Introduction
Nasopharyngeal cancer (NPC) derives from the
epithelial lining of the nasopharynx (1). Although the incidence rate of NPC is
relatively low in the majority of the world, representing about
0.7% of the global cancer burden, it is comparatively higher in
southern China and Southeast Asia with ~7.3% (2,3).
Previous studies have revealed that inheritance, environmental
influences and the Epstein-Barr virus serve essential functions in
NPC development (4–9).
The GATA family contains GATA1, GATA2, GATA3, GATA4,
GATA5 and GATA6 (10). They all
contain a zinc finger domain, which can bind to the specific
consensus DNA sequence, 5′-A/TGATAA/G-3′ (11). Multiple reports demonstrated that the
GATA family serves essential functions in cell proliferation and
differentiation (12,13). The GATA family is separated into two
subfamilies; the GATA1/2/3 subfamily and the GATA4/5/6 subfamily.
Several previous studies have indicated that GATA4 regulates gene
transcription by binding to GATA elements (14,15).
Additionally, GATA4 was demonstrated to interact with other
transcription factors, including Tbx5 and Nkx2-5 (16). GATA4 acts as a transcriptional
regulator, regulating hypertrophic growth of the heart (17). Defects or mutations of GATA4
contribute to a series of cardiac problems, including abnormal
ventral folding, congenital heart disease as well as hypoplasia of
the ventricular myocardium (18).
Additionally, GATA4 is crucial to the gastrointestinal, respiratory
and reproductive systems and cancer (19). In addition, GATA4 contributes to
cancer development and serves as a potent prognostic predictor
(20–23). However, the molecular mechanism of
GATA4 in NPC remains poorly understood.
Tumor metastasis occurs due to
epithelial-mesenchymal transition (EMT) in cancer cells (24,25). EMT
is a crucial phenotypic event; it regulates embryonic development,
wound healing and tissue remodeling (26). The main characteristics of EMT are
decreased epithelial markers and increased mesenchymal markers
(26). Several studies have
indicated that GATA4 regulates EMT in many types of cancer
(27,28). However, the function of GATA4 in NPC
remains unknown.
The current study demonstrated that GATA4 was
upregulated in NPC and the expression of GATA4 was associated with
tumor size, metastasis and pathological grade. Additionally, the
upregulation of GATA4 was associated with a poor prognosis in NPC.
In addition, GATA4 enhanced EMT through the regulation of
SLUG expression in NPC and facilitated cell invasion.
Furthermore, cell proliferation was also demonstrated to be
regulated by GATA4. In conclusion, the current study indicated that
GATA4 acts as an oncogene in NPC.
Materials and methods
Cell culture and tissue samples
The human NPC cell line, 5-8F, was purchased from
the Cell Bank of Type Culture Collection of Chinese Academy of
Sciences (Shanghai, China). Cells were maintained in RPMI 1640
medium (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 10% fetal bovine serum (FBS; HyClone; GE
Healthcare Life Sciences, Logan, UT, USA) and 1%
penicillin-streptomycin solution at 37°C with 5% CO2.
The human immortalized normal nasopharyngeal epithelial cell line,
NP69 obtained from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China), was cultured in Keratinocyte-SFM medium
supplemented with epidermal growth factor (EGF; both Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) at 37°C with 5%
CO2.
The adjacent normal tissue (ANT; 0.25 cm from tumor
tissue) and tumor tissue samples were obtained from patients who
were diagnosed with NPC. A total of 42 pairs of ANT and NPC
specimens were collected from Department of Otorhinolaryngology,
The Second Affiliated Hospital of Zhejiang University School of
Medicine (Hangzhou, China) between April 2015 and October 2016. A
total of 19 males and 23 females participated in the study with a
mean age of 55.3±4.3 years (range, 35–82 years). The current study
was approved by The Ethics Committee of The Second Affiliated
Hospital of Zhejiang University School of Medicine. All patients
provided written informed consent.
Transfection
Cells (3×105) were transfected with 2.5
µg pcDNA3.1 empty vector (Youbio, Hunan, China), 2.5 µg FLAG-GATA4
(pcDNA3.1-GATA4; Youbio) or 30 nM small interfering (si)RNA
scrambled (SCR) or GATA4 siRNA (siGATA4; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according the manufacturer's protocol.
Following 48 h transfection at 37°C, cells were collected by
centrifugation (800 × g, 4°C, 3 min) and used for the further
experiments. The sequences of siRNA were as follows: SCR,
5′-UUCUCCGAACGUGUCACGU-3′; siGATA4,
5′-AATCTCGTAGATATGTTTGAC-3′.
Western blotting
Whole protein was extracted from cells and tissues
using radioimmunoprecipitation assay buffer [50 mM Tris-HCl (pH
7.4), 150 mM NaCl and 1% NP-40] with protease inhibitor cocktail
(Roche Diagnostics, Basel, Switzerland). The concentration of
protein was measured using a bicinchoninic acid kit (Pierce; Thermo
Fisher Scientific, Inc.). Subsequently, equal amount of proteins
(40 µg) were separated by 8% SDS-PAGE and then transferred onto
polyvinylidene difluoride membranes. The membranes were blocked by
5% non-fat milk at room temperature for 1 h, and then incubated
with primary antibodies at 4°C overnight. The membranes were washed
three times and incubated with horseradish peroxidase
(HRP)-conjugated secondary antibodies for 1 h at room temperature.
Finally, blots were visualized using an enhanced chemiluminescence
kit (Pierce; Thermo Fisher Scientific, Inc.). The antibodies used
were as follows: Rabbit anti-GATA4 (1:2,000; cat, 19530-1-AP;
Proteintech Group, Inc., Chicago, IL, USA), rabbit anti-TWIST
(1:1,000; cat: 25465-1-AP; Proteintech Group, Inc.), rabbit
anti-a-catenin (1:3,000; cat, 12831-1-AP; Proteintech Group, Inc.),
rabbit antibodies from the EMT kit (1:2,000; cat, 9783; Cell
Signaling Technology, Inc., Danvers, MA, USA) and mouse
anit-β-actin (1:3,000; cat, 6276; Abcam, Cambridge, UK), goat
anti-rabbit (HRP-conjugated; 1:5,000; cat, ab205718; Abcam) and
goat anti-mouse (HRP-conjugated; 1:3,000; cat, ab205719; Abcam).
