Introduction
Breast cancer is one of the most prevalent malignant
tumors in females. It typically occurs between the ages of 40 and
60 years and has a high prevalence in females prior to and
following the menopause (1). In
2016, breast cancer accounted for 29% of cancer diagnoses in
females and became the leading cause of female associated mortality
in females in the United States (2,3). The
causes of breast cancer are complex (4), therefore it is necessary to identify a
novel treatment strategy.
Cyclin E1 (CCNE1) gene is a member of the highly
conserved cyclin family. It encodes G1/S-specific cyclin-E1 protein
and is a positive regulator in the cell cycle (5). The overexpression of CCNE1 in tumors
may contribute to tumorigenesis (6,7). CCNE1
is critical in ovarian cancer tumor growth and proliferation
(6). Additionally, it is a novel
translocation partner of the immunoglobulin heavy chain locus and a
novel oncogene in B cell lymphomagenesis (8). CCNE1 was first implicated in breast
cancer (9,10). The abnormal expression of CCNE1 is
associated with high proliferation rates, particularly in the
estrogen receptor-phenotype in breast cancer (11). Stratified analysis has previously
indicated that CCNE1 may be a factor and molecular marker for
breast cancer and survival in the Chinese Han population (11).
It has been demonstrated previously that microRNAs
(miRNAs or miRs) serve a role in the regulation of CCNE1. miR-15a
has been identified as an anti-tumor miRNA in breast cancer via
targeting of CCNE1 (5). In addition,
the human hepatocellular carcinoma cell cycle was arrested at G1 to
S transition by miR-7 and similarly, CCNE1 was identified as its
target gene (12). miR-16-1 also
serves a vital role in the regulation of cell cycle processes and
downregulates the expression of CCNE1 in cervical cancer (13). The present study aimed to investigate
the effects of miR-483-3p on cell growth, apoptosis, migration,
invasion and the cell cycle in breast cancer via targeting
CCNE1.
Materials and methods
Cell culture and transfection
Three breast cancer cell lines, MCF-7, T47D,
MDA-MB-231 and normal breast cell line MCF-10A (America Type
Culture Collection, Manassas, VA, USA), were incubated with
RPMI-1640 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific,
Inc.), in an atmosphere of 5% CO2 at 37°C. Cells were
then seeded in 6-well plates (5×105 cells/well) at a
cell density of 70–90% and transfected using
Lipofectamine® 3000 reagent (Thermo Fisher Scientific,
Inc.) and 50 nM mature miR-483-3p mimics (UCACUCCUCUCCUCCCGUCUU;
Guangzhou RiboBio Co., Ltd., Guangzhou, China) for 48 h according
to the manufacturer's protocols. A non-homologous miRNA mimics
control (cat. no. miR01201-1-5; Guangzhou RiboBio Co., Ltd.) was
used as a negative control.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
T47D, MDA-MB-231, MCF-7 and MCF-10A cells were
cultured in 6 well plates (5×105 cells/well) and total
RNA was isolated using a High Purity Total RNA Extraction kit
(Bioteke Corporation, Beijing, China). Total RNA was then
reverse-transcribed into cDNA using a PrimeScript RT Master Mix
(Takara Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's protocol. qPCR was performed at 95°C for an initial
10 min, followed by 40 cycles of denaturation at 95°C for 30 sec,
annealing at 55°C for 30 sec and extension at 72°C for 30 sec using
SYBR Premix Ex Taq (Takara Biotechnology Co., Ltd.). mRNA relative
expression levels were calculated using the 2−ΔΔCq
method (14). GAPDH was used as the
endogenous control for calculating the gene expression and U6 was
used as a control for microRNA expression. The primers used for
qPCR are presented in Table I. The
expression of miR-483-3p was the lowest in MCF-7 cells. Thus
subsequent experiments were only performed on MCF-7 cells.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|
| CCNE1 |
TTCTTGAGCAACACCCTCTTCTGCAGCC |
TCGCCATATACCGGTCAAAGAAATCTTGTGCC |
| CDK2 |
CTGCTTCCTGTTGGCTCTTTCT |
CTTTGTTTCTGCCTTCTCTCCT |
| NPAT |
TACACAGGTTACTCGACCAAGT |
GCGTCTACCGGGAGACATTAAA |
| U6 |
CTCGCTTCGGCAGCACA |
AACGCTTCACGAATTTGCGT |
| miR-483-3p |
ACACTCCAGCTGGGTCACTCCTCTCCTCC | TGGTGTCGTGGAGTCG |
| GAPDH |
GCACCGTCAAGGCTGAGAAC |
TGGTGAAGACGCCAGTGGA |
Western blot analysis
Total protein was extracted from MCF-7 cells
(1×107) using radioimmunoprecipitation buffer containing
1% phenylmethane sulfonyl fluoride protease inhibitor (Beyotime
Institute of Biotechnology, Haimen, China). A BCA Protein assay kit
was then used to determine protein concentration. Proteins (30
µg/lane) were separated by 10% SDS-PAGE and transferred to a
polyvinylidene difluoride membrane (EMD Millipore, Billerica, MA,
USA). The membrane was then blocked with 3% bovine serum albumin
(Biosharp, Shanghai, China) at room temperature for 2 h and
incubated with primary antibodies against CCNE1 (1:500; cat no.
