Introduction
Coronary atherosclerosis is a chronic disease
characterized by ongoing progression in response to systemic risk
factors and local pro-atherogenic stimuli (1). Atherosclerosis is the primary cause of
coronary heart disease, cerebral infarction and peripheral vascular
disease (2). Notably,
atherosclerotic cardiovascular disease remains the leading cause of
human fatality worldwide (3).
Vascular smooth muscle cells (VSMCs) have a critical role in
atherosclerotic plaque formation. Abnormal proliferation and
migration of VSMCs within the intima contributes to initial
atherosclerotic plaque formation (4). Notably, VSMCs can form fibrous caps to
stabilize vulnerable plaques at advanced stages (5). Furthermore, a previous study has
indicated the regulation of atherosclerosis at tissue and molecular
levels is associated with VSMCs (6).
Several reports have demonstrated that platelet-derived growth
factor-BB (PDGF-BB) can stimulate VSMC proliferation and migration
during vascular injury (7,8). Therefore, exploring the underlying
molecular mechanism involved in the modulation of PDGF-BB-dependent
VSMC proliferation and migration, including PDGF-BB-dependent human
coronary artery smooth muscle cell (HCASMC) proliferation and
migration, may be of great scientific interest.
MicroRNAs (miRs) are a type of endogenous non-coding
RNA that are 22 nucleotides in length (9). By binding directly to the
3′-untranslated region (UTR) of target mRNAs, miRs contribute to
mRNA degradation or transcriptional inhibition, thus regulating
gene expression (10). Accumulating
evidence has indicated that miRs serve an important role in VSMC
proliferation, migration and remodeling. For example, miR-26a is
significantly increased following PDGF-BB treatment in VSMCs, and
is involved in VSMC phenotypic transition by targeting Smad1
(11). Notably, miR-24 effectively
regulated vascular remodeling and reduced the level of inflammatory
factors in a diabetic rat model (12). A previous study indicated that
overexpression of miR-612 inhibited VSMC proliferation and
migration through inducing cell cycle arrest at G1 stage, and RAC-β
serine/threonine-protein kinase (AKT2) was identified as a direct
target of miR-612 (13).
Furthermore, miR-448 has been identified to be highly expressed in
VSMCs from coronary atherosclerotic plaques compared with normal
VSMCs (14). Notably, miR-448
promoted the proliferation and migration of VSMCs by targeting
myocyte enhancer factor 2C (14).
There are fewer reports concerning the role of miR-365b-3p. A
recent study indicated that overexpression of miR-365b-3p
upregulated p21 and p27 and induced cell cycle arrest in
G1 phase and cell apoptosis (15). However, the role of miR-365b-3p in
the HCASMCs is poorly understood.
A disintegrin and metalloproteinase with
thrombospondin motifs 1 (ADAMTS1) is the member of the ADAMTS
family, a family of extracellular proteases (16). ADAMTS1, which is regulated by
peroxisome proliferator-activated receptor, promotes the
proliferation and migration of VSMCs, inducing atherosclerosis and
vascular thrombosis (17). A recent
study suggested that miR-362-3p directly binds to ADAMTS1 and
inhibits the proliferation and migration of VSMCs (18). However, there is no research to
clarify the association between ADAMTS1 and miR-365b-3p. In present
study, the effect of PDGF-BB treatment was explored on miR-365b-3p
and ADAMTS1 expression in HCASMCs. Functional assays were performed
to assess cell the proliferation and migration of HCASMCs.
TargetScan Human 3.1 (www.targetscan.org) and the dual-luciferase reporter
assay were used to identify whether ADAMTS1 is a potential target
of miR-365b-3p.
Materials and methods
Cell culture and treatment
HCASMCs (cat. no. C0175C; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) were cultured in Dulbecco's modified
Eagle's medium (DMEM; Thermo Fisher Scientific, Inc.) supplemented
with 10% fetal bovine serum (Thermo Fisher Scientific, Inc.) at
37°C in a humidified atmosphere containing 5% CO2. For
the PDGF-BB group, HCASMCs were treated with 30 ng/ml PDGF-BB
(Thermo Fisher Scientific, Inc.) for 6 h at 37°C.
Transfection and groups
miR-365b-3p mimic and negative control (NC) mimic
were obtained from GenePharma (Shanghai, China). Lipofectamine 2000
(Thermo Fisher Scientific, Inc.) was used to perform cell
transfection, in accordance with the manufacturer's protocol. Cells
were collected at 48 h post-transfection and grouped as follows:
Control, miR-NC (100 nmol/l), miR-365b-3p mimic (100 nmol/l),
PDGF-BB (20 ng/ml), PDGF-BB + miR-NC and PDGF-BB + miR-365b-3p
mimic. For the PDGF-BB + miR-NC and PDGF-BB + miR-365b-3p mimic
groups, the PDGF-BB was added at 24 h post-transfection. The
sequences of mimics were as follows: miR-365b-3p mimic sense,
5′-UAAUGCCCCUAAAAAUCCUUAU-3′ and antisense,
5′-AAGGAUUUUUAGGGGCAUUATT-3′; negative control mimic sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). For miR, the TaqMan
MicroRNA Reverse Transcription kit (Takara Biotechnology Co., Ltd.,
Dalian, China) was used to obtain cDNA and a TaqMan Reverse
Transcription kit (Takara Biotechnology Co., Ltd.) was used for
mRNA. RT-qPCR was performed using a Perfect Real Time SYBR Premix
Ex Taq kit (Takara Biotechnology Co., Ltd.) with an ABI 7500
thermocycler (Thermo Fisher Scientific, Inc.). All kits were used
according to the manufacturer's protocol. The reaction conditions
for PCR were as follows: Pre-degeneration at 95°C for 3 min and 40
cycles of 95°C for 30 sec and 60°C for 30 sec. Independent
experiments were repeated three times. The relative expression
levels of mRNAs were analyzed using the 2−ΔΔCq method
(19). U6 and GAPDH were used as
control for the expression of miR-365b-3p and ADAMTS1,
respectively. The primers used were as follows: miR-365b-3p
stem-loop primer,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTGCACTGGATACGACATAAGG-3′, forward,
5′-TAATGCCCCTAAAAAT-3′ and reverse, 5′-CCAGTGCAGGGTCCGAGGT-3′; U6
stem-loop primer,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTGCACTGGATACGACAAAATATGG-3′, forward,
5′-TGCGGGTGCTCGCTTCGGCAGC-3′ and reverse,
5′-CCAGTGCAGGGTCCGAGGT-3′; ADAMTS1 forward,
5′-GGATGGCTGATGTTGGAA-3′ and reverse, 5′-CATTAAGGCTGGCACACT-3′; and
GAPDH forward, 5′-CTGGGCTACACTGAGCACC-3′ and reverse,
5′-AAGTGGTCGTTGAGGGCAATG-3′.