All the experiments were repeated at least three times.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from tissue samples and
cells using a Qiagen RNeasy Mini kit (Qiagen GmbH, Hilden, Germany)
according to the manufacturer's protocol. Then, cDNA was reverse
transcribed from mRNA at 42°C for 30 min using a PrimeScript RT-PCR
kit (Takara Bio, Inc., Otsu, Japan). Subsequently, Power
SYBR™ Green PCR Master Mix (Thermo Fisher Scientific,
Inc.) was used to perform qPCR analyses in an Applied Biosystems
7500 (Applied Biosystems; Thermo Fisher Scientific, Inc.).
Thermocycling conditions were as follows: 5 min at 98°C, followed
by 34 cycles of denaturation at 98°C for 30 sec, annealing at 57°C
for 30 sec and extension at 72°C for 35 sec. Primers were as
follows: GATA4 forward, 5′-CCCAATCTCGTAGATATGTTTGAC-3′ and reverse,
5′-CCGTTCATCTTGTGGTAGAG-3′; E-cadherin forward,
5′-AAACATCATTGATGCAGACC-3′ and reverse,
5′-GATAGATTCTTGGGTTGGGTC-3′; α-catenin forward,
5′-TGTTACACAGGTTACAACCCT-3′ and reverse,
5′-GCAGCCTTCATCAAATCACC-3′; N-cadherin forward,
5′-CAAAGCCTGGAACATATGTG-3′ and reverse, 5′-GTTTGAAAGGCCATATGTGG-3′;
vimentin forward, 5′-CTCCACGAAGAGGAAATCCA-3′ and reverse,
5′-GATTTGTACCATTCTTCTGCCT-3′; Slug forward,
5′-ACACATACAGTGATTATTTCCC-3′ and reverse,
5′-ACTGTAGTCTTTCCTCTTCAT-3′; Snail forward,
5′-TCTAATCCAGAGTTTACCTTCCAG-3′ and reverse,
5′-TGAAGTAGAGGAGAAGGACGA-3′; Twist forward,
5′-GTACATCGACTTCCTCTACC-3′ and reverse,
5′-GAAACAATGACATCTAGGTCTC-3′; ZEB1 forward,
5′-TTACACCTTTGCATACAGAACCC-3′ and reverse,
5′-TTTACGATTACACCCAGACTGC-3′; GAPDH forward,
5′-ATTTCCTGGTATGACAACGA-3′ and reverse,
5′-TTGATGGTACATGACAAGGTG-3′. GAPDH was used as an internal control.
Relative mRNA levels were calculated using the 2−ΔΔCq
method (29). All the experiments
were repeated at least three times.
Transwell invasion assay
A Transwell invasion assay was performed using
Transwell chambers (8 µm pore size; Costar; Corning Incorporated,
Corning, NY, USA) according to the manufacturer's protocol. In
brief, GATA4 was overexpressed or knocked down in 5-8F cells.
Following a 48-h transfection, 20,000 cells were placed into the
upper chamber, which was coated with Matrigel (BD Biosciences, San
Jose, CA, USA). The cells in the upper chamber were maintained with
serum-free RPMI medium 1640, the lower plate was filled with RPMI
medium 1640 which contained 10% FBS. Following a 16-h incubation at
37°C, cells on the undersurface were fixed with 10% methanol at
room temperature for 10 min and stained with 0.5% crystal violet at
room temperature for 10 min. Six fields were randomly selected and
cells were counted under a light microscope (magnification, ×20).
The number of invaded cells was normalized to the control group.
All experiments were repeated at least three times.
Dual-luciferase reporter assay
The promoter region sequences of SLUG and
TWIST were cloned into a pGL3-basic plasmid (Biofeng,
Beijing, China). For the dual-luciferase reporter assay, 5-8F cells
were co-transfected with 0.5 µg Renilla, 0, 0.5, 1 or 2 µg
pcDNA3.1-GATA4 (Vigene Biosciences, Rockville, MD, USA) and 2 µg
pGL3-SLUG or 2 µg pGL3-TWIST using Lipofectamine 2000 (Thermo
Fisher Scientific, Inc.). Renilla was used to normalize the
firefly luciferase activity in each well. Following transfection
for 24 h at 37°C, the Dual-Luciferase® Reporter Assay
System (Promega Corporation, Madison, WI, USA) was used to measure
the relative luciferase activity. All the experiments were repeated
at least three times.
Cell counting kit-8 (CCK-8) assay
To investigate the effect of GATA4 on cell
proliferation in NPC, 5-8F cells transfected with the empty vector,
FLAG-GATA4, scramble small interfering (si)RNA (SCR) or GATA4 siRNA
(siGATA4) were placed into 96 well plate at a density of 2,000
cells per well and maintained at 37°C. Then, the cell viability
rate was measured using CCK-8 (Beyotime Institute of Biotechnology,
Haimen, China) at different time points (0, 24, 48 and 72 h). CCK-8
solution (20 µl) was added to each well and incubated at 37°C for
60 min; the absorbance of each well was measured at 450 nm. All the
experiments were repeated at least three times.
Colony formation assay
Transfected 5-8F cells (5×103) were
seeded in 6-well plates and maintained in RPMI 1640 containing 10%
FBS for 2 weeks at 37°C. Cells were then fixed with 10% methanol at
room temperature for 10 min and stained with 0.5% crystal violet at
room temperature for 10 min. The number of colonies was counted
under a light microscope (magnification, ×20). All the experiments
were repeated at least three times.
Wound healing assay
A wound-healing assay was performed to determine the
effect of GATA4 on cell migration. Briefly, GATA4 was overexpressed
or knocked down in 5-8F cells. When the cells were grown to 90–100%
confluence in RPMI 1640 at 37°C, a 200-µl pipette tip was used to
scratch the cells. Next, cells were incubated with serum-free RPMI
1640 at 37°C and the migration distance was observed under a light
microscope (magnification, ×20) at different time points (0 and 36
h). The relative migrating distance was the ratio of migration
distance at 36 h and the distance measured at 0 h. All the
experiments were repeated at least three times.
