Introduction
Age-associated vascular diseases are accompanied by
structural and functional changes in blood vessels. A body of
evidence suggested that the behavior of vascular smooth muscle
cells (VSMCs) is modified with ageing (1), and senescent VSMCs have been observed
in atherosclerotic plaques of patients with atherosclerosis
(2). Thus, senescent VSMCs
contribute to ageing and age-associated cardiovascular diseases.
Although permanent inhibition of cell proliferation is thought to
be a traditional hallmark of cellular senescence, this procedure is
accompanied by changes in the expression of replicative
senescence-associated genes, such as p53, p21 and
senescence-associated β-galactosidase (SA-β-gal) (3).
Mammalian target of rapamycin (mTOR) is a
serine/threonine protein kinase and its functions rely on forming
two distinct complexes, mTOR complex 1 (mTORC1) and mTORC2
(4). Dysfunctional mTOR signaling
has been considered a central integral mechanism linked to ageing
(5). AMP-activated protein kinase
(AMPK) is a highly conserved heterotrimeric kinase complex.
Activation of AMPK leads to the inhibition of mTORC1 via a tuberous
sclerosis complex 2 (TSC2)-dependent or -independent mechanism
(6). In animal models with impaired
mTORC1 signaling, AMPK is highly activated (7), suggesting a negative crosstalk between
these two pathways. AMPK/TSC2/mTOR is a classic upstream pathway of
mTORC1 signaling, which affects the phosphorylation of downstream
factors, such as ribosome protein subunit 6 kinase 1 (S6K1), to
regulate cell metabolism, proliferation, differentiation and
autophagy. However, the role of mTOR and particularly that of its
downstream effector S6K1 in age-associated vascular diseases has
remained largely elusive. A previous study has demonstrated that
inhibition of the mTORC1/S6K1 pathway by rapamycin or activation of
AMPK by 5-aminoimidazole-4-carboxamide riboside (AICAR) negatively
regulates mTORC1 signaling and attenuated pressure overload-induced
cardiac hypertrophy in vivo (8). Conversely, a deficiency in AMPK
expression was reported to enhance mTOR/S6K1 signaling and
exacerbate myocardial hypertrophy in response to pressure overload
(9). However, the role of the
AMPK/mTOR signaling in VSMC senescence has remained elusive.
The purpose of the present study was to explore the
roles of AMPK/TSC2/mTOR signaling in the replicative senescence of
VSMCs and to provide a theoretical basis for the clinical
prevention and treatment of vascular aging.
Materials and methods
Cell culture
Human VMSCs were purchased from the American Type
Culture Collection (ATCC; Manassas, VA, USA; ATCC® no.
CRL-1999). Cells were cultured in Vascular Cell Basal Medium
(ATCC). When cells reached 90% confluence, 20% of the cells were
re-plated and cultured until they reached 90% confluence again for
the next passage and were then passaged every 3–4 days. The VSMC
phenotype was confirmed by positive immunohistochemical staining
for α-smooth muscle actin using anti-α-SMA antibody (catalogue no.
55135-1-AP; 1:100 dilution; Proteintech, Chicago, IL, USA)
following the manufacture's protocol.
Cell treatments
Human VSMCs were divided into five treatment groups:
i) Young cell group [5th passage (P5)], ii) replicative senescence
group (P15), iii) replicative senescence + rapamycin group [10 nM
rapamycin (Calbiochem Corp; EMD Millipore, Billerica, MA, USA) was
added to the medium from P5-15], iv) replicative senescence +
Compound C group [20 µm/l AMPK inhibitor Compound C (Calbiochem
Corp; EMD Millipore) was added from P5-15] and v) replicative
senescence + AICAR group [0.1 mM AMPK activator AICAR (Calbiochem
Corp; EMD Millipore) was added from P5-15].