Western blot analysis
Total protein extraction was performed using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China). The protein concentration was
measured using the BCA kit (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Protein samples (20 µg/lane) were separated using 10%
SDS-PAGE and blotted onto polyvinylidene difluoride membranes
(Thermo Fisher Scientific, Inc.). The membranes were blocked in 5%
non-fat milk for 1 h at room temperature, followed by incubation
overnight at 4°C with the indicated antibodies against ADAMTS1
(cat. no. MAB 1810; 1:1,000; EMD Millipore; Billerica, MA, USA) and
GAPDH (cat. no. AB 2302; 1:2,000; EMD Millipore). Subsequently, the
membranes were incubated with secondary rabbit anti-mouse
IgG-horseradish peroxidase antibody (cat. no. sc-358914; 1:5,000;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) at room
temperature for a further 2 h. Chemiluminescent signals were
visualized using the enhanced chemiluminescence detection reagent
(EMD Millipore). The relative protein expression was analyzed using
ImageJ software 1.4 (National Institutes of Health, Bethesda, MD,
USA).
Cell Counting kit-8 (CCK-8) assay
The CCK-8 assay was used to assess cell
proliferation. For the CCK-8 assay, cells were seeded in 96-well
plates (5×103 cells/well) and incubated at 37°C for 24
h. Cells were then treated with PDGF-BB and transfected with
miR-365b-3p mimic or NC. At 0, 24, 48 and 72 h post-PDGF-BB
treatment and/or transfection, cell proliferation indices were
measured using a CCK-8 kit (cat no. C0038; Beyotime Institute of
Biotechnology, Shanghai, China), according to the manufacturer's
protocol. The optical density was measured at 450 nm.
Transwell and wound healing
assays
Transwell and wound healing assays were used to
assess cell migration. For the Transwell assay, cells of each
groups (control, miR-NC, miR-365b-3p mimic, PDGF-BB, PDGF-BB +
miR-NC and PDGF-BB + miR-365b-3p mimic) were seeded in the upper
chambers with 200 µl serum-free DMEM at density of 5×103
cells/ml, and the lower chamber was filled with 500 µl serum-free
DMEM. Following an incubation time of 24 h, the migrated cells on
the lower face of the chamber membrane were fixed at room
temperature for 30 min with 4% formaldehyde and stained with 0.1%
crystal violet at room temperature for 30 min. Cells were counted
under a light microscope (magnification, ×200; Nikon Corporation,
Tokyo, USA).
For the wound healing assay, HCASMCs
(5×103 cells/well) were seeded into 6-well plates. At 24
h post-transfection, a scratched wound was created using a pipette
tip. The migrated cells were determined under an inverted
microscope at 0 and 24 h. Image Pro Plus software (version 6.0;
Media Cybernetics, Inc., Rockville, MD, USA) was used to measure
the widths of the scratch wounds.
Dual-luciferase reporter assay
TargetScan Human 3.1 (www.targetscan.org) was used to predict the potential
target of miR-365b-3p. The mutant (MUT) type of AKT2 3′-UTR was
constructed using a Fast Mutagenesis System kit (TransGen Biotech,
Beijing, China), according to the manufacturer's protocol. The wild
type (WT) or MUT type of AKT2 3′-UTR was then inserted into the
Firefly luciferase reporter vector pGL3-promoter (Promega
Corporation, Madison, WI, USA) to generate the recombinant vector
pGL3-ADAMTS1-3′-UTR-WT (3′-UTR-WT) or pGL3-ADAMTS1-3′-UTR-MUT
(3′-UTR-MUT). HCASMCs were cultured on 24-well plates for 24 h
prior to co-transfection with 50 ng of 3′-UTR-WT or 3′-UTR-MUT
vector and 20 µM miR-365b-3p mimics or NC control. At 48 h
post-transfection, the Dual-Luciferase Reporter Assay System
(Promega Corporation) was used to determine the luciferase
activity, and the Renilla luciferase activity was normalized to the
Firefly luciferase activity.
Statistical analysis
Data are presented as the mean ± standard deviation.
GraphPad Prism (version 6; GraphPad software, Inc., La Jolla, CA,
USA) was used to perform the statistical analyses. The differences
between two groups were analyzed using the Student's t-test.
Comparisons between multiple groups were determined by one-way
analysis of variance followed by Bonferroni's multiple comparison
tests. P<0.05 was considered to indicate a statistically
significant difference.