ChIP assay
A ChIP assay was performed using an EZ-ChIP kit
(Merck KGaA, Darmstadt, Germany) according to the manufacturer's
protocol. In brief, 5-8F cells (~3×106) were used at a
density of 80–90% confluence and were fixed with 1% formaldehyde
(Beyotime Institute of Biotechnology) at 37°C for 30 min. Following
the addition of lysis buffer containing RNase A (Merck KGaA), the
lysate was sonicated (20 kHz; amplitude, 40%; 20 cycles, 1 sec on
and 1 sec off; 4°C) to break cross-linked chromatin into 200 to
1,000-bp fragments, ChIP assays were performed using 5 µl
anti-GATA4 antibody (cat. no. 19530-1-AP; Proteintech Group, Inc.)
at 4°C overnight. Protein G agarose beads (60 µl) were used to
purify antibody-bound DNA fragments at 4°C over 1 h. Beads were
then washed with each 1 ml low salt immune complex wash buffer,
high salt immune complex wash buffer, LiCl immune complex wash
buffer and TE buffer (all Merck KGaA) at 4°C. DNA was isolated from
the immunoprecipitates using a DNA kit (Tiangen Biotech Co., Ltd.)
and quantified using RT-qPCR analysis with the specific primer
pairs: Slug, forward 5′-CTGGATTATGCCTCTGTGAT-3′ and reverse
5′-TGGTATTTATTTGCTGGTAG-3′; twist, forward
5′-AAGGGATGGACCTGAAACGG-3′ and reverse 5′-GGCAAACTGGAAGCAGCAAA-3′.
The reaction mixture contained 2 µl sample, 1 µl primer (10 nM), 10
µl 2X MIX buffer (TransGen Biotech Co., Ltd., Beijing, China) and 7
µl RNase-free. RT-qPCR conditions were as follows: 5 min at 98°C,
followed by 35 cycles of denaturation at 98°C for 30 sec, annealing
at 56°C for 30 sec and extension at 72°C for 20 sec. Input DNA and
DNA immunoprecipitated by anti-IgG antibody served as a positive
and negative control, respectively. The results were calculated
using the 2−ΔΔCq method (29).
Statistical analysis
All data were presented as mean + standard
deviation. SPSS (version 18.0; SPSS, Inc., Chicago, IL, USA) was
used to analyze data. The association between GATA4 expression and
clinical information was analyzed by Pearson's χ2 test.
Kaplan-Meier plots and log-rank tests were used to analyze the
rates of survival. Comparisons between two groups were analyzed by
Student's t-test. Multiple groups were analyzed by one-way analysis
of variance followed by Tukey's test. P<0.05 was considered to
indicate a statistically significant difference.
Results
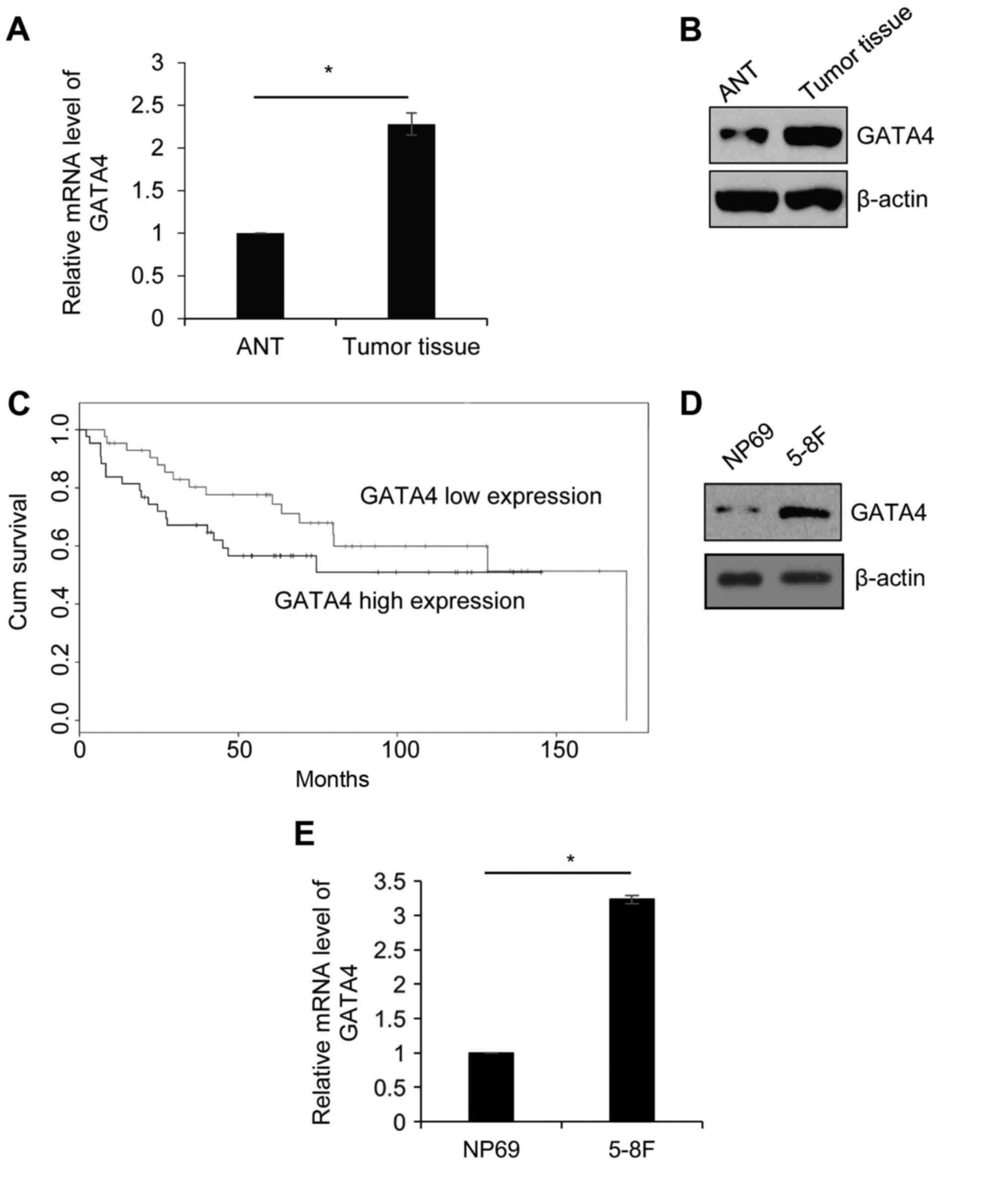
GATA4 is upregulated in NPC tissues
and the cell line, 5-8F
In order to explore the function of GATA4 in NPC, 42
paired NPC tissue and ANT samples were collected from patients who
were diagnosed with NPC. The expression of GATA4 was detected by
RT-qPCR. As presented in Fig. 1A,
the expression of GATA4 in tumor tissue was significantly higher
compared with its expression in ANT. Western blotting was performed
to detect the protein level of GATA4. Consistent with the RT-qPCR
analysis, the expression of GATA4 was markedly higher in tumor
tissue compared with ANT (Fig. 1B).