Western blot analysis
Cells were lysed and the protein concentration was
measured using a BCA Protein Assay kit (Beyotime Institute of
Biotechnology, Haimen, China). Total protein (20 µg per lane) was
separated by 12% SDS-PAGE and transferred onto nitrocellulose
membranes (Pierce; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). After blocking with 5% non-fat milk for 45 min, membranes
were incubated with primary antibodies overnight at 4°C, followed
by incubation with secondary antibody Anti-mouse IgG, HRP-linked
Antibody (catalogue no. 7076) or anti-rabbit IgG, HRP-linked
antibody (catalogue no. 7074) both at a dilution of 1:2,000,
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA)
for 2 h at room temperature. Reactive proteins were visualized
using chemiluminescent reagents (Pierce; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). As a loading control, the blots were
probed with GAPDH antibody (catalogue no. SC-365062; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). Primary antibodies for
AMPKα, (catalogue no. 5831), p-mTOR Ser2481 (catalogue
no. 2974), phospho (p)-AMPKα Thr172 (catalogue no.
2535), mTOR (catalogue no. 2972), p-mTOR Ser2448
(catalogue no. 2971), TSC2 (catalogue no. 3612) and p-TSC2
(catalogue no. 3615) were purchased from Cell Signaling Technology,
Inc. Primary antibodies for p-mTOR Thr2446 (catalogue
no. ab63552), ALP (ab133602) S6K1 (catalogue no. ab32529),
p-S6K1Thr389 (catalogue no. ab2571) and P21 (catalogue
no. ab102635) were purchased from Abcam (Cambridge, MA, USA).
Primary antibodies for osteocalcin (catalogue no. SC-74495) and p53
(catalogue no. SC-6243), were purchased from Santa Cruz
Biotechnology, Inc.
SA-β-gal activity
SA-β-gal activity was measured according to a
previously established protocol (10). SA-β-gal staining was performed using
a senescence-galactosidase staining kit (Cell Signaling Technology,
Inc., Danvers, MA, USA) by following the manufacturer's manual.
After the blue staining became visible in either experimental or
control cells, 500 cells in each treatment group were counted under
a light microscope.
Statistical analysis
Values are expressed as the mean ± standard
deviation. Statistical analysis was performed using SPSS v17.0
(SPSS, Inc., Chicago, IL, USA). Differences between two groups were
analyzed using the Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Inhibition of the AMPK/TSC2/mTOR
pathway inhibits replicative senescence of VSMCs
Human VSMCs were treated as described in the Methods
section. The percentage of SA-β-gal-positive cells was increased in
the replicative senescence, replicative senescence + rapamycin,
replicative senescence + Compound C and replicative senescence +
AICAR groups compared with that of young cells (P<0.001;
Fig. 1A and B). The senescent cells
presented as irregular or triangular shapes with obvious
pseudopodia and larger overall size compared with that of normal
cells. Intracellular vacuoles were seen in a proportion of the
senescent cells. The percentage of SA-β-gal positive cells was
decreased in the replicative senescence + rapamycin (P<0.001),
replicative senescence + CC (P<0.001) and replicative senescence
+ AICAR (P<0.001) groups compared with that in the replicative
senescence group. The percentage of SA-β-gal-positive cells in the
replicative senescence + Compound C group was significantly higher
than that in the replicative senescence + rapamycin and replicative
senescence + AICAR groups (P<0.05, P<0.01, respectively). The
percentage of SA-β-gal positive cells in young cell group was
significantly lower than that in all other groups (P<0.05;
Fig. 1A and B).