Results
PDGF-BB treatment promotes HCASMC
proliferation and migration and inhibits miR-365b-3p
expression
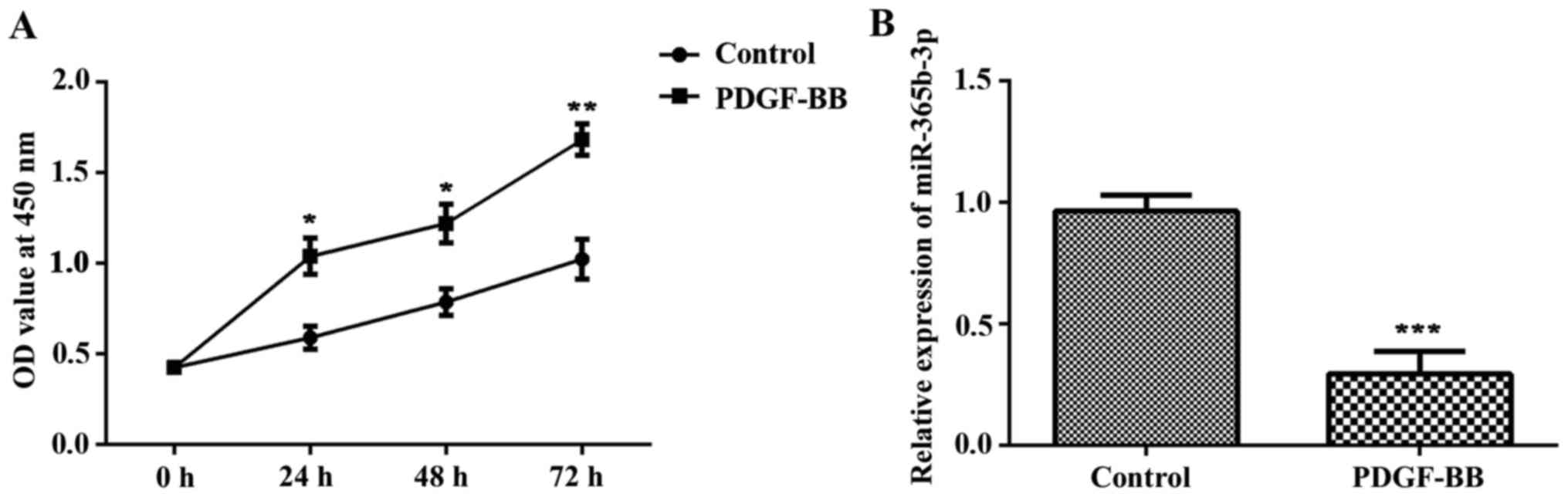
In the present study, HCASMCs were treated with
PDGF-BB for 6 h. Cells without any treatment were used as the
control group. CCK-8 analysis revealed that the proliferation of
HCASMCs was significantly increased in the PDGF-BB group compared
with the control group at 24, 48 and 72 h post-PDGF-BB treatment
(Fig. 1A). RT-qPCR assay was used to
detect the expression of miR-365b-3p. The results indicated that
the expression of miR-365b-3p was significantly decreased in the
PDGF-BB group compared with that of the control (Fig. 1B). These data demonstrated that
PDGF-BB treatment promotes cell proliferation and migration of
HCASMCs and inhibits the expression of miR-365b-3p.
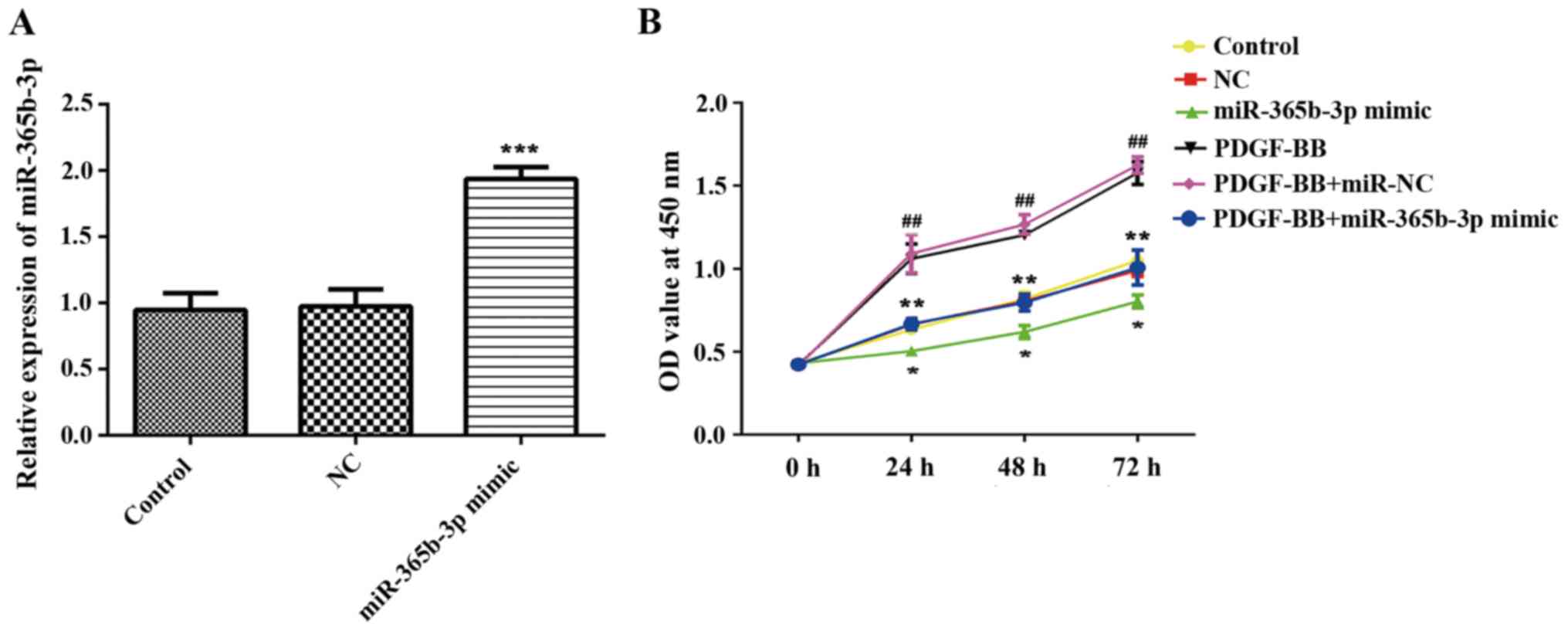
Overexpression of miR-365b-3p inhibits