Additionally, the association between GATA4 expression and
clinicopathology of NPC was analyzed. High GATA4 expression was
significantly associated with large tumor size, metastasis and high
pathological grade (Table I). In
addition, according to survival curves analysis, the patients with
high GATA4 expression had lower survival rates compared with the
patients who had low GATA4 expression (Fig. 1C). Next, the expression of GATA4 was
further investigated in the NPC cell line, 5-8F, and the human
immortalized normal nasopharyngeal epithelial cell line, NP69, was
used as control. The results demonstrated that the protein and mRNA
levels of GATA4 were higher in 5-8F cells compared with that of
NP69 cells (Fig. 1D and E). The
results indicated that GATA4 serves an essential function in
NPC.
 | Table I.Clinicopathological variables in 42
nasopharyngeal cancer patients. |
Table I.
Clinicopathological variables in 42
nasopharyngeal cancer patients.
|
|
| GATA4 protein
expression, n |
|
|---|
|
|
|
|
|
|---|
| Variables | Patients, n | Low (n=13) | High (n=29) | P-value |
|---|
| Sex |
|
|
| 0.555 |
|
Male | 19 | 5 | 14 |
|
|
Female | 23 | 8 | 15 |
|
| Age, years |
|
|
| 0.588 |
|
<55 | 20 | 7 | 13 |
|
|
≥55 | 22 | 6 | 16 |
|
| Tumor size,
diameter |
|
|
| 0.038 |
| Small
(<1 cm) | 16 | 8 | 8 |
|
| Large
(≥1 cm) | 26 | 5 | 21 |
|
| Pathological
stage |
|
|
| 0.032 |
|
I–II | 13 | 7 | 6 |
|
|
III–IV | 29 | 6 | 23 |
|
| Metastasis |
|
|
| 0.015 |
|
Yes | 30 | 6 | 24 |
|
| No | 12 | 7 | 5 |
|
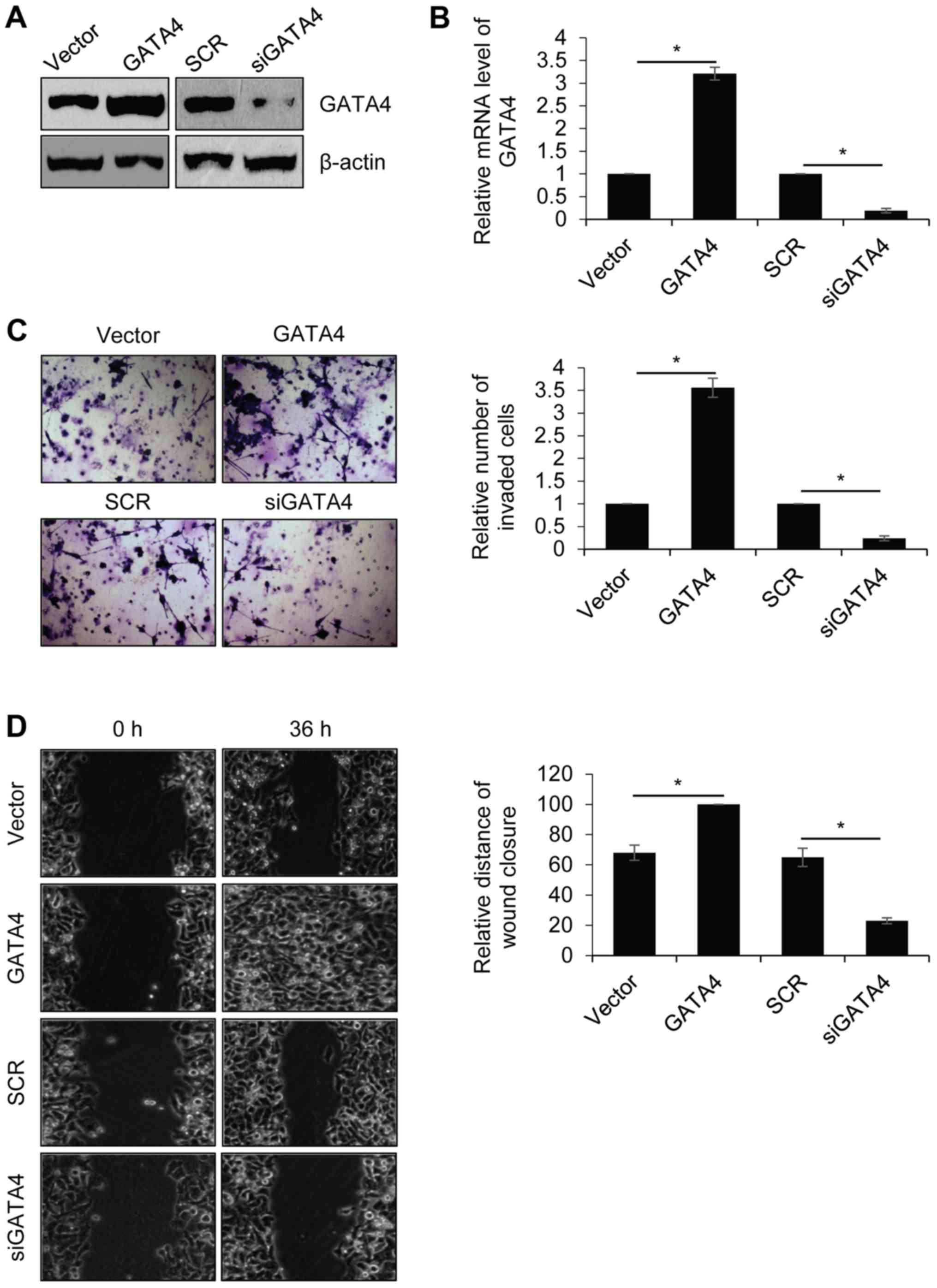
Overexpression of GATA4 promotes NPC
cell migration and invasion
Previous studies by our group indicated that GATA4
was positively correlated with metastasis (22,30).
Therefore, the role of GATA4 in the regulation of NPC cell
migration and invasion was explored. The human highly metastatic
NPC cell line, 5-8F, was used for further experiments. 5-8F cells
were transfected with the empty vector, FLAG-GATA4, SCR or siGATA4.
