Introduction
Gastric cancer is the most common malignant tumor of
digestive tract, and its morbidity and mortality are the second
highest in all malignant tumors in the world (1–3). After
clinical treatments, the five-year survival rate of patients with
early gastric cancer can reach 98%, but patients with advanced
gastric cancer have poor prognosis due to distant metastasis
(4). Tumor recurrence and metastasis
are the major causes of death in patients with gastric cancer
(5). Because the early symptoms of
gastric cancer are not obvious and the disease lack of specific
diagnostic markers, most patients are in the middle and late stage
of gastric cancer at diagnosis, and postoperative recurrence and
metastasis are easy to occur (6).
Therefore, researches on the molecular mechanism of recurrence and
metastasis of gastric cancer is of great significance for its
clinical diagnosis and treatment.
Dapper, antagonist of β-catenin (Dact) gene is a
signal pathway regulating molecule that exerts its function by
negatively regulating Wnt/β-catenin and Nodal/TGFβ signaling
pathways (7,8). Wnt/β-catenin and Nodal/TGFβ signaling
pathways participate in the regulation of the occurrence and
development of tumors, and Dact gene can therefore play a role in
the occurrence and development of tumors by affecting these
signaling pathways (9). Dact2 gene
is a member of Dact family that plays a role in tumors by
participating in the negative regulation of both Wnt/β-catenin and
Nodal/TGFβ signaling pathways (10).
As a tumor-suppressor gene in esophageal cancer, liver cancer, and
lymphoma, Dact2 inhibits the occurrence and development of tumors
by regulating the methylation of gene promoters (11–13).
However, it is still unknown whether Dact2 is regulated by other
factors. A study on the methylation in hepatocellular carcinoma and
gastric cancer tissues shows that promoter methylation is not found
in some patients with low expression of Dact2 (14), suggesting that Dact2 may be regulated
by other factors.
MicroRNA (miRNA or miR) is a class of small
non-encoding RNA molecules (18–22 nucleotides) that bind with the
3′-untranslated region (UTR) of mRNA to inhibit its translation
(15). miRNA plays important
regulatory roles in post-transcriptional levels of genes, and
widely participates in the proliferation, aging, apoptosis,
migration, differentiation, and drug resistance of cells (16). Studies show that the expression of a
variety of miRNA molecules is abnormal in gastric cancer, and
closely related with the occurrence and development of the disease.
For example, miR-1 and miR-200c act as tumor-suppressor genes in
gastric cancer, and the downregulation of their expression promotes
the migration of gastric cancer cells (17,18). In
addition, miR-33b-5p enhances the sensitivity of gastric cancer
cells to chemotherapeutic drugs and improves their clinical
efficacy (19). Of note,
Wnt/β-catenin signaling pathway that is regulated by Dact2, is also
regulated by various miRNA molecules (20). Therefore, miRNA is likely to be
involved in the regulation of Dact2 gene. In the present study, we
investigate the miRNA molecule that regulates Dact2 gene at tissue
and cellular levels, and try to elucidate the mechanism of action
of the miRNA molecule.
Patients and methods
Bioinformatics
Using miRNA molecule online prediction software
Targetscan 7.1 (www.targetscan.org/vert_71/), we predicted the miRNA
molecules that might regulate the expression of Dact2 gene
following the instructions on the website.
Patients
A total of 42 patients who received surgical
resection of gastric cancer tissues at our hospital between
December 2014 and February 2016 were included in the present study
(Table I). The 42 cases of resected
gastric cancer tissues were sliced and stained with hematoxylin and
eosin, and examined by two pathologists independently. Patients
with lymphatic metastasis were included into N1 group, while those
without lymphatic metastasis were included into N0 group. According
to 2003 WHO cancer classification criteria, 17 cases were included
into stage I, 12 cases were included into stage II, 10 cases were
included into stage III, and 3 cases were included into stage IV.
Among all 42 patients, 31 patients had moderate or high
differentiation, and 11 patients had low differentiation.
Tumor-adjacent tissues 5 cm away from tumor tissues were also
collected as controls. The tissue samples were frozen in liquid
nitrogen and stored at −80°C. Clinical information and pathological
data of the patients were collected. All procedures were approved
by the Ethics Committee of Harbin Medical University. Written
informed consents were obtained from all patients or their
families.
 | Table I.The clinic characteristics of 42
gastric cancer patients. |
Table I.
The clinic characteristics of 42
gastric cancer patients.
| Indexes | Number of
patients |
|---|
| Sex |
|
| Male | 27 |
|
Female | 15 |
| Age (year) |
|
| ≤60 | 26 |
| ≥60 | 16 |
| Tumor size (cm) |
|
| ≤5 | 30 |
| ≥5 | 12 |
| TNM stages |
|
| I | 17 |
| II | 12 |
| III | 10 |
| IV | 3 |
| Differentiation |
|
| Well | 16 |
|
Moderate | 15 |
| Poor | 11 |
| Lymphatic
metastasis |
|
| Yes | 23 |
| No | 19 |
Cells
Mixed gastric adenocarcinoma type Gastric cancer
MKN28 line was a derivative of MKN74 cells (which are also a
gastric adenocarcinoma cell line) (21). was cultured in RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS) at 37°C and 5%
CO2. When reaching 80–90% confluency, the cells were
passaged. The medium was replaced every two days. Cells with
passage numbers 3–6 were used for experiments.
Mixed gastric adenocarcinoma type MKN28 cells were
divided into negative control (NC) group, miR-214 mimics group and
miR-214 inhibitor group. On the day before transfection, mixed
gastric adenocarcinoma type MKN28 cells (2×105) in
log-phase growth were seeded onto 24-well plates containing
antibiotics-free RPMI-1640 medium supplemented with 10% FBS. When
reaching 70% confluency, 1.5 µl miR-214 mimics/inhibitor (20
pmol/µl; RiboBio, Guangzhou, China) and 1 µl Lipofectamine 2000
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) were added into
two individual vials containing 50 µl Opti Memi medium,
respectively. Five min later, the liquids in the two vials were
mixed together before standing still for another 20 min. Then, the
mixture was added onto the cells for an incubation of 6 h before
changing to RPMI-1640 medium supplemented with 10% FBS. The cells
were cultured at 37°C and 5% CO2 for 48 h before
use.
