Introduction
Type 1 diabetes mellitus (T1DM) is an
insulin-dependent type of diabetes (1). One of the key mechanisms of T1DM
development is autoimmune-mediated destruction of the pancreatic β
cells with consequent severe insulin deficiency (2–5).
Patients with T1DM rely on daily insulin injections, while excess
insulin injections may put patients at risk of hypoglycemia.
Therefore, it is necessary to investigate the detailed mechanisms
of T1DM.
MicroRNAs (miRs) are a class of non-coding,
single-stranded small RNAs of 21–25 nucleotides in length, which
regulate the expression of target genes through binding with their
mRNA. Certain miRs are involved in pathological processes,
including kidney diseases (6).
Plasma miRs have been reported to be stable even under harsh
conditions, including boiling, long-term storage at room
temperature, high or low pH and multiple freeze-thaw cycles
(7,8). Various studies have suggested that
certain miRs have key roles in diabetes, including miR-23 and
miR-144 (9,10). miR-192 has a key function in the
formation and progression of kidney diseases (11,12).
Krupa et al (13) indicated
that the reduced expression of serum miR-192 deteriorated kidney
diseases through facilitating the progression of kidney fibrosis.
In addition, a previous study has indicated that miR-192 is
aberrantly expressed in prediabetes (14), suggesting that miR-192 takes part in
the development of diabetes. miR-192 has been reported to have a
key role in DM and cardiovascular diseases (15). However, the detailed function of
miR-192 in T1DM has remained to be elucidated.
Glucagon-like peptide-1 (GLP-1) is a potent insulin
secretagogue and promotes glucose-induced insulin secretion
(16,17). Furthermore, recent studies have
indicated that GLP-1 reduces food intake by interacting with the
hypothalamus (18,19). Several studies using cellular models
of animal origin have investigated the regulation of GLP-1
secretion, including the STC-1 murine enteroendocrine cell line
(20), fetal rat intestinal cell
cultures (21), isolated canine L
cells (22) and GLUTag (23).
The present study indicated that miR-192 was
upregulated in T1DM patients and streptozotocin (STZ)-induced rats,
and the expression of miR-192 was associated with the age and
glucose concentration. miR-192 suppressed the GLP-1 expression in
NCI-H716 cells. In addition, miR-192 inhibited the proliferation of
pancreatic β-cell lines and insulin secretion. Taken together, the
present study indicated that miR-192 has a key function in T1DM and
suggested that miR-192 may serve as a novel molecular therapeutic
target of T1DM.
Materials and methods
Cell line and culture conditions
Human NCI-H716 cells and NIT-1 mouse insulinoma β
cells were obtained from the American Type Culture Collection
(Manassas, VA, USA). NCI-H716 cells were grown in high-glucose
Dulbecco's modified Eagle's medium (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 100 IU/ml penicillin,
100 g/ml streptomycin, 2 mm L-glutamine and 10% fetal bovine serum
(FBS; HyClone; GE Healthcare Life Sciences). NIT-1 cells were
cultured in Ham's F12K medium with 100 IU/ml penicillin, 100 g/ml
streptomycin, 2 mM L-glutamine and 10% FBS.
Animals
A total of 24 male Sprague-Dawley rats (weight,
250–300 g; age, 7–8 weeks) were obtained from the Experimental
Animal Center of The Third Affiliated Hospital of Sun Yat-sen
University (Guangzhou, China). The animal experiments were approved
by the Animal Care Committee of The Third Affiliated Hospital of
Sun Yat-sen University. Mice were housed under a 12 h light/dark
cycle in a temperature (22.0±1.0°C; 0.05% CO2) and
humidity (40–60%) controlled room and were provided with ad
libitum access to rodent chow (Teklad 7001; 4.4%; Envigo Teklad
Global Diets, Huntington, UK) and water. STZ (60 mg/kg;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in 0.1 mol/l citrate
buffer was used to induce Type 1 diabetes mellitus through single
intraperitoneal (i.p) injection as reported previously (24). Control animals received buffer only.
The blood glucose concentration was assessed using a commercial
glucometer (Accu-Chek Sensor; Roche Inc., Mannheim, Germany) at
four days after STZ injection. The animals with blood glucose
levels of >16 mmol/l were considered diabetic and were selected
for the experiments. The STZ+insulin group received 1.5 UI
Novolin®N (Novo Nordisk, Gatwick, West Sussex, UK)
insulin twice a day.
Patients and samples
All patients and healthy volunteers provided written
informed consent. The present study was approved by the Ethics
Committee of The Third Affiliated Hospital of Sun Yat-sen
University. Patients were diagnosed according to the criteria of
the American Diabetes Association (25). A total of 73 patients with T1DM (30
male, 43 female; age range, 9–29 years; average age, 16±3.12
years). Patients were excluded from the present study if that had
clinical cardiovascular disease. Healthy control subjects were
recruited from age- and gender-matched healthy blood donors. Blood
samples were collected from The Third Affiliated Hospital of Sun
Yat-sen University between April 2014 and October 2016 and were
stored at −80°C prior to use.