HCASMC proliferation and migration
To explore the function of miR-365b-3p in HCASMCs,
miR-365b-3p mimic was transfected in HCASMCs with or without
PDGF-BB treatment, and NC mimic was used as negative control
(Fig. 2A). A CCK-8 assay revealed
that overexpression of miR-365b-3p in HCASMCs with or without
PDGF-BB treatment significantly inhibited cell proliferation when
compared with NC or PDGF-BB+NC groups, respectively, at 24, 48 and
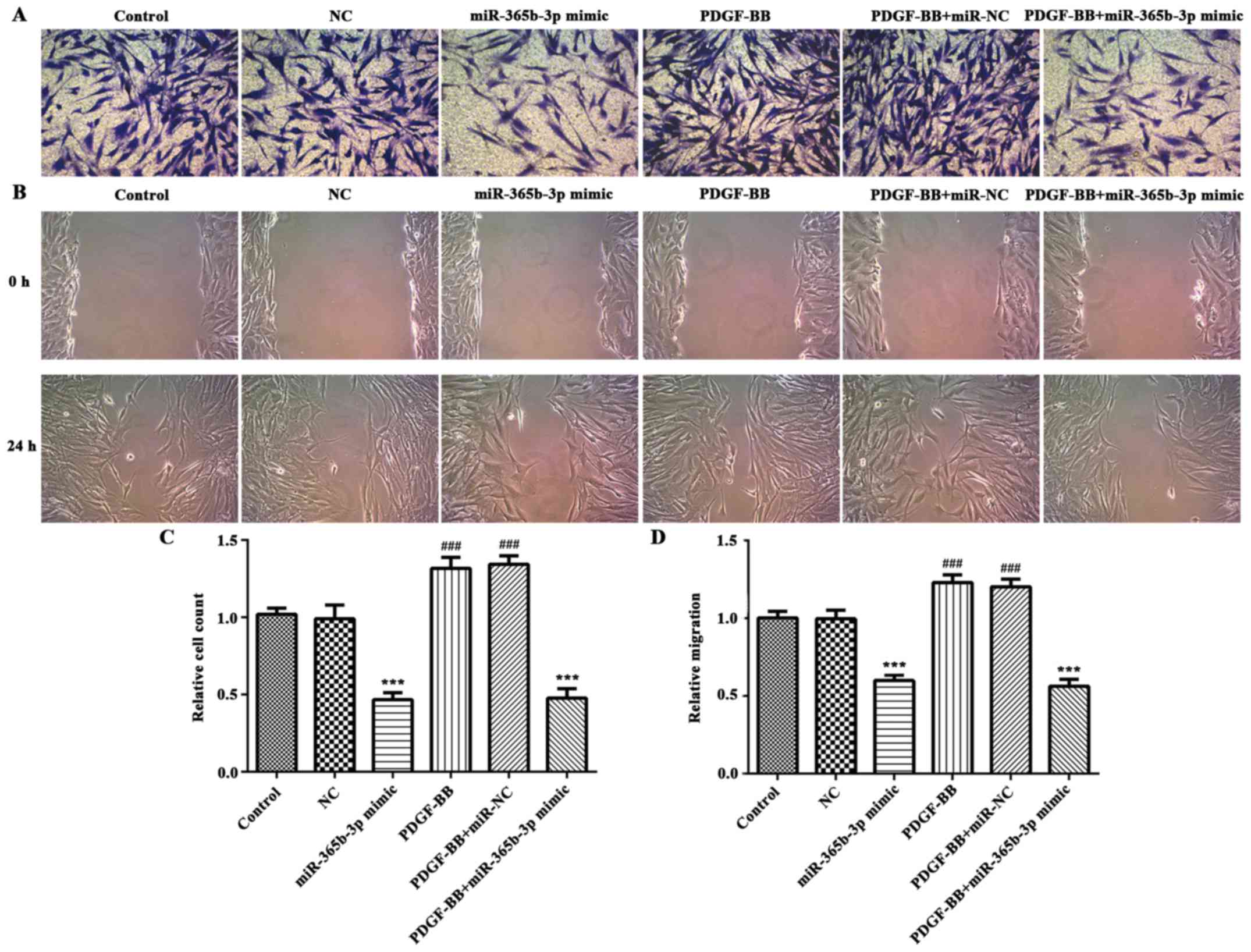
72 h (Fig. 2B). Transwell assays
were performed to assess cell migration. The results indicated that
overexpression of miR-365b-3p significantly attenuated the
upregulation of PDGF-BB-induced HCASMC migration (Fig. 3). The wound healing assay exhibited
the similar results (Fig. 3). Taken
together, these results suggested that overexpression of
miR-365b-3p inhibits HCASMC proliferation and migration.