For rescue experiments, mixed gastric adenocarcinoma
type MKN28 cells (2×105) in miR-NC and miR-214 inhibitor
groups were seeded into 24-well plates containing antibiotics-free
RPMI-1640 medium supplemented with 10% FBS. When reaching 60%
confluency, mixed gastric adenocarcinoma type MKN28 cells in
miR-214 inhibitor group were infected by sh-DACT2 plasmid (Hanbio
Biotechnology Co., Ltd., Shanghai, China), while cells in miR-NC
group were infected by 0.5 µg NC plasmid. After being cultured at
37°C and under 5% CO2 for 6 h, the medium was refreshed
to newly made RPMI-1640 medium containing 10% FBS before
cultivation for 72 h. Then, RPMI-1640 medium containing 1 µg/ml
puro was added before incubation for 72 h.
Quantitative real-time polymerase
chain reaction (qRT-PCR)
Gastric cancer and tumor-adjacent tissues (100 mg)
were ground into powder using liquid nitrogen before addition of 1
ml TRIzol isolation reagent (Thermo Fisher Scientific, Inc.) for
lysis. After lysis, total RNA was extracted using phenol chloroform
method. The purity of RNA was determined by A260/A280 using
ultraviolet spectrophotometry (Nanodrop ND2000; Thermo Scientific,
Inc.). Then, cDNA was obtained by reverse transcription using
miScript II RT kit (Qiagen, Hilden, Germany) from 1 µg RNA and
stored at −20°C.
qRT-PCR was performed using miScript
SYBR® Green PCR kit (Qiagen) and the reaction system was
composed of 10 µl qRT-PCR-Mix, 0.5 µl upstream primer
(5′-ACAGCAGGCACAGACAGGCAGT-3′), 0.5 µl downstream primer (universal
primer provided by the kit), 2 µl cDNA and 7 µl ddH2O.
Reaction protocol was initial denaturation at 95°C for 10 min, and
40 cycles of 95°C for 1 min and 60°C for 30 sec.
Cell-Counting kit (CCK)-8 assay
The sample cells were inoculated in 96-well plates
at a density of 2,000/well. At 0, 24, 48 and 72 h, 20 µl CCK-8 (5
g/l; Beyotime Institute of Biotechnology, Beijing, China) was added
onto the cells. After being incubated at 37°C for 2 h, absorbance
(490 nm) of each well was determined, and cell proliferation curves
were plotted. Each group was tested in 3 replicate wells and the
values were averaged.
Transwell assay
Matrigel chambers (Corning Inc., Corning, NY, USA)
were used to determine the migration and invasion abilities of
cells. Matrigel was first diluted with serum-free RPMI-1640 medium
at a ratio of 1:2. In upper chamber, 50 µl diluted Matrigel was
added and kept at 37°C for 1 h. Then, 1×105 cells and
200 µl serum-free RPMI-1640 medium were added into the upper
chamber. In the lower chamber, 500 µl RPMI-1640 medium supplemented
with 10% FBS was added. After incubation for 24 h, the cells in
upper chamber were wiped by cotton swab. Then, the chamber was
fixed using 4% formaldehyde for 10 min at room temperature, and
then subjected to Giemsa's staining for 1 min. After washing for 3
times, cells that moved to the other side of the chamber were
counted under a microscope (5 fields; magnification, ×200) to
evaluate migration and invasion abilities.
Western blot analysis
Cells in each group were trypsinized and collected.
Then, cold Radio-Immunoprecipitation Assay (RIPA) lysis buffer (600
µl; Beyotime Institute of Biotechnology) was mixed with the
samples. Then, the mixture was lysed for 30 min on ice, and then
centrifuged at 12,000 rpm and 4°C for 10 min. Bicinchoninic acid
(BCA) protein concentration determination kit (RTP7102; Real-Times
Biotechnology Co., Ltd., Beijing, China) was used to determine
protein concentration in the supernatant. After mixing protein
samples (6 µl) with 5X sodium dodecyl sulfate loading buffer, the
mixture was denatured by boiling in water bath for 10 min.
Afterwards, 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (100 V) was performed using the samples. Then, the
proteins were electro-transferred to polyvinylidene difluoride
(PVDF) membranes on ice (250 mA, 1 h) before being blocked with 50
g/l skimmed milk at room temperature for 1 h. Afterwards, rabbit
anti-human β-catenin and DACT2 polyclonal primary antibodies (both
1:1,000) and mouse anti-human GAPDH primary antibody (1:4,000; both
Abcam, Cambridge, UK) were added onto the membranes before
incubation at 4°C overnight. Then, the membrane was extensively
washed with phosphate-buffered saline with Tween-20 (PBST) for 5
times of 5 min, and incubated with goat anti-mouse horseradish
peroxidase-conjugated secondary antibodies (1:4,000; Abcam) at room
temperature for 1 h. Subsequently, the membrane was washed with
PBST for 5 times of 5 min before the membrane was developed with
enhanced chemiluminescence detection kit (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) for imaging. We used Image lab v3.0
software (Bio-Rad Laboratories, Inc., Hercules, CA, USA) to acquire
and analyze imaging data. The relative expression of target
proteins was expressed with the ratio against GAPDH.
Dual luciferase reporter assay
Wild-type (WT) and mutant seeding regions of miR-214
in 3′-UTR of DACT2 gene were chemically synthesized before adding
Spe-1 and HindIII restriction sites, and then cloned into
pMIR-REPORT luciferase reporter plasmids (0.5 µg) with WT or mutant
3′-UTR DNA sequences, which were transfected together with miR-214
mimics into HEK293T cells. Following incubation for 24 h, cells
were processed using dual luciferase reporter assay kit according
to the manufacturer's manual (Beyotime Institute of Biotechnology),
and fluorescence intensity was determined by GloMax 20/20
luminometer (Promega, Fitchburg, WI, USA). Fluorescence values of
each group of were measured using renilla fluorescence activity as
internal reference.
Statistical analysis
All results were analyzed using SPSS 17.0
statistical software (SPSS, Inc., Chicago, IL, USA), and all data
were shown as means ± SD. Comparison between groups was performed
using group t-test. The results among multiple groups were compared
using one-way analysis of variance followed by Dunnett's test as
the post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of miR-214 is elevated, but
expression of Dact2 mRNA is decreased in gastric cancer tissues,
being closely correlated with the invasion, metastasis, occurrence
and development of gastric cancer
To search miRNA molecules that might participate in
the regulation of Dact2 gene, Targetscan 7.1 website was used.