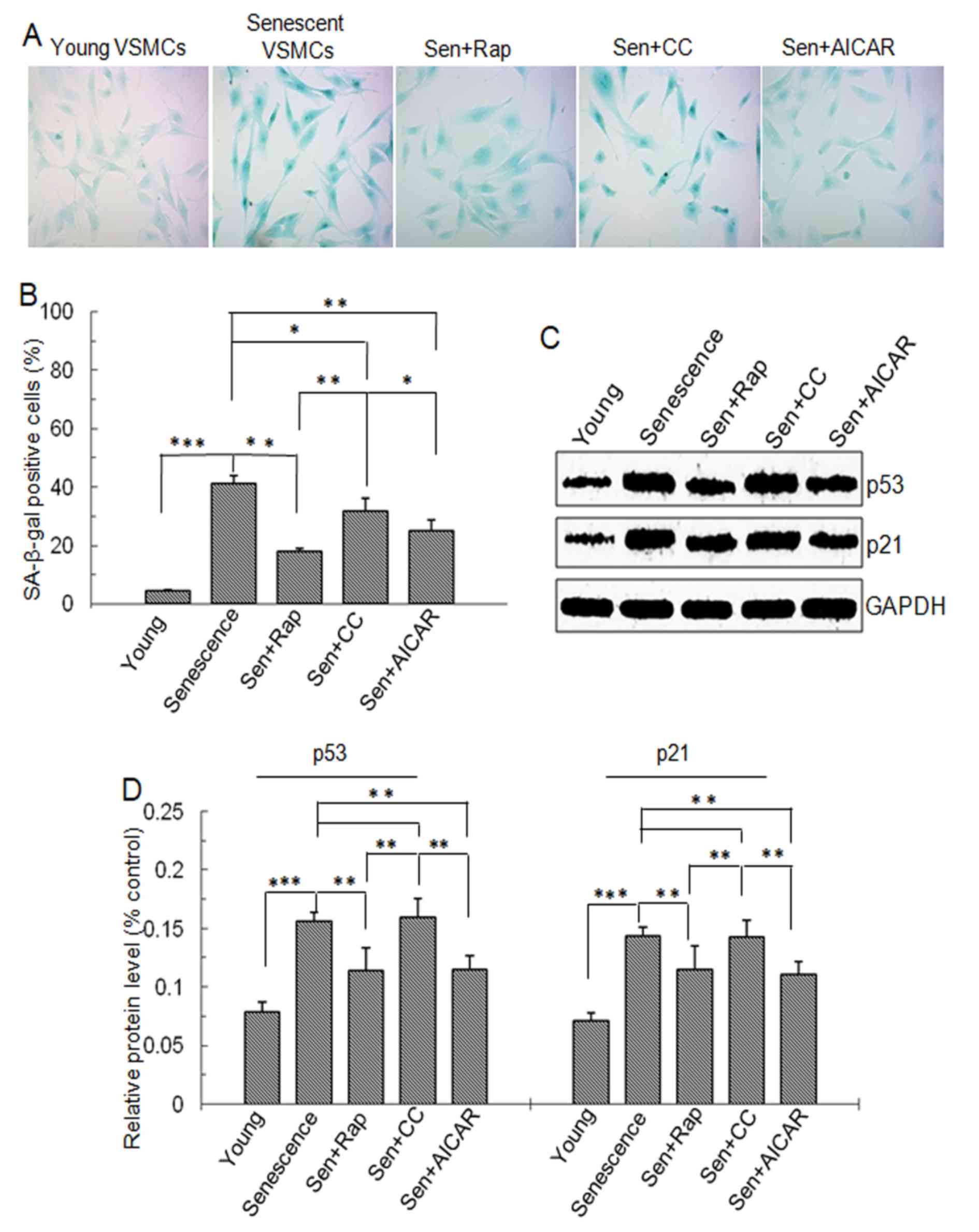 | Figure 1.SA-β-gal staining and p53 and p21
protein level. Vascular smooth muscle cells were divided into five
groups: Young cells, senescent cells, Sen + rap, Sen + CC and Sen +
AICAR group. (A) SA-β-gal staining (blue). Magnification ×200. (B)
Quantification SA-β-gal-stained cells. (C) Representative western
blots displaying p53 and p21 protein expression. (D)
Semi-quantitative analysis of p53 and p21 protein levels in C.
Values are expressed as the mean ± standard deviation (n=4).
*P<0.05, **P<0.01, ***P<0.001 between two indicated
groups. Sen, senescent cells; Rap, rapamycin; CC, Compound C;
SA-β-gal, senescence-associated β-galactosidase, AICAR,
5-aminoimidazole-4-carboxamide riboside (agonist of adenosine
monophosphate-activated protein kinase). |
p53 and p21 are senescence-associated proteins.