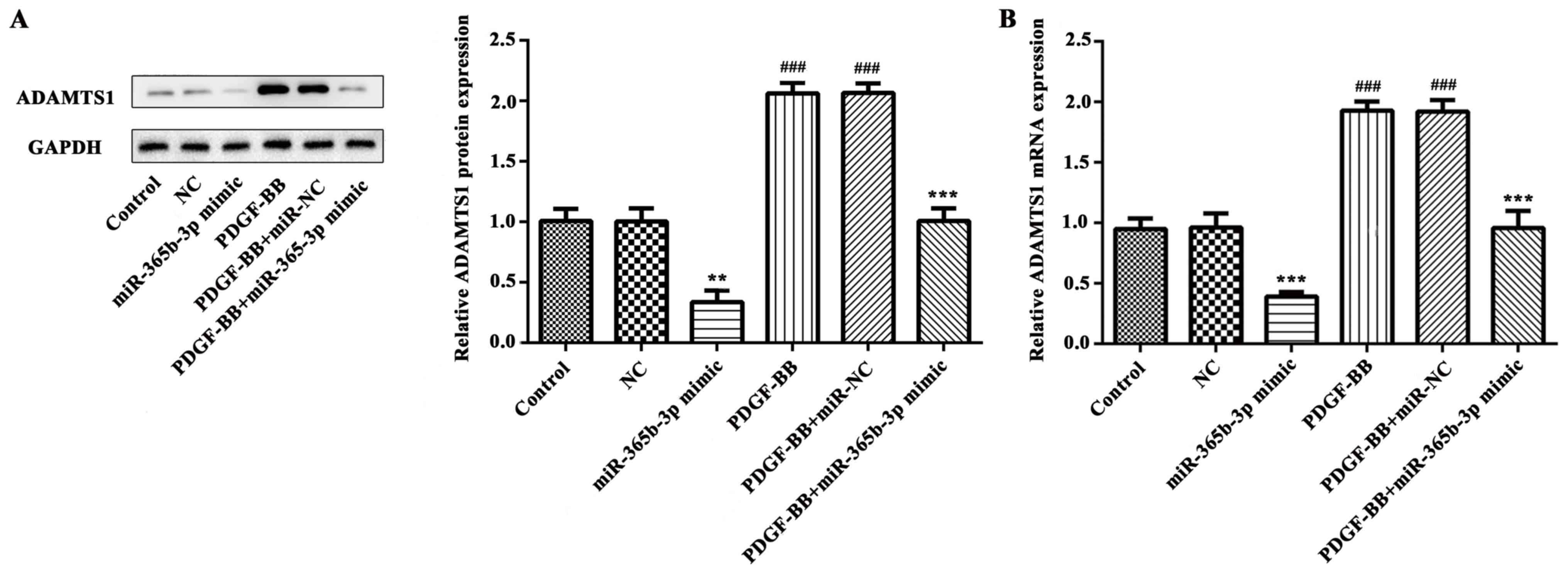
miR-365b-3p is responsible for the
PDGF-BB-mediated increase of ADAMTS1 expression
In order to further study the underlying mechanism,
western blot analysis and RT-qPCR were performed to detect the
expression levels of ADAMTS1. The data indicated that PDGF-BB
treatment significantly increased ADAMTS1 expression at protein and
mRNA levels (Fig. 4). However, the
expression of ADAMTS1 protein was significantly decreased in the
miR-365b-3p group compared with the NC group (Fig. 4A) and relative mRNA expression levels
exhibited similar results (Fig. 4B).
In the PDGF-BB+miR-365-3p group, ADAMTS1 mRNA and protein
expression levels were significantly decreased compared with the
PDGF-BB+NC group (Fig. 4A and B).
These results revealed that miR-365b-3p may downregulate the
PDGF-BB-induced expression of ADAMTS1 by contributing to the
degradation of mRNA.
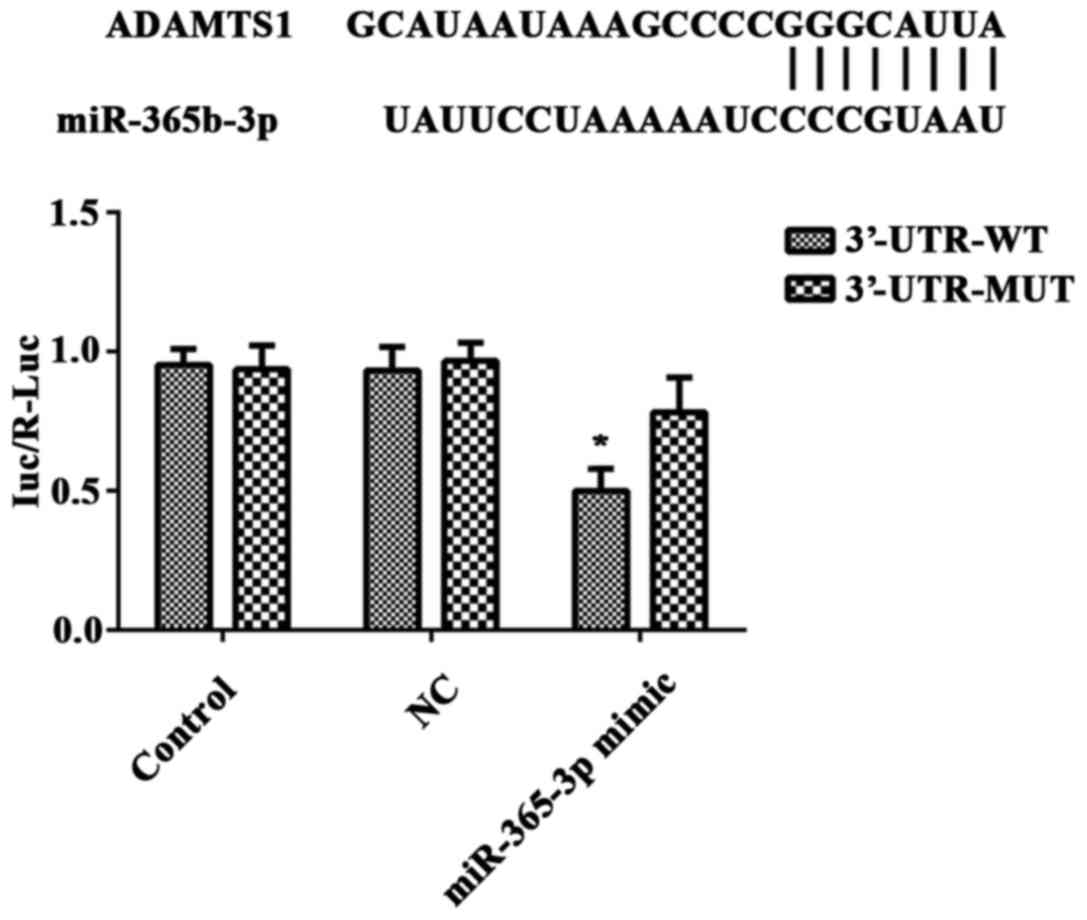
ADAMTS1 is a direct target of
miR-365b-3p
Protein and mRNA experiments indicated that ADAMTS1
may be a target gene of miR-365b-3p. To confirm this prediction,
the recombinant vector pGL3-ADAMTS1-3′-UTR-WT (3′-UTR-WT) or
pGL3-ADAMTS1-3′-UTR MUT type (3′-UTR-MUT) and miR-365b-3p mimic or
NC mimic were co-transfected in HCASMCs, and a luciferase reporter
gene assay was performed. The luciferase activity was significantly
downregulated in miR-365b-3p mimic-transfected HCASMCs transfected
with 3′-UTR-WT luciferase reporter plasmid when compared with those
transfected with 3′-UTR-MUT luciferase reporter plasmid (Fig. 5). However, no significant difference
was identified between the groups transfected with the NC mimic
(Fig. 5). These findings indicated
that miR-365b-3p directly binds to the 3′-UTR of ADAMTS1 mRNA in
HCASMCs.