Search of Dact2 on the website showed that miR-214 was likely to be
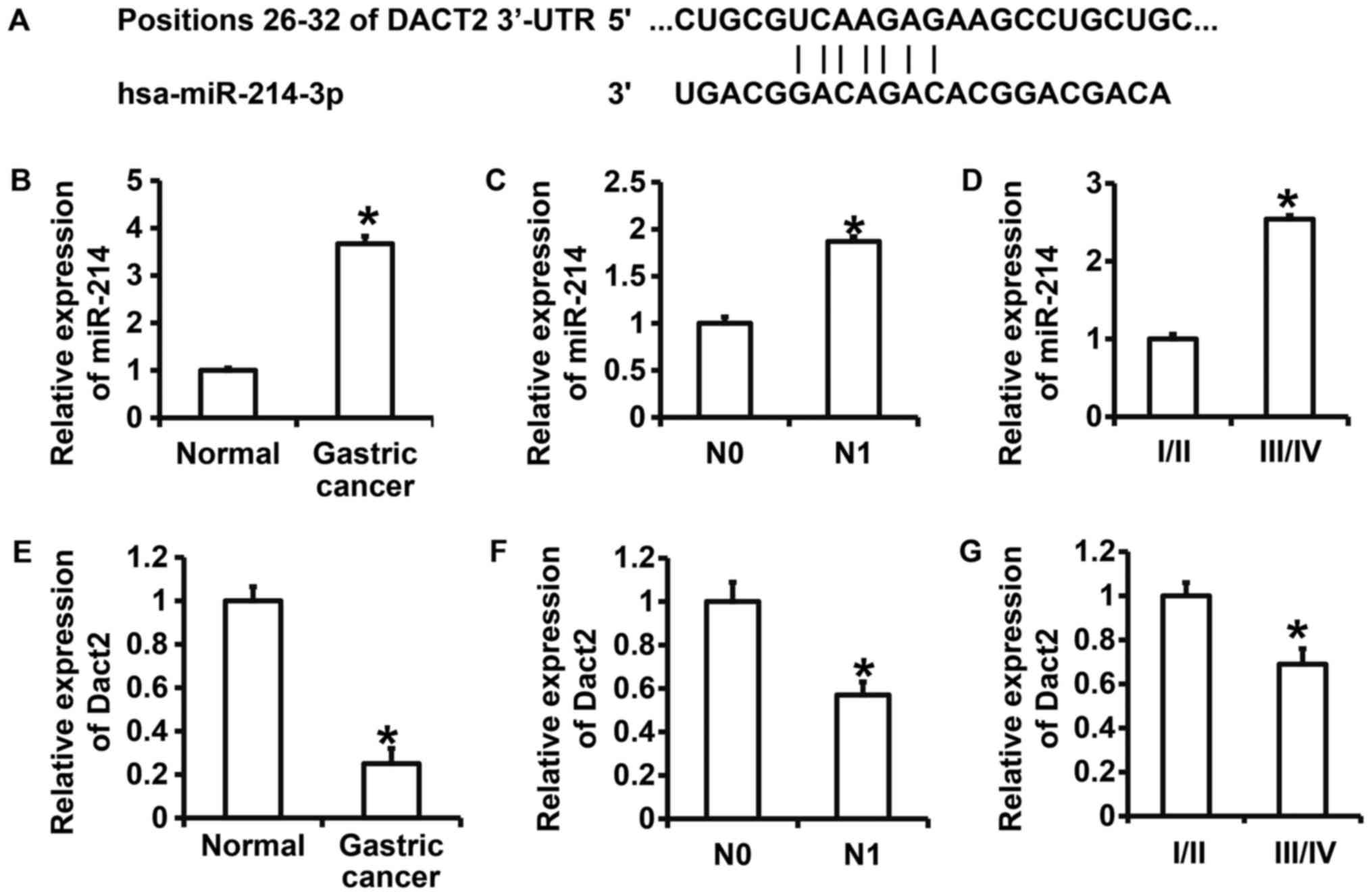
involved in the regulation of Dact2 expression (Fig. 1). To further test the expression of
miR-214 and Dact2 in gastric cancer, qRT-PCR was performed. The
data showed that expression of miR-214 in gastric cancer tissues
was significantly higher than that in tumor-adjacent tissues
(P<0.05) (Fig. 1B). In addition,
miR-214 expression in N1 group was significantly higher than that
in N0 group (P<0.05) (Fig. 1C).
Of note, miR-214 expression in gastric cancer tissues from patients
at stages III/IV was significantly higher than that from patients
at stages I/II (P<0.05) (Fig.
1D). By contrast, the expression of Dact2 mRNA in gastric
cancer tissues was significantly decreased than that in
tumor-adjacent tissues (P<0.05) (Fig.
1E). The expression of Dact2 mRNA in gastric cancer tissues in
N1 group was significantly lower than that in N0 group (P<0.05)
(Fig. 1F). Moreover, the expression
of Dact2 mRNA in gastric cancer tissues from patients at stages
III/IV was significantly reduced than that from patients at stages
I/II (P<0.05) (Fig. 1G). The
results suggest that expression of miR-214 is elevated, but
expression of Dact2 mRNA is decreased in gastric cancer tissues,
being closely correlated with the invasion, metastasis, occurrence
and development of gastric cancer.
miR-214 promotes, but Dact2 inhibits
the proliferation of gastric cancer MKN28 cells in vitro
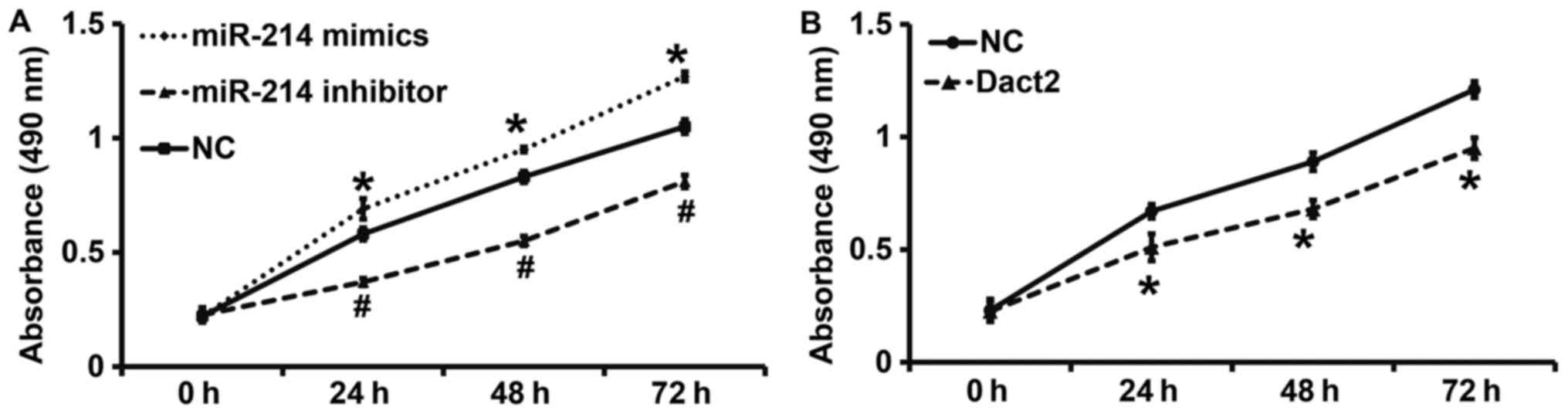
To examine the proliferation of mixed gastric
adenocarcinoma type MKN28 cells, CCK-8 assay was carried out. The
data showed that the absorbance of mixed gastric adenocarcinoma
type MKN28 cells transfected with miR-214 mimics was significantly
higher than that from cells in NC group at all time points
(P<0.05), while the absorbance of the cells transfected with
miR-214 inhibitor was significantly lower than that from cells in
NC group at all time points (P<0.05) (Fig. 2A). By contrast, the absorbance of
mixed gastric adenocarcinoma type MKN28 cells with overexpression
of Dact2 gene was significantly lower than that of cells in NC
group at all time points (P<0.05) (Fig. 2B). The results indicate that miR-214
promotes, but Dact2 inhibits the proliferation of gastric cancer
MKN28 cells in vitro.