Cell transfection
The mimics negative control (pre-NC), miR-192 mimics
(pre-miR-192), inhibitor negative control (NC-inh) and miR-192
inhibitors (miR-192 inh) were purchased from Ribobio, Inc
(Guangzhou, China). Prior to transfection, the cells were grown to
70–80% confluence in 6-well plates. The mimics and inhibitor were
transfected using Lipofectamine® 2000 according to the
manufacturer's protocol (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). After transfection for 48 h, the
expression of miR-192 was determined by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
RT-qPCR
Total RNA was extracted from cells or tissues using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The RNA concentration was measured
using a NanoDrop® ND-1000 spectrophotometer (Thermo
Fisher Scientific, Inc.). The complementary (c)DNA was synthesized
using the High Capacity cDNA Reverse Transcription kit (Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
RT-qPCR was performed using SYBR-Green PCR Master Mix (Takara,
Tokyo, Japan) on an ABI 7500 system (Applied Biosystems; Thermo
Fisher Scientific, Inc.). Duplicate samples were normalized to
GAPDH expression. The relative expression of indicated genes was
presented as the mean value and calculated with the formula
2−ΔΔCq (26). The primers
used were as follows: GLP-1 forward, 5′-CATCAAATGCAGACTTGCCA-3′ and
reverse, 5′-ACCTCATTGTTGACAAAGCAG-3′; cyclin D1 forward,
5′-CCTCTAAGATGAAGGAGACCA-3′ and reverse,
5′-AATGAACTTCACATCTGTGGC-3′; cyclin E1 forward,
5′-ATGTTGACTGCCTTGAATTTCC-3′ and reverse,
5′-ACCACTGATACCCTGAAACC-3′; caspase-9 forward,
5′-CTCTTCCTTTGTTCATCTCCT-3′ and reverse,
5′-CAGGATGTAAGCCAAATCTG-3′; GAPDH forward,
5′-ATGATGACATCAAGAAGGTGGT-3′ and reverse,
5′-TTGTCATACCAGGAAATGAGCT-3′. For detecting miR-192 expression,
miR-192 and U6 small nuclear (sn)RNA PCR primers were purchased
from RiboBio (cat. no. miRQ0000517-1-2) and U6 snRNA was used as an
internal control. The cycling conditions for all reactions were as
follows: 98°C for 1 min, 57°C for 90 sec, and 72°C for 90 sec over
32 cycles, followed by a 5-min extension step at 72°C.
Western blot analysis
Following treatment, the protein was extracted from
the cells or tissues using radioimmunoprecipitation assay lysis
buffer (Sangon Biotech, Shanghai, China) for 45 min at 4°C,
followed by centrifugation at 13,000 × g for 10 min at 4°C.
Subsequently, the protein concentration was measured using a
bicinchoninic acid kit (Pierce; Thermo Fisher Scientific, Inc.).
Equal amounts of protein (45 µg) were separated by 10% SDS-PAGE and
then transferred onto polyvinylidene fluoride membranes (Bio-Rad
Laboratories, Hercules, CA, USA). The membranes were blocked with
5% non-fat milk in Tris-buffered saline supplemented with 0.1%
Tween-20 (TBST) for 60 min at room temperature and were
subsequently incubated with the primary antibodies, anti-GLP-1
(cat. no. ab23468; 1:2,000 dilution; Abcam, Cambridge, UK) and
β-actin (cat. no. ab8226; 1:5,000 dilution; Abcam) at 4°C
overnight. The membranes were washed with TBST three times and then
incubated with horseradish peroxidase-conjugated secondary
immunoglobulin G (cat. no. ab97023; 1:5,000 dilution; Abcam) at
room temperature for 60 min. The specific protein bands were
visualized using an enhanced chemiluminescence kit (EMD Millipore,
Billerica, MA, USA) according to the manufacturer's protocol. The
results were quantified using Image J software (version 1.48;
National Institutes of Health, Bethesda, MD, USA).
Assay of extracellular GLP-1
After transfection with miR-192 mimics or
inhibitors, 3×105 NCI-H716 cells were seeded in 6-well
plates with 2 ml medium. After 48 h, the supernatants were
collected and used to analyze the extracellular GLP-1 levels using
an ELISA kit (cat. no. EZGLP1T-36K; EMD Millipore) according to the
protocol of a previous study (27).
Each experiment was repeated three times.
Cell proliferation assay
Following treatment, ~3,000 NIT-1 cells were placed
into 96-well plates and incubated at 37°C with 5% CO2.
Every 24 h, the cell proliferation was detected using a Cell
Counting Kit-8 (CCK-8) kit (Beyotime Institute of Biotechnology,
Haimen, China) according to the manufacturer's instructions. The
absorbance at 450 nm was measured using a Varioskan Flash (Thermo
Fisher Scientific, Inc.). Each experiment was repeated three
times.
Colony formation assay
To assess the effects of miR-192 on the growth of
pancreatic β cells, ~5,000 NIT-1 cells per well were seeded into
6-well plates and incubated for 14 days. The medium was replaced
every two days. After the incubation, the colonies were fixed with
methanol at room temperature for 15 min and then stained with 0.1%
crystal violet (Sigma-Aldrich; Merck KGaA) at room temperature for
15 min. The number of colonies was counted under a light
microscope. Each experiment was repeated three times.
Analysis of cell apoptosis
Annexin V-propidium iodide (PI) staining was used to
detect cell apoptosis. In brief, after transfection of NIT-1 cells
with miR-192 mimics or inhibitors for 48 h, the cells were
collected and washed with PBS for three times. The cells were
stained with 5 µl annexin V-fluorescein isothiocyanate at room
temperature for 20 min in the dark in binding buffer
(Sigma-Aldrich; Merck KGaA), 10 µl propidium iodide was then added
and the cells were incubated at room temperature for 10 min.
Finally, cell apoptosis was assessed using a FACSVerse flow
cytometer (BD Biosciences, San Jose, CA, USA). Each experiment was
repeated three times.
Statistical data analysis
Data analysis was performed with Prism 6 (GraphPad
Inc., La Jolla, CA, USA). Values are expressed as the mean ±
standard deviation. The correlation between the clinicopathological
features of type 1 diabetic patients and miR-192 expression was
assessed using the χ2 test. A two-tailed Student's
t-test was used to analyze differences between two groups. The
correlation between blood glucose levels and miR-192 expression was
determined by Pearson's correlation coefficient. P<0.05 was
considered to indicate a statistically significant difference.