Discussion
In the present study, it was demonstrated that
treatment of PDGF-BB promoted HCASMC proliferation and migration.
Jönsson-Rylander et al (17)
have suggested that ADAMTS1 promotes the proliferation and
migration of VSMCs. Considering this, western blot analysis and
RT-qPCR assays were performed to assess ADAMTS1 expression in the
present study. The results suggested that PDGF-BB treatment
decreased ADAMTS1 expression at protein and mRNA levels. Rescue
experiments indicated that PDGF-BB treatment significantly
downregulated the expression of miR-365b-3p in HCASMCs. TargetScan
Human 3.1 was used to identify that ADAMTS1 is a potential target
of miR-365b-3p. Based on the findings, miR-365b-3p mimic was
transfected in HCASMCs to further explore the potential role of
this miR in atherosclerosis. Western blot analysis and RT-qPCR
assay results revealed that miR-365b-3p inhibited ADAMTS1
expression at protein and mRNA levels. The dual-luciferase reporter
assay results confirmed that miR-365b-3p could directly bind to
ADAMTS1. Furthermore, functional assay results indicated that
miR-365b-3p inhibited HCASMC proliferation and migration. Taken
together, the present findings suggested that overexpression of
miR-365b-3p significantly attenuated PDGF-BB-induced proliferation
and migration of HCASMCs through directly targeting ADAMTS1.
The extracellular matrix (ECM) is associated with
various physiological and pathological conditions, including
embryogenesis, cell-to-cell interactions, hemostasis, cell
migration and apoptosis (20,21).
ADAMTS1 remodels the ECM through catalyzing proteoglycan
degradation (22). Since destruction
of ECM is thought to be responsible for the initiation of cancer
(23,24), multiple studies have focused on the
function of ADAMTS1 in cancer. Dysregulation of ADAMTS1 have been
reported in various types of cancer, including prostate, breast,
lung, colon and liver cancer (25–30).
Studies have revealed that ADAMTS1 has a role in the development of
atherosclerosis (17,31). Atherosclerosis is a complicated
process stimulated by environmental and genetic factors (18) that is the leading cause of coronary
heart disease, cerebral infarction and peripheral vascular disease
(3). Subendothelial retention of
atherogenic lipoproteins serves crucial roles in the initiation of
atherosclerosis (32). Versican, a
proteoglycan substrate of ADAMTS1, has a been demonstrated to have
a critical role in this process. Notably, the degradation of
versican by ADAMTS1 has been revealed to contribute to
atherosclerosis (33). A recent
study conducted by Vorkapic et al (34) has also demonstrated the association
between the cleavage of ADAMTS1 substrates and atherosclerosis
development. Furthermore, in an acute aortic dissection (AAD) mouse
model, it was suggested that ADAMTS1 promotes the progression of
AAD by degrading versican in macrophages and neutrophils (35). Jönsson-Rylander et al
(17) suggested that ADAMTS1 may
promote VSMC migration by cleaving ECM proteins in the mouse
carotid artery flow cessation model. Taken together, a growing
number of studies suggest that ADAMTS1 has a crucial role in the
initiation and development of atherosclerosis. Although numerous
animal model and tissue experimentation studies have investigated
ADAMTS1, the underlying mechanisms at the molecular level are still
poorly understood.
miRs are non-coding RNAs that are small in size but
have significant effects. It is well acknowledged that miRs are
involved in gene expression at the post-transcriptional level and
serve a role in in plethora of biological processes (10,36). In
recent years, miRs have received much attention for their function
in cardiovascular disease, including atherosclerosis (37,38).
With further study, a number of miRs have been identified to be
associated with the regulation of ADAMTS1. For example, a previous
study suggested that the expression of ADAMTS1 was inhibited by
miR-142 in human endothelial progenitor cells, and the following
upregulation of the endothelial nitric oxide synthase expression
serves an important role in the angiogenesis (39). Previous research on breast cancer
indicated that ADAMTS1 is also a target of miR-365, and
downregulation of this miR inhibits cell proliferation and cell
invasion (40). Other miRs, such as
miR-21 (41), miR-378 (42) and miR-181d (43), have been demonstrated to have the
function of regulating ADAMTS1 expression and serve their
respective roles in different biological processes. However, to the
best of our knowledge no research has clarified the association
between ADAMTS1 and miR-365b-3p. The present findings suggested
that miR-365b-3p inhibited ADAMTS1 expression by directly targeting
ADAMTS1. The findings of the present study suggest the possible
molecular pathway of ADAMTS1 in its involvement in
atherosclerosis.
Notably, there are a few limitations of the present
study. First, the experiments in the current study were
investigated the mechanism at a cell level and thus, as there was
not clinical data, lack further validation. Second, miRs are
involved in complex regulatory networks in cells. A single mRNA
molecule may be regulated by multiple miRs, and each miR may be
involved in a number of gene regulatory mechanisms (44,45). It
is hard to say whether there are other genes regulated by the
overexpression of miR-365b-3p that could impact the proliferation
and migration of HCASMCs. Further studies should be performed to
clarify these issues.
In conclusion the data obtained in the present study
demonstrated that PDGF-BB treatment significantly downregulated the
expression of miR-365b-3p in HCASMCs. Furthermore, overexpression
of miR-365b-3p significantly attenuated PDGF-BB-induced HCASMC
proliferation and migration through directly targeting ADAMTS1.
These results provide novel evidence regarding the role of miRs in
coronary atherosclerosis and advance our understanding of the
mechanistic regulation of ADAMTS1.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YQ analyzed the data, wrote and revised the
manuscript. YQ and NZ collected the data and designed the
study.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Weber C and Noels H: Atherosclerosis:
Current pathogenesis and therapeutic options. Nat Med.
17:1410–1422. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lusis AJ: Atherosclerosis. Nature.
407:233–241. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Roth GA, Johnson C, Abajobir A, Abd-Allah
F, Abera SF, Abyu G, Ahmed M, Aksut B, Alam T, Alam K, et al:
Global, regional, and national burden of cardiovascular diseases
for 10 causes, 1990 to 2015. J Am Coll Cardiol. 70:1–25. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Johnson JL: Emerging regulators of
vascular smoothmuscle cell function in the development and
progression of atherosclerosis. Cardiovasc Res. 103:452–460. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ross R: Atherosclerosis-an inflammatory
disease. N Engl J Med. 340:115–126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hutcheson JD, Goettsch C, Bertazzo S,
Maldonado N, Ruiz JL, Goh W, Yabusaki K, Faits T, Bouten C, Franck
G, et al: Genesis and growth of extracellular-vesicle-derived
microcalcification in atherosclerotic plaques. Nat Mater.
15:335–343. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Millette E, Rauch BH, Kenagy RD, Daum G
and Clowes AW: Platelet-derived growth factor-BB transactivates the
fibroblast growth factor receptor to induce proliferation in human
smooth muscle cells. Trends Cardiovasc Med. 16:25–28. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Raines EW: PDGF and cardiovascular
disease. Cytokine Growth Factor Rev. 15:237–254. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moss EG: MicroRNAs: Hidden in the genome.
Curr Biol. 12:R138–R140. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang X, Dong M, Wen H, Liu X, Zhang M, Ma
L, Zhang C, Luan X, Lu H and Zhang Y: MiR-26a contributes to the
PDGF-BB-induced phenotypic switch of vascular smooth muscle cells
by suppressing Smad1. Oncotarget. 8:75844–75853. 2017.PubMed/NCBI
|
|
12
|
Yang J, Zeng P, Yang J, Liu X, Ding J,
Wang H and Chen L: MicroRNA-24 regulates vascular remodeling via
inhibiting PDGF-BB pathway in diabetic rat model. Gene. 659:67–76.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen C, Yan Y and Liu X: microRNA-612 is
downregulated by platelet-derived growth factor-BB treatment and
has inhibitory effects on vascular smooth muscle cell proliferation
and migration via directly targeting AKT2. Exp Ther Med.
15:159–165. 2018.PubMed/NCBI
|
|
14
|
Zhang RH, Sui L, Hong XJ, Yang M and Li
WM: MiR-448 promotes vascular smooth muscle cell proliferation and
migration in through directly targeting MEF2C. Environ Sci Pollut
Res Int. 24:22294–22300. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang J, Wang X, Wu G, Hou D and Hu Q:
miR-365b-3p, down-regulated in retinoblastoma, regulates cell cycle
progression and apoptosis of human retinoblastoma cells by
targeting PAX6. FEBS Lett. 587:1779–1786. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang BL: ADAMTS: A novel family of
extracellular matrix proteases. Int J Biochem Cell Biol. 33:33–44.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jönsson-Rylander AC, Nilsson T,
Fritsche-Danielson R, Hammarström A, Behrendt M, Andersson JO,
Lindgren K, Andersson AK, Wallbrandt P, Rosengren B, et al: Role of
ADAMTS-1 in atherosclerosis: Remodeling of carotid artery,
immunohistochemistry, and proteolysis of versican. Arterioscler
Thromb Vasc Biol. 25:180–185. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li M, Liu Q, Lei J, Wang X, Chen X and
Ding Y: miR-362-3p inhibits the proliferation and migration of
vascular smooth muscle cells in atherosclerosis by targeting
ADAMTS1. Biochem Biophys Res Commun. 493:270–276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu P, Weaver VM and Werb Z: The
extracellular matrix: A dynamic niche in cancer progression. J Cell
Biol. 196:395–406. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vong S and Kalluri R: The role of stromal
myofibroblast and extracellular matrix in tumor angiogenesis. Genes
Cancer. 2:1139–1145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuno K, Okada Y, Kawashima H, Nakamura H,
Miyasaka M, Ohno H and Matsushima K: ADAMTS-1 cleaves a cartilage
proteoglycan, aggrecan. FEBS Lett. 478:241–245. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rucci N, Sanità P and Angelucci A: Roles
of metalloproteases in metastatic niche. Curr Mol Med. 11:609–622.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hart IR and Saini A: Biology of tumour
metastasis. Lancet. 339:1453–1457. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tan Ide A, Ricciardelli C and Russell DL:
The metalloproteinase ADAMTS1: A comprehensive review of its role
in tumorigenic and metastatic pathways. Int J Cancer.
133:2263–2276. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gustavsson H, Tesan T, Jennbacken K, Kuno
K, Damber JE and Welén K: ADAMTS1 alters blood vessel morphology
and TSP1 levels in LNCaP and LNCaP-19 prostate tumors. Bmc Cancer.
10:2882010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fletcher SJ, Sacca PA, Pistone-Creydt M,
Coló FA, Serra MF, Santino FE, Sasso CV, Lopez-Fontana CM, Carón
RW, Calvo JC and Pistone-Creydt V: Human breast adipose tissue:
Characterization of factors that change during tumor progression in
human breast cancer. J Exp Clin Cancer Research. 36:262017.
View Article : Google Scholar
|
|
28
|
Kohno T, Otsuka A, Girard L, Sato M,
Iwakawa R, Ogiwara H, Sanchez-Cespedes M, Minna JD and Yokota J: A
Catalog of genes homozygously deleted in human lung cancer and the
candidacy of PTPRD as a tumor suppressor gene. Genes Chromosomes
Cancer. 49:342–352. 2010.PubMed/NCBI
|
|
29
|
Mullany LE, Herrick JS, Wolff RK and
Slattery ML: Single nucleotide polymorphisms within MicroRNAs,
MicroRNA targets, and MicroRNA biogenesis genes and their impact on
colorectal cancer survival. Genes Chromosomes Cancer. 56:285–295.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee YJ, Koch M, Karl D, Torres-Collado AX,
Fernando NT, Rothrock C, Kuruppu D, Ryeom S, Iruela-Arispe ML and
Yoon SS: Variable inhibition of thrombospondin 1 against liver and
lung metastases through differential activation of
metalloproteinase ADAMTS1. Cancer Res. 70:948–956. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
De Schutter A, Lavie CJ and Milani RV: The
impact of obesity on risk factors and prevalence and prognosis of
coronary heart disease-the obesity paradox. Prog Cardiovasc Dis.
56:401–408. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Williams KJ: Arterial wall chondroitin
sulfate proteoglycans: Diverse molecules with distinct roles in
lipoprotein retention and atherogenesis. Curr Opin Lipidol.
12:477–487. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wight TN: Versican: A versatile
extracellular matrix proteoglycan in cell biology. Curr Opin Cell
Biol. 14:617–623. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vorkapic E, Folkesson M, Magnell K,
Bohlooly YM, Länne T and Wågsäter D: ADAMTS-1 in abdominal aortic
aneurysm. PLoS One. 12:e01787292017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao Y, Wu W, Yu C, Zhong F, Li G, Kong W
and Zheng J: A disintegrin and metalloproteinase with
thrombospondin motif 1 (ADAMTS1) expression increases in acute
aortic dissection. Sci China Life Sci. 59:59–67. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wahid F, Shehzad A, Khan T and Kim YY:
MicroRNAs: Synthesis, mechanism, function, and recent clinical
trials. Biochim Biophys Acta. 1803:1231–1243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bayoumi AS, Aonuma T, Teoh JP, Tang YL and
Kim IM: Circular noncoding RNAs as potential therapies and
circulating biomarkers for cardiovascular diseases. Acta Pharmacol
Sin. 39:1100–1109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pordzik J, Pisarz K, De Rosa S, Jones AD,
Eyileten C, Indolfi C, Malek L and Postula M: The potential role of
platelet-related microRNAs in the development of cardiovascular
events in high-risk populations, including diabetic patients: A
review. Front Endocrinol (Lausanne). 9:742018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang HW, Li H, Yan H and Liu BL:
MicroRNA-142 promotes the expression of eNOS in human peripheral
blood-derived endothelial progenitor cells in vitro. Eur Rev Med
Pharmacol Sci. 20:4167–4175. 2016.PubMed/NCBI
|
|
40
|
Li M, Liu L, Zang W, Wang Y, Du Y, Chen X,
Li P, Li J and Zhao G: miR365 overexpression promotes cell
proliferation and invasion by targeting ADAMTS-1 in breast cancer.
Int J Oncol. 47:296–302. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu L, Yin H, Huang M, He J, Yi G, Wang Z
and Qian H: miR-21 promotes pulmonary fibrosis in rats via
down-regulating the expression of ADAMTS-1. Xi Bao Yu Fen Zi Mian
Yi Xue Za Zhi. 32:1636–1640. 2016.(In Chinese). PubMed/NCBI
|
|
42
|
Pan B, Toms D, Shen W and Li J:
MicroRNA-378 regulates oocyte maturation via the suppression of
aromatase in porcine cumulus cells. Am J Physiol Endocrinol Metab.
308:E525–E534. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen SZ, Ning LF, Xu X, Jiang WY, Xing C,
Jia WP, Chen XL, Tang QQ and Huang HY: The miR-181d-regulated
metalloproteinase Adamts1 enzymatically impairs adipogenesis via
ECM remodeling. Cell Death Differ. 23:1778–1791. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Anglicheau D, Muthukumar T and
Suthanthiran M: MicroRNAs: Small RNAs with big effects.
Transplantation. 90:105–112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Witkos TM, Koscianska E and Krzyzosiak WJ:
Practical aspects of microRNA target prediction. Curr Mol Med.
11:93–109. 2011. View Article : Google Scholar : PubMed/NCBI
|