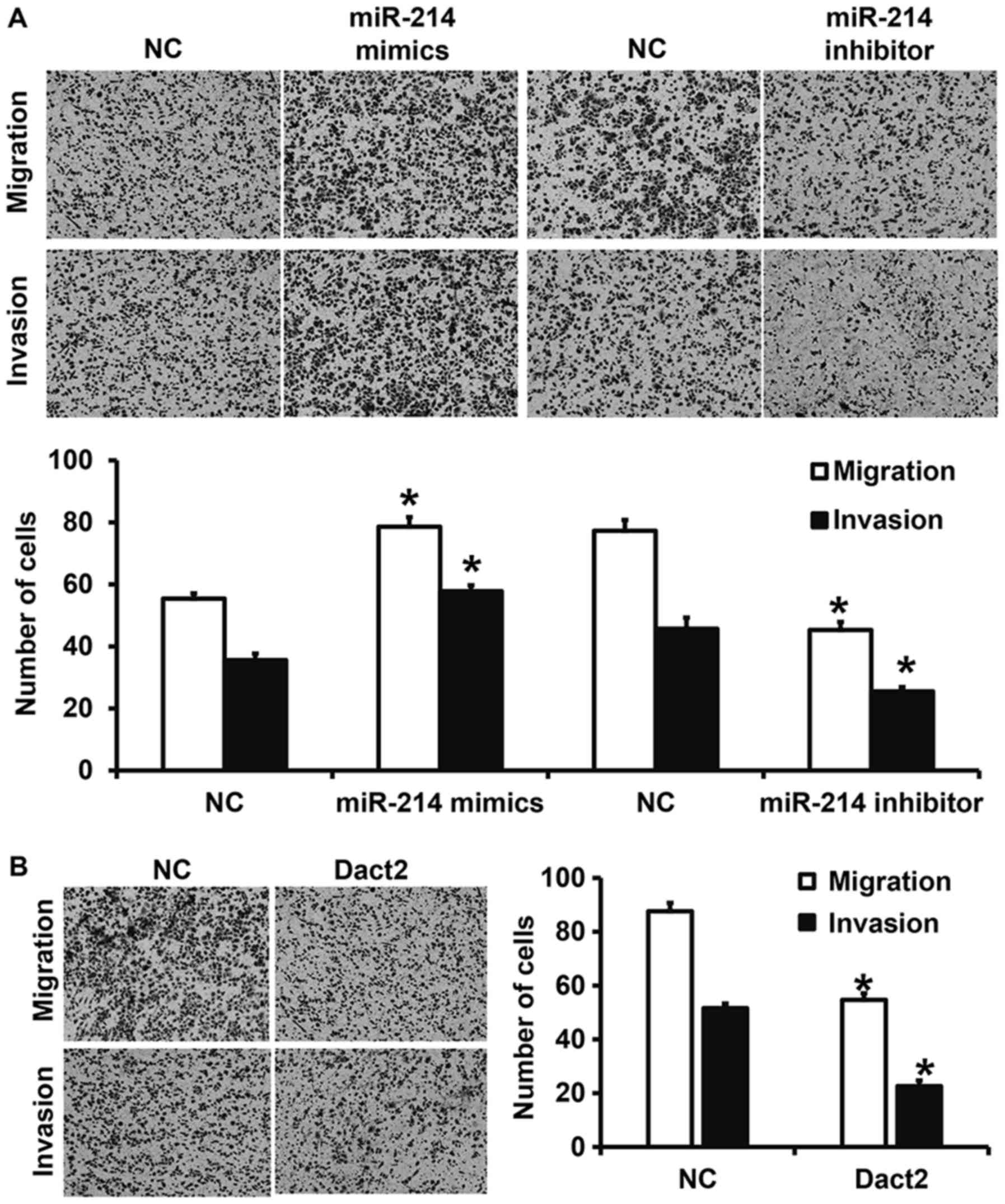
Downregulation of miR-214 expression or upregulation
of Dact2 expression inhibits the migration and invasion of mixed
gastric adenocarcinoma type MKN28 cells. To investigate migration
and invasion abilities of mixed gastric adenocarcinoma type MKN28
cells, Transwell assay was employed. The data showed that the
numbers of cells in miR-214 mimics group that crossed chamber
membrane in migration and invasion assays were increased than those
in NC group (P<0.05). By contrast, cell counts in miR-214
inhibitor group that crossed chamber membrane in migration and
invasion assays were lower than those in NC group (P<0.05).
Similarly, cell count in miR-214 mimics group that crossed chamber
membrane in invasion assay was reduced than that in miR-NC group
(P<0.05) (Fig. 3A). Moreover, the
numbers of cells in Dact2 group that crossed chamber membrane in
migration and invasion assays were significantly reduced compared
with those in NC group (P<0.05) (Fig.
3B). The results suggest that downregulation of miR-214
expression or upregulation of Dact2 expression inhibits the
migration and invasion of mixed gastric adenocarcinoma type MKN28
cells.