Results
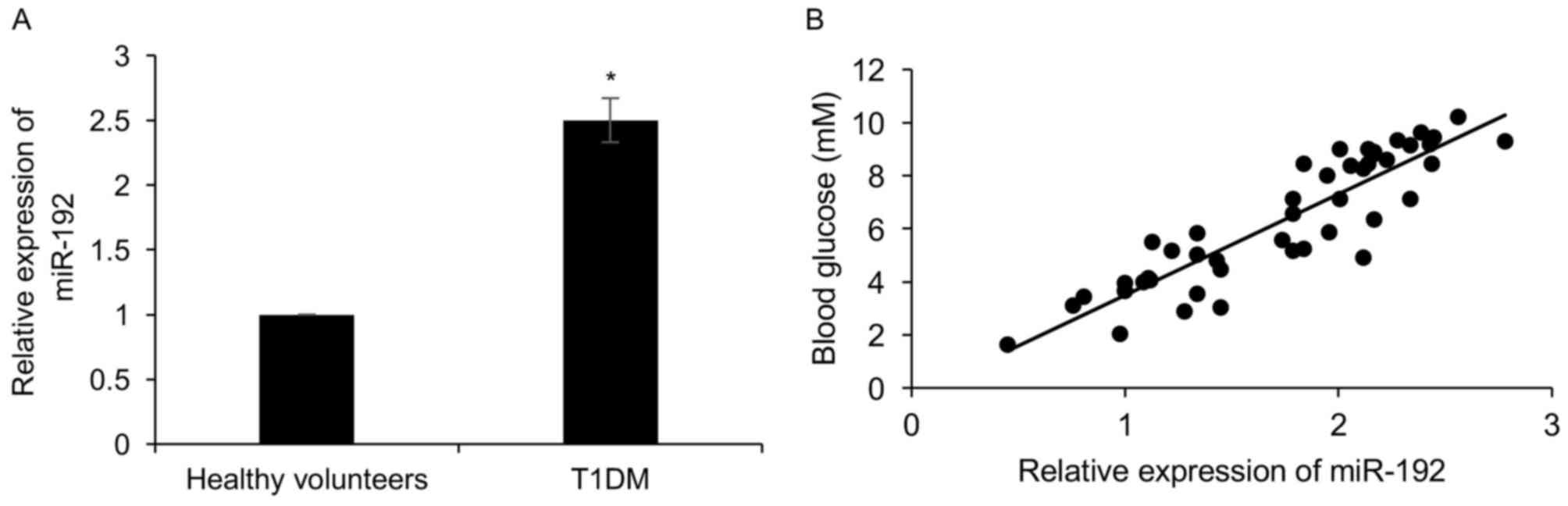
Expression of miR-192 is obviously
upregulated in type 1 diabetic patients
To investigate the role of miR-192 in diabetes,
RT-qPCR analysis was performed to determine the expression of
miR-192 in the plasma of 73 healthy volunteers and 46 T1DM
patients, revealing that compared with the healthy volunteers
group, miR-192 was obviously increased 2.5-fold in T1DM patients
(Fig. 1A). In addition, the patients
with high expression of miR-192 had a higher glucose concentration
than those with low expression of miR-192 (r=0.78; Fig. 1B). Subsequently, the correlation
between the expression of miR-192 and patients' clinicopathological
characteristics was analyzed. As presented in Table I, high expression miR-192 was closely
associated with age, but not with gender (Table I).
 | Table I.Clinicopathological parameters in 73
type 1 diabetic patients stratified by miR-192 expression
status. |
Table I.
Clinicopathological parameters in 73
type 1 diabetic patients stratified by miR-192 expression
status.
|
|
| miR-192
expression |
|
|---|
|
|
|
|
|
|---|
| Parameter | No. (%) | Low (n=25) (%) | High (n=48)
(%) | P-value |
|---|
| Age (years) |
|
|
|
|
|
<20 | 31 (42.5) | 15 (20.5) | 16 (20) | 0.029 |
|
≥20 | 42 (57.5) | 10 (13.7) | 32 (43.8) |
|
| Sex |
|
|
|
|
|
Male | 30 (41.1) | 12 (16.4) | 18 (24.7) | 0.387 |
|
Female | 43 (58.9) | 13 (17.8) | 30 (41.1) |
|
| Glucose
concentration (mM) |
|
|
|
|
|
<7.0 | 24 (32.9) | 16 (21.9) | 8 (11.0) | <0.001 |
|
>7.0 | 49 (67.1) | 9 (12.3) | 40 (54.8) |
|
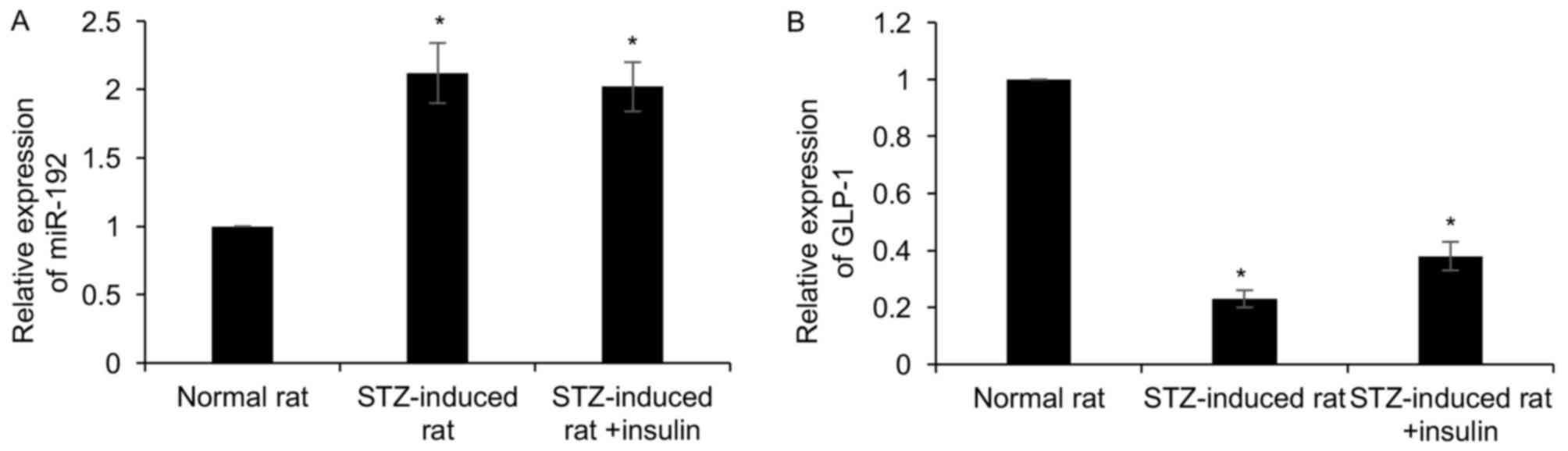
miR-192 is obviously upregulated in an
STZ-induced rat model of T1DM
To determine the role of miR-192 in T1DM,
STZ-induced model rats of T1DM were established. The diabetic
features of the model rats, including plasma blood glucose levels
and body weight, were detected. Blood samples were collected from
the rats at 4 weeks post-STZ injection. Results demonstrated that
body weight decreased and blood glucose levels were improved
following STZ injection; however, an insulin injection only
partially offset the effect of STZ, compared with control group
(Table II). RT-qPCR analysis was
performed to detect the expression of miR-192 compared with that in
normal control rats, which revealed that miR-192 was increased
2.2-fold in STZ-induced rats, but insulin had no effect on miR-192
expression (Fig. 2A). Subsequently,
the expression of GLP-1, a potent insulin secretagogue, was
detected in STZ-induced rats, which suggested that GLP-1 was
downregulated in STZ-induced rats compared with that in normal rats
(Fig. 2B). Of note, insulin