ab3927), cyclin-dependent kinase 2 (CDK2; 1:1,000; cat no.
ab32147), nuclear protein ataxia-telangiectasia (NPAT; 1:1,000; cat
no. ab70595), phosphorylated (p)-NPAT (1:1,000; cat no. ab70595)
and GAPDH (1:5,000; cat no. ab8245; all Abcam, Cambridge, UK)
overnight at 4°C. Following washing with 1X Tween20 in Tris
buffered solution three times, samples were incubated with a goat
anti-mouse horseradish peroxide (HRP)-conjugated secondary antibody
(1:5,000; cat no. ab205719) or goat anti-rabbit HRP-conjugated
secondary antibody (1:5,000; cat no. ab205718; both Abcam) at room
temperature for 2 h. The membrane was visualized using an enhanced
chemiluminescence kit (Beyotime Institute of Biotechnology, Haimen,
China).
Cell proliferation assay
MCF-7 cells (1×105) were seeded in a
96-well plate and cultured at 37°C with 5% CO2 and
transfected with miRNA mimics or negative controls. A cell counting
kit (CCK)-8 kit (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was
subsequently used to measure cell proliferation. Briefly, following
transfection and incubation at 37°C for 24 h, 10 µl CCK-8 solution
was added to each well of the plate. Following 4 h incubation,
absorbance was measured at a wavelength of 450 nm using a
microplate reader.
Wound healing assay
The migration ability of MCF-7 cells was detected
using a wound healing assay. MCF-7 cells (5×105/well)
were seeded in 6-well dishes. Following transient transfection,
cells were incubated for 12 h in a humidified incubator at 37°C
with 5% CO2. A ‘wound’ was created in each culture using
a sterilized 1 ml pipette. The debris was washed with PBS and cells
were incubated with serum-free RPMI-1640 medium at 37°C for 48 h.
The cell migration area was quantified by observing images taken
under a light microscope (Olympus Corporation, Tokyo, Japan) at a
magnification of ×100. Images were analyzed by ImageJ software
(version 1.48; National Institutes of Health, Bethesda, MD, USA).
Migration was quantified by counting the cell numbers from five
fields in the wound area. Experiments were carried out in
triplicate, at least three times.
Migration assay
MCF-7 cell migration was investigated using 24-well
plates. Briefly, following transfection, 2.5×105 cells
were resuspended and incubated in RPMI-1640 medium without serum
and seeded in the upper chamber of a Transwell insert at 37°C for
48 h. RPMI 1640 containing FBS medium was added to the lower
chamber. Cells were fixed using 4% paraformaldehyde at room
temperature for 30 min and stained using 0.1% crystal violet
(Sigma-Aldrich; Merck KGaA) at room temperature for 20 min. The
migratory status of MCF-7 cells was observed under a light
microscope at a magnification of ×200 and assessed by the relative
number of migrated cells.
Bioinformatics, vector construction
and dual luciferase reporter assay
The sequence of CCNE1 3′-untranslated region (UTR)
was predicted to interact with miR-483-3p using TargetScan
(www.targetscan.org/vert_71/). The
fragments of CCNE1, containing a wild-type 3′UTR
(CCAACAUGGUGAAACCCCGUCUC) or mutated (CCAACAUGGUGAAAC
UUUAGCUC), were inserted into the psiCHECK™−2
vector (Promega Corporation, Madison, WI, USA). Then MCF-7 cells
were transfected using Lipofectamine® 3,000 reagent
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocols. Following 24 h incubation of MCF-7 cells in RPMI-1640
medium at 37°C, luciferase activity was measured using the Dual-Glo
luciferase assay system (Promega Corporation). Renilla
luciferase activity served as the internal control.