Western blotting and RT-qPCR were performed to determine GATA4
expression in 5-8F cells following transfection. The results
revealed that the protein and mRNA levels of GATA4 were markedly
(Fig. 2A) and significantly
(Fig. 2B) increased, respectively,
in 5-8F cells transfected with FLAG-GATA4 compared with the vector
group. The protein and mRNA levels of GATA4 were markedly and
significantly decreased, respectively, in 5-8F cells transfected
with siGATA4 compared with the SCR group. Transwell and wound
healing assays were performed to determine the effect of GATA4 on
cell invasion and migration. As demonstrated in Fig. 2C, overexpression of GATA4
significantly promoted the invasion of 5-8F cells compared with the
vector group. Additionally, GATA4 inhibition significantly
inhibited the invasion of 5-8F cells compared with the SCR group.
In addition, overexpression of GATA4 significantly facilitated cell
migration in 5-8F cells compared with the vector group, whereas
GATA4 inhibition significantly suppressed cell migration in 5-8F
cells compared with the SCR group (Fig.
2D). These results demonstrated that GATA4 promotes NPC cell
migration and invasion.
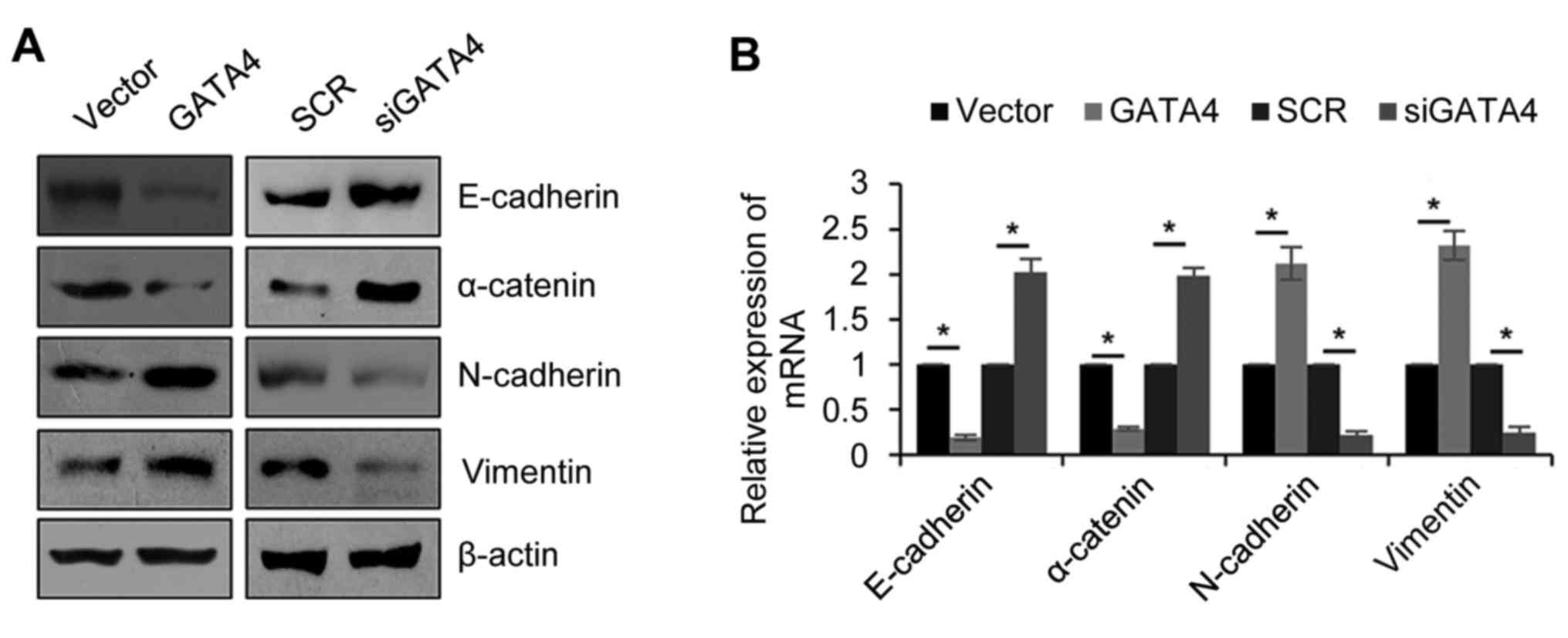
GATA4 facilitates EMT in NPC
cells
EMT is a complex process, which is associated with
metastasis. Therefore, the effect of GATA4 on EMT was explored. The
5-8F cells were transfected with the empty vector, FLAG-GATA4, SCR
or siGATA4. Western blotting and RT-qPCR were then used to
determine the GATA4 expression in the cells following transfection.
As expected, the protein levels of the epithetical markers
(E-cadherin and α-catenin) were markedly decreased and mesenchymal
markers (N-cadherin and vimentin) were markedly increased when
GATA4 was overexpressed compared with the vector group (Fig. 3A). Additionally, knocking down GATA4
expression markedly increased the epithetical markers and decreased
the mesenchymal markers compared with the SCR group. The results
were similar and significant when the mRNA levels were analyzed
(Fig. 3B). These results revealed
that GATA4 facilitates EMT in NPC cells.