treatment had little effect on GLP-1 expression (Fig. 2B). Taken together, miR-192 was
upregulated in STZ-induced rats, and negatively correlated with the
expression of GLP-1.
| Table II.Body weight and plasma glucose in the
experimental rats (4 weeks). |
Table II.
Body weight and plasma glucose in the
experimental rats (4 weeks).
| Group | No. | Body weight
(g) | Blood glucose
(mmol/l) |
|---|
| Control | 8 | 377.1±18.62 | 8.35±0.19 |
| Diabetic (4
weeks) | 8 |
264.3±31.46a |
23.3±3.31a |
| Diabetic + insulin
(4 weeks) | 8 | 348.3±19.30 | 8.36±2.72 |
miR-192 suppresses GLP-1 expression in
NCI-H716 cells
To identify the downstream target genes of miR-192,
potential targets of miR-192 were searched with a public algorithm,
TargetScan (http://www.targetscan.org). It was
indicated that miR-192 binds to a sequence in the 3′-UTR of GLP-1.
It is known that miRs bind to sequences in the 3′-UTRs of their
target genes, resulting in either inhibition of target mRNA
translation or mRNA degradation. To verify whether miR-192
suppresses GLP-1 expression, possibly by directly binding to the
putative binding site in the 3′-UTR of its mRNA, the effect of
miR-192 on the expression of GLP-1 mRNA and protein was first
detected. For this purpose, human NCI-H716 cells were used
(28), which are widely used to
assess the mechanisms underlying the secretion of GLP-1 (29). The NCI-H716 cell line, derived from a
poorly differentiated adenocarcinoma of human cecum (30), has been described to have certain
endocrine features, in particular the formation of secretory
granules and chromogranin A expression (31). Furthermore, this cell line expresses
several neurohormonal receptors, including receptors for gastrin,
serotonin and somatostatin (32).
NCI-H716 cells were transfected with miR-192 mimics or miR-192
inhibitors, and the expression of miR-192 was determined by RT-qPCR
analysis. As presented in Fig. 3A and
B, miR-192 was overexpressed or knocked down in NCI-H716 cells
transfected with pre-miR-192 or miR-192-inh, respectively (Fig. 3A and B). In addition, the mRNA and
protein levels of GLP-1 were determined by RT-qPCR and western blot
analysis, respectively. The results demonstrated that the mRNA and
protein levels of GLP-1 were downregulated when miR-192 was
overexpressed. Conversely, miR-192 inhibition obviously increased
the expression of GLP-1 (Fig. 3C and
D). The extracellular GLP-1 levels were also assessed, which
demonstrated that ectopic expression of miR-192 decreased the
extracellular GLP-1 levels; however, miR-192 inhibition
significantly increased the extracellular GLP-1 levels (Fig. 3E). The present results therefore
reveal that miR-192 suppresses GLP-1 expression in NCI-H716
cells.