Cell cycle assay
Following transfection, MCF-7 cells were harvested
when they reached 80% confluence and fixed with 70% ice-cold
ethanol at 4°C for 1 h. Cells were washed with PBS and stained with
0.1% (m/v) propidium iodide (BD Biosciences, Franklin Lakes, NJ,
USA) in the dark at room temperature for 1 h. The cell cycle was
analyzed by flow cytometry using a FACSCalibur flow cytometer (BD
Biosciences). The percentages of cells in the G0/G1, S and G2/M
phases were then determined by FlowJo software (version 7.6.1;
FlowJo LLC, Ashland, OR, USA).
Statistical analysis
Data are expressed as the mean ± standard error of
the mean and experiments were repeated in triplicate. Statistical
differences were identified using Student's t-test or one-way
analysis of variance followed by the Dunnett's t-test. Statistical
analysis was performed using SPSS 20.0 (IBM Corp., Armonk, NY,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-483-3p is downregulated in breast
cancer cells
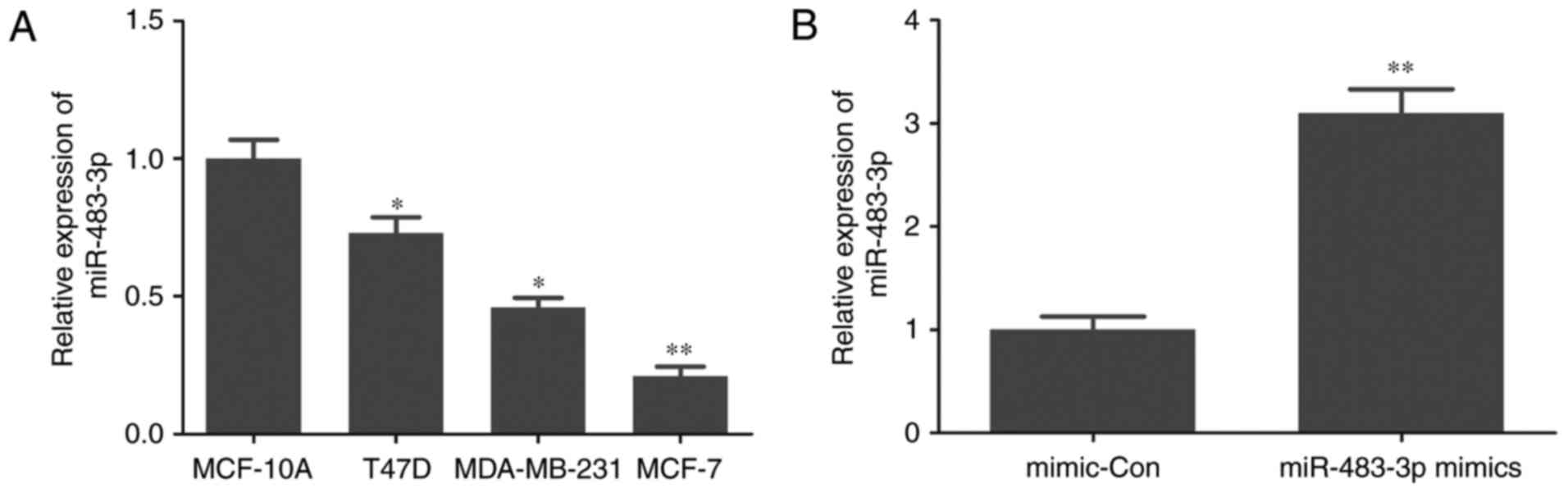
RT-qPCR was performed to determine the level of
miR-483-3p expression in normal breast cell and breast cancer
cells. The expression of miR-483-3p was significantly reduced in
T47D, MDA-MB-231 and MCF-7 breast cancer cells compared with
MCF-10A normal breast cells (Fig.
1A). These results suggest that miR-483-3p may serve a role in
breast cancer progression.