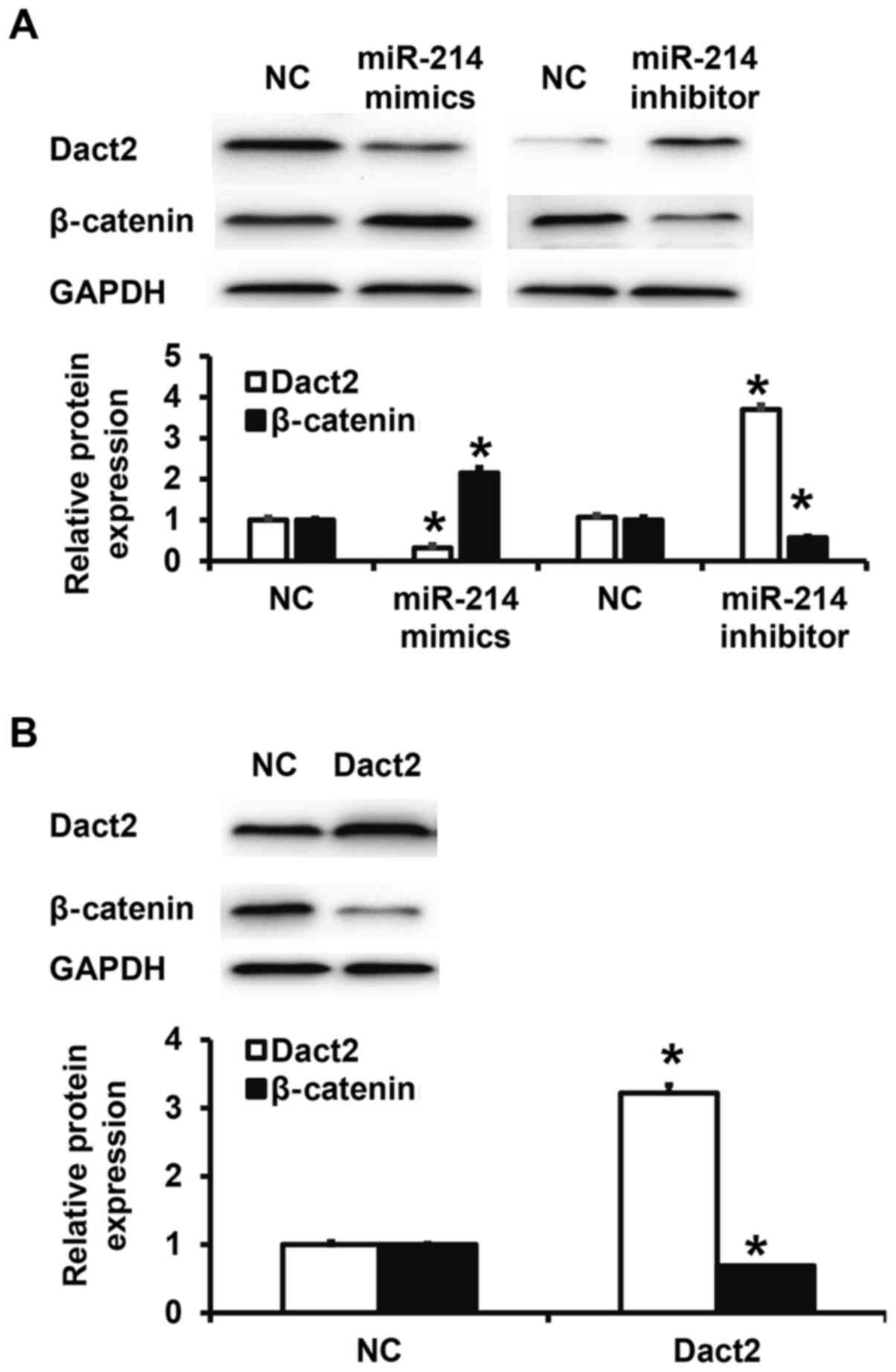
miR-214 regulates the expression of Dact2 protein
and its downstream β-catenin protein in mixed gastric
adenocarcinoma type MKN28 cells. To test Dact2 and β-catenin
protein expression in mixed gastric adenocarcinoma type MKN28
cells, western blotting was performed. Quantification of western
blots showed that Dact2 protein expression in miR-214 mimics group
was significantly reduced than that in NC group (P<0.05), while
β-catenin expression in miR-214 mimics group was significantly
higher than that in NC group (P<0.05). By contrast, Dact2
protein expression in miR-214 inhibitor group was significantly
higher than that in NC group (P<0.05), while β-catenin
expression in miR-214 inhibitor group was significantly lower than
that in NC group (P<0.05) (Fig.
4A). Moreover, transfection with Dact2 plasmid significantly
elevated Dact2 expression in mixed gastric adenocarcinoma type
MKN28 cells (P<0.05), but significantly reduced β-catenin
expression (P<0.05) (Fig. 4B).
The results indicate that miR-214 regulates the expression of Dact2
protein and its downstream β-catenin protein in mixed gastric
adenocarcinoma type MKN28 cells.
