Introduction
Infectious respiratory diseases, including
mycoplasma pneumonia and influenza, present a serious threat to
public health (1,2). The nasal mucosa is the primary invasion
site for a number of pathogenic microorganisms and nasal
immunization has the potential to increase mucosal immunity
(3). The anatomy of the porcine
respiratory system is similar to that of humans (more so than mice)
and is considered to be a good animal model (4,5). The
tonsils are located near the nasal cavity, which is understood to
serve the same role as the Waldeyer's ring in humans (6). Waldeyer's ring is located at the
entrance to the digestive and respiratory tracts where it acts as a
key component in the mucosal-associated lymphoid system (7). B and T cells are located within the
nasal-associated lymphoid tissue and tonsils with
antigen-presenting cells (APCs), including dendritic cells (DCs),
interspersed among them (8,9). The nasal mucosa and tonsils are
considered to be a potential target for nasal vaccines as they
serve an important role in antigen recognition and immune
activation following intranasal administration, which is similar to
gut-associated lymphoid tissue in the gastrointestinal tract
(10,11).
There are high numbers of DCs distributed under the
nasal epithelium and, as the most ubiquitous professional APCs in
the nasal mucosa and tonsils, they serve an important role in
immune surveillance (12,13). DCs are able to present antigens and
activate T cell responses (14,15).
Probiotics are defined as live microorganisms that
confer a health benefit when administered at adequate
concentrations (16). It has
previously been suggested that probiotic bacteria interact with
epithelial and immune cells from the innate immune system to
reinforce the mucosal barrier (17).
Probiotic bacteria may be sampled by DCs as whole cells or their
antigenic fragments (17).
Bacillus subtilis is a common probiotic with good biological
safety that is able to effectively antagonize pathogenic bacteria
(18). It has been reported that
B. subtilis effectively enhances the innate immunity of the
intestinal tract; however, whether B. subtilis is able to
enhance the innate immunity of the nasal mucosa and tonsils remains
unknown (19). The spores of B.
subtilis have been successfully used for antigen delivery and
they may also be used as a mucosal adjuvant for the H5N1 vaccine,
in which they significantly enhance the effect of intranasal
immunization (20,21). The mechanism by which B.
subtilis spores act as a potential immune adjuvant and whether
this effect is associated with immune cells in the nasal mucosa and
tonsils require further investigation.
In the present study, pigs were selected as the
animal model to investigate the effects of B. subtilis on
immune cells in the nasal mucosa and tonsils. Changes to the immune
cells in the nasal mucosa and tonsils following intranasal
administration with B. subtilis were observed. The results
of the present study may be valuable for further study into an
intranasal vaccines with B. subtilis as a mucosal
adjuvant.
Materials and methods
Animals and reagents
A total of 6 (age, 1-month-old; weight, 13–16 kg)
and 8 (age, 1-day-old; weight, 1.10–1.30 kg) male cross-bred
Duroc/Landrace/Yorkshire piglets were obtained from Jiangsu Huai'an
Pig Farm (Huai'an, China). A total of 2 cross-bred
Landrace/Yorkshire sows (age, 1 year and 6 months; weight, 140–145
kg; Jiangsu Huai'an Pig Farm) were used to nurse the piglets. The
pigs and piglets were housed in Jiangsu Huai'an Pig Farm with a
constant humidity (60%) and temperature (26°C) at 12 h light/dark
cycle with free access to water and food. B. subtilis WB800
was obtained from the College of Plant Protection Nanjing
Agricultural University (Nanjing, China).
Intranasal administration
A total of 6 1-month-old male piglets were randomly
equally divided into two groups (n=3) as follows: The control group
was intranasally administrated with PBS (500 µl/nostril) and the
B. subtilis group was intranasally administrated with
Dylight 488 (Life Technologies; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA)-labeled B. subtilis WB800 spores
(108 cfu/nostril). The procedure was repeated after 1 h.
Piglets were euthanized by intravenous injection of sodium
pentobarbital (100 mg/kg) 2 h later as previously described
(22). All procedures were approved
by the Institutional Animal Care and Use Committee of Nanjing
Agricultural University (Nanjing, China) and followed the National
Institutes of Health guidelines for the performance of animal
experiments. Following sacrifice, the pigs were decapitated, the
lower jaw and skin were removed and the muscles around the nasal
cavity were removed. The nasal cavities, nasopharyngeal tonsils and
soft palate tonsils were removed, fixed in 4% paraformaldehyde at
room temperature for 6 h, embedded in optimal cutting temperature
compound (Sakura Finetek USA, Inc., Torrance, CA, USA) and cut into
8-µm-thick sections for immunofluorescence staining.
A total of 8 1-day-old male piglets were bred in a
pathogen free environment with the aforementioned conditions and
randomly divided into two groups (n=4): The control group and the
B. subtilis group. While the piglets were suckling, sows had
free access to water and feed (containing no antibiotics). No pigs
succumbed during the experiment. The B. subtilis group was
intranasally administrated with 200 µl/nostril B. subtilis
WB800 (1010 cfu/ml) on day 3, 500 µl/nostril on day 12
and 1 ml/nostril on day 21; these doses were selected from previous
studies (22–26). The control group was treated with the
same volumes of PBS. On day 28 the piglets were euthanized by
intravenous injection of sodium pentobarbital. The nasal cavities,
nasopharyngeal tonsils and soft palate tonsils were extracted and
fixed in Bouin's fluid (Nanjing SenBeiJia Biological Technology
Co., Ltd., Nanjing, China) for 48 h at 20–25°C for histological
analysis, or stored at −80°C for the detection of interleukin
(IL)-1β, tumor necrosis factor (TNF)-α, IL-8, IL-6, porcine
β-defensin 2 (pBD-2), toll-like receptor (TLR)-2, TLR-9 mRNA using
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR). The nasal cavities were subsequently decalcified by
dipping them in formic acid decalcifying solution (Nanjing
SenBeiJia Biological Technology, Co., Ltd.) for 1 week. The tissues
were embedded in paraffin, cut into 5-µm-thick sections, dewaxed in
xylene, rehydrated in a graded series of ethanol (100, 90, 80 and
70%, each for 1 min) and washed in PBS for immunohistochemistry,
immunofluorescence, and hematoxylin and eosin (H&E) staining.
The sections were immersed in hematoxylin for 30 sec, PBS for 5 min
and eosin for 5 sec. The sections were dehydrated through
increasing concentrations of ethanol (70, 80, 90 and 100%, each for
1 min) and xylene. All processes are performed at room
temperature.
Immunohistochemistry
The paraffin embedded sections were dewaxed in
xylene and rehydrated in a graded series of ethanol (100, 90, 80
and 70%, each for 1 min). Sections were subsequently poached in a
citrate buffer (pH 6) at 90–95°C for 15 min to retrieve antigens.
The sections were treated with 0.3% hydrogen peroxide at room
temperature for 15 min to quench endogenous peroxidase. The
sections were blocked with 5% normal goat serum, 5% bovine serum
albumin (BSA) and 5% normal rabbit serum (all Boster Biological
Technology, Pleasanton, CA, USA) for 30 min at room temperature.
The sections were subsequently incubated with primary antibodies
1–4 listed in Table I for 12 h at
4°C. Biotinylated secondary antibodies 1–4 listed in Table I were added to the sections for 1 h
at room temperature. Positive cells were visualized by treatment
with diaminobenzidine (DAB) for 60 min at room temperature, the
sections were then sealed with neutral balata. The negative control
was treated in the same way except the primary antibodies were
omitted. The sections were visualized using a light microscope
(Olympus CX23; Olympus Corporation, Tokyo, Japan) at a
magnification of ×400. A total of 15 fields of each tissue from
each piglet were assessed for the statistical analysis.
 | Table I.Information of antibodies. |
Table I.