miR-483-3p decreases MCF-7 cell
proliferation
The effect of miR-483-3p on cell proliferation in
breast cancer was investigated by transfecting MCF-7 cells with
miR-483-3p mimics (Fig. 1B). The
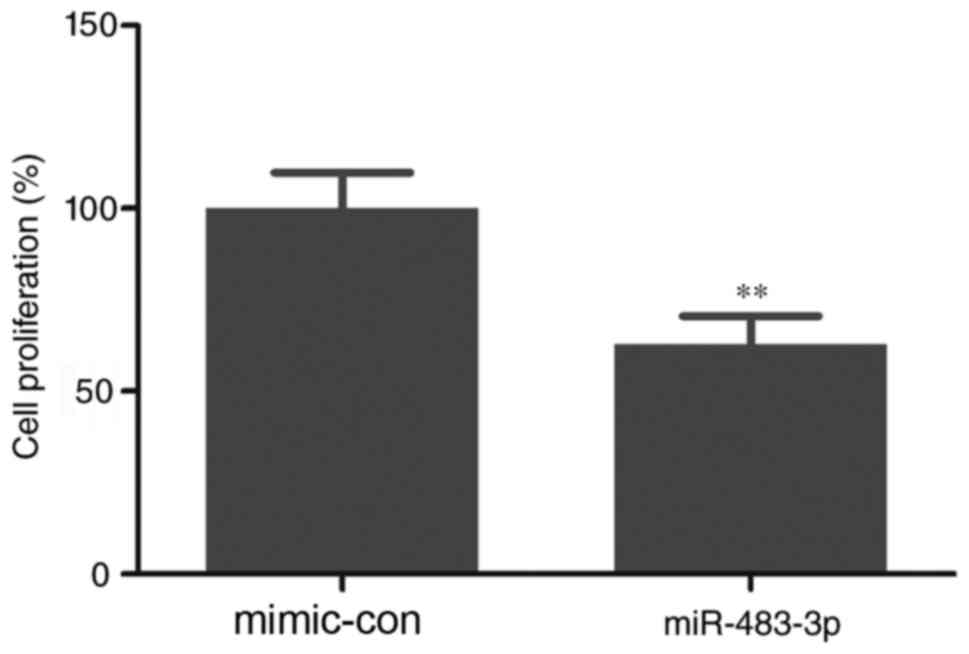
results of the CCK-8 assay indicated that overexpression of
miR-483-3p significantly inhibited cell proliferation compared with
the mimic control group (Fig. 2). In
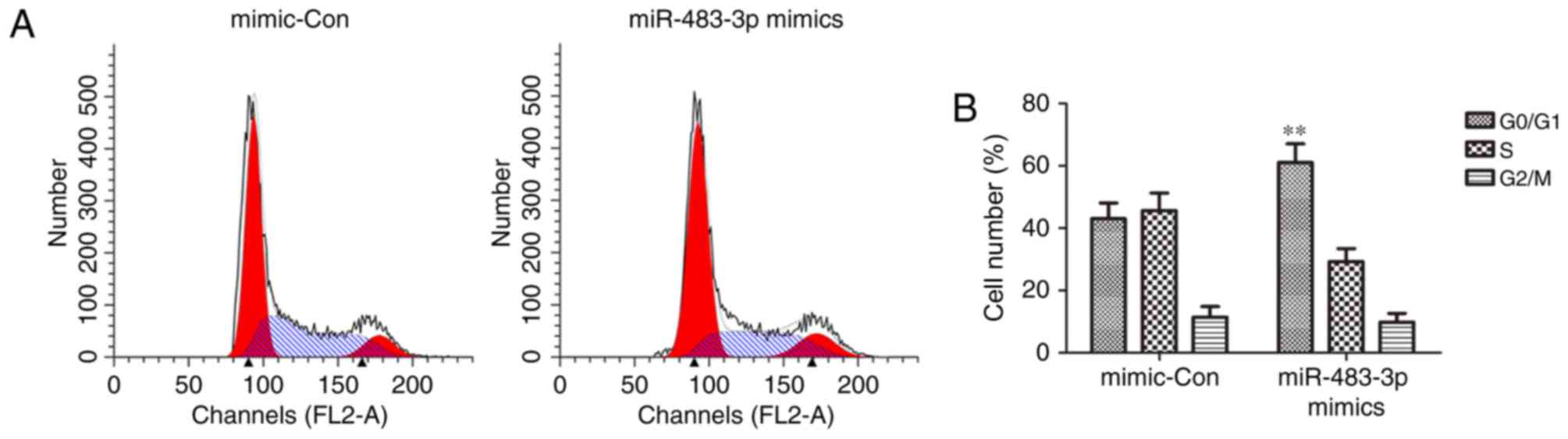
addition, the cell cycle assay demonstrated that overexpression of
miR-483-3p significantly retained MCF-7 cells in the G1 phase,
preventing them from entering the S phase (Fig. 3). These results suggest that
miR-483-3p regulates MCF-7 cell proliferation.
miR-483-3p downregulates the
expression of CCNE1, CDK2, NPAT and p-NPAT
RT-qPCR and western blotting were used to measure
the expression of CCNE1, CDK2 and p-NPAT following transfection, to
further analyze the regulation of the cell cycle by miR-483-3p in
MCF-7 cells. The results of RT-qPCR demonstrated that miR-483-3p
overexpression significantly downregulated the expression of CCNE1,
CDK2 and NPAT mRNA compared with that in the mimic control group
(Fig. 4A-C). The results of western
blotting also suggested that miR-483-3p overexpression markedly
downregulated the expression of CCNE1, CDK2, NPAT and p-NPAT
protein compared with the mimic control group (Fig. 4). These results, therefore indicated
that miR-483-3p suppressed MCF-7 cell proliferation.