Western blot analysis demonstrated that p53 and p21 protein levels
were significantly higher in cells with replicative senescence
compared with those in young cells. However, in the rapamycin- and
AICAR-treated senescent cells, p53 and p21 protein levels were
significantly lowered (P<0.01) and were significantly lower than
those in the Compound C-treated senescent cells (P<0.01;
Fig. 1C and D). These findings
suggested that mTOR signaling was involved in the replicative
senescence of VSMCs and that rapamycin and AICAR inhibits
replicative senescence.
The AMPK/TSC2/mTOR signaling pathway
regulates the replicative senescence of VSMCs
The mTOR protein expression and the levels of p-mTOR
(S2448), p-mTOR (T2446) and p-S6K1 protein
were significantly higher in the replicative senescence,
replicative senescence + rapamycin, replicative senescence +
Compound C and replicative senescence + AICAR groups compared with
those in young cells (Fig. 2). The
levels in the rapamycin- and AICAR- treated replicative senescence
groups were significantly lower than those in the replicative
senescence and Compound C-treated replicative senescence groups
(Fig. 2). No significant differences
in p-mTOR (S2481) and S6K1 levels were observed between
the 5 groups (Fig. 2).
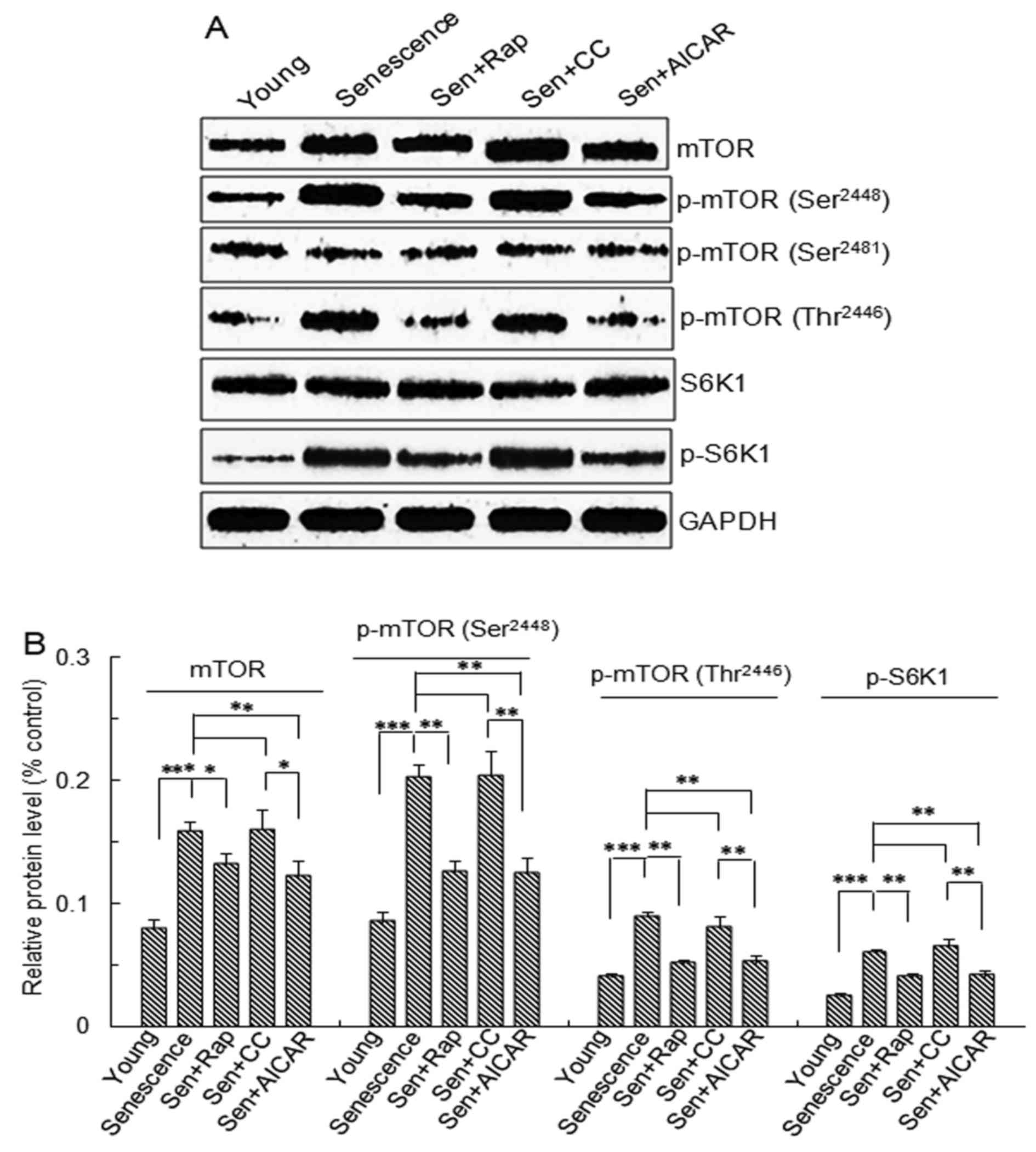 | Figure 2.mTOR and S6K1 protein expression and
phosphorylation. Vascular smooth muscle cells were divided into
five groups: Young cells, senescent cells, Sen + rap, Sen + CC and
Sen + AICAR group. (A) Representative western blots of mTOR, p-mTOR
(Ser2448), p-mTOR (Thr2446), p-mTOR
(Ser2481), S6K1 and p-S6K1 protein levels. (B)