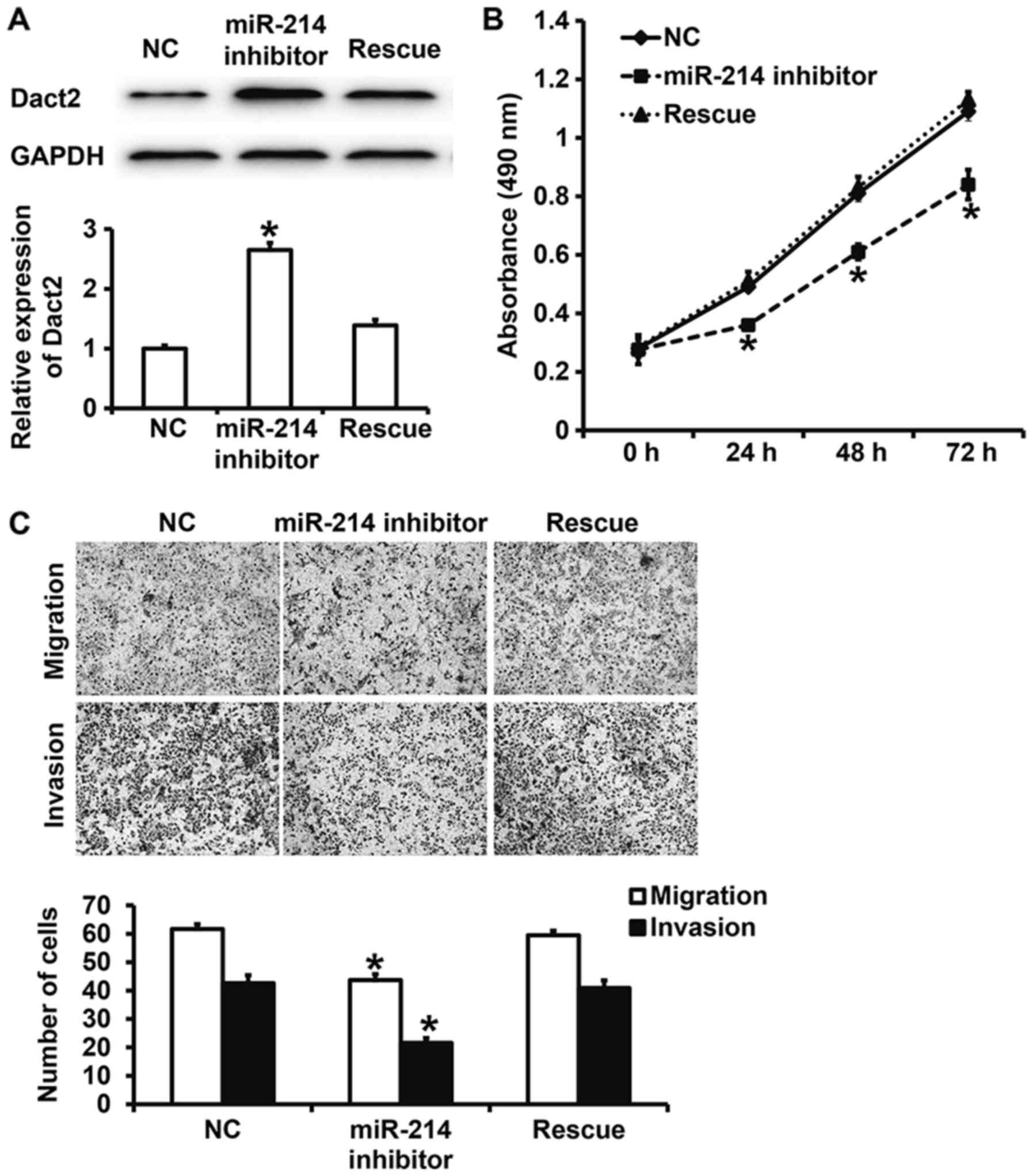
Dact2 reverses the effect of miR-214
on the proliferation, migration and invasion of gastric cancer
MKN28 cells
To test how the regulation of Dact2 by miR-214
affects biological functions of mixed gastric adenocarcinoma type
MKN28 cells, we silenced and rescued the expression of miR-214.
Western blotting showed that Dact2 expression in miR-214 inhibitor
group was significantly higher than that in NC group (P<0.05),
while that in rescue group was significantly lower than that in
miR-214 inhibitor group (P<0.05) (Fig. 5A). CCK-8 assay showed that the
absorbance of cells in miR-214 inhibitor group was significantly
reduced than that in NC group at all time points (P<0.05), while
that in rescue group was significantly higher than that in miR-214
inhibitor group at all time points (P<0.05), reaching a level
similar to NC group (Fig. 5B).
Moreover, Transwell assay showed that the numbers of cells in
miR-214 inhibitor group that crossed chamber membrane in migration
and invasion assays were significantly lower than those in NC
group, respectively (P<0.05), while those in rescue group were
significantly higher than those in miR-214 inhibitor group
(P<0.05) (Fig. 5C). The results
suggest that Dact2 reverses the effect of miR-214 on the
proliferation, migration and invasion of mixed gastric
adenocarcinoma type MKN28 cells.
miR-214 can bind with the 3′-UTR
seeding region of Dact2 mRNA to regulate its expression
To identify the interaction between miR-214 and the
3′-UTR of Dact2 mRNA, dual luciferase reporter assay was performed.
The fluorescence value of cells co-transfected with miR-214 mimics
and pMIR-REPORT-WT luciferase reporter plasmids was significantly
lower than that in NC group (P<0.05). By contrast, the
fluorescence value of cells co-transfected with miR-214 mimics and
pMIR-REPORT-mutant luciferase reporter plasmids was not
significantly different from that in NC group (P>0.05) (Fig. 6). The result indicates that miR-214
can bind with the 3′-UTR seeding region of Dact2 mRNA to regulate
its expression.
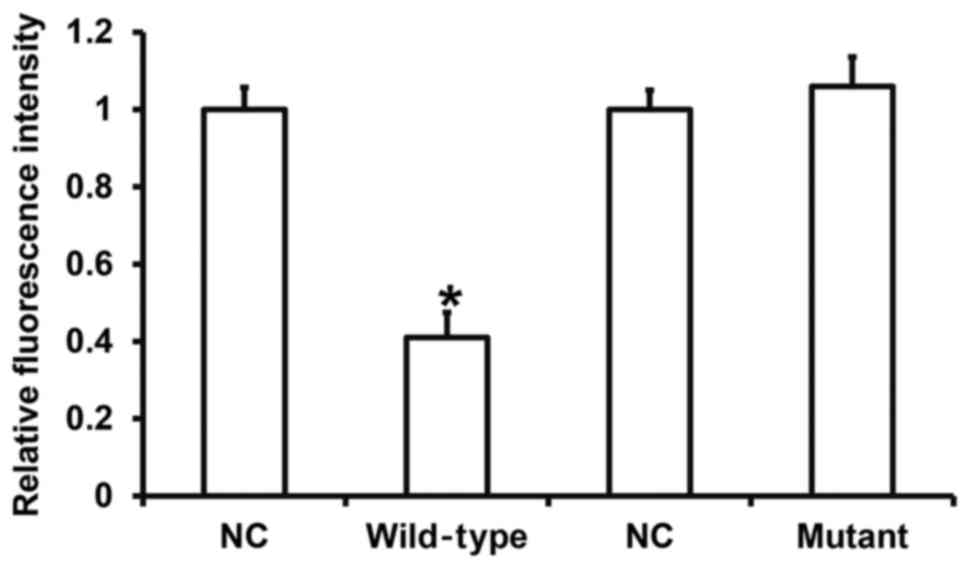 | Figure 6.Identification of interaction between
miR-214 and Dact2 mRNA using dual luciferase reporter assay. WT and
mutant seeding regions of miR-214 in 3′-UTR of DACT2 gene were
chemically synthesized before adding Spe-1 and HindIII restriction
sites, and then cloned into pMIR-REPORT luciferase reporter
plasmids (0.5 µg) with WT or mutant 3′-UTR DNA sequences, which
were transfected together with miR-214 mimics into HEK293T cells.