Information of antibodies.
|
| Primary
antibodies | Secondary
antibodies |
|---|
|
|
|
|
|---|
| No. | Name | Cat. no. | Dilution | Supplier | Name | Cat. no. | Dilution | Supplier |
|---|
| 1 | Rabbit anti-pig
CD3 | ab16669 | 1:400 | Abcam, Cambridge,
MA, USA | SABC-POD (rabbit
IgG) kit | SA1022 | 1:1 | Boster Biological
Technology, Pleasanton, CA, USA |
| 2 | Goat anti-pig
IgA | A100-102P | 1:200 | Bethyl
Laboratories, Inc., Montgomery, TX, USA | SABC-POD (goat IgG)
Kit | SA1023 | 1:1 | Boster Biological
Technology |
| 3 | Mouse anti-pig
CD4 | sc1140 | 1:200 | Santa Cruz
Biotechnology, Inc., Dallas, TX, USA | SABC-POD (mouse
IgG) kit | SA1021 | 1:1 | Boster Biological
Technology |
| 4 | Mouse anti-pig
CD8 | NBP1-28237 | 1:200 | Novus Biologicals,
LLC, Littleton, CO, USA | SABC-POD (mouse
IgG) kit | SA1021 | 1:1 | Boster Biological
Technology |
| 5 | PE-conjugated goat
anti-pig CD11b | ab62817 | 1:200 | Abcam, Cambridge,
MA, USA | AlexaFluor
594-conjugated donkey anti-goat IgG2b | A-11058 | 1:200 | Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA |
| 6 | FITC-conjugated
mouse anti-pig MHC II | MCA2314F | 1:200 | Bio-Rad
Laboratories, Inc., Hercules, CA, USA | AlexaFluor
488-conjugated donkey anti-mouse IgG1 | A-21202 | 1:200 | Invitrogen; Thermo
Fisher Scientific, Inc. |
| 7 | Rabbit anti-human
TLR-2 | PA1-41045 | 1:1,000 | Thermo Fisher
Scientific, Inc., Waltham, MA, USA | HRP-conjugated goat
anti-rabbit IgG | A6154 | 1:5,000 | Sigma Aldrich;
Merck KGaA, Darmstadt, Germany |
| 8 | Rabbit anti-human
TLR-2 | PA5-20202 | 1:1,000 | Thermo Fisher
Scientific, Inc., Waltham, MA, USA | HRP-conjugated goat
anti-rabbit IgG | A6154 | 1:5,000 | Sigma Aldrich;
Merck KGaA |
| 9 | Mouse anti-pig
GAPDH | AF0006 | 1:1,000 | Beyotime Institute
of Biotechnology, Haimen, China | HRP-conjugated goat
anti-mouse IgG | A9917 | 1:5,000 | Sigma Aldrich;
Merck KGaA |
Immunofluorescence and confocal
microscopy
The frozen sections were incubated with 5% BSA for
20 min at room temperature and subsequently incubated with
PE-conjugated goat anti-pig CD11b primary antibodies (Table I) at 4°C for 12 h. The sections were
washed in PBS and incubated with AlexaFluor 594-conjugated donkey
anti-goat immunoglobulin G (IgG)2b secondary antibodies at room
temperature for 1 h (Table I). The
sections were subsequently stained with DAPI solution (Invitrogen;
Thermo Fisher Scientific, Inc.) at room temperature for 5 min. The
negative control was treated in the same way except the primary
antibodies were omitted. The sections were visualized using a Zeiss
LSM710 confocal microscope (Zeiss AG, Oberkochen, Germany) at a
magnification of ×1,000.
The paraffin embedded sections were rinsed and
subjected to antigen retrieval as described above. The serum
incubation and antibody incubations were the same as for the frozen
sections described above. The primary antibodies used were
PE-conjugated goat anti-pig CD11b and fluorescein
isothiocyanate-conjugated mouse anti-pig major histocompatibility
complex (MHC)II antibodies (Table
I). The fluorescent secondary antibodies used were AlexaFluor
594-conjugated donkey anti-goat IgG2b and AlexaFluor 488-conjugated
donkey anti-mouse IgG1 (Table I). A
total of 15 different fields of each tissue type were assessed from
each piglet for statistical analysis by using Image Pro Plus
software (version 6.0; Media Cybernetics, Inc., Rockville, MD,
USA).
RNA isolation and RT-qPCR
Total RNA was extracted from mucosa segments using a
TRIzol™ Plus RNA Purification kit (Thermo Fisher
Scientific, Inc.). Total RNA (2 mg) reverse transcribed using
PrimeScript™ RT-PCR kit (Takara Biotechnology Co., Ltd.,
Dalian, China) according to the manufacturer's protocol. Reverse
transcription was performed at 37°C for 50 min and then at 70°C for
15 min. A total of 2 µl diluted cDNA (vol/:vol, 1:20) was used for
RT-qPCR analysis, which was performed with an ABI 7500 PCR system
(Life Technologies; Thermo Fisher Scientific, Inc.) using
SYBR-Green qPCR Master Mix (Takara Biotechnology Co., Ltd., Dalian,
China) according to the manufacturer's protocol. The thermocycling
protocol was 95°C for 30 sec, followed by 40 cycles at 95°C for 5
sec and 60°C for 31 sec. The data was normalized to the β-actin
housekeeping gene to account for repeated measures. The specific
primers used for PCR are listed in Table II. The porcine TNF-α, IL-8, IL-1β,
IL-6, pBD-2, TLR-2 and TLR-9 primers were taken from Deng et
al (27) and Khoufache et
al (28). PCR products were
analyzed using a melting curve and the results (fold change) were
calculated using the 2−ΔΔCq method (29).
| Table II.Oligonucleotide polymerase chain
reaction primers. |
Table II.
Oligonucleotide polymerase chain
reaction primers.
| Gene | GenBank accession
number | Primers sequence
(5′-3′) | Orientation |
|---|
| TNF-α | X57321.1 |
CACCACGCTCTTCTGCCTACTGC | Forward |
|
|
|
TCGGCTTTGACATTGGCTACAA | Reverse |
| IL-8 | NM213867 |
TAGGACCAGAGCCAGGAAGA | Forward |
|
|
|
GAACTGCAGCCTCACAGAGA | Reverse |
| IL-1β | M86725.1 |
AAGTGATGGCTAACTACGGTGAC | Forward |
|
|
|
ATCTGCCTGATGCTCTTGTTCC | Reverse |
| IL-6 | M86722.1 |
TGGATAAGCTGCAGTCACAG | Forward |
|
|
|
ATTATCCGAATGGCCCTCAG | Reverse |
| pBD-2 | AY506573.1 |
ACCTGCTTACGGGTCTTG | Forward |
|
|
|
CTCTGCTGTGGCTTCTGG | Reverse |
| TLR-2 | AB072190 |
ACATGAAGATGATGTGGGCC | Forward |
|
|
|
TAGGAGTCCTGCTCACTGTA | Reverse |
| TLR-9 | AB071394 |
GTGGAACTGTTTTGGCATC | Forward |
|
|
|
CACAGCACTCTGAGCTTTGT | Reverse |
| β-actin | U07786.1 |
CATCACCATCGGCAACGA | Forward |
|
|
|
GCGTAGAGGTCCTTCCTGATGT | Reverse |
ELISA and western blot analysis
Tissue samples (50 mg) were homogenized in PBS using
a Brinkman homogenizer and subjected to three freeze-thaw cycles
(−20°C and 37°C). The homogenates were subsequently centrifuged for
5 min at 5,000 × g at 4°C. Levels of IL-1β, IL-8 and TNF-α proteins
in the supernatant were analyzed using IL-1β (cat. no. 70-EK101B2),
IL-8 (cat. no. 70-EK1082) and TNF-α (cat. no. 70-EK1822) ELISA kits
[all Hangzhou MultiSciences (Lianke) Biotech, Co, Ltd., Hangzhou,
China] according to the manufacturer's protocol. Each sample was
run in triplicate.
Tissue samples (50 mg) were homogenized in 500 µl
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Shanghai, China) with a Brinkman Homogenizer. Total
protein was extracted using a Protein Extraction kit (Beyotime
Institute of Biotechnology) and protein content was measured using
a bicinchoninic acid assay according to the manufacturer's protocol
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). The homogenates
were subsequently centrifuged for 15 min at 14,000 × g at 4°C. The
supernatant was diluted in a 5:1 ratio with SDS-PAGE Sample Loading
Buffer (5X; Beyotime Institute of Biotechnology) and heated to 98°C
for 10 min. Proteins (40 mg/lane) were separated in an 8% gel via
SDS-PAGE. The protein was then transferred onto a 0.45 µm-pore
polyvinylidene difluoride membrane (Immun-Blot®; Bio-Rad
Laboratories, Inc., Hercules, CA, USA) at 100 V for 1 h. The
membranes were blocked with 7% skim milk in PBS with 0.1% Tween-20
at room temperature for 2 h. The TLR-2 and TLR-9 proteins were
detected using western blot analysis with rabbit anti-human TLR-2
and rabbit anti-human TLR-9 polyclonal antibodies (Table I) for 10 h at 4°C, followed by
horseradish peroxidase-conjugated goat anti-rabbit IgG secondary
antibodies (Table I) for 1 h at room
temperature. The signals were detected using Supersignal West Pico
kit (Thermo Fisher Scientific, Inc.) and subjected to an ImageQuant
LAS-4000 imaging system (GE Healthcare Bio-Sciences, Pittsburgh,
PA, USA). The intensity of the bands in terms of density was
measured by Image Pro Plus version 6.0 (Media Cybernetics, Inc.)
and normalized against GAPDH expression. Three independent
experiments and appropriate gel exposures yielded very similar
results for each treatment modality.
Statistical analysis
Measurements and cell counts were performed using
Image Pro Plus version 6.0 (Media Cybernetics, Inc.). Results are
expressed as the mean ± standard error of the mean. One-way
analysis of variance, followed by Duncan's test, was employed to
determine statistical differences among multiple groups and t-test
was employed to determine the same between two groups. Histological
scores were analyzed by the Mann-Whitney U test. P<0.05
indicated that the difference between groups was statistically
significant.
Results
Analysis of B. subtilis location
following intranasal administration
Dylight 488-labeled B. subtilis were observed
in the lamina propria of the nasal mucosa, nasopharyngeal tonsils
and soft palate tonsils, and the concentration of B.
subtilis was markedly higher in the nasopharyngeal tonsil
compared with the nasal mucosa or the soft palate tonsils (Fig. 1A). B. subtilis were also
observed inside the CD11b+ DCs (Fig. 1A-b). The morphological structure of
the tissue did not appear to change and the epithelial integrity
remained intact following intranasal administration with B.
subtilis (Fig. 1B and C), which
indicates that intranasal administration with B. subtilis
did not cause pathological changes to the tissues.
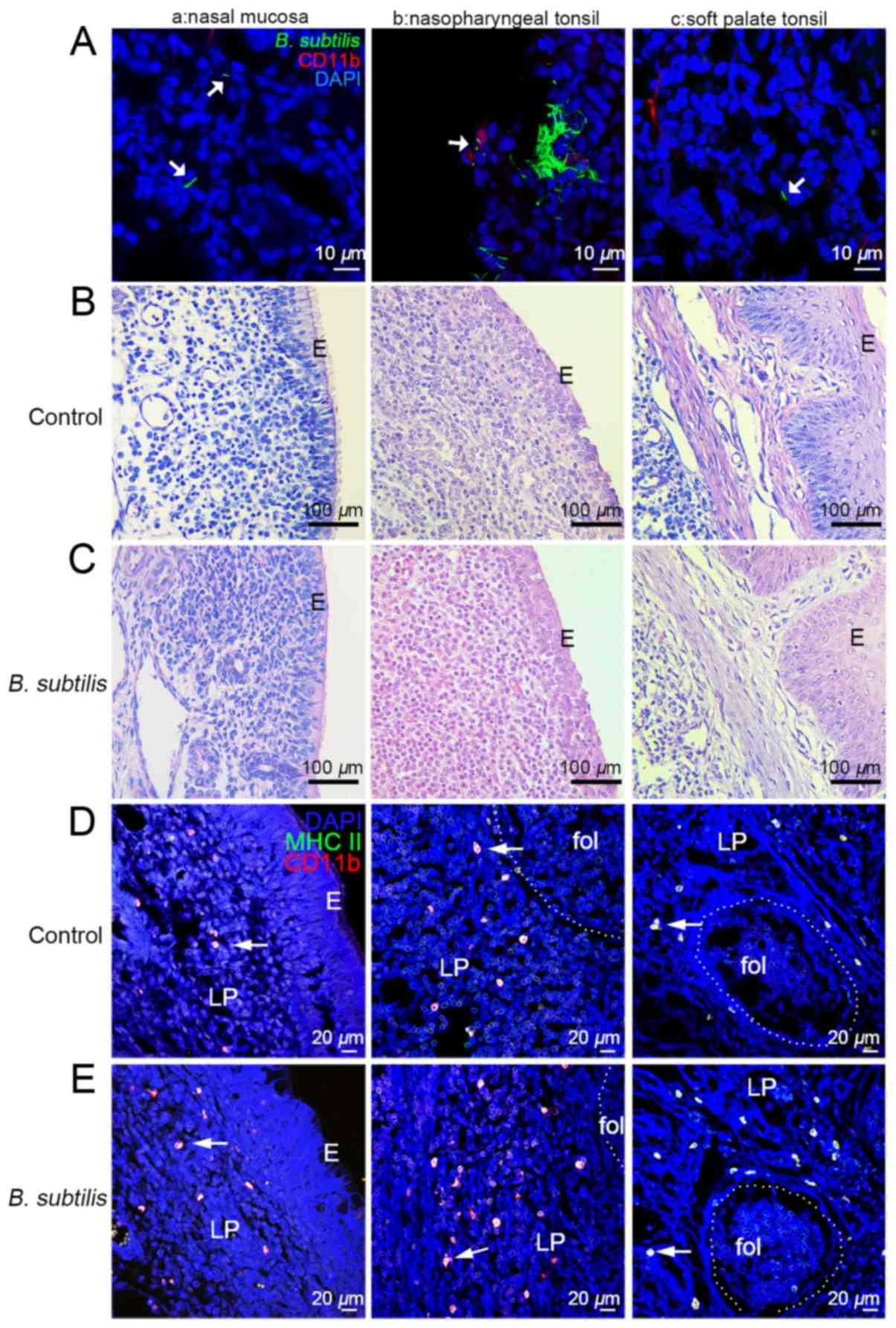 | Figure 1.Effects of intranasal administration
of B. subtilis on DCs in the nasal mucosa, nasopharyngeal
tonsil and soft palate tonsils of piglets. (A) Immunofluorescent
staining was performed to detect the prescence of Dylight
488-labeled B. subtilis (green) in the (A-a) lamina propria
of nasal mucosa, (A-b) nasopharyngeal tonsil and (A-c) soft palate
tonsil. CD11b+ DCs were labelled as red and DAPI was
labelled as blue. (n=3/group). Hematoxylin and eosin staining was
performed on (B) control samples and (C) B. subtilis samples
in the (B-a and C-a) nasal mucosa, (B-b and C-b) nasopharyngeal
tonsil and (B-c and C-c) soft palate tonsil. Immunofluorescent
staining was performed on (D) control samples and (E) B.
subtilis samples. CD11b+ (red) and MHC
II+ (green) DCs (arrows) were detected in the (D-a and
E-a) nasal mucosa, (D-b and E-b) nasopharyngeal tonsil, (D-c and
E-c) and soft palate tonsil. CD11b+ MHC II+
DCs were labelled as orange/yellow and DAPI was labelled as blue
(n=4/group). E, epithelium; LP, lamina propria; fol, lymphoid
follicle; DCs, dendritic cells; CD, cluster of differentiaion. |
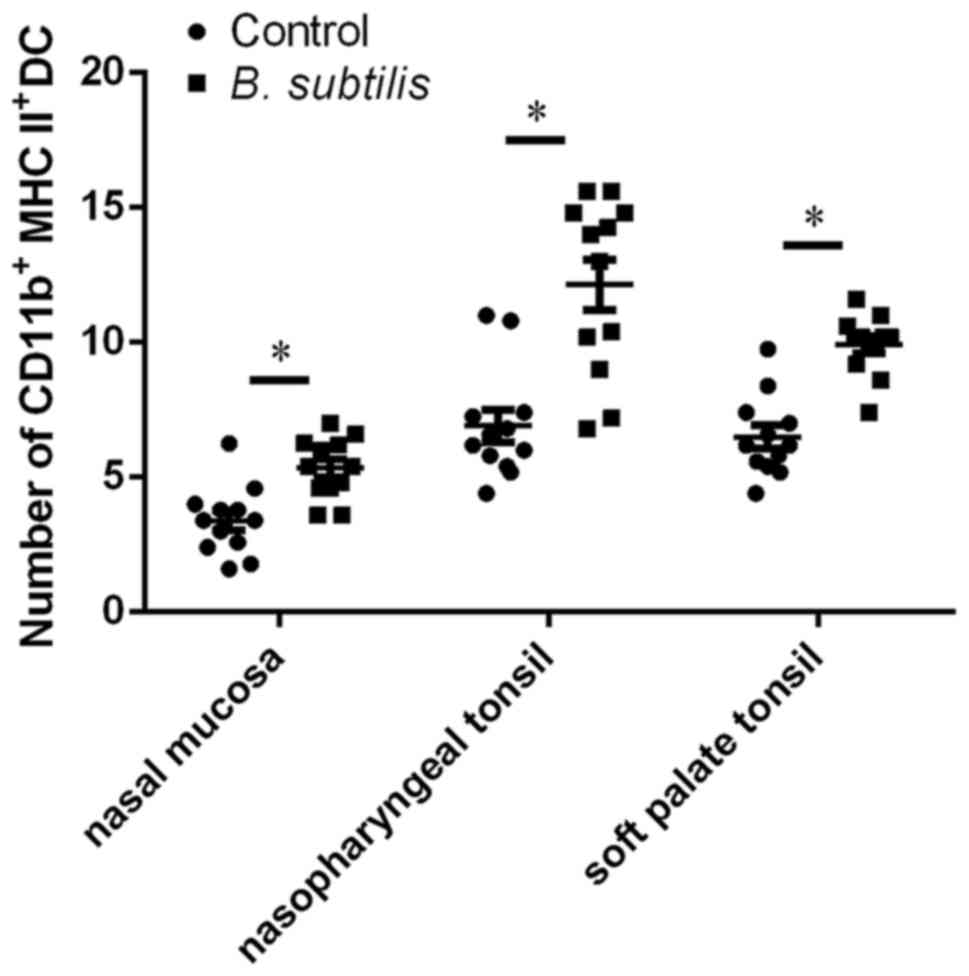
Effect of B. subtilis on DCs in the
nasal mucosa and tonsils
Immunofluorescent analysis via dual staining with
antibodies specific to the DC markers to detected DCs.
CD11b+ cells were stained red, MHC II+ cells
were stained green and double positive cells were stained bright
yellow (arrows; Fig. 1D and E).
There were several double positive cells and they were located in
the lamina propria particularly around the lymphatic follicles
(fol; dotted circles). The DCs were round or polygonal and
distributed in the lamina propria particularly around the lymphatic
follicles. CD11b+ MHC II+ DCs were
significantly increased in the nasal mucosa, nasopharyngeal tonsil
and soft palate tonsil following administration with B.
subtilis compared with the control (P<0.05; Fig. 2).
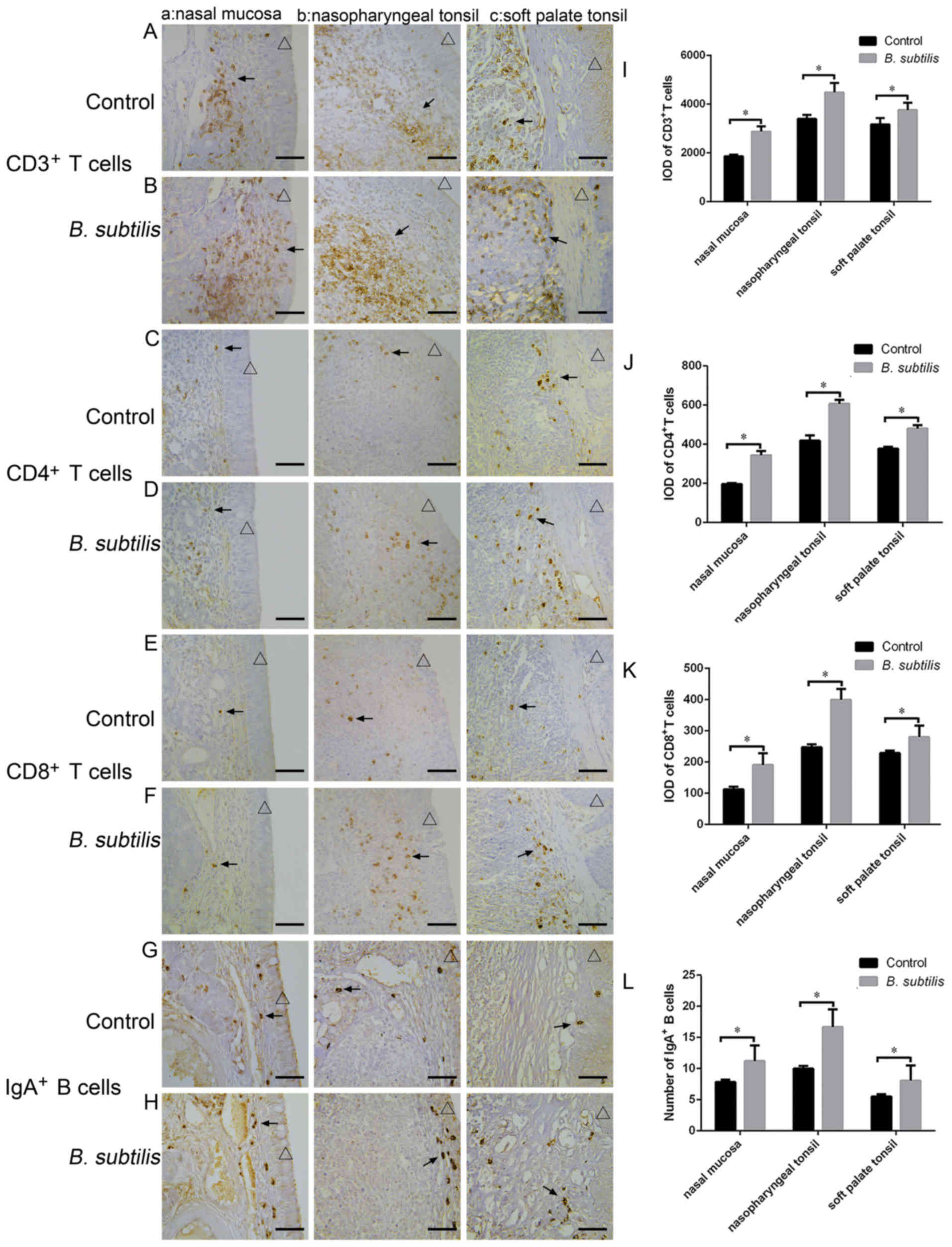
Effect of B. subtilis on T cells in
the nasal mucosa and tonsils
Immunohistochemical staining was performed on the
paraffin embedded samples (Fig. 3).
The results revealed that CD3+, CD4+ and
CD8+ T cells were widely distributed in the lamina
propria of the nasal mucosa, soft palate tonsils and nasopharyngeal
tonsils (Fig. 3A-F). The cells
positively stained for CD3+, CD4+ and
CD8+ appeared brown. The number of CD3+,
CD4+ and CD8+ T cells in the nasal mucosa,
nasopharyngeal tonsil and soft palate tonsil increased
significantly following the nasal administration of B.
subtilis compared with the control (P<0.05; Fig. 3I-K).
Effect of B. subtilis on
IgA+ B cells in the nasal mucosa and tonsils
Immunohistochemical staining revealed that the
IgA+ B cells (stained brown) were round or oval in shape
and primarily distributed in the lamina propria of the nasal
mucosa, soft palate tonsils and nasopharyngeal tonsils (Fig. 3G-H). Following the intranasal
administration of B. subtilis, the number of IgA+
B cells was significantly increased in the nasal mucosa,
nasopharyngeal tonsils and soft palate tonsils at 28 days post
administration compared with the control (P<0.05; Fig. 3L).
Expression of IL-1β, IL-6, IL-8,
TNF-α, pBD-2, TLR-2 and TLR-9 mRNA
RT-qPCR was performed to determine the mRNA levels
of IL-1β, IL-6, IL-8, TNF-α, pBD-2, TLR-2 and TLR-9 in the tissue
samples (Fig. 4). The expression of
IL-6 mRNA in the nasal cavities and tonsils increased significantly
following the intranasal administration of B. subtilis
compared with the control (P<0.05; Fig. 4D). The expression of pBD-2, TLR-2 and
TLR-9 mRNA also significantly increased following the
administration of B. subtilis, but only in the tonsils
(P<0.05; Fig. 4E-G). These
results indicate that the administration of B. subtilis
stimulates nasal mucosal immunity. However, no statistical
difference in mRNA expression was observed for IL-1β, TNF-α and
IL-8 between the B. subtilis group and the control group in
the nasal mucosa, nasopharyngeal tonsil or soft palate tonsil
(Fig. 4A-C).
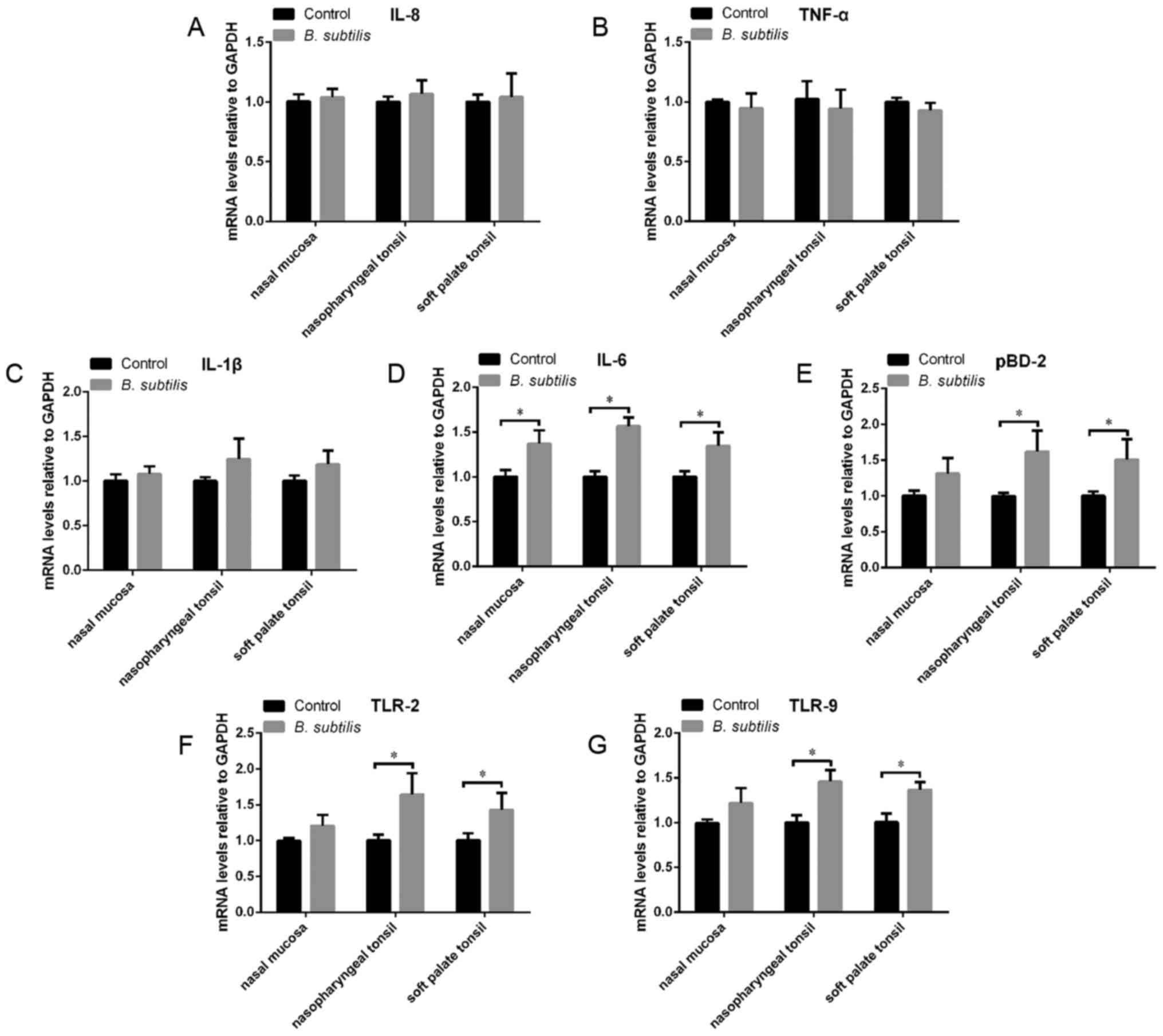 | Figure 4.mRNA expression of (A) IL-8, (B)
TNF-α, (C) IL-1β, (D) IL-6, (E) pBD-2, (F) TLR-2 and (G) TLR-9 in
the nasal mucosa, nasopharyngeal tonsils and soft palate tonsils.
Data are presented as the mean ± standard error of the mean
(n=4/group) and the error bars indicate the standard error.
*P<0.05. IL, interleukin; TLR, toll-like receptor; TNF, tumor
necrosis factor; pBD, porcine β-defensin 2. |
Protein expression of IL-1β, IL-8,
TNF-α, TLR-2, TLR-9
No statistical differences in the protein expression
of IL-1β, TNF-α and IL-8 were observed between the B.
subtilis group and the control group in the nasal mucosa,
nasopharyngeal tonsil or soft palate tonsil (Fig. 5A-C). The TLR-2 and TLR-9 protein
levels in the nasal mucosa, nasopharyngeal tonsils and soft palate
tonsils were significantly increased following intranasal
administration of B. subtilis (P<0.05; Fig. 5D-I).
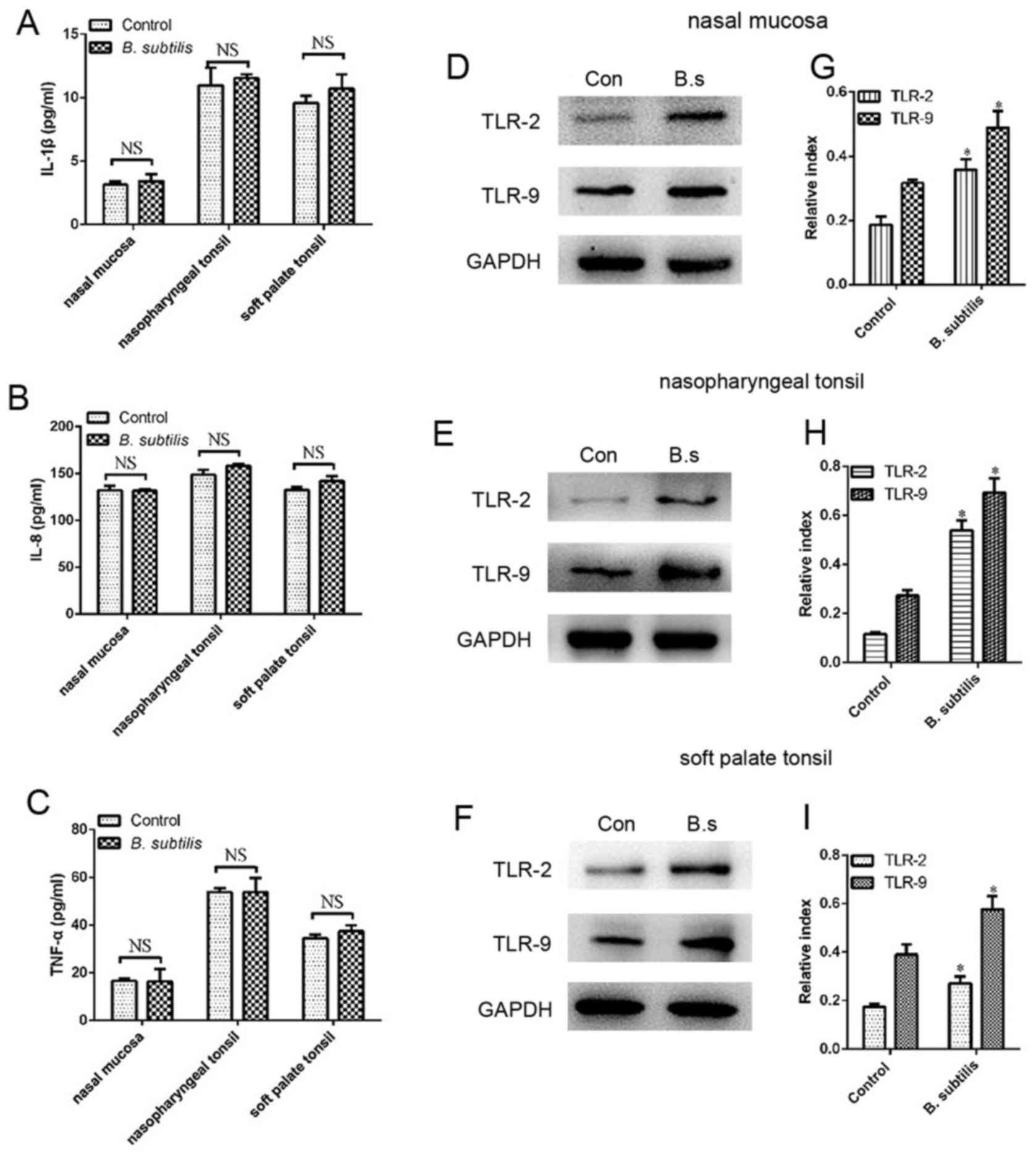 | Figure 5.Protein expression of IL-1β, IL-8,
TNF-α, TLR-2 and TLR-9 in the nasal mucosa, nasopharyngeal tonsils
and soft palate tonsils. ELISA analysis of (A) IL-1β, (B) IL-8 and
(C) TNF-α in the nasal mucosa, nasopharyngeal tonsils and soft
palate tonsils. Western blot analysis of TLR-2 and TLR-9 protein
expression in the (D) nasal mucosa, (E) nasopharyngeal tonsils and
(F) soft palate tonsils. TLR-2 and TLR-9 protein expression in the
(G) nasal mucosa, (H) nasopharyngeal tonsils and (I) soft palate
tonsils were normalized by comparison with GAPDH and the relative
index was determined in comparison with the control. Values
represent the mean ± standard error of the mean (n=4/group) and the
error bars indicate the standard error. *P<0.05. Con, the
control group; B.s, the B. subtilis group; IL, interleukin;
TLR, toll-like receptor; TNF, tumor necrosis factor. |
Discussion
The innate immune system represents the first line
of defense against pathogen invasion; the innate immunity of the
respiratory tract in pigs has previously been studied due to its
similarities with humans (30).
Innate immunity functions through interactions between immune cells
and TLRs, which are primarily expressed on APCs (31). The innate immune system promotes the
activation of the adaptive immune system (32) and therefore serves an important role
in the body's immune response. TLRs link the innate immune system
with specific adaptive immunity (33). TLR-2 has a wide range of ligands,
including lipoproteins and lipoteichoic acids from bacteria
(34). TLR-9 is able to recognize
bacterial DNA, which contains CpG motifs, and activate APCs and
native B cells (35). In the present
study, treatment with B. subtilis significantly increased
TLR-2 and TLR-9 protein expression in the nasal mucosa,
nasopharyngeal tonsils and soft palate tonsils. The results
revealed that immunomodulation induced by B. subtilis WB800
primarily depends on extracellular macromolecules TLRs.
Secretory IgA, which is the main antibody for local
mucosal immunity, is secreted by B cells (36). It may combine with antigens in the
digestive and respiratory tracts and mediate virus neutralization
in infected epithelial cells, or adhere to the mucosa to prevent
pathogen translocation (37). DCs
are the most ubiquitously distributed professional APCs (38) and are widely distributed throughout
the secondary lymphoid organs (39).
As the primary APCs in the nasal mucosa and tonsils, the
protrusions of DCs may extend across the epithelium to uptake
antigens and present them to T and B cells (40,41). The
maturation of DCs is a complex process during which they express
surface molecules and cytokines, which are important for T-cell
activation (42). It has previously
been suggested that DCs were associated with the proliferation of
CD4+ T cells, and that DCs may serve as APCs and
upregulate CD4+ T cells in the nasal mucosa (40). The present study demonstrated that
B. subtilis WB800 treatment significantly increased immune
cells, indicating that it enhanced the local immunity of the
piglets. A previous study has demonstrated that pBD-2 inhibits
pathogens in vitro (27). T
cells may produce cytokines that promote the maturation and
differentiation of B lymphocytes into antibody producing cells
(43,44).
A previous study revealed that probiotics may
upregulate the protein expression of pBD-2 in the saliva and
intestines of piglets (45). In
addition, it has been reported that the administration of
probiotics increases the expression of IL-6 in the nasal mucosa,
nasopharyngeal tonsil and soft palate tonsil (27,46,47).
Based on this previous research, the authors of the present study
hypothesized that the protein expression of pBD-2 and IL-6 would
increase following the administration of B. subtilis. The
focus of the present study was local immunity; however, the number
of tissue samples was limited. To combat this, the expression of
IL-1β, IL-8, TNF-α, TLR-2 and TLR-9 protein was measured in the
nasal mucosa, nasopharyngeal tonsils and soft palate tonsils, all
of which are key sites for local mucosal immunity. The present
study revealed that IL-6 mRNA was significantly increased in the
nasal mucosa, nasopharyngeal tonsil and soft palate tonsil
following intranasal administration of B. subtilis. This may
be due to the regulative capability of B. subtilis in
improving mucosal immune responses by inducing the production of
IL-6. No statistical differences were observed in levels of
pro-inflammatory factors TNF-α, IL-1β and IL-8 between the B.
subtilis group and the control. These results indicate that the
increase in immune cells and cytokines was not caused by
inflammatory reactions. A previous study has reported that B.
subtilis may effectively modulate the intestinal microbiota by
enhancing the proliferation of beneficial bacteria and inhibiting
potential pathogens, including Staphylococcus and
Escherichia-Shigella, in the intestinal tract (48). A previous study reported that B.
subtilis reduced Salmonella enteritidis attachment to
the surfaces of intestinal epithelial cells (49). Based on this previous research, the
authors of the present study hypothesized that B. subtilis
had the ability to modulate the nasal microbiota and enhance the
local mucosal immunity.
In conclusion, intranasal administration of B.
subtilis increased the number of immune cells in the nasal
mucosa, nasopharyngeal tonsils and soft palate tonsils, which
enhanced the nasal mucosal and tonsillar immunity of piglets. The
present study provides a basis for the further study of intranasal
immunization with B. subtilis as a mucosal adjuvant. It also
lays the foundation for the potential intranasal administration of
B. subtilis in humans, which may enhance the immune response
of human nasal mucosa against respiratory diseases.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 31172302) and a
project funded by the Priority Academic Program Development of
Jiangsu Higher Education Institutions.
Availability of data and materials
Not applicable.
Authors' contributions
YY and QY conceived and designed the study and wrote
the paper. YY, YJ, and JY performed the experiments. YY, YJ, JY,
and QY reviewed and edited the manuscript. All authors read and
approved the manuscript.
Ethics approval and consent to
participate
All procedures were approved by the Institutional
Animal Care and Use Committee of Nanjing Agricultural University
(Nanjing, China) and followed the National Institutes of Health
guidelines for the performance of animal experiments.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Meng Y, Yang Y, Lu W, Wang Y, Qian F, Wang
X, Zhang Z and Wang W: The inhibition of Platycodin D on Mycoplasma
pneumoniae proliferation and its effect on promoting cell growth
after anti-Mycoplasma pneumoniae treatment. Front Cell Infect
Microbiol. 4:1922015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim TK, Bheda-Malge A, Lin Y, Sreekrishna
K, Adams R, Robinson MK, Bascom CC, Tiesman JP, Isfort RJ and
Gelinas R: A systems approach to understanding human rhinovirus and
influenza virus infection. Virology. 486:146–157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Persson J, Zhang Y, Olafsdottir TA, Thörn
K, Cairns TM, Wegmann F, Sattentau QJ, Eisenberg RJ, Cohen GH and
Harandi AM: Nasal immunization confers high avidity neutralizing
antibody response and immunity to primary and recurrent genital
herpes in guinea pigs. Front Immunol. 7:6402016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang J, Dai L, Yu QH and Yang Q:
Histological and anatomical structure of the nasal cavity of Bama
minipigs. PLoS One. 12:e01739022017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amuguni H, Lee S, Kerstein K, Brown D,
Belitsky B, Herrmann J, Keusch G, Sonenshein A and Tzipori S:
Sublingual immunization with an engineered Bacillus subtilis strain
expressing tetanus toxin fragment C induces systemic and mucosal
immune responses in piglets. Microbes Infect. 14:447–456. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hellings P, Jorissen M and Ceuppens JL:
The Waldeyer's ring. Acta Otorhinolaryngol Belg. 54:237–241.
2000.PubMed/NCBI
|
|
7
|
Casteleyn C, Breugelmans S, Simoens P and
Van den Broeck W: The tonsils revisited: Review of the anatomical
localization and histological characteristics of the tonsils of
domestic and laboratory animals. Clin Dev Immunol. 2011:4724602011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cisney ED, Fernandez S, Hall SI, Krietz GA
and Ulrich RG: Examining the role of nasopharyngeal-associated
lymphoreticular tissue (NALT) in mouse responses to vaccines. J Vis
Exp: 3960. 2012. View
Article : Google Scholar
|
|
9
|
Casteleyn C, Broos AM, Simoens P and Van
den Broeck W: NALT (nasal cavity-associated lymphoid tissue) in the
rabbit. Vet Immunol Immunopathol. 133:212–218. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pabst R: Mucosal vaccination by the
intranasal route. Nose-associated lymphoid tissue (NALT)-Structure,
function and species differences. Vaccine. 33:4406–4413. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brandtzaeg P and Pabst R: Let's go
mucosal: Communication on slippery ground. Trends Immunol.
25:570–577. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Khaiboullina SF, Morzunov SP, Hall MR, De
Meirleir KL, Rizvanov AA and Lombardi VC: Human dendritic cells
transfected with a human papilloma virus-18 construct display
decreased mobility and upregulated cytokine production. Int J
Oncol. 43:1701–1709. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rate A, Bosco A, McKenna KL, Holt PG and
Upham JW: Airway epithelial cells condition dendritic cells to
express multiple immune surveillance genes. PLoS One. 7:e449412012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakahara T, Moroi Y, Uchi H and Furue M:
Differential role of MAPK signaling in human dendritic cell
maturation and Th1/Th2 engagement. J. Dermatol Sci. 42:1–11. 2006.
View Article : Google Scholar
|
|
15
|
Jamin A, Gorin S, Le Potier MF and
Kuntz-Simon G: Characterization of conventional and plasmacytoid
dendritic cells in swine secondary lymphoid organs and blood. Vet
Immunol Immunopathol. 114:224–237. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fijan S: Microorganisms with claimed
probiotic properties: An overview of recent literature. Int J
Environ Res Public Health. 11:4745–4767. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Galdeano CM, de Moreno de LeBlanc A,
Vinderola G, Bonet ME and Perdigón G: Proposed model: Mechanisms of
immunomodulation induced by probiotic bacteria. Clin Vaccine
Immunol. 14:485–492. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Toure Y, Ongena M, Jacques P, Guiro A and
Thonart P: Role of lipopeptides produced by Bacillus subtilis GA1
in the reduction of grey mould disease caused by Botrytis cinerea
on apple. J Appl Microbiol. 96:1151–1160. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang JM, La Ragione RM, Nunez A and
Cutting SM: Immunostimulatory activity of Bacillus spores. FEMS
Immunol Med Microbiol. 53:195–203. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liang J, Fu J, Kang H, Lin J, Yu Q and
Yang Q: Comparison of 3 kinds of Toll-like receptor ligands for
inactivated avian H5N1 influenza virus intranasal immunization in
chicken. Poult Sci. 92:2651–2660. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song M, Hong HA, Huang JM, Colenutt C,
Khang DD, Nguyen TV, Park SM, Shim BS, Song HH, Cheon IS, et al:
Killed Bacillus subtilis spores as a mucosal adjuvant for an H5N1
vaccine. Vaccine. 30:3266–3277. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mou C, Zhu L, Yang J, Xu W, Cheng X and
Yang Q: Immune responses induced by recombinant Bacillus subtilis
expressing the hemagglutinin protein of H5N1 in chickens. Sci Rep.
6:384032016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Upadhaya SD, Shanmugam SK, Kang DK and Kim
IH: Preliminary assessment on potentials of probiotic B. subtilis
RX7 and B. methylotrophicus C14 strains as an immune modulator in
Salmonella-challenged weaned pigs. Trop Anim Health Prod.
49:1065–1070. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang W, Zhu YH, Zhou D, Wu Q, Song D,
Dicksved J and Wang JF: Oral administration of a select mixture of
Bacillus probiotics affects the gut microbiota and goblet cell
function following Escherichia coli challenge in newly weaned pigs
of genotype MUC4 that are supposed to be enterotoxigenic E. coli
F4ab/ac receptor negative. Appl Environ Microbiol. 83:pii: e02747.
–16. 2017. View Article : Google Scholar
|
|
25
|
Sheng QK, Zhou KF, Hu HM, Zhao HB, Zhang Y
and Ying W: Effect of Bacillus subtilis natto on meat quality and
skatole content in TOPIGS pigs. Asian-Australas J Anim Sci.
29:716–721. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shi C, Zhang Y, Lu Z and Wang Y:
Solid-state fermentation of corn-soybean meal mixed feed with
Bacillus subtilis and Enterococcus faecium for degrading
antinutritional factors and enhancing nutritional value. J Anim Sci
Biotechnol. 8:502017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Deng J, Li Y, Zhang J and Yang Q:
Co-administration of Bacillus subtilis RJGP16 and Lactobacillus
salivarius B1 strongly enhances the intestinal mucosal immunity of
piglets. Res Vet Sci. 94:62–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khoufache K, Cabaret O, Farrugia C,
Rivollet D, Alliot A, Allaire E, Cordonnier C, Bretagne S and
Botterel F: Primary in vitro culture of porcine tracheal epithelial
cells in an air-liquid interface as a model to study airway
epithelium and Aspergillus fumigatus interactions. Med Mycol.
48:1049–1055. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Loving CL, Osorio FA, Murtaugh MP and
Zuckermann FA: Innate and adaptive immunity against porcine
reproductive and respiratory syndrome virus. Vet Immunol
Immunopathol. 167:1–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tu S, Zhong D, Xie W, Huang W, Jiang Y and
Li Y: Role of toll-like receptor signaling in the pathogenesis of
graft-versus-host diseases. Int J Mol Sci. 17:pii: E1288. 2016.
View Article : Google Scholar
|
|
32
|
Romo MR, Pérez-Martinez D and Ferrer
Castillo C: Innate immunity in vertebrates: An overview.
Immunology. 148:125–139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang EJ and Lu MJ: Toll-like receptor
(TLR)-mediated innate immune responses in the control of hepatitis
B virus (HBV) infection. Med. Microbiol Immunol. 204:11–20. 2015.
View Article : Google Scholar
|
|
34
|
Modlin RL: Mammalian toll-like receptors.
Ann Allerg Asthma Immunol. 88:543–548. 2002. View Article : Google Scholar
|
|
35
|
Giordani L, Sanchez M, Libri I, Quaranta
MG, Mattioli B and Viora M: IFN-α amplifies human naive B cell
TLR-9-mediated activation and Ig production. J Leukocyte Biol.
86:261–271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luo Q, Cui HM, Peng X, Fang J, Zuo Z, Deng
J, Liu J and Deng Y: Intestinal IgA+ cell numbers as well as IgA,
IgG, and IgM contents correlate with mucosal humoral immunity of
broilers during supplementation with high fluorine in the diets.
Biol Trace Elem Res. 154:62–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu B, Cui H, Peng X, Fang J, Zuo Z, Deng J
and Huang J: Toxicological effects of nickel chloride on IgA+ B
cells and sIgA, IgA, IgG, IgM in the intestinal mucosal immunity in
broilers. Int J Environ Res Public Health. 11:8175–8192. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gao Q, Zhao S, Qin T, Yin Y, Yu Q and Yang
Q: Effects of inactivated porcine epidemic diarrhea virus on
porcine monocyte-derived dendritic cells and intestinal dendritic
cells. Res Vet Sci. 106:149–158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Durand M and Segura E: Dendritic cell
subset purification from human tonsils and lymph nodes. Methods Mol
Biol. 1423:89–99. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fukuyama Y, Tokuhara D, Sekine S, Kataoka
K, Markham JD, Irwin AR, Moon GH, Tokuhara Y, Fujihashi K, Davydova
J, et al: Notch-ligand expression by NALT dendritic cells regulates
mucosal Th1- and Th2-type responses. Biochem Biophys Res Commun.
418:6–11. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Qin T, Yin Y, Wang X, Liu H, Lin J, Yu Q
and Yang Q: Whole inactivated avian Influenza H9N2 viruses induce
nasal submucosal dendritic cells to sample luminal viruses via
transepithelial dendrites and trigger subsequent DC maturation.
Vaccine. 33:1382–1392. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Rajan Sundara S and Longhi MP: Dendritic
cells and adipose tissue. Immunology. 149:353–361. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lammers A, Wieland WH, Kruijt L, Jansma A,
Straetemans T, Schots A, den Hartog G and Parmentier HK: Successive
immunoglobulin and cytokine expression in the small intestine of
juvenile chicken. Dev Comp Immunol. 34:1254–1262. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Moutsopoulos NM, Nares S, Nikitakis N,
Rangel Z, Wen J, Munson P, Sauk J and Wahl SM: Tonsil epithelial
factors may influence oropharyngeal human immunodeficiency virus
transmission. Am J Pathol. 171:571–579. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang J, Deng J, Li Y and Yang Q: The
effect of Lactobacillus on the expression of porcine β-defensin-2
in the digestive tract of piglets. Livestock Sci. 138:259–265.
2011. View Article : Google Scholar
|
|
46
|
Zhou D, Zhu YH, Zhang W, Wang ML, Fan WY,
Song D, Yang GY, Jensen BB and Wang JF: Oral administration of a
select mixture of Bacillus probiotics generates Tr1 cells in weaned
F4ab/acR-pigs challenged with an F4+ ETEC/VTEC/EPEC strain. Vet
Res. 46:952015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Cilieborg MS, Thymann T, Siggers R, Boye
M, Bering SB, Jensen BB and Sangild PT: The incidence of
necrotizing enterocolitis is increased following probiotic
administration to preterm pigs. J Nutr. 141:223–230. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li Y, Xu Q, Huang Z, Lv L, Liu X, Yin C,
Yan H and Yuan J: Effect of Bacillus subtilis CGMCC 1.1086 on the
growth performance and intestinal microbiota of broilers. J Appl
Microbiol. 120:195–204. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Thirabunyanon M and Thongwittaya N:
Protection activity of a novel probiotic strain of Bacillus
subtilis against Salmonella Enteritidis infection. Res Vet Sci.
93:74–81. 2012. View Article : Google Scholar : PubMed/NCBI
|