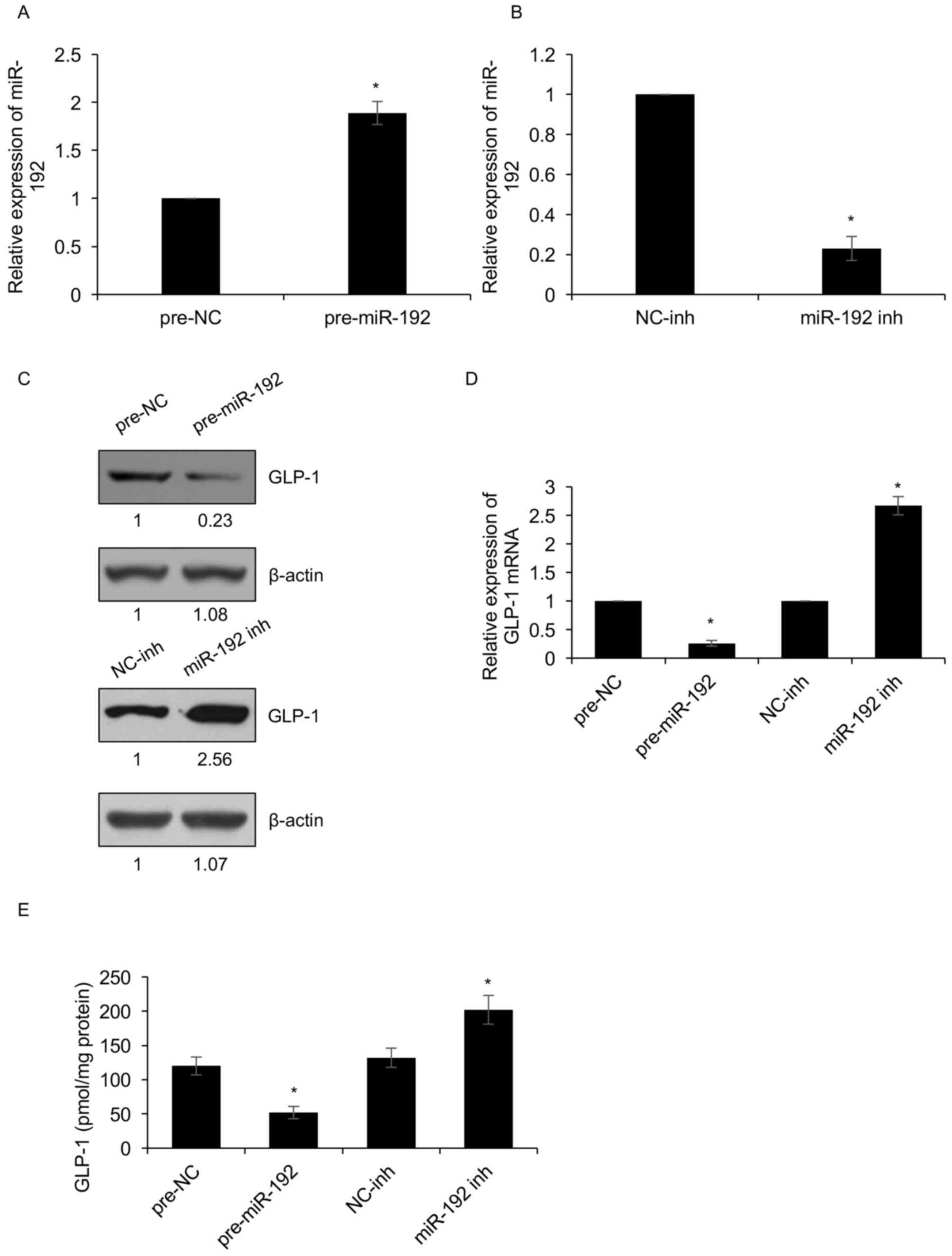 | Figure 3.miR-192 suppresses GLP-1 expression in
NCI-H716 cells. (A and B) Expression of miR-192 in NCI-H716 cells
(A) following transfection with pre-NC or pre-miR-192 and (B)
NC-inh or miR-192 inh for 48 h as determined by RT-qPCR. (C and D)
Protein and mRNA levels of GLP-1 in NCI-H716 cells after
transfection with miR-192 overexpression vector, inhibitor or the
respective controls for 48 h as determined by (C) western blot
analysis and (D) RT-qPCR, respectively. (E) GLP-1 secretion in
NCI-H716 cells treated as above was determined with a GLP-1 ELISA
kit. *P<0.05, pre-miR-192 vs. pre-NC, miR-192 inh vs. NC-inh.
NC, negative control; pre-miR-192, miR-192 mimics; inh, inhibitor;
miR, microRNA; GLP-1, glucagon-like peptide-1; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction. |
miR-192 suppresses the proliferation
of pancreatic β-cell lines and insulin secretion
To further investigate the functions of miR-192 in
the pancreatic β-cell development process, miR-192 was first
overexpressed or knocked down in NIT-1 pancreatic β cells. After
transfection for 48 h, the expression of miR-192 was detected by
RT-qPCR. As presented in Fig. 4A,
miR-192 was markedly increased when NIT-1 cells were transfected
with miR-192 mimics when compared with that in the control groups,
while miR-192 was significantly downregulated when NIT-1 cells were
transfected with miR-192 inhibitors. Subsequently, a CCK-8 assay
was used to determine the effect of miR-192 on cell proliferation,
which revealed that overexpression of miR-192 suppressed cell
proliferation, while inhibition of miR-192 promoted cell
proliferation (Fig. 4B).
Furthermore, a colony formation assay confirmed that ectopic
expression of miR-192 suppressed the cologenicity of NIT-1 cells,
whereas inhibition of miR-192 facilitated it (Fig. 4C). In addition, in response to
glucose stimulation, ectopic expression of miR-192 obviously
decreased the insulin secretion, while inhibition of miR-192
markedly increased it (Fig. 4D).
T1DM is an autoimmune disease, and to detect whether miR-192
regulates apoptosis of pancreatic β-cells, a flow cytometric assay
was performed, which revealed that ectopic expression of miR-192
increased the percentage of apoptotic NIT-1 cells compared with
that in the control groups, whereas miR-192 inhibition decreased
the percentage of apoptotic cells (Fig.
