:732016.PubMed/NCBI
|
|
14
|
Höpfl G, Ogunshola O and Gassmann M: HIFs
and tumors-causes and consequences. Am J Physiol Regu.
286:R608–R623. 2004.
|
|
15
|
Masoud GN and Li W: HIF-1ü pathway: Role,
regulation and intervention for cancer therapy. Acta Pharm Sin B.
5:378–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu D and Yotnda P: Induction and testing
of hypoxia in cell culture. J Vis Exp. 12:28992011.
|
|
17
|
Eisenberg E and Levanon EY: Human
housekeeping genes, revisited. Trends Genet. 29:569–574. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sørensen BS, Knudsen A, Wittrup CF,
Nielsen S, Aggerholm-Pedersen N, Busk M, Horsman M, Høyer M,
Bouchelouche PN, Overgaard J and Alsner J: The usability of a
15-gene hypoxia classifier as a universal hypoxia profile in
various cancer cell types. Radiother Oncol. 116:346–351. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wenger RH, Kurtcuoglu V, Scholz CC, Marti
HH and Hoogewijs D: Frequently asked questions in hypoxia research.
Hypoxia (Auckl). 3:35–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bensaad K, Favaro E, Lewis CA, Peck B,
Lord S, Collins JM, Pinnick KE, Wigfield S, Buffa FM, Li JL, et al:
Fatty acid uptake and lipid storage induced by HIF-1alpha
contribute to cell growth and survival after hypoxia-reoxygenation.
Cell Rep. 9:349–365. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang YB, Wang X, Meister EA, Gong KR, Yan
SC, Lu GW, Ji XM, Shao G, et al: The effects of CoCl2 on HIF-1α
protein under experimental conditions of autoprogressive hypoxia
using mouse models. Int J Mol Sci. 15:10999–11012. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chiche J, Ilc K, Laferrière J, Trottier E,
Dayan F, Mazure NM, Brahimi-Horn MC and Pouysségur J:
Hypoxia-inducible carbonic anhydrase IX and XII promote tumor cell
growth by counteracting acidosis through the regulation of the
intracellular pH. Cancer Res. 69:358–368. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zuo J, Wen J, Lei M, Wen M, Li S, Lv X,
Luo Z and Wen G: Hypoxia promotes the invasion and metastasis of
laryngeal cancer cells via EMT. Med Oncol. 33:152016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guimarães TA, Farias LC, Santos ES, de
Carvalho Fraga CA, Orsini LA, de Freitas Teles L, Feltenberger JD,
de Jesus SF, de Souza MG, Santos SH, et al: Metformin increases PDH
and suppresses HIF-1α under hypoxic conditions and induces cell
death in oral squamous cell carcinoma. Oncotarget. 7:55057–55068.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zeng HL, Zhong Q, Qin YL, Bu QQ, Han XA,
Jia HT and Liu HW: Hypoxia-mimetic agents inhibit proliferation and
alter the morphology of human umbilical cord-derived mesenchymal
stem cells. BMC Cell Biol. 12:322011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li J, Zhang D, Ward KM, Prendergast GC and
Ayene IS: Hydroxyethyl disulfide as an efficient metabolic assay
for cell viability in vitro. Toxicol In Vitro. 26:603–612. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Depoix C, Barret LA, Hubinont C and
Debieve F: Viability of primary term cytotrophoblast cell culture
in normoxia and hypoxia. Mol Hum Reprod. 19:29–34. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jose C, Bellance N and Rossignol R:
Choosing between glycolysis and oxidative phosphorylation: A
tumor's dilemma? Biochim Biophys Acta. 1807:552–561. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rodríguez-Enríquez S, Carreño-Fuentes L,
Gallardo-Pérez JC, Saavedra E, Quezada H, Vega A, Marín-Hernández
A, Olín-Sandoval V, Torres-Márquez ME and Moreno-Sánchez R:
Oxidative phosphorylation is impaired by prolonged hypoxia in
breast and possibly in cervix carcinoma. Int J Biochem Cell Biol.
42:1744–1751. 2010. View Article : Google Scholar : PubMed/NCBI
|