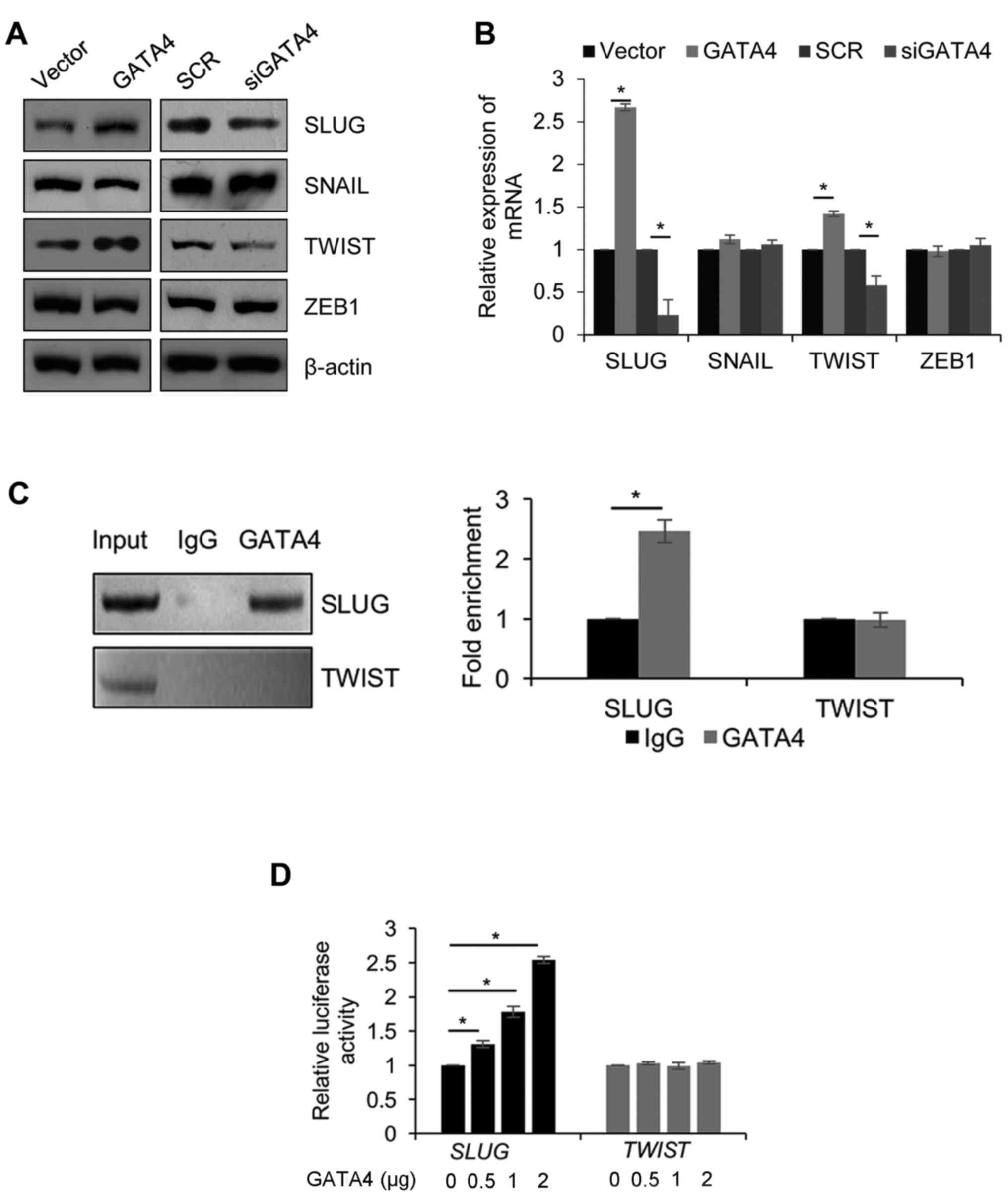
SLUG is transcriptionally activated by
GATA4
To further decipher the molecular mechanisms of
GATA4 in EMT, the expression of genes that serve roles in EMT
(SLUG, SNAIL, TWIST and ZEB1) were analyzed to identify whether
they are regulated by GATA4. Western blotting demonstrated that the
protein expression of SLUG and TWIST was increased when GATA4 was
overexpressed; in contrast, the protein expression of SLUG and
TWIST was decreased when GATA4 was depleted (Fig. 4A). The results were similar and
significant when the mRNA levels were analyzed (Fig. 4B). However, the expression of SNAIL
and ZEB1 did not significantly change at the protein and mRNA
level. Thus, it was assumed that GATA4 transcriptionally regulated
SLUG and TWIST. To validate this hypothesis, a
chromatin immunoprecipitation (ChIP) assay was performed to detect
the interaction between GATA4, and promoter regions of SLUG
and TWIST. The results of the ChIP assay demonstrated that
GATA4 directly bound the promoter region of SLUG, but not
the promoter region of TWIST (Fig. 4C). Subsequently, a dual-luciferase
reporter assay was performed in 5-8F cells, which were
co-transfected with pGL3-SLUG or pGL3-TWIST and Renilla, and
incubated with increasing concentrations of pcDNA3.1-GATA4. GATA4
significantly increased the luciferase activity of the pGL3-SLUG
group, but not in the pGL3-TWIST group (Fig. 4D). These results suggested that GATA4
transcriptionally activates SLUG. Although GATA4 did not
transcriptionally regulate TWIST, GATA4 may regulate
TWIST through the regulation of other proteins.
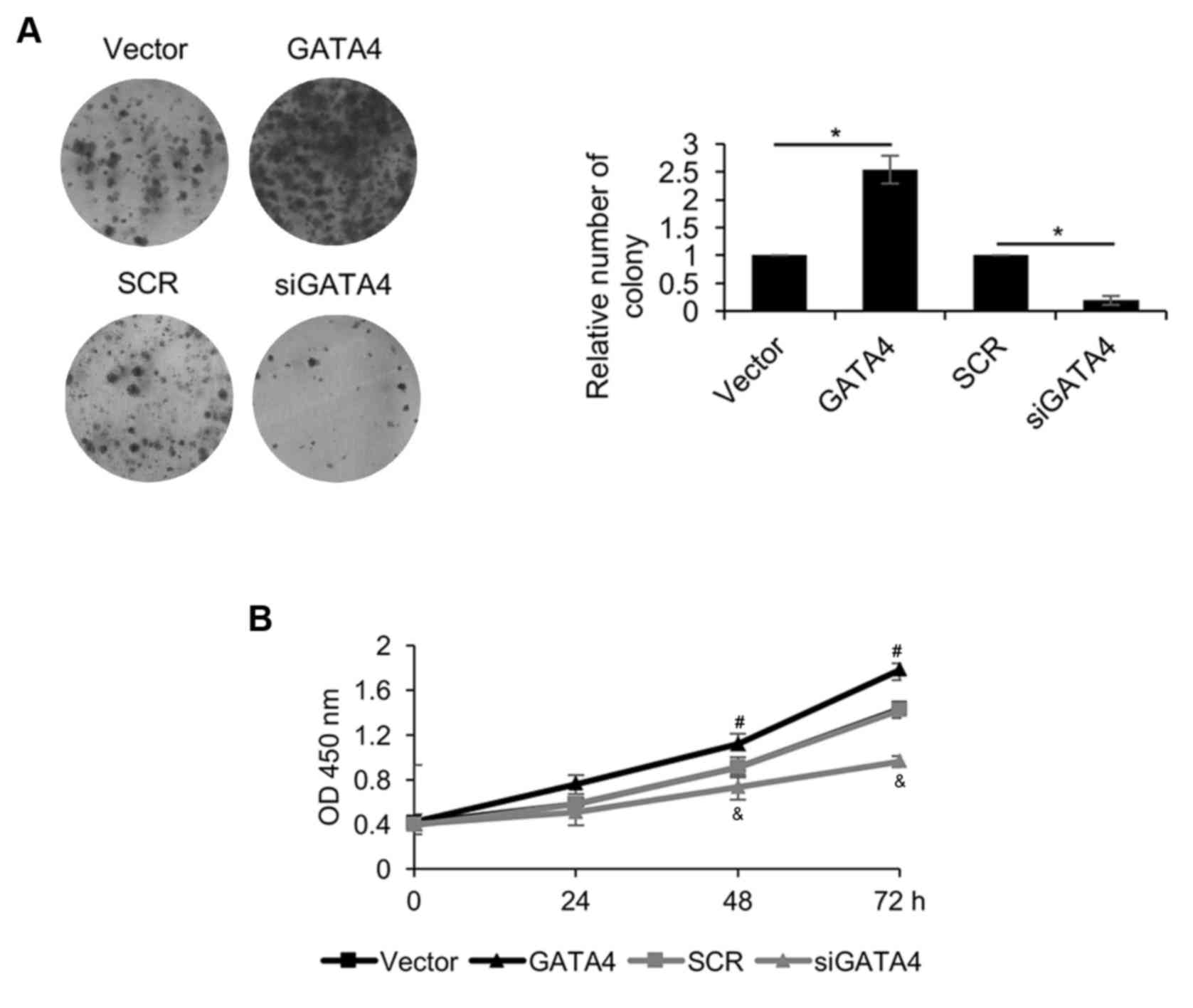
GATA4 promotes cell proliferation in
NPC cells
Due to the association between GATA4 expression and
tumor size, the role of GATA4 in the regulation of NPC cell
proliferation was explored using CCK-8 and colony formation assays.
The colony formation assay revealed that overexpression of GATA4 in
5-8F cells significantly increased the number of colonies compared
with the vector group, but GATA4 inhibition in 5-8F cells
significantly decreased the number of colonies compared with the
SCR group (Fig. 5A). The CCK-8 assay
demonstrated that overexpression of GATA4 significantly promoted
cell proliferation compared with the vector group (Fig. 5B). However, GATA4 inhibition
significantly suppressed 5-8F cell proliferation compared with the
SCR group. These results suggested that GATA4 promotes cell
proliferation in NPC cells.
Discussion
The results of the current study revealed that GATA4
was upregulated in patients' tissue samples and the 5-8F NPC cell
line. In addition, the increased expression of GATA4 in these
samples and cells was associated with larger tumor size, high
pathological grade, metastasis and poor prognosis. The survival
curve analysis indicated that patients with high GATA4 expression
survived for ~145 months and patients with low GATA4 expression
survived for ~175 months, suggesting that GATA4 may be a novel
predictor of NPC. Metastasis is an important event in cancer
development, previous reports demonstrated that nasopharyngeal
carcinoma frequently led to lymph node metastasis (31,32). As
stated above, the current study demonstrated that high GATA4
expression was associated with metastasis. Several functional
experiments, including Transwell invasion and wound healing assays,
were performed to investigate the mechanism of GATA4 on cell
migration and invasion. The results indicated that GATA4
significantly promoted migration and invasion in 5-8F cells.
EMT is a crucial phenotypic event; it regulates
embryonic development, wound healing and tissue remodeling
(33,34). A number of studies have indicated
that GATA4 regulates EMT in several cancers (27,28).
However, the role of GATA4 in EMT remains unclear. In order to
explore whether GATA4 regulates EMT, GATA4 was overexpressed or
knocked down in 5-8F cells. It was demonstrated that the
overexpression of GATA4 increased the expression of N-cadherin and
vimentin, whereas the expression of E-cadherin and α-catenin was
decreased. Conversely, GATA4 inhibition increased the expression of
E-cadherin and α-catenin, but the expression of N-cadherin and
vimentin was decreased. Therefore, GATA4 promoted EMT in 5-8F
cells.
EMT is regulated by several transcription factors,
including SLUG, SNAIL, TWIST and ZEB1 (35–38).
Subsequently, ChIP and luciferase reporter assays demonstrated that
GATA4 transcriptionally activated SLUG. Although previous
studies have demonstrated that TWIST is also regulated by
GATA4 (39), the current study did
not identify GATA4 as a transcriptional regulator of TWIST.
Thus, it is assumed that GATA4 may regulate TWIST through
other proteins. In summary, GATA4 promotes EMT through
transcriptionally activating SLUG in NPC.
Cell proliferation is also an important event in
cancer development (40). Due to the
association between the expression of GATA4 and tumor size, it was
hypothesized that GATA4 may regulate cancer cell proliferation. As
expected, the current study revealed that GATA4 facilitated cell
proliferation in 5-8F cells.
To the best of our knowledge, the current study is
the first study to demonstrate that GATA4 is upregulated in NPC and
serves as an oncogene. The high expression of GATA4 predicted a
poor prognosis for patients with NPC. Additionally, GATA4 promoted
metastasis and proliferation in NPC cells. Therefore, GATA4 may be
a novel biomarker for NPC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YZ and BY conceived and designed the present study.
YZ, HC and BY performed the experiments. YZ and BY wrote the paper.
All authors reviewed and edited the manuscript. The final version
of the manuscript was read and approved by all authors, and each
author believes that the manuscript represents honest work.
Ethics approval and consent to
participate
The current study has been approved by The Ethics
Committee of The Second Affiliated Hospital of Zhejiang University
School of Medicine. All patients were informed that their tissues
would be used in the current study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zeng Z, Huang H, Zhang W, Xiang B, Zhou M,
Zhou Y, Ma J, Yi M, Li X, Li X, et al: Nasopharyngeal carcinoma:
Advances in genomics and molecular genetics. Sci China Life Sci.
54:966–975. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wei WI and Sham JS: Nasopharyngeal
carcinoma. Lancet. 365:2041–2054. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xiong W, Zeng ZY, Xia JH, Xia K, Shen SR,
Li XL, Hu DX, Tan C, Xiang JJ, Zhou J, et al: A susceptibility
locus at chromosome 3p21 linked to familial nasopharyngeal
carcinoma. Cancer Res. 64:1972–1974. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zeng Z, Zhou Y, Zhang W, Li X, Xiong W,
Liu H, Fan S, Qian J, Wang L, Li Z, et al: Family-based association
analysis validates chromosome 3p21 as a putative nasopharyngeal
carcinoma susceptibility locus. Genet Med. 8:156–160. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zeng Z, Huang H, Huang L, Sun M, Yan Q,
Song Y, Wei F, Bo H, Gong Z, Zeng Y, et al: Regulation network and
expression profiles of Epstein-Barr virus-encoded microRNAs and
their potential target host genes in nasopharyngeal carcinomas. Sci
China Life Sci. 57:315–326. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yan Q, Zeng Z, Gong Z, Zhang W, Li X, He
B, Song Y, Li Q, Zeng Y, Liao Q, et al: EBV-miR-BART10-3p
facilitates epithelial-mesenchymal transition and promotes
metastasis of nasopharyngeal carcinoma by targeting BTRC.
Oncotarget. 6:41766–41782. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Song Y, Li X, Zeng Z, Li Q, Gong Z, Liao
Q, Li X, Chen P, Xiang B, Zhang W, et al: Epstein-Barr virus
encoded miR-BART11 promotes inflammation-induced carcinogenesis by
targeting FOXP1. Oncotarget. 7:36783–36799. 2016.PubMed/NCBI
|
|
9
|
Zeng Z, Fan S, Zhang X, Li S, Zhou M,
Xiong W, Tan M, Zhang W and Li G: Epstein-Barr virus-encoded small
RNA 1 (EBER-1) could predict good prognosis in nasopharyngeal
carcinoma. Clin Transl Oncol. 18:206–211. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Simon MC: Gotta have GATA. Nat Genet.
11:9–11. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Martin DI, Zon LI, Mutter G and Orkin SH:
Expression of an erythroid transcription factor in megakaryocytic
and mast cell lineages. Nature. 344:444–447. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Molkentin JD: The zinc finger-containing
transcription factors GATA-4, −5, and −6. Ubiquitously expressed
regulators of tissue-specific gene expression. J Biol Chem.
275:38949–38952. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ayanbule F, Belaguli NS and Berger DH:
GATA factors in gastrointestinal malignancy. World J Surg.
35:1757–1765. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
White RA, Dowler LL, Pasztor LM, Gatson
LL, Adkison LR, Angeloni SV and Wilson DB: Assignment of the
transcription factor GATA4 gene to human chromosome 8 and mouse
chromosome 14: Gata4 is a candidate gene for Ds (disorganization).
Genomics. 27:20–26. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Engels M, Span PN, Mitchell RT, Heuvel
JJTM, Marijnissen-van Zanten MA, van Herwaarden AE, Hulsbergen-van
de Kaa CA, Oosterwijk E, Stikkelbroeck NM, Smith LB, et al: GATA
transcription factors in testicular adrenal rest tumours. Endocr
Connect. 6:866–875. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang WY, Heng HH and Liew CC: Assignment
of the human GATA4 gene to 8p23.1-->p22 using fluorescence in
situ hybridization analysis. Cytogenet Cell Genet. 72:217–218.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Perrino C and Rockman HA: GATA4 and the
two sides of gene expression reprogramming. Circ Res. 98:715–716.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McCulley DJ and Black BL: Curr Top Dev
Biol. 100:253–277. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Suzuki YJ: Cell signaling pathways for the
regulation of GATA4 transcription factor: Implications for cell
growth and apoptosis. Cell Signal. 23:1094–1099. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Färkkilä A, Andersson N, Bützow R, Leminen
A, Heikinheimo M, Anttonen M and Unkila-Kallio L: HER2 and GATA4
are new prognostic factors for early-stage ovarian granulosa cell
tumor-a long-term follow-up study. Cancer Med. 3:526–536. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lu H, Huang S, Zhang X, Wang D, Zhang X,
Yuan X, Zhang Q and Huang Z: DNA methylation analysis of SFRP2,
GATA4/5, NDRG4 and VIM for the detection of colorectal cancer in
fecal DNA. Oncol Lett. 8:1751–1756. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takagi K, Moriguchi T, Miki Y, Nakamura Y,
Watanabe M, Ishida T, Yamamoto M, Sasano H and Suzuki T: GATA4
immunolocalization in breast carcinoma as a potent prognostic
predictor. Cancer Sci. 105:600–607. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chia NY, Deng N, Das K, Huang D, Hu L, Zhu
Y, Lim KH, Lee MH, Wu J, Sam XX, et al: Regulatory crosstalk
between lineage-survival oncogenes KLF5, GATA4 and GATA6
cooperatively promotes gastric cancer development. Gut. 64:707–719.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang MH, Wu MZ, Chiou SH, Chen PM, Chang
SY, Liu CJ, Teng SC and Wu KJ: Direct regulation of TWIST by
HIF-1alpha promotes metastasis. Nat Cell Biol. 10:295–305. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gumireddy K, Li A, Gimotty PA,
Klein-Szanto AJ, Showe LC, Katsaros D, Coukos G, Zhang L and Huang
Q: KLF17 is a negative regulator of epithelial-mesenchymal
transition and metastasis in breast cancer. Nat Cell Biol.
11:1297–1304. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
van Tuyn J, Atsma DE, Winter EM, van der
Velde-van Dijke I, Pijnappels DA, Bax NA, Knaän-Shanzer S,
Gittenberger-de Groot AC, Poelmann RE, van der Laarse A, et al:
Epicardial cells of human adults can undergo an
epithelial-to-mesenchymal transition and obtain characteristics of
smooth muscle cells in vitro. Stem Cells. 25:271–278. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Campbell K, Whissell G, Franch-Marro X,
Batlle E and Casanova J: Specific GATA factors act as conserved
inducers of an endodermal-EMT. Dev Cell. 21:1051–1061. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Han Q, Xu X, Li J, Wang J, Bai L, Wang A,
Wang W and Zhang B: GATA4 is highly expressed in childhood acute
lymphoblastic leukemia, promotes cell proliferation and inhibits
apoptosis by activating BCL2 and MDM2. Mol Med Rep. 16:6290–6298.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chan JY, Chow VL, Wong ST and Wei WI:
Surgical salvage for recurrent retropharyngeal lymph node
metastasis in nasopharyngeal carcinoma. Head Neck. 35:1726–1731.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kang M, Zhou P, Wei T, Zhao T, Long J, Li
G, Yan H, Feng G, Liu M, Zhