Introduction
Sepsis is a clinical syndrome defined by
physiological changes with a systemic inflammation response, and it
remains a major medical challenge in pediatric and critical care
medicine (1,2). Sepsis is often caused by documented or
suspected infection with bacteria that produce lipopolysaccharide
(LPS), and septic shock is the progression of those physiological
changes to the extent that delivery of oxygen and metabolic
substrate to tissues is compromised (3). Sepsis is usually diagnosed when the
symptoms of the systemic inflammatory response syndrome (SIRS)
develop. SIRS is defined clinically by the activation of the
immune/inflammatory response, resulting in abnormal temperature or
leukocyte count (4). Biomarkers have
been identified that have the potential to diagnose, monitor,
stratify and predict the outcome of these syndromes, for example,
C-reactive protein and interleukin (IL)-18 elevation have been used
as biomarkers indicating sepsis (5,6).
However, many clinicians in the intensive care unit still face the
challenge of diagnosing and accurately assessing the risk of
outcome. More importantly, initiating appropriate therapy for
sepsis may help improve patient management and decrease
sepsis-related morbidity and mortality.
IL-31 is a cytokine derived from T cells, namely T
helper 2 (Th2) cells, that shares several structural and functional
characteristics with IL-6, oncostatin M, leukemia inhibitory factor
and cardiotrophin-1 (7). IL-31
signals through a receptor complex comprised of GPL (also known as
gp130-like receptor or IL-31RA) and oncostatin M receptor (OSMR)
(8–10). GPL/OSMR signaling is a strong
activator of signal transducer and activator of transcription STAT3
and STAT5, and also activates STAT1, Janus kinase (JAK)1 and JAK2
signaling pathways (11).
IL-31-regulated immune responses have been implicated in skin
physiology and inflammatory skin diseases (12). Research has reported that IL-31 may
negatively regulate lung type 2 inflammation by IL-31/IL-31
receptor (R) interaction (13). In
addition, IL-31-IL-31R interactions limit the magnitude of Th2
cytokine-dependent immunity and inflammation following intestinal
helminth infection (14). However,
the effect of IL-31 on sepsis and its underlying mechanisms remain
uncertain. NLR family, pyrin domain-containing 3 (NLRP3, CIAS1,
PYPAF or cryopyrin, is a cytosolic member of the NLRP family of
proteins expressed in leukocytes, particularly neutrophils
(15). As a component of the
inflammasome, NLRP3 activates caspase 1 and causes the maturation
cleavage of pro-IL-1β and pro-IL-18 (16). Defects in NLRP3 may cause
autoinflammatory syndromes. A subgroup of the nucleotide-binding
domain, leucine-rich repeat containing (NLR) proteins are key
mediators of the inflammasome. IL-1β functions in the systemic
responses to infection and chronic and acute inflammation (17). IL-1β (p17) is the mature active form
of pro-IL-1β (p35) that has to be cleaved by caspase 1 at Asp116
(15,18). The present study examined the role
IL-31 on experimental sepsis and investigated the mechanisms of
IL-31 efficacy responsible for the regulation of NLRP3 as well as
IL-1β.
Materials and methods
Mouse strains and reagents
C57BL/6 mice (50 female mice, 20–30 g and 6–10 weeks
old) were obtained from Vital River Laboratories Co., Ltd.,
(Beijing, China) and were housed in plastic cages (3–4 mice per
cage) with free access to drinking filtered tap water and food
under controlled conditions of humidity (50±10%), light (12-h
light/dark cycle) and temperature (23±2°C). Mice were maintained in
a specific pathogen free facility in accordance with institutional
guidelines. The protocol was approved by the Committee on the
Ethics of Animal Experiments of Capital Medical University
(Beijing, China).
Septic shock model
To induce in vivo cytokine secretion, 7-week
old female mice were injected intraperitoneally with LPS (10 mg/kg
body weight) (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and
their health status was monitored at regular intervals. A total of
6 h after the injection, the peritoneal cavities were washed with
0.8 ml phosphate-buffered saline (PBS) containing 1% fetal bovine
serum (FBS, Invitrogen; Thermo Fisher Scientific, Inc., Waltham,
MA, USA). Cytokines in the peritoneal lavage fluids and in the sera
were then measured by ELISA. The cecal ligation and puncture (CLP)
technique was used to induce intraabdominal sepsis in mice
(19). All the mice received LPS
injection or CLP operation, and 6–8 mice were checked in each
group. Control mice were injected with PBS compared with IL-31
injection after LPS or CLP treatment. The mice were divided into
the following groups: Vehicle control group (IL-31-), which
received PBS injection + LPS or CLP operation; IL-31 treatment
group (IL-31+), which received IL-31 injection + LPS or CLP
operation. In total, four groups (PBS+LPS, IL-31+LPS, PBS+CLP and
IL-31+CLP) were used for the study (6–8 mice per group).
Cells
In preparation for the isolation of peritoneal
macrophages as described previously (20), mice were intraperitoneally injected
with 1 ml 4% thioglycollate (Sigma-Aldrich; Merck KGaA, B2551), and
peritoneal exudate cells were isolated from the peritoneal cavity 4
days post-injection. The cells were then incubated at 37°C for 6 h
and washed three times with Hank's Balanced Salt Solution (HBSS;
Thermo Fisher Scientific, Inc.). The remaining adherent cells were
used as the peritoneal macrophages described in previous
experiments (20). Unless otherwise
indicated, the macrophages were primed with 200 ng/ml LPS from
Escherichia coli 0111:B4 (Sigma-Aldrich; Merck KGaA) for 4 h
at 37°C before stimulation with 5 mM adenosine triphosphate (ATP;
Sigma-Aldrich; Merck KGaA, A6419) for 30 min at 37°C.
Human peripheral blood monocytes (PBMC) were
obtained from healthy donors who provided written informed consent.
The cells were adjusted to 5×106 cells/ml and
resuspended in RPMI-1640 culture medium (Invitrogen; Thermo Fisher
Scientific, Inc.) supplemented with 50 mg/ml gentamicin, 2 mM
L-glutamine and 1 mM pyruvate. Human THP-1 cells were purchased
from the China Center for Type Culture Collection (Wuhan, China).
THP-1 cells were cultured in RPMI-1640 supplemented with 10% FBS,
1% HEPES, 1% L-glutamine, and 50 µg of cefotaxime. The cells were
treated with 5 nM phorbol myristate acetate (Sigma-Aldrich; Merck
KGaA) overnight and then washed three times. Cells were rested 3
days following chemical differentiation to ensure that they
reverted to a resting phenotype.
Proteins and antibodies
All reagents used in the present study were from
Sigma-Aldrich (Merck KGaA), unless stated otherwise. Recombinant
murine IL-31 was obtained from PeproTech Company (Suzhou, China). A
total of 6–8 mice for each IL-31 treatment group were used in the
three groups of experiments: i) Survival study of IL-31 treatment
and control treatment (8 mice in PBS+LPS vs. 8 mice in IL-31+LPS
for survival rate check); ii) cytokine analysis of LPS-induced
sepsis for the IL-31 treatment and control treatment (8 mice in
PBS+LPS vs. 8 mice in IL-31+LPS for cytokine analysis); and iii)
cytokine analysis of CLP operation for the IL-31 treatment and
control treatment (8 mice in PBS+CLP vs. 8 mice in IL-31+CLP for
cytokine analysis). A total of 100 µg of IL-31 cytokine in PBS for
one time i.p. injection was given per mouse in vivo. Control
mice were injected with PBS vehicle compared with IL-31 injection
after LPS or CLP treatment. Functional anti-human IL-31 antibody
(cat. no. AF2824, 1 µg/ml; R&D Systems China Co., Ltd.,
Shanghai, China) and anti-human IL-31RA antibody (Nemolizumab; cat.
no. TAB-439CQ, 1 µg/ml; Creative Biolabs; New York, NY, USA).
ELISA
Mouse (m)TNFα (cat. no. MTA00B), mIL-18 (cat. no.
7625), mIL-1β (cat. no. MLB00C), human (h)IL-1β (cat. no. DLB50),
hIL-6 (cat. no. D6050) and hTNF-α (cat. no. DTA00C) ELISA kits for
cytokine detection in the peritoneal lavage fluids, sera or cell
culture supernatant were obtained from R&D Systems China Co.,
Ltd. To detect low levels of cytokines in the samples, a standard
curve was obtained by diluted standard reagents.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from cell culture was extracted using
TRIzol reagent (Invitrogen, Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and subjected to RT-qPCR in triplicates, using
SYBR Green chemical dye (Toyobo Life Science, Osaka, Japan).
Relative expression levels of IL-31, IL-31RA,
NLRP1/3/6 and tumor necrosis factor (TNF) were calculated using the
2−ΔΔCq method (21)
normalized to the internal control, β-actin. To quantify cytokine
mRNA, the assays were performed in 2 µg total RNA for reverse
transcription using cDNA synthesis kits (Invitrogen; Thermo Fisher
Scientific, Inc., cat. no. 11756050). Reverse transcription was
performed at 50°C for 30 min. qPCR was performed using a RT-qPCR
kit (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermal cycling conditions used for the assays were as follows: 1
cycle at 94°C for 15 min, followed by 40 cycles at 94°C for 15 sec,
55°C for 30 sec and 72°C for 30 sec. The following primer sequences
were used: IL-31, forward 5′-TCGGTCATCATAGCACATCTGGAG-3′ and
reverse 5′-GCACAGTCCCTTTGGAGTTAAGTC-3′; IL-31RA, forward
5′-AGAATGTTCCAGATACAATGG-3′ and reverse
5′-CGAAGCATGCATACTAAAGGAA-3′; NLRP1, forward
5′-GGACCTCATGGTGGTTACTTTC-3′ and reverse 5′-TCCCAGGGGCCGTAAACTT-3′;
NLRP3, forward 5′-ATTACCCGCCCGAGAAAGG-3′ and reverse
5′-CATGAGTGTGGCTAGATCCAAG-3′; NLRP6, forward
5′-TCTCTCCGTGTCAGCGTTCA-3′ and reverse
5′-CGGAAGAGCCGATTAAAAGTGT-3′; TNF, forward
5′-TCCCCAAAGGGATGAGAAGTTC-3′ and reverse
5′-TCATACCAGGGTTTGAGCTCAG-3′; and β-actin, forward
5′-ATGGGTCAGAAGGACTCCTACG-3′ and reverse
5′-AGTGGTACGACCAGAGGCATAC-3′.
Statistical analysis
Data were presented as the mean ± standard deviation
of three independent experiments. The statistical comparisons
between the different treatments were performed using an unpaired
Student's t-test. Analysis was performed using SPSS software,
Version 22.0 (IBM Corp., Armonk, NY, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
IL-31 protects against experimental
sepsis in vivo
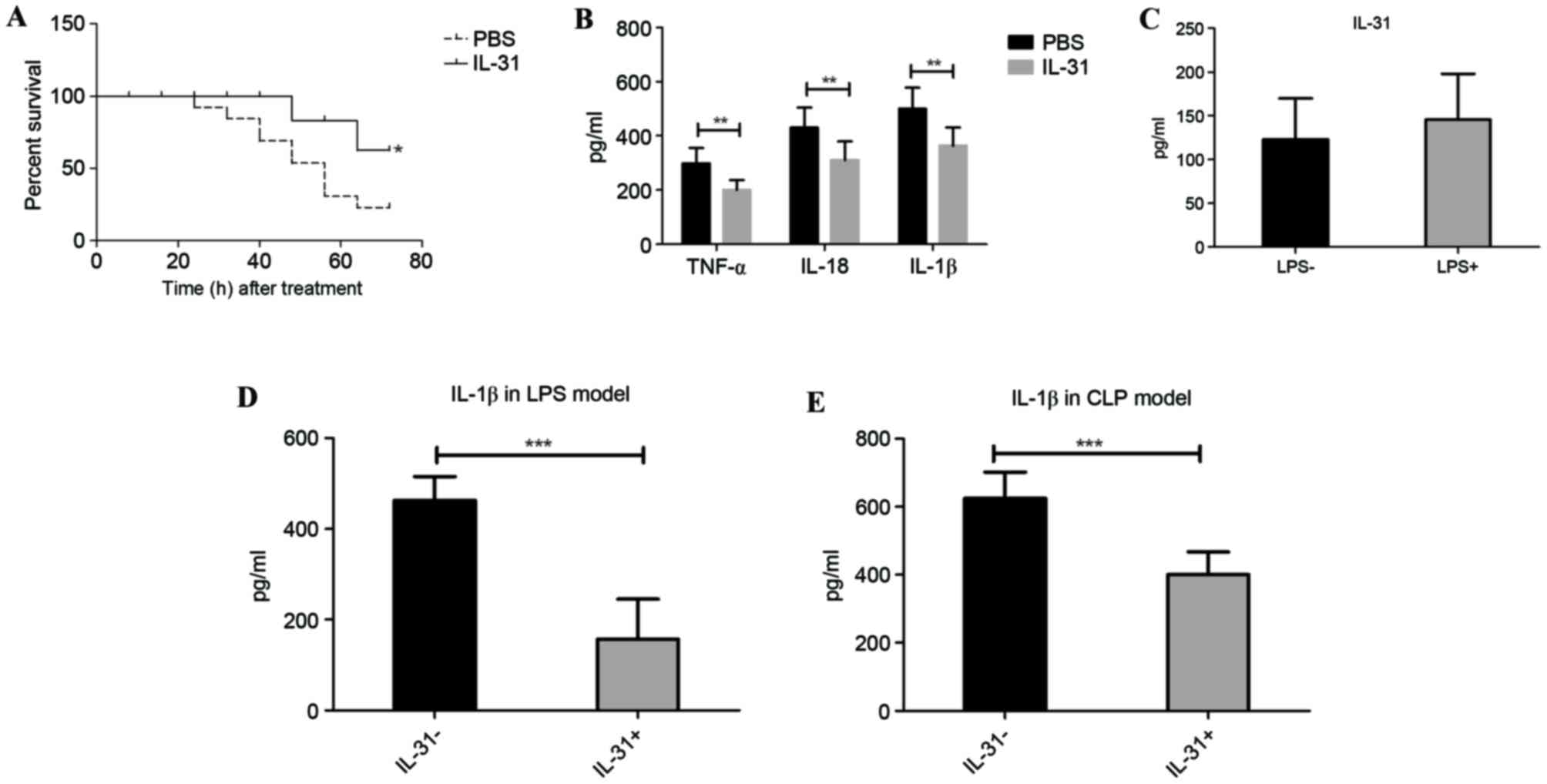
In LPS-induced sepsis, the susceptibility of mice
treated with PBS or IL-31 cytokine was compared. Mice that received
the high dose of LPS and that were treated with recombinant murine
IL-31 injection demonstrated a significantly lower mortality rate
than the vehicle control-treated (PBS) mice from 24 to 72 h
(P=0.0473; Fig. 1A). Furthermore,
the septic shock marker and inflammatory cytokine levels in the
serum, including TNF-α, IL-18 and IL-1β, were significantly
suppressed by IL-31 injection compared with treatment with PBS
(P<0.01; Fig. 1B). There was no
significant difference in IL-31 production following LPS challenge
(Fig. 1C), whereas the IL-1β level
in the peritoneal lavage fluid was significantly downregulated by
IL-31 treatment compared with no IL-31 treatment (P<0.001;
Fig. 1D), suggesting that IL-31 may
regulate IL-1β activation-related signaling in vivo.
Furthermore, to confirm whether IL-31 was protective against
infection and septic shock, IL-31 production in the CLP model of
sepsis was measured. As expected, mice that received IL-31
treatment demonstrated a significant reduction of IL-1β in the CLP
model compared with the vehicle control group (P<0.001; Fig. 1E).
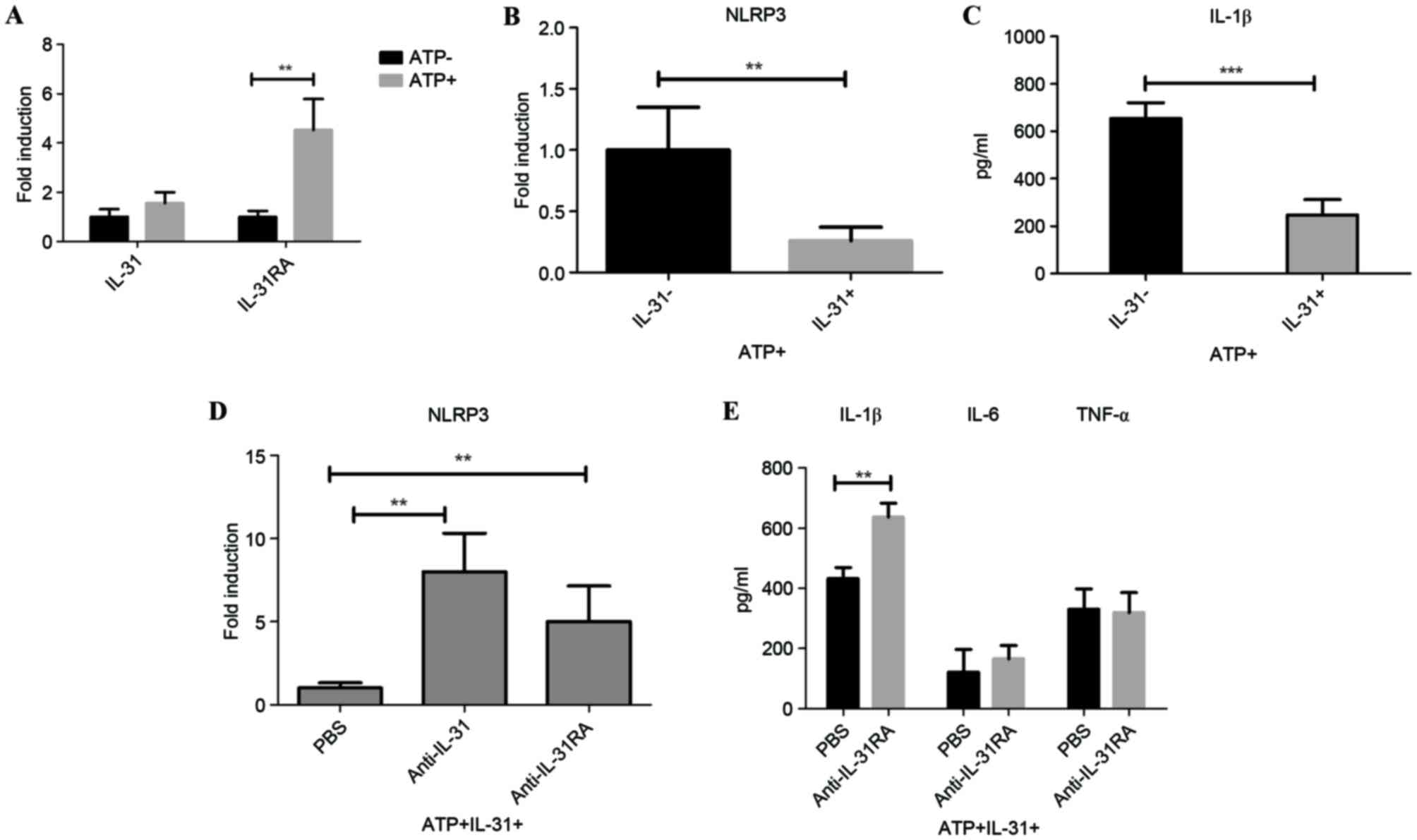
IL-31 regulates IL-1β production by
targeting NLRP3 inflammasome transcription in vitro
As IL-31 may be involved in IL-1β production in
vivo, the role of IL-31 on IL-1β signaling was examined in
vitro. RT-qPCR results demonstrated that IL-31RA mRNA was
significantly induced following ATP stimulation on the peritoneal
macrophages (P<0.01); however, there was no significant change
of IL-31 expression on the cell following ATP induction (Fig. 2A). ATP is an activator of the NLRP3
inflammasome. To analyze the IL-31-IL-1β signaling axis, the in
vitro mRNA expression of NLRP3 following IL-31 treatment was
measured. Results demonstrated that IL-31 was able to significantly
reduce the expression of NLRP3 (P<0.01; Fig. 2B). Furthermore, IL-31 also
significantly decreased LPS-induced IL-1β secretion in ATP-treated
cells (P<0.001; Fig. 2C).
To investigate the role of IL-31 signaling in human
cells, a specific antibody that directly blocks IL-31 (anti-IL-31)
or IL-31R (IL-31RA antibody) was utilized. NLRP3 mRNA expression
increased significantly following IL-31 signal blocking with
neutralizing antibody targeting IL-31/IL-31RA compared with cells
treated with PBS (P<0.01; Fig.
2D), suggesting that IL-31 inhibited IL-1β activation through
decreasing the NLRP3 inflammasome. In concordance with this,
IL-31RA antibody significantly enhanced the level of IL-1β compared
with the level in cells treated with PBS (P<0.01; Fig. 2E), without significantly affecting
the levels of Toll-like receptor signaling-related IL-6 and TNF-α
in the supernatants of the cell culture.
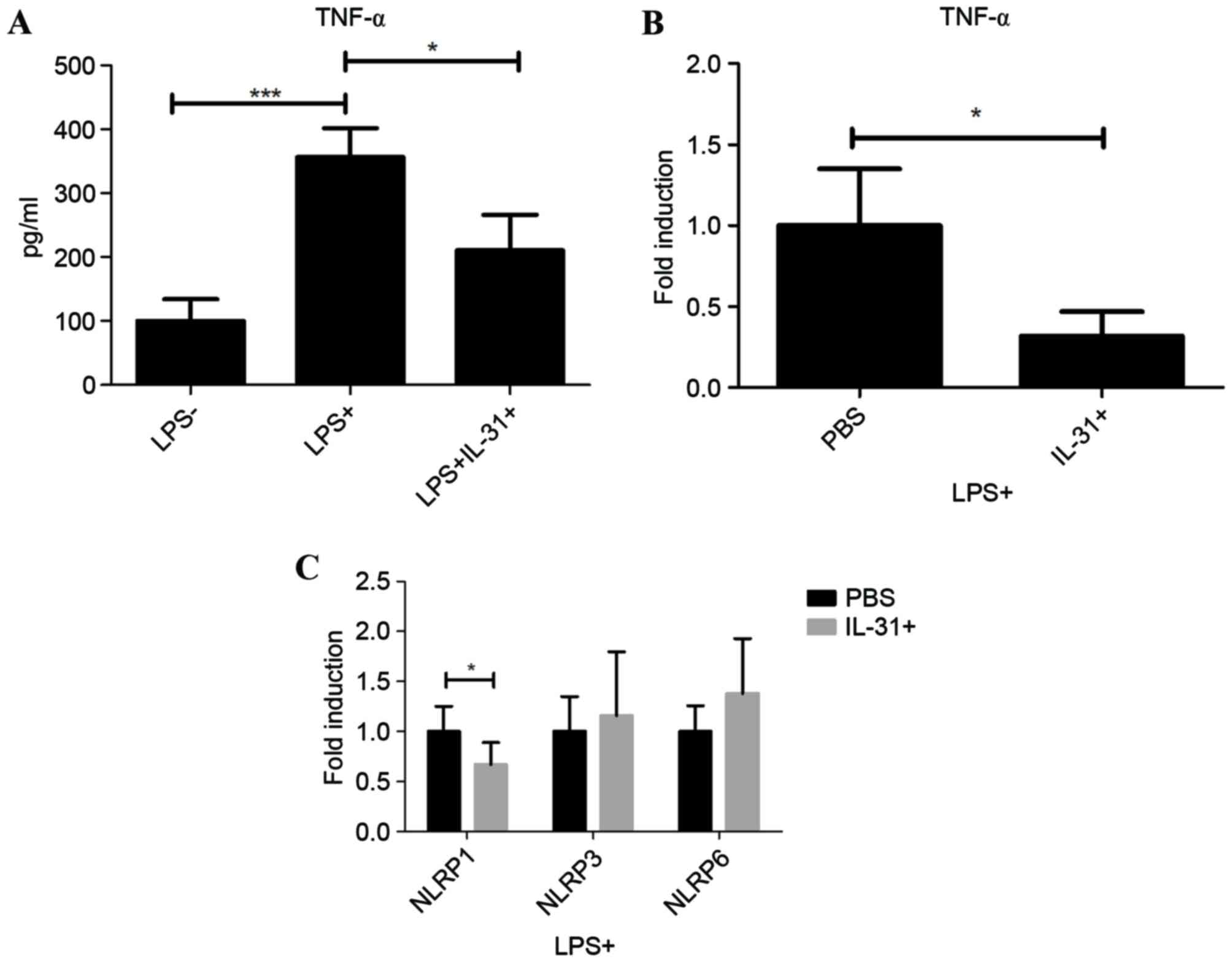
IL-31 modulates NLRP1 in human
PBMC
To systematically investigate the effect of IL-31 on
septic shock, LPS-primed human PBMC from healthy donors were
cultured and treated with IL-31 in vitro. As a typical
inflammatory cytokine in sepsis, TNF-α was significantly induced in
the supernatants by LPS treatment compared with when LPS was not
used (P<0.001), suggesting the activation of the LPS pathway in
human PBMC (Fig. 3A). Following
IL-31 treatment, it was demonstrated that IL-31 was able to
significantly inhibit the protein expression and mRNA expression of
TNF-α following LPS stimulation compared with PBS treatment
(P<0.05; Fig. 3B), suggesting a
regulatory role of IL-31 on PBMC. However, results indicated that
IL-31 negatively regulated the transcription of inflammasome
subtype NLRP1 significantly compared with PBS treatment
(P<0.05), without significantly affecting NLRP3 or NLRP6 in the
macrophages in human PBMC (Fig. 3C).
These results suggested a dual role of IL-31 on the septic shock in
mice and human cells; IL-31 may regulate the NLRP3-IL-1β in the
macrophage and also mediate NLRP1 on the LPS-primed human PBMC,
exhibiting therapeutic effects on septic shock.
Discussion
Currently, a patient is diagnosed with sepsis when
symptoms of SIRS develop, and SIRS results from documented or
suspected infection (22). Although
various clinical studies have identified monitoring markers in
sepsis and respiratory infection (23,24),
effective therapeutic approaches are lacking. One of the earliest
discovered biomarkers used to diagnose infection was C-reactive
protein and procalcitonin, which are often elevated in the serum
during sepsis (3). However, research
has demonstrated that inflammatory cytokines, including IL-6, TNF-α
and IL-1, have important roles in the disease (3). Recently, IL-8 and IL-18 have also been
used as newer candidate biomarkers. It was reported that IL-8 was
identified in genome-wide expression profiling in pediatric septic
shock as an effective predictor of outcome in children with septic
shock who were receiving standard care (25). On the other hand, IL-18 is a cytokine
produced by activated macrophages and is involved in
infection-induced cell immunity (26). However, further study is required to
clarify the utility of this biomarker in the diagnosis of sepsis.
Meanwhile, the role of cytokines in experimental sepsis need to be
elucidated for drug discovery for septic shock
In the present study, novel cytokine IL-31 functions
were identified regarding the immune response. IL-31 was identified
as an inflammatory cytokine induced by activated cluster of
differentiation (CD)4+ T-helper cells and has an
important role in the pathogenesis of atopic dermatitis and
allergic diseases in human eosinophils (8–10). The
glycoprotein 130 (gp130) family constitutes the signaling receptors
for the IL-6/IL-12 family of cytokines, such as IL-6, IL-12, IL-23,
leukemia inhibitory factor and oncostatin M, many of which have
important pro- and anti-inflammatory functions in immune cells
(27). The most recent addition to
this family was the gp130-like monocyte receptor or IL-31RA, which
heterodimerizes with OSMR to form the IL-31R signaling complex
(28). Signaling through IL-31R
primarily results in the phosphorylation of STAT in the JAK-STAT
signaling pathway (29). The ligand
for IL-31R, IL-31, is predominantly expressed by activated the
CD4+ Th2 subset. Therefore, the interactions of
IL-31-IL-31R may regulate various allergic and infectious diseases
(30). For example, anti-IL-31R
antibody has been demonstrated to be a potential therapeutic option
for treating itch and dermatitis in mice; scratching behavior was
inhibited by treatment with anti-IL-31Rα-neutralizing antibody,
BM095 (9). In the lung, macrophages
derived from IL-31RA-knockout mice promoted enhanced
ovalbumin-specific CD4+ T cell proliferation and
purified naive CD4+ T cells from IL-31RA knockout mice
exhibited enhanced proliferation and expression of Th2 cytokines
(13). Furthermore, IL-31RA-knockout
mice also exhibited increased Th2 cytokine responses in the
mesenteric lymph nodes and elevated serum immunoglobulin (Ig)E and
IgG1 levels compared with wild type mice with intestinal helminth
infection (14). IL-31RA-knockout
mice also displayed enhanced goblet cell hyperplasia and a notable
increase in secretion of goblet cell-derived resistin-like molecule
β into the intestinal lumen (14).
In the present study, it was demonstrated that IL-31
treatment was able to rescue the symptoms of septic shock by
reducing inflammatory cytokines, particularly IL-1β in serum and
peritoneal lavage fluid. In vitro data indicated that IL-31
inhibited the expression of NLRP3 in macrophages, thus reducing
IL-1β secretion following LPS treatment. In the human T cell line,
it was demonstrated that anti-human IL-31 neutralizing antibody or
anti-human IL-31RA neutralizing antibody consistently enhanced the
expression of NLRP3 as well as IL-1β, indicating the involvement of
IL-31-IL-31R-NLRP3-IL-1 signaling in ATP-stimulated LPS-mediated
inflammation in vitro. In contrast, IL-31 suppressed TNF
activation through another inflammasome subtype, NLRP1, in human
PBMC, which was a different signaling pathway in this specific cell
type. In conclusion, the present data demonstrated that IL-31 may
be a potential therapeutic target for the treatment of sepsis and
septic shock.
Acknowledgements
Not applicable.
Funding
This study was supported by a grant from the project
of the Natural Science Foundation of Beijing (grant no.
7153169).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XYG and CW designed and performed the experiments,
analyzed the data, and wrote the manuscript. XSZ performed the
evaluation and analysis. FPL, BS and XFZ assisted with performing
the experiments.
Ethics approval and consent to
participate
The animal protocol was approved by the Committee on
the Ethics of Animal Experiments of Capital Medical University
(Beijing, China.).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kumar A, Ellis P, Arabi Y, Roberts D,
Light B, Parrillo JE, Dodek P, Wood G, Kumar A, Simon D, et al:
Initiation of inappropriate antimicrobial therapy results in a
fivefold reduction of survival in human septic shock. Chest.
136:1237–1248. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schuetz P, Maurer P, Punjabi V, Desai A,
Amin DN and Gluck E: Procalcitonin decrease over 72 hours in US
critical care units predicts fatal outcome in sepsis patients. Crit
Care. 17:R1152013. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Standage SW and Wong HR: Biomarkers for
pediatric sepsis and septic shock. Expert Rev Anti Infect Ther.
9:71–79. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dellinger RP, Levy MM, Carlet JM, Bion J,
Parker MM, Jaeschke R, Reinhart K, Angus DC, Brun-Buisson C, Beale
R, et al: Surviving sepsis campaign: International guidelines for
management of severe sepsis and septic shock: 2008. Crit Care Med.
36:296–327. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alqahtani MF, Smith CM, Weiss SL, Dawson
S, Ranaivo Ralay H and Wainwright MS: Evaluation of new diagnostic
biomarkers in pediatric sepsis: Matrix Metalloproteinase-9, tissue
inhibitor of metalloproteinase-1, mid-regional pro-atrial
natriuretic peptide, and adipocyte fatty-acid binding protein. PLoS
One. 11:e01536452016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gucyetmez B and Atalan HK: C-reactive
protein and hemogram parameters for the non-sepsis systemic
inflammatory response syndrome and sepsis: What do they mean? PLoS
One. 11:e01486992016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gearing DP, Comeau MR, Friend DJ, Gimpel
SD, Thut CJ, McGourty J, Brasher KK, King JA, Gillis S, Mosley B,
et al: The IL-6 signal transducer, gp130: An oncostatin M receptor
and affinity converter for the LIF receptor. Science.
255:1434–1437. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hänel KH, Pfaff CM, Cornelissen C, Amann
PM, Marquardt Y, Czaja K, Kim A, Lüscher B and Baron JM: Control of
the physical and antimicrobial skin barrier by an IL-31-IL-1
signaling network. J Immunol. 196:3233–3244. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kasutani K, Fujii E, Ohyama S, Adachi H,
Hasegawa M, Kitamura H and Yamashita N: Anti-IL-31 receptor
antibody is shown to be a potential therapeutic option for treating
itch and dermatitis in mice. Br J Pharmacol. 171:5049–5058.
2014.PubMed/NCBI
|
|
10
|
Hwang JS, Kim GC, Park E, Kim JE, Chae CS,
Hwang W, Lee C, Hwang SM, Wang HS, Jun CD, et al: NFAT1 and JunB
cooperatively regulate IL-31 gene expression in CD4+ T cells in
health and disease. J Immunol. 194:1963–1974. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kasraie S, Niebuhr M and Werfel T:
Interleukin (IL)-31 activates signal transducer and activator of
transcription (STAT)-1, STAT-5 and extracellular signal-regulated
kinase 1/2 and down-regulates IL-12p40 production in activated
human macrophages. Allergy. 68:739–747. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kunsleben N, Rüdrich U, Gehring M, Novak
N, Kapp A and Raap U: IL-31 induces chemotaxis, calcium
mobilization, release of reactive oxygen species, and CCL26 in
eosinophils, which are capable to release IL-31. J Invest Dermatol.
135:1908–1911. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Perrigoue JG, Li J, Zaph C, Goldschmidt M,
Scott P, de Sauvage FJ, Pearce EJ, Ghilardi N and Artis D:
IL-31-IL-31R interactions negatively regulate type 2 inflammation
in the lung. J Exp Med. 204:481–487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Perrigoue JG, Zaph C, Guild K, Du Y and
Artis D: IL-31-IL-31R interactions limit the magnitude of Th2
cytokine-dependent immunity and inflammation following intestinal
helminth infection. J Immunol. 182:6088–6094. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schmidt RL and Lenz LL: Distinct licensing
of IL-18 and IL-1β secretion in response to NLRP3 inflammasome
activation. PLoS One. 7:e451862012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brydges SD, Broderick L, McGeough MD, Pena
CA, Mueller JL and Hoffman HM: Divergence of IL-1, IL-18, and cell
death in NLRP3 inflammasomopathies. J Clin Invest. 123:4695–4705.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
van de Veerdonk FL, Netea MG, Dinarello CA
and Joosten LA: Inflammasome activation and IL-1β and IL-18
processing during infection. Trends Immunol. 32:110–116. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ketelut-Carneiro N, Silva GK, Rocha FA,
Milanezi CM, Cavalcanti-Neto FF, Zamboni DS and Silva JS: IL-18
triggered by the Nlrp3 inflammasome induces host innate resistance
in a pulmonary model of fungal infection. J Immunol. 194:4507–4517.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rittirsch D, Huber-Lang MS, Flierl MA and
Ward PA: Immunodesign of experimental sepsis by cecal ligation and
puncture. Nat Protoc. 4:31–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ding AH, Nathan CF and Stuehr DJ: Release
of reactive nitrogen intermediates and reactive oxygen
intermediates from mouse peritoneal macrophages. Comparison of
activating cytokines and evidence for independent production. J
Immunol. 141:2407–2412. 1988.PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Caserta S, Kern F, Cohen J, Drage S,
Newbury SF and Llewelyn MJ: Circulating plasma microRNAs can
differentiate human sepsis and systemic inflammatory response
syndrome (SIRS). Sci Rep. 6:280062016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Müller B, Becker KL, Schächinger H,
Rickenbacher PR, Huber PR, Zimmerli W and Ritz R: Calcitonin
precursors are reliable markers of sepsis in a medical intensive
care unit. Crit Care Med. 28:977–983. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bozza FA, Salluh JI, Japiassu AM, Soares
M, Assis EF, Gomes RN, Bozza MT, Castro-Faria-Neto HC and Bozza PT:
Cytokine profiles as markers of disease severity in sepsis: A
multiplex analysis. Crit Care. 11:R492007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wong HR, Cvijanovich N, Wheeler DS, Bigham
MT, Monaco M, Odoms K, Macias WL and Williams MD: Interleukin-8 as
a stratification tool for interventional trials involving pediatric
septic shock. Am J Respir Crit Care Med. 178:276–282. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wei XQ, Leung BP, Niedbala W, Piedrafita
D, Feng GJ, Sweet M, Dobbie L, Smith AJ and Liew FY: Altered immune
responses and susceptibility to Leishmania major and Staphylococcus
aureus infection in IL-18-deficient mice. J Immunol. 163:2821–2828.
1999.PubMed/NCBI
|
|
27
|
Klein C, Wüstefeld T, Assmus U, Roskams T,
Rose-John S, Müller M, Manns MP, Ernst M and Trautwein C: The
IL-6-gp130-STAT3 pathway in hepatocytes triggers liver protection
in T cell-mediated liver injury. J Clin Invest. 115:860–869. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin MW, Lee DD, Liu TT, Lin YF, Chen SY,
Huang CC, Weng HY, Liu YF, Tanaka A, Arita K, et al: Novel IL31RA
gene mutation and ancestral OSMR mutant allele in familial primary
cutaneous amyloidosis. Eur J Hum Genet. 18:26–32. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Edukulla R, Singh B, Jegga AG, Sontake V,
Dillon SR and Madala SK: Th2 Cytokines augment IL-31/IL-31RA
interactions via STAT6-dependent IL-31RA expression. J Biol Chem.
290:13510–13520. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lei Z, Liu G, Huang Q, Lv M, Zu R, Zhang
GM, Feng ZH and Huang B: SCF and IL-31 rather than IL-17 and BAFF
are potential indicators in patients with allergic asthma. Allergy.
63:327–332. 2008. View Article : Google Scholar : PubMed/NCBI
|