4E). To further investigate the mechanisms by which miR-192
inhibits cell proliferation and induces apoptosis, it was assessed
whether miR-192 regulated the expression of cell cycle- and
apoptosis-associated genes, including cyclin D1, cyclin E1 and
caspase-9. As presented in Fig. 4F,
miR-192 had no effect on the mRNA expression of cyclin D1, cyclin
E1 and caspase-9 (Fig. 4F). Taken
together, the present results reveal that miR-192 suppresses the
proliferation of pancreatic β cells through facilitating cell
apoptosis, thereby reducing insulin secretion.
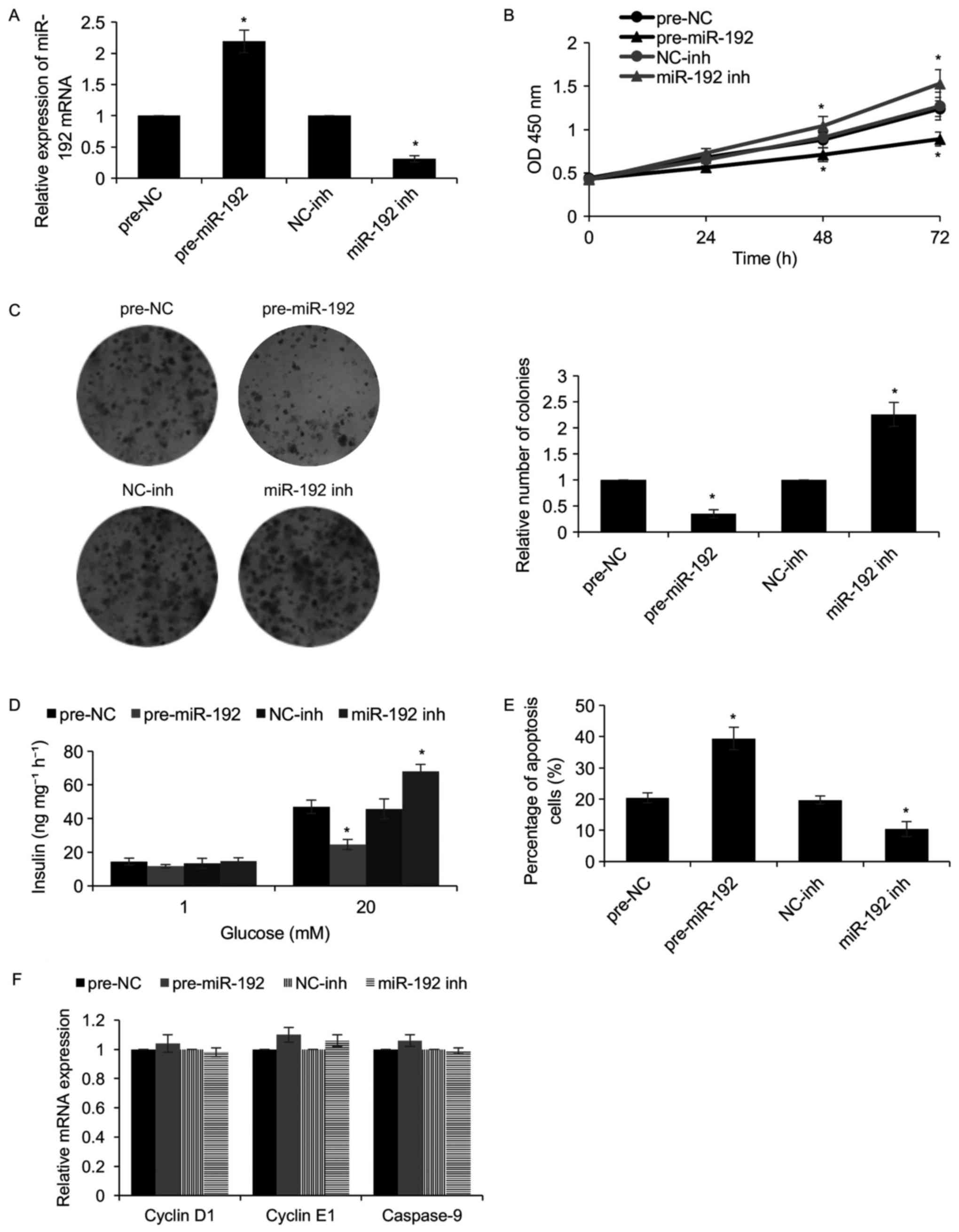 | Figure 4.miR-192 suppresses the proliferation
of a pancreatic β-cell line and the secretion of insulin. miR-192
was overexpressed or knocked down in NIT-1 cells by transfection
for 48 h. (A) The levels of miR-192 were determined by RT-qPCR. (B)
A Cell Counting Kit-8 assay was used to determine the effect of
miR-192 on cell proliferation. (C) A colony formation assay was
used to determine the effect of miR-192 on cell proliferation
(magnification, ×20). (D) The cells were incubated with 1 or 20 mM
glucose for 100 min, and glucose-stimulated insulin secretion was
determined in each group. Each experiment was repeated for three
times. (E) The apoptotic rate in the different groups was assessed
to determine the effect of miR-192 on cell apoptosis. (F) The
expression of cyclin D1, cyclin E1 and caspase-9 was determined by
RT-qPCR. *P<0.05, pre-miR-192 vs. pre-NC, miR-192 inh vs.
NC-inh. NC, negative control; pre-miR-192, miR-192 mimics; inh,
inhibitor; miR, microRNA; GLP-1, glucagon-like peptide-1; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction; OD,
optical density. |
Discussion
T1DM is an insulin-dependent type of diabetes
(1). One of the key
pathophysiological mechanisms of T1DM is the autoimmune-mediated
destruction of pancreatic β-cells with consequent severe insulin
deficiency (2–5).
miR-192 has been identified to have several key
roles in diabetic kidney disease (33); however, the detailed functions of
miR-192 in T1DM have remained to be elucidated.
The present study revealed that miR-192 was
upregulated in T1DM patients and STZ-induced rats. Furthermore,
insulin treatment had no effect on the expression of miR-192 in
STZ-induced rats. In addition, the expression of miR-192 was
closely associated with age and high glucose levels. To further
assess the effect of miR-192 in T1DM, the pancreatic β cell line
NIT-1 was used. A colony formation assay and a CCK-8 assay were
performed to determine the effect of miR-192 on cell proliferation,
which revealed that miR-192 suppressed the proliferation of
pancreatic β cell lines and insulin secretion. Furthermore, miR-192
was indicated to promote β-cell apoptosis. A recent study has
demonstrated that miR-192 suppresses cell proliferation and induces
apoptosis in human rheumatoid arthritis fibroblast-like
synoviocytes and lung cancer (34,35). The
present results were consistent with the previous ones, as it was
indicated that miR-192 promoted T1DM through suppressing pancreatic
β-cell proliferation and facilitating cell apoptosis. However, the
detailed mechanisms by which miR-192 affects cell proliferation and
apoptosis have remained elusive. It was then detected whether
miR-192 regulates certain cell cycle- and apoptosis-associated
proteins, including cyclin D1, cyclin E1 and caspase-9, but as it
appeared to have no effect the mRNA expression of these genes, it
was assumed that miR-192 regulates the proliferation and apoptosis
of pancreatic β cells through other genes/proteins.
GLP-1 is a potent insulin secretagogue and promotes
glucose-induced insulin secretion (16,17). As
NCI-H716 cells were previously used to investigate GLP-1 (29), they were also used in the present
study. Furthermore, NIT-1 cells were employed to investigate the
effect of miR-192 on pancreatic β cells, including their
proliferation and apoptosis. The results indicated that miR-192
suppresses GLP-1 expression, indicating miR-192 promoted T1DM also
through negative regulation of GLP-1. However, the direct binding
of miR-192 to the 3′UTR of GLP-1 mRNA should be assessed using a
luciferase assay in future studies.
In conclusion, to the best of our knowledge, the
present study indicates that miR-192 is upregulated in T1DM and
determined the role of miR-192 in T1DM. miR-192 was identified as a
driver in T1DM. Mechanisms via which miR-192 suppresses pancreatic
β-cell proliferation and insulin secretion were provided, and it
was demonstrated that miR-192 suppresses GLP-1 expression in
NCI-H716 cells. Additionally, miR-192 also suppresses cellular
proliferation and promotes apoptosis in NIT-1 cells. It was
suggested that miR-192 is a novel diagnostic biomarker for
T1DM.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WP and JW conceived and designed the study. WP, YZ,
FX JY and CZ performed the experiments. WP and JY wrote the paper.
WP and JW reviewed and edited the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethical approval and consent to
participate
The animal experiments were approved by the Animal
Care Committee of The Third Affiliated Hospital of Sun Yat-sen
University (Guangzhou, China). All patients and healthy volunteers
provided written informed consent. The patient study has been
approved by the Ethics Committee of The Third Affiliated Hospital
of Sun Yat-sen University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vargas R, Repke JT and Ural SH: Type 1
diabetes mellitus and pregnancy. Rev Obstet Gynecol. 3:92–100.
2010.PubMed/NCBI
|
|
2
|
Achenbach P, Bonifacio E and Ziegler AG:
Predicting type 1 diabetes. Curr Diab Rep. 5:98–103. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Herold KC, Vignali DA, Cooke A and
Bluestone JA: Type 1 diabetes: Translating mechanistic observations
into effective clinical outcomes. Nat Rev Immunol. 13:243–256.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bluestone JA, Herold K and Eisenbarth G:
Genetics, pathogenesis and clinical interventions in type 1
diabetes. Nature. 464:1293–1300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang L and Eisenbarth GS: Prediction and
prevention of Type 1 diabetes mellitus. J Diabet. 3:48–57. 2011.
View Article : Google Scholar
|
|
6
|
Wu H, Kong L, Zhou S, Cui W, Xu F, Luo M,
Li X, Tan Y and Miao L: The role of microRNAs in diabetic
nephropathy. J Diabetes Res. 2014:9201342014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K,
Guo J, Zhang Y, Chen J, Guo X, et al: Characterization of microRNAs
in serum: A novel class of biomarkers for diagnosis of cancer and
other diseases. Cell Res. 18:997–1006. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mitchell PS, Parkin RK, Kroh EM, Fritz BR,
Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Noteboom J, O'Briant
KC, Allen A, et al: Circulating microRNAs as stable blood-based
markers for cancer detection. Proc Natl Acad Sci USA.
105:10513–10518. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Z, Chen H, Si H, Li X, Ding X, Sheng
Q, Chen P and Zhang H: Serum miR-23a, a potential biomarker for
diagnosis of pre-diabetes and type 2 diabetes. Acta Diabetol.
51:823–831. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Karolina DS, Armugam A, Tavintharan S,
Wong MT, Lim SC, Sum CF and Jeyaseelan K: MicroRNA 144 impairs
insulin signaling by inhibiting the expression of insulin receptor
substrate 1 in type 2 diabetes mellitus. PLoS One. 6:e228392011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chung AC, Huang XR, Meng X and Lan HY:
miR-192 mediates TGF-beta/Smad3-driven renal fibrosis. J Am Soc
Nephrol. 21:1317–1325. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kato M, Arce L and Natarajan R: MicroRNAs
and their role in progressive kidney diseases. Clin J Am Soc
Nephrol. 4:1255–1266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Krupa A, Jenkins R, Luo DD, Lewis A,
Phillips A and Fraser D: Loss of MicroRNA-192 promotes fibrogenesis
in diabetic nephropathy. J Am Soc Nephrol. 21:438–447. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Párrizas M, Brugnara L, Esteban Y,
González-Franquesa A, Canivell S, Murillo S, Gordillo-Bastidas E,
Cussó R, Cadefau JA, García-Roves PM, et al: Circulating miR-192
and miR-193b are markers of prediabetes and are modulated by an
exercise intervention. J Clin Endocrinol Metab. 100:E407–E415.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Asgeirsdóttir SA, van Solingen C, Kurniati
NF, Zwiers PJ, Heeringa P, van Meurs M, Satchell SC, Saleem MA,
Mathieson PW, Banas B, et al: MicroRNA-126 contributes to renal
microvascular heterogeneity of VCAM-1 protein expression in acute
inflammation. Am J Physiol Renal Physiol. 302:F1630–F1639. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Drucker DJ: Glucagon-like peptides.
Diabetes. 47:159–169. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Drucker DJ, Philippe J, Mojsov S, Chick WL
and Habener JF: Glucagon-like peptide I stimulates insulin gene
expression and increases cyclic AMP levels in a rat islet cell
line. Proc Natl Acad Sci USA. 84:3434–3438. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Turton MD, O'Shea D, Gunn I, Beak SA,
Edwards CM, Meeran K, Choi SJ, Taylor GM, Heath MM, Lambert PD, et
al: A role for glucagon-like peptide-1 in the central regulation of
feeding. Nature. 379:69–72. 1996. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Flint A, Raben A, Astrup A and Holst JJ:
Glucagon-like peptide 1 promotes satiety and suppresses energy
intake in humans. J Clin Invest. 101:515–520. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Abello J, Ye F, Bosshard A, Bernard C,
Cuber JC and Chayvialle JA: Stimulation of glucagon-like peptide-1
secretion by muscarinic agonist in a murine intestinal endocrine
cell line. Endocrinology. 134:2011–2017. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Brubaker PL and Vranic M: Fetal rat
intestinal cells in monolayer culture: A new in vitro system to
study the glucagon-like immunoreactive peptides. Endocrinology.
120:1976–1985. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Buchan AM, Barber DL, Gregor M and Soll
AH: Morphologic and physiologic studies of canine ileal
enteroglucagon-containing cells in short-term culture.
Gastroenterology. 93:791–800. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brubaker PL, Schloos J and Drucker DJ:
Regulation of glucagon-like peptide-1 synthesis and secretion in
the GLUTag enteroendocrine cell line. Endocrinology. 139:4108–4114.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Remor AP, de Matos FJ, Ghisoni K, da Silva
TL, Eidt G, Búrigo M, de Bem AF, Silveira PC, de León A, Sanchez
MC, et al: Differential effects of insulin on peripheral
diabetes-related changes in mitochondrial bioenergetics:
Involvement of advanced glycosylated end products. Biochim Biophys
Acta. 1812:1460–1471. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
American Diabetes Association, : Diagnosis
and classification of diabetes mellitus. Diabetes Care. 35 Suppl
1:S64–S71. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lo SH, Cheng KC, Li YX, Chang CH, Cheng JT
and Lee KS: Development of betulinic acid as an agonist of TGR5
receptor using a new in vitro assay. Drug Des Devel Ther.
10:2669–2676. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cordier-Bussat M, Bernard C, Levenez F,
Klages N, Laser-Ritz B, Philippe J, Chayvialle JA and Cuber JC:
Peptones stimulate both the secretion of the incretin hormone
glucagon-like peptide 1 and the transcription of the proglucagon
gene. Diabetes. 47:1038–1045. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Reimer RA, Darimont C, Gremlich S,
Nicolas-Métral V, Rüegg UT and Macé K: A human cellular model for
studying the regulation of glucagon-like peptide-1 secretion.
Endocrinology. 142:4522–4528. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Park JG, Oie HK, Sugarbaker PH, Henslee
JG, Chen TR, Johnson BE and Gazdar A: Characteristics of cell lines
established from human colorectal carcinoma. Cancer Res.
47:6710–6718. 1987.PubMed/NCBI
|
|
31
|
de Bruïne AP, Dinjens WN, van der Linden
EP, Pijls MM, Moerkerk PT and Bosman FT: Extracellular matrix
components induce endocrine differentiation in vitro in NCI-H716
cells. The Am J Pathol. 142:773–782. 1993.PubMed/NCBI
|
|
32
|
de Bruïne AP, Dinjens WN, Pijls MM, vd
Linden EP, Rousch MJ, Moerkerk PT, de Goeij AF and Bosman FT:
NCI-H716 cells as a model for endocrine differentiation in
colorectal cancer. Virchows Arch B Cell Pathol Incl Mol Pathol.
62:311–320. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang X, Liu S, Zhang R, Sun B, Zhou S,
Chen R and Yu P: Microribonucleic acid-192 as a specific biomarker
for the early diagnosis of diabetic kidney disease. J Diabetes
Investigat. September 22–2017.(Epub ahead of print).
|
|
34
|
Li S, Jin Z and Lu X: MicroRNA-192
suppresses cell proliferation and induces apoptosis in human
rheumatoid arthritis fibroblast-like synoviocytes by downregulating
caveolin 1. Mol Cell Biochem. 432:123–130. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feng S, Cong S, Zhang X, Bao X, Wang W, Li
H, Wang Z, Wang G, Xu J, Du B, et al: MicroRNA-192 targeting
retinoblastoma 1 inhibits cell proliferation and induces cell
apoptosis in lung cancer cells. Nucleic Acids Res. 39:6669–6678.
2011. View Article : Google Scholar : PubMed/NCBI
|