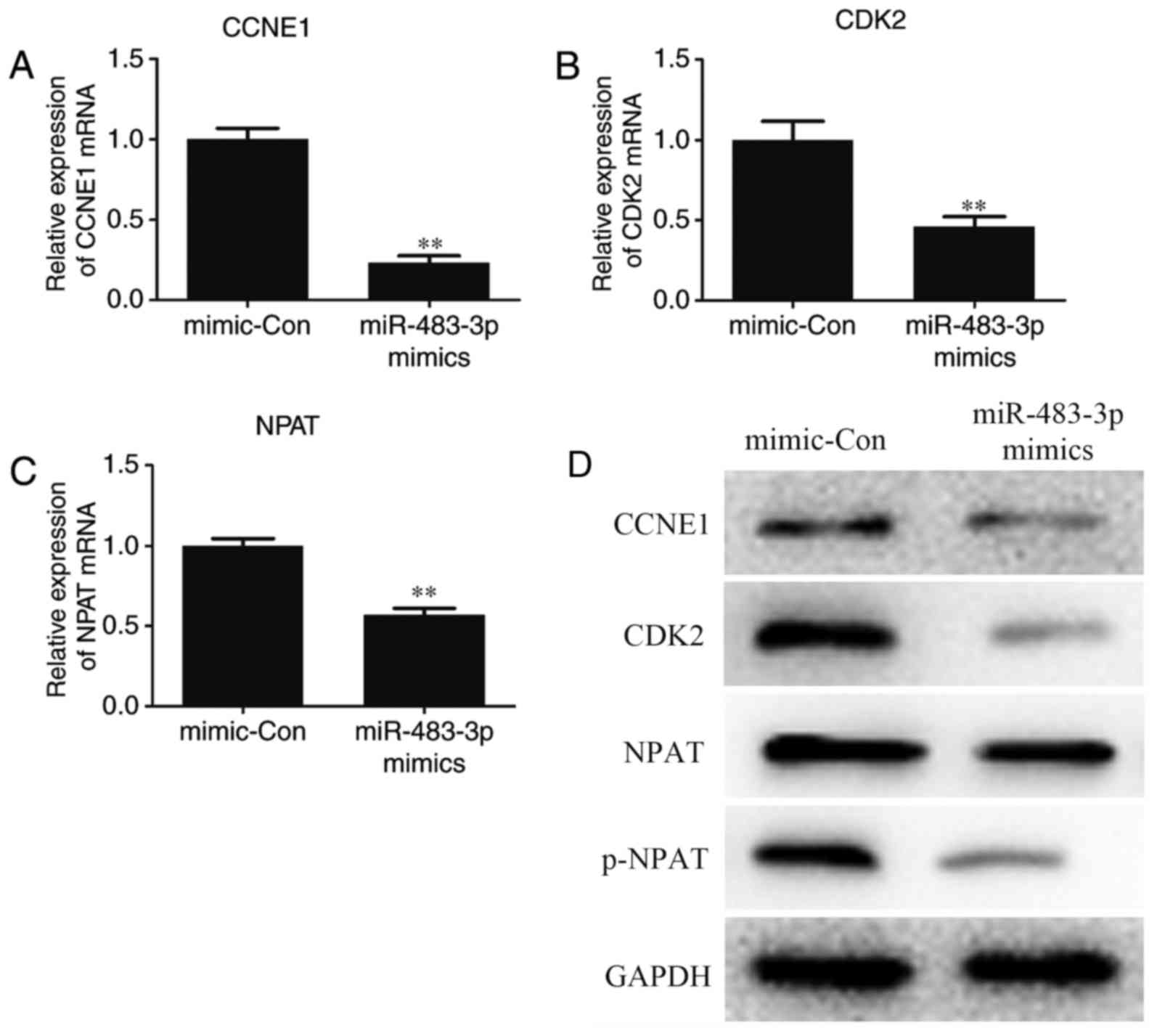 | Figure 4.Overexpression of miR-483-3p inhibits
the expression of CCNE1, CDK2, and NPAT mRNA and protein. MCF-7
cells were transfected with miR-483-3p mimics. The expression of
(A) CCNE1 (B) CDK2 and (C) NPAT mRNA was measured using reverse
transcription-quantitative polymerase chain reaction. (D) The
expression of CCNE1, CDK2, NPAT and p-NPAT protein was measured
using western blotting. Data are expressed as the mean ± standard
error of the mean and experiments were repeated in triplicate.