Following incubation for 24 h, cells were processed using dual
luciferase reporter assay kit according to the manufacturer's
manual, and fluorescence intensity was determined by GloMax 20/20
luminometer. Fluorescence values of each group of were measured
using renilla fluorescence activity as internal reference.
*P<0.05 compared with respective NC group. NC, negative control;
WT, wild-type; miRNA, microRNA; Dact2, dapper, antagonist of
β-catenin 2; UTR, untranslated region. |
Discussion
Gastric cancer is a systemic disease, and its
recurrence and metastasis are the major causes of poor prognosis
(4). Similar to other solid tumors,
the proliferation and metastasis of gastric cancer cells are
regulated by multiple genes and multiple factors (22,23).
miRNA has powerful post-transcriptional regulation function and a
wide application prospect in tumor therapy. It is reported that
many miRNA molecules are expressed abnormally in gastric cancer
cells and play important roles in the occurrence and development of
gastric cancer (24).
Wnt/β-catenin signaling pathway is abnormally
activated in many tumors, and promotes tumor formation,
maintenance, invasion, metastasis and drug resistance (25). As an important negative regulator of
Wnt/β-catenin signaling pathway, Dact2 has become a focus among
researchers (26). The deletion of
Dact2 leads to the activation of Wnt/β-catenin signaling pathway,
and facilitates the occurrence and development of colon cancer,
esophageal cancer and liver cancer. Studies show that the deletion
of Dact2 is related to the methylation of promoter region. For
example, methylation of the Dact2 gene promoter leads to the
silencing of Dact2 gene, and promotes epithelial mesenchymal
transition and cytoskeletal rearrangement in breast cancer
(27). Moreover, aberrant
methylation of Dact2 promoter in squamous cell carcinoma of
esophagus is the key reason for silenced expression of Dact2, and
this ultimately promotes the invasion and metastasis of squamous
cell carcinoma (28). However,
promoter methylation does not necessarily exist in tumor tissues
with downregulated Dact2 expression, suggesting that Dact2 has
other regulatory mechanisms.
miRNA molecules are key regulators of mRNA at
posttranscriptional levels, and they are almost involved in all
physiological and pathological activities of the body. Studies have
shown that a variety of miRNA molecules have been involved in the
regulation of Wnt/β-catenin signaling pathway. For example, Liu
et al discover that miR-155 promotes the proliferation and
metastasis of SW-480 cells by regulating Wnt/β-catenin signaling
pathway (29). Wei et al find
that miR-638 inhibits the metastasis of cervical cancer cells by
regulating Wnt/β-catenin signaling pathway (30). The discovery of these miRNA molecules
provides new targets for the regulation of Wnt/β-catenin signaling
pathway. In the present study, we use bioinformatics to predict
that miR-214 may regulate Dact2 gene. Using qRT-PCR, we identified
upregulated expression of miR-214 in gastric cancer tissues, which
is positively related with clinical staging and lymphatic
metastasis. Moreover, expression of Dact2 gene is downregulated,
and negatively correlated with clinical staging and lymphatic
metastasis. Cellular functional experiments show that miR-214, as
an oncogene, promotes the proliferation, migration and invasion of
gastric cancer cells. By contrast, Dact2 acts as a tumor-suppressor
gene that inhibits the proliferation, migration and invasion of
gastric cancer cells. These results demonstrate that the function
and expression of miR-214 are reversely related with Dact2,
suggesting that miR-214 may exert its biological functions via
Dact2. Western blotting shows that miR-214 regulates the expression
of Dact2. Furthermore, rescue experiments demonstrate that Dact2
inhibits the tumor-promoting effect of miR-214. Indeed, dual
luciferase reporter assay shows that miR-214 directly binds with
the 3′-UTR of Dact2 mRNA.
In conclusion, miR-214 downregulates the expression
of Dact2 gene, activates Wnt/β-catenin signaling pathway, and
promotes the proliferation, migration and invasion of gastric
cancer. Therefore, miR-214 is a potential molecular therapeutic
target and biomarker for gastric cancer. In our future studies, we
will investigate the correlation between the expression of miR-214
and Dact2 in tissues, and test the function and mechanism of action
of miR-214 at animal level.
Acknowledgements
The authors would like to thank their department and
research team for their help and dedication.
Funding
Project of Heilongjiang Provincial Health Bureau
(grant no. 2011-094).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ designed the study. WF, YF and SG were
responsible for performing experiments. LZ and WF analyzed the
data. All authors collaborated to interpret results and develop the
manuscript. The final version of the manuscript has been read and
approved by all authors.
Ethics approval and consent to
participate
All procedures performed in the current study were
approved by the Ethics Committee of Harbin Medical University.
Written informed consent was obtained from all patients or their
families.
Patient consent for publication
Written informed consents for publication of any
associated data and accompanying images were obtained from all
patients or their parents, guardians or next of kin.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wei TT, Wang LL, Yin JR, Liu YT, Qin BD,
Li JY, Yin X, Zhou L and Zhong RQ: Relationship between red blood
cell distribution width, bilirubin, and clinical characteristics of
patients with gastric cancer. Int J Lab Hematol. 39:497–501. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Choi HI, Choi JP, Seo J, Kim BJ, Rho M,
Han JK and Kim JG: Helicobacter pylori-derived extracellular
vesicles increased in the gastric juices of gastric adenocarcinoma
patients and induced inflammation mainly via specific targeting of
gastric epithelial cells. Exp Mol Med. 49:e3302017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhuo C, Ying M, Lin R, Wu X, Guan S and
Yang C: Negative lymph node count is a significant prognostic
factor in patient with stage IV gastric cancer after palliative
gastrectomy. Oncotarget. 8:71197–71205. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Singh P, Toom S and Huang Y: Anti-claudin
18.2 antibody as new targeted therapy for advanced gastric cancer.
J Hematol Oncol. 10:1052017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin XL, Xu Q, Tang L, Sun L, Han T, Wang
LW and Xiao XY: Regorafenib inhibited gastric cancer cells growth
and invasion via CXCR4 activated Wnt pathway. PLoS One.
12:e01773352017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sosa Arias LA, Orduz Cuspoca AF and Gómez
Bernal BM: Deregulation of microRNAs in gastric cancer: Up
regulation by miR-21 and miR-106. Rev Gastroenterol Peru. 37:65–70.
2017.(In Spanish). PubMed/NCBI
|
|
7
|
Komsky-Elbaz A and Roth Z: Effect of the
herbicide atrazine and its metabolite DACT on bovine sperm quality.