Semi-quantitative analysis of mTOR, p-mTOR (Ser2448),
p-mTOR (Thr2446) and p-S6K1 protein levels in A. Values
are expressed as the mean ± standard deviation (n=4). *P<0.05,
**P<0.01, ***P<0.001 between two indicated groups. Sen,
senescent cells; Rap, rapamycin; CC, Compound C; p-mTOR,
phosphorylated mammalian target of rapamycin; S6K1, ribosomal
protein S6 kinase 1; AICAR, 5-aminoimidazole-4-carboxamide riboside
(agonist of adenosine monophosphate-activated protein kinase). |
To further investigate the upstream signaling of
mTOR, AMPK and TSC2 in the treatment groups were examined by
western blot analysis. The results demonstrated that p-AMPKα
Thr172 and p-TSC2 protein levels were significantly
lower in the replicative senescence, replicative senescence +
rapamycin, replicative senescence + Compound C and replicative
senescence +AICAR groups compared with those in young cells
(Fig. 3). p-AMPKα Thr172
and p-TSC2 protein levels in the Compound C-treated replicative
senescence group were significantly lower than those in the AICAR-
and rapamycin-treated replicative senescence groups (Fig. 3). p-AMPKα Thr172 and
p-TSC2 protein levels in the rapamycin- and AICAR-treated
replicative senescence groups were significantly higher than those
in the replicative senescence group. p-TSC2 protein levels in the
Compound C-treated replicative senescence group were significantly
lower than those in the replicative senescence group (P<0.05;
Fig. 3B).
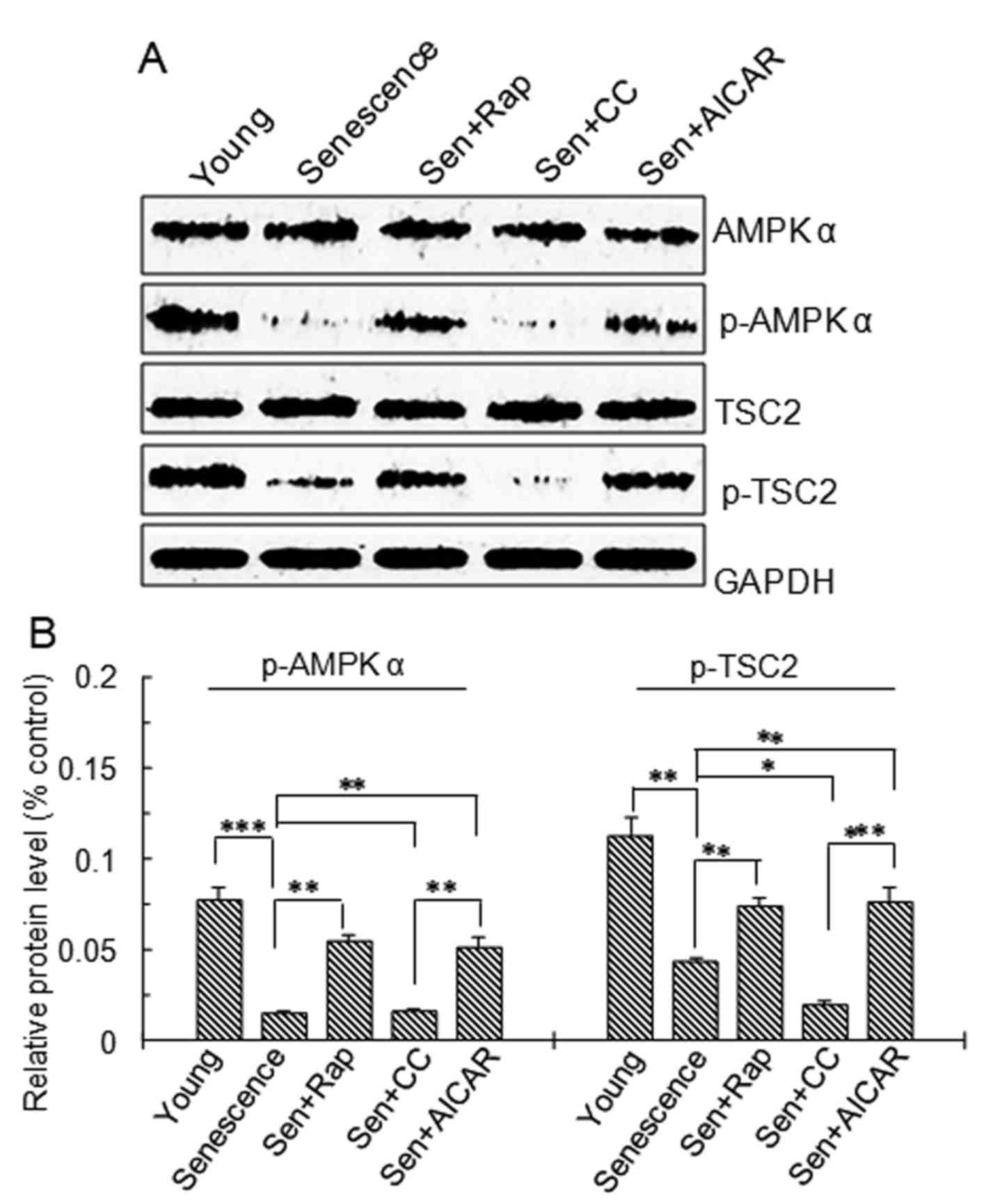 | Figure 3.AMPK and TSC2 protein expression and
phosphorylation. Vascular smooth muscle cells were divided into
five groups: Young cells, senescent cells, Sen + rap, Sen + CC and