**P<0.01 vs. the mimic-con group. miR-483-3p, microRNA-483-3p;
CCNE1, cyclin E1; CDK2, cyclin-dependent kinase 2; NPAT, nuclear
protein ataxia-telangiectasia; p, phosphorylated mimic-con,
mimic-control group. |
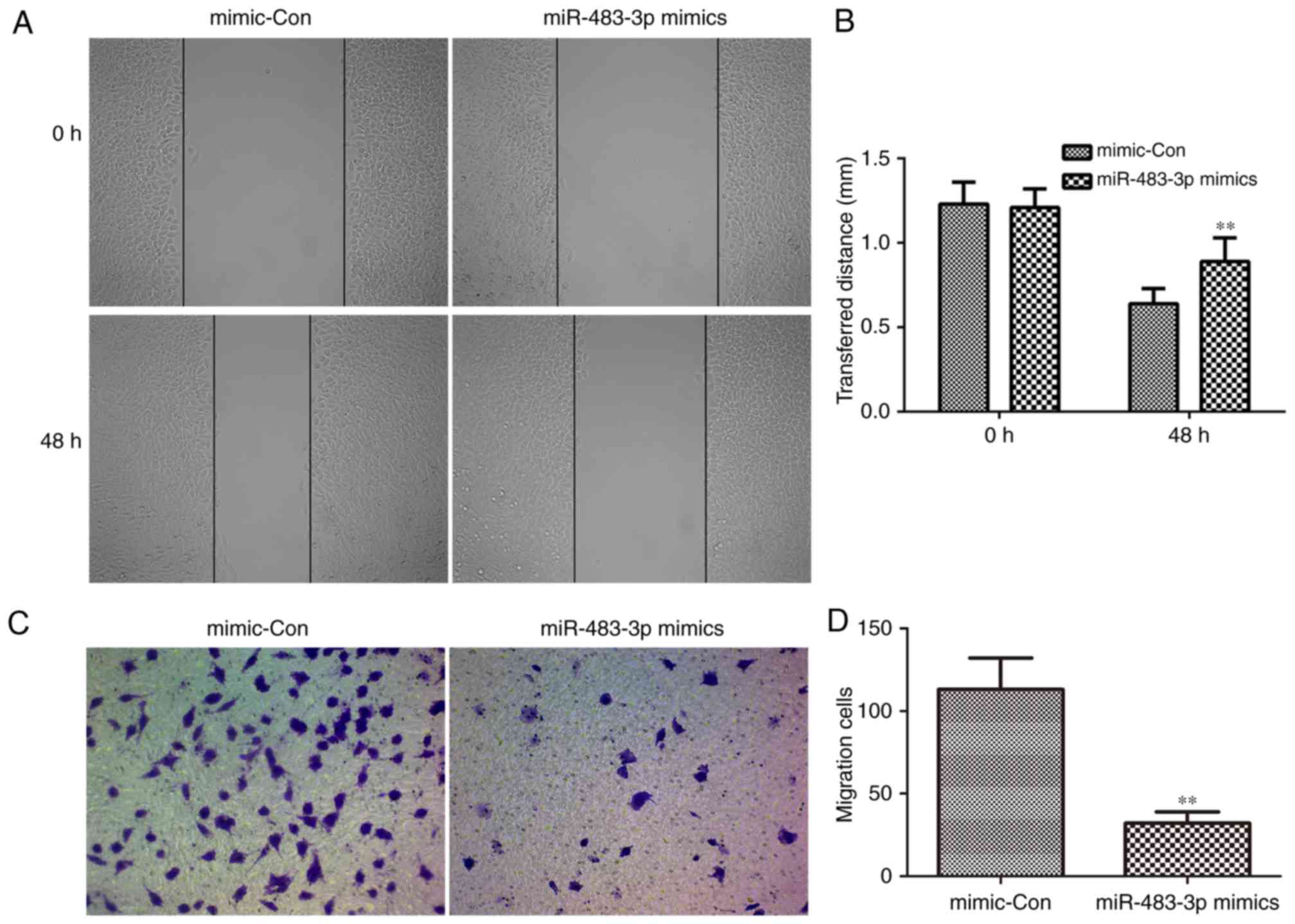
miR-483-3p inhibits the migration
ability of MCF-7 cells
The role of miR-483-3p in the migration of breast
cancer cells was demonstrated using wound healing and Transwell
assays to observe the migration of MCF-7 cells following
transfection. miR-483-3p overexpression significantly inhibited the
wound healing ability of MCF-7 cells compared with the mimic
control group (Fig. 5A and B). In
addition, miR-483-3p overexpression also significantly reduced the
migration ability of MCF-7 cells compared with the mimic control
group (Fig. 5C and D). These
findings indicated that miR-483-3p attenuates the migration ability
of MCF-7 cells.