Reprod Toxicol. 67:15–25. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schubert FR, Sobreira DR, Janousek RG,
Alvares LE and Dietrich S: Dact genes are chordate specific
regulators at the intersection of Wnt and Tgf-β signaling pathways.
BMC Evol Biol. 14:1572014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sensiate LA, Sobreira DR, Da Veiga FC,
Peterlini DJ, Pedrosa AV, Rirsch T, Joazeiro PP, Schubert FR,
Collares-Buzato CB, Xavier-Neto J, et al: Dact gene expression
profiles suggest a role for this gene family in integrating Wnt and
TGF-β signaling pathways during chicken limb development. Dev Dyn.
243:428–439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang S, Dong Y, Zhang Y, Wang X, Xu L,
Yang S, Li X, Dong H, Xu L, Su L, et al: DACT2 is a functional
tumor suppressor through inhibiting Wnt/β-catenin pathway and
associated with poor survival in colon cancer. Oncogene.
34:2575–2585. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schussel JL, Kalinke LP, Sassi LM, de
Oliveira BV, Pedruzzi PA, Olandoski M, Alvares LE, Garlet GP and
Trevilatto PC: Expression and epigenetic regulation of DACT1 and
DACT2 in oral squamous cell carcinoma. Cancer Biomark. 15:11–17.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu Y, Yan W, Liu X, Jia Y, Cao B, Yu Y, Lv
Y, Brock MV, Herman JG, Licchesi J, et al: DACT2 is frequently
methylated in human gastric cancer and methylation of DACT2
activated Wnt signaling. Am J Cancer Res. 4:710–724.
2014.PubMed/NCBI
|
|
13
|
Jia Y, Yang Y, Brock MV, Zhan Q, Herman JG
and Guo M: Epigenetic regulation of DACT2, a key component of the
Wnt signalling pathway in human lung cancer. J Pathol. 230:194–204.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang X, Yang Y, Liu X, Herman JG, Brock
MV, Licchesi JD, Yue W, Pei X and Guo M: Epigenetic regulation of
the Wnt signaling inhibitor DACT2 in human hepatocellular
carcinoma. Epigenetics. 8:373–382. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Manne RK, Agrawal Y, Bargale A, Patel A,
Paul D, Gupta NA, Rapole S, Seshadri V, Subramanyam D, Shetty P and
Santra MK: A microRNA/Ubiquitin ligase feedback loop regulates
slug-mediated invasion in breast cancer. Neoplasia. 19:483–495.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Michael JV, Wurtzel JGT, Mao GF, Rao AK,
Kolpakov MA, Sabri A, Hoffman NE, Rajan S, Tomar D, Madesh M, et
al: Platelet microparticles infiltrating solid tumors transfer
miRNAs that suppress tumor growth. Blood. 130:567–580. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xie M, Dart DA, Guo T, Xing XF, Cheng XJ,
Du H, Jiang WG, Wen XZ and Ji JF: MicroRNA-1 acts as a tumor
suppressor microRNA by inhibiting angiogenesis-related growth
factors in human gastric cancer. Gastric Cancer. 21:41–54. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li M, Gu K, Liu W, Xie X and Huang X:
MicroRNA-200c as a prognostic and sensitivity marker for platinum
chemotherapy in advanced gastric cancer. Oncotarget. 8:51190–51199.
2017.PubMed/NCBI
|
|
19
|
Yang X, Zhao Q, Yin H, Lei X and Gan R:
MiR-33b-5p sensitizes gastric cancer cells to chemotherapy drugs
via inhibiting HMGA2 expression. J Drug Target. 25:653–660. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shen X, Pan B, Zhou H, Liu L, Lv T, Zhu J,
Huang X and Tian J: Differentiation of mesenchymal stem cells into
cardiomyocytes is regulated by miRNA-1-2 via WNT signaling pathway.
J Biomed Sci. 24:292017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Capes-Davis A, Theodosopoulos G, Atkin I,
Drexler HG, Kohara A, MacLeod RA, Masters JR, Nakamura Y, Reid YA,
Reddel RR and Freshney RI: Check your cultures! A list of
cross-contaminated or misidentified cell lines. Int J Cancer.
127:1–8. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zeng XQ, Wang J and Chen SY: Methylation
modification in gastric cancer and approaches to targeted
epigenetic therapy (Review). Int J Oncol. 50:1921–1933. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hayakawa Y, Fox JG and Wang TC: The
origins of gastric cancer from gastric stem cells: Lessons from
mouse models. Cell Mol Gastroenterol Hepatol. 3:331–338. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jafari N and Abediankenari S: MicroRNA-34
dysregulation in gastric cancer and gastric cancer stem cell.
Tumour Biol. 39:10104283177016522017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang M, Chen C, Geng J, Han D, Wang T,
Xie T, Wang L, Wang Y, Wang C, Lei Z and Chu X: Targeting KDM1A
attenuates Wnt/β-catenin signaling pathway to eliminate
sorafenib-resistant stem-like cells in hepatocellular carcinoma.
Cancer Lett. 398:12–21. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo YL, Shan BE, Guo W, Dong ZM, Zhou Z,
Shen SP, Guo X, Liang J and Kuang G: Aberrant methylation of DACT1
and DACT2 are associated with tumor progression and poor prognosis
in esophageal squamous cell carcinoma. J Biomed Sci. 24:62017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xiang T, Fan Y, Li C, Li L, Ying Y, Mu J,
Peng W, Feng Y, Oberst M, Kelly K, et al: DACT2 silencing by
promoter CpG methylation disrupts its regulation of
epithelial-to-mesenchymal transition and cytoskeleton
reorganization in breast cancer cells. Oncotarget. 7:70924–70935.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang M, Linghu E, Zhan Q, He T, Cao B,
Brock MV, Herman JG, Xiang R and Guo M: Methylation of DACT2
accelerates esophageal cancer development by activating Wnt
signaling. Oncotarget. 7:17957–17969. 2016.PubMed/NCBI
|
|
29
|
Liu N, Jiang F, Han XY, Li M, Chen WJ, Liu
QC, Liao CX and Lv YF: MiRNA-155 promotes the invasion of
colorectal cancer SW-480 cells through regulating the
Wnt/β-catenin. Eur Rev Med Pharmacol Sci. 22:101–109.
2018.PubMed/NCBI
|
|
30
|
Wei H, Zhang JJ and Tang QL: MiR-638
inhibits cervical cancer metastasis through Wnt/β-catenin signaling
pathway and correlates with prognosis of cervical cancer patients.
Eur Rev Med Pharmacol Sci. 21:5587–5593. 2017.PubMed/NCBI
|