Sen + AICAR group. (A) Representative western blots of AMPKα,
p-AMPKα (Thr172), TSC2 and p-TSC2 protein levels. (B)
Semi-quantitative analysis of AMPKα, p-AMPKα (Thr172),
TSC2 and p-TSC2 protein levels in A. Values are expressed as the
mean ± standard deviation (n=4). *P<0.05, **P<0.01,
***P<0.001 between two indicated groups. Sen, senescent cells;
Rap, rapamycin; CC, Compound C; p-AMPK, phosphorylated adenosine
monophosphate-activated protein kinase; AICAR,
5-aminoimidazole-4-carboxamide riboside (agonist of adenosine
monophosphate-activated protein kinase); TSC2, tuberous sclerosis
complex 2. |
Discussion
Calcification of VSMCs is widely observed in a
variety of cardiovascular diseases. However, VSMC calcification has
also been revealed to be a manifestation of cellular senescence. In
other words, replicative senescence of VSMCs enhances
age-associated medial artery calcification (11,12). A
previous study revealed that the AMPK–TSC2–mTOR–S6K1 signal pathway
is involved in osteoblastic differentiation and calcification of
VSMCs and this may be markedly inhibited following adiponectin
treatment (13). Tan et al
(14) demonstrated that inhibition
of the PI3K/Akt/mTOR/S6K1 signal pathway decreased replicative
senescence in human VSMCs. GLP-1 analogue liraglutide may also
attenuate osteoblastic differentiation and calcification of human
VSMCs through regulating PI3K/Akt/mTOR/S6K1 signaling (15). The present study demonstrated that
AMPK/TSC2/mTOR/S6K1 signaling is involved in replicative senescence
of VSMCs. In the present study, replicative senescence of VSMCs was
induced in vitro by passaging for 15 generations.
Significant increases in SA-β-gal activity, p53 and p21 protein
expression as well as increase in cell size were observed in VSMCs
at 15 passages. Cell volume increases, changes in the expression of
replicative senescence-associated genes, such as p53 and p21, and
SA-β-gal activity are widely thought of as markers of cellular
senescence (3). Thus, the
replicative senescence model of VSMCs was successfully established
by extended passaging. Application of rapamycin, an mTORC1
inhibitor, significantly inhibited the replicative senescence of
VSMCs. Further molecular study revealed that the expression of
mTOR, as well as the phosphorylation of mTOR at Ser2448
and Thr2446 was significantly increased in VSMCs with
replicative senescence. Of note, rapamycin significantly inhibited
mTOR expression and phosphorylation. The present findings suggested
that the mTOR pathway has a role in VSMC senescence. Of note, to
the best of our knowledge, the present study was the first to
reveal that mTOR phosphorylation at Ser2448 at
Thr2446, but not at Ser2481, regulates
replicative senescence of VSMCs.
S6K1 is a specific downstream signaling protein and
substrate of mTORC1, which regulates S6K1 activation through its
phosphorylation at Thr389. In the present study, AICAR
and rapamycin applied during extended passaging inhibited mTOR
signaling and S6K1 phosphorylation as well as senescence of VSMCs.
These findings suggested that the mTOR/S6K1 pathway is involved in
the process of replicative senescence of VSMCs. A previous study
demonstrated that the activation of AMPK leads to the inhibition of
mTORC1 via a TSC2-dependent or -independent mechanism (6). To validate whether AMPK/TSC2 signaling
occurs upstream of mTOR activation, AMPK and TSC2 protein
expression and phosphorylation were determined. In senescent VSMCs,
AMPK and TSC2 protein phosphorylation, but not their expression,
was significantly decreased. Application of the AMPK activator
AICAR significantly inhibited SA-β-gal activity as well as p53 and
p21 protein expression, suggesting that AMPK activator inhibited
replicative senescence in VSMCs. However, application of AMPK
inhibitor Compound C did not significantly accelerate replicative
senescence. This result may require further comparative observation
using a small number of passages. Further molecular study
identified that AMPK activator AICAR significantly increased TSC2
phosphorylation, while inhibiting mTOR expression and
phosphorylation at Ser2448 and Thr2446
residues as well as S6K1 phosphorylation, while the effect of AMPK
inhibitor Compound C on mTOR signing was not significant. These
findings suggested that the downregulation of AMPK/TSC2 signaling
and subsequent upregulation of mTOR/S6K1 signaling are involved in
the senescence of VSMCs in vitro.
Of note, the mTOR inhibitor rapamycin significantly
inhibited mTOR expression and phosphorylation accompanied by a
significant increase in AMPKα and TSC2 phosphorylation. A previous
study using an animal model demonstrated that AMPK is highly
activated if mTORC1 signaling is impaired (16), suggesting that mTOR signaling exerts
a feedback regulation on AMPK/TSC2/mTOR signaling. These findings
suggested that mTOR signaling has an important role in the
replicative senescence of VSMCs and that activation of the
AMPK/TSC2/mTOR/S6K1 pathway may delay the replicative senescence of
VSMCs.
In conclusion, the AMPK/TSC2/mTOR/S6K1 signaling
pathway has a key role in the replicative senescence of VSMCs.
Interference with mTOR signaling may be used to inhibit replicative
senescence of VSMCs.
Acknowledgements
The present study was supported by the National
Basic Research Program of China (grant no. 2014CB910500), the
National Natural Science Foundation of China (grant nos. 81501212,
81370931) and the Hunan Province Special Health Research Projects
(grant no. A2015-04).