CCNE1 is the target of miR-483-3p
The molecular mechanism of miR-483-3p in regulating
cell proliferation was determined by predicting the targets of
miR-483-3p using TargetScan. The results indicated that CCNE1 was a
target of miR-483-3p (Fig. 6A). A
dual luciferase reporter assay demonstrated that miR-483-3p
overexpression significantly reduced dual-luciferase activity in
the wild type group compared with the mutant group (Fig. 6B). These results therefore confirmed
that CCNE1 is a target of miR-483-3p.
Discussion
The role of miRs as tumor suppressors targeting
molecules and signaling pathways in breast cancer, including the
lethal-7, miR-125, miR-200, miR-205 and miR-206 families, has been
previously suggested (15).
miR-483-3p has also been reported to be associated with breast
cancer and may be a featured biomarker in breast cancer when using
an miRNA microarray (16).
miR-483 was initially identified in the human fetal
liver (16) and is located within
intron 2 of the insulin-like growth factor 2 locus (17). It serves important roles in the
mechanism of many diseases, including alcoholic hepatitis (18), diabetic cardiomyopathy (19), ovarian carcinoma (20) and pancreatic cancer (21). In the present study, the effect of
miR-483-3p in breast cancer was investigated and the results
demonstrated that miR-483-3p was downregulated in breast cancer
cell lines, especially in MCF7 cells. These results suggest the
potential tumor suppressor function of miR-483-3p. Therefore, MCF7
cells were transfected with miR-483-3p mimics and it was
demonstrated that overexpression of miR-483-3p decreased cell
proliferation and suppressed cell migration.
It was also demonstrated that miR-483-3p
overexpression suppressed MCF-7 cells in the G1 phase from entering
the S phase. CCNE1 forms a complex with CDK2 and CCNE1-CDK2
regulates the cell cycle, as it is a key kinase complex in G1-S
transition (22). It induces cells
to initiate DNA synthesis via phosphorylation of their downstream
substrates, including NPAT, thereby allowing cells to irreversibly
enter the S phase (23). Hence, to
further investigate the effects of miR-483-3p overexpression on the
cell cycle, the expression of CCNE1, CDK2 and p-NPAT protein was
detected using western blot analysis. The results demonstrated that
miR-483-3p overexpression markedly reduced the expression of CCNE1,
CDK2 and p-NPAT protein.
Cyclin E1 is encoded by the CCNE1 gene (Gene ID:
898) and belongs to the cyclin family (11). Bioinformatics analysis predicted that
the CCNE1 gene was a target gene of miR-483-3p. In addition, the
results of the dual-luciferase reporter assay indicated that CCNE1
is a direct target of miR-483-3p.
In conclusion, the results of the present study
indicated that miR-483-3p is a key factor in breast cancer and
inhibits G1-S transition in the cell cycle of MCF-7 cells. These
findings suggest that miR-483-3p has a tumor suppression role in
breast cancer and CCNE1 is its direct target. Therefore, miR-483-3p
may be a novel therapy for the treatment of breast cancer.
References
|
1
|
Dodiyi-Manuel A and Wakama IE:
Predispositions of carcinoma of the breast: A review. Niger J Med.
23:7–12. 2014.PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Panieri E: Breast-cancer awareness in
low-income countries. Lancet Oncol. 14:274–275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang BN, Cao XC, Chen JY, Chen J, Fu L,
Hu XC, Jiang ZF, Li HY, Liao N, Liu DG, et al: Guidelines on the
diagnosis and treatment of breast cancer (2011 edition). Gland
Surg. 1:39–61. 2012.PubMed/NCBI
|
|
5
|
Luo Q, Li X, Li J, Kong X, Zhang J, Chen
L, Huang Y and Fang L: MiR-15a is underexpressed and inhibits the
cell cycle by targeting CCNE1 in breast cancer. Int J Oncol.
43:1212–1218. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakayama N, Nakayama K, Shamima Y,
Ishikawa M, Katagiri A, Iida K and Miyazaki K: Gene amplification
CCNE1 is related to poor survival and potential therapeutic target
in ovarian cancer. Cancer. 116:2621–2634. 2010.PubMed/NCBI
|
|
7
|
Coradini D, Boracchi P, Oriana S,
Biganzoli E and Ambrogi F: Differential expression of genes
involved in the epigenetic regulation of cell identity in normal
human mammary cell commitment and differentiation. Chin J Cancer.
33:501–510. 2014.PubMed/NCBI
|
|
8
|
Nagel I, Akasaka T, Klapper W, Gesk S,
Böttcher S, Ritgen M, Harder L, Kneba M, Dyer MJ and Siebert R:
Identification of the gene encoding cyclin E1 (CCNE1) as a novel
IGH translocation partner in t(14;19)(q32;q12) in diffuse large
B-cell lymphoma. Haematologica. 94:1020–1023. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Keyomarsi K and Pardee AB: Redundant
cyclin overexpression and gene amplification in breast cancer
cells. Proc Natl Acad Sci USA. 90:1112–1116. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sutherland RL and Musgrove EA: Cyclins and
breast cancer. J Mammary Gland Biol Neoplasia. 9:95–104. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Han JY, Wang H, Xie YT, Li Y, Zheng LY,
Ruan Y, Song AP, Tian XX and Fang WG: Association of germline
variation in CCNE1 and CDK2 with breast cancer risk, progression
and survival among Chinese Han women. PLoS One. 7:e492962012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang X, Hu S, Zhang X, Wang L, Zhang X,
Yan B, Zhao J, Yang A and Zhang R: MicroRNA-7 arrests cell cycle in
G1 phase by directly targeting CCNE1 in human hepatocellular
carcinoma cells. Biochem Biophys Res Commun. 443:1078–1084. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zubillaga-Guerrero MI, Ldel Alarcon-Romero
C, Illades-Aguiar B, Flores-Alfaro E, Bermúdez-Morales VH, Deas J
and Peralta-Zaragoza O: MicroRNA miR-16-1 regulates CCNE1 (cyclin
E1) gene expression in human cervical cancer cells. Int J Clin Exp
Med. 8:15999–16006. 2015.PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Asghari F, Haghnavaz N, Baradaran B,
Hemmatzadeh M and Kazemi T: Tumor suppressor microRNAs: Targeted
molecules and signaling pathways in breast cancer. Biomed
Pharmacother. 81:305–317. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang M, Liu D, Li W, Wu X, Gao C and Li
X: Identification of featured biomarkers in breast cancer with
microRNA microarray. Arch Gynecol Obstet. 294:1047–1053. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Veronese A, Lupini L, Consiglio J, Visone
R, Ferracin M, Fornari F, Zanesi N, Alder H, D'Elia G, Gramantieri
L, et al: Oncogenic role of miR-483-3p at the IGF2/483 locus.
Cancer Res. 70:3140–3149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu H, French BA, Li J, Tillman B and
French SW: Altered regulation of miR-34a and miR-483-3p in
alcoholic hepatitis and DDC fed mice. Exp Mol Pathol. 99:552–557.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qiao Y, Zhao Y, Liu Y, Ma N, Wang C, Zou
J, Liu Z, Zhou Z, Han D, He J, et al: miR-483-3p regulates
hyperglycaemia-induced cardiomyocyte apoptosis in transgenic mice.
Biochem Biophys Res Commun. 477:541–547. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Arrighetti N, Cossa G, De Cecco L, Stucchi
S, Carenini N, Corna E, Gandellini P, Zaffaroni N, Perego P and
Gatti L: PKC-alpha modulation by miR-483-3p in platinum-resistant
ovarian carcinoma cells. Toxicol Appl Pharmacol. 310:9–19. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hao J, Zhang S, Zhou Y, Hu X and Shao C:
MicroRNA 483-3p suppresses the expression of DPC4/Smad4 in
pancreatic cancer. FEBS Lett. 585:207–213. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Deng M, Li F, Ballif BA, Li S, Chen X, Guo
L and Ye X: Identification and functional analysis of a novel
cyclin e/cdk2 substrate ankrd17. J Biol Chem. 284:7875–7888. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao J, Kennedy BK, Lawrence BD, Barbie
DA, Matera AG, Fletcher JA and Harlow E: NPAT links cyclin E-Cdk2
to the regulation of replication-dependent histone gene
transcription. Genes Dev. 14:2283–2297. 2000. View Article : Google Scholar : PubMed/NCBI
|