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Monk BA and George SJ: The effect of
ageing on vascular smooth muscle cell behaviour-a mini-review.
Gerontology. 61:416–426. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yin H and Pickering JG: Cellular
senescence and vascular disease: Novel routes to better
understanding and therapy. Can J Cardiol. 32:612–623. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Unterluggauer H, Hütter E, Voglauer R,
Grillari J, Vöth M, Bereiter-Hahn J, Jansen-Dürr P and Jendrach M:
Identification of cultivation-independent markers of human
endothelial cell. senescence in vitro. Biogerontology. 8:383–397.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hay N and Sonenberg N: Upstream and
downstream of mTOR. Genes Dev. 18:1926–1945. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zoncu R, Efeyan A and Sabatini DM: mTOR:
From growth signal integration to cancer, diabetes and ageing. Nat
Rev Mol Cell Biol. 12:21–35. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kawaguchi T, Hayakawa M, Koga H and
Torimura T: Effects of fucoidan on proliferation, AMP-activated
protein kinase, and downstream metabolism- and cell
cycle-associated molecules in poorly differentiated human hepatoma
HLF cells. Int J Oncol. 46:2216–2222. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Long YC, Cheng Z, Copps KD and White MF:
Insulin receptor substrates Irs1 and Irs2 coordinate skeletal
muscle growth and metabolism via the Akt and AMPK pathways. Mol
Cell Biol. 31:430–441. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li HL, Yin R, Chen D, Liu D, Wang D, Yang
Q and Dong YG: Long-term activation of adenosine
monophosphateactivated protein kinase attenuates
pressure-overload-induced cardiac hypertrophy. J Cell Biochem.
100:1086–1099. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang P, Hu X, Xu X, Fassett J, Zhu G,
Viollet B, Xu W, Wiczer B, Bernlohr DA, Bache RJ and Chen Y: AMP
activated protein kinase-alpha 2 deficiency exacerbates
pressure-overload-induced left ventricular hypertrophy and
dysfunction in mice. Hypertension. 52:918–924. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dimri GP, Lee X, Basile G, Acosta M, Scott
G, Roskelley C, Medrano EE, Linskens M, Rubelj I, Pereira-Smith O,
et al: A biomarker that identifies senescent human cells in culture
and in aging skin in vivo. Proc Natl Acad Sci USA. 92:9363–9367.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nakano-Kurimoto R, Ikeda K, Uraoka M,
Nakagawa Y, Yutaka K, Koide M, Takahashi T, Matoba S, Yamada H,
Okigaki M and Matsubara H: Replicative senescence of vascular
smooth muscle cells enhances the calcification through initiating
the osteoblastic transition. Am J Physiol Heart Circ Physiol.
297:H1673–H1684. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iijima K: Bone and calcium update;
diagnosis and therapy of bone metabolism disease update. Regulatory
mechanism of mammalian sirtuin SIRT1 in vascular calcification:
Impact of vascular smooth muscle cell senescence. Clin Calcium.
21:53–60. 2011.(In Japanese). PubMed/NCBI
|
|
13
|
Zhan JK, Wang YJ, Wang Y, Tang ZY, Tan P,
Huang W and Liu YS: Adiponectin attenuates the osteoblastic
differentiation of vascular smooth muscle cells through the
AMPK/mTOR pathway. Exp Cell Res. 323:352–358. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tan P, Wang YJ, Li S, Wang Y, He JY, Chen
YY, Deng HQ, Huang W, Zhan JK and Liu YS: The PI3K/Akt/mTOR pathway
regulates the replicative senescence of human VSMCs. Mol Cell
Biochem. 422:1–10. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhan JK, Wang YJ, Wang Y, Tang ZY, Tan P,
Huang W and Liu YS: The protective effect of GLP-1 analogue in
arterial calcification through attenuating osteoblastic
differentiation of human VSMCs. Int J Cardiol. 189:188–193. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Inoki K, Kim J and Guan KL: AMPK and mTOR
in cellular energy homeostasis and drug targets. Annu Rev Pharmacol
Toxicol. 52:381–400. 2012. View Article : Google Scholar : PubMed/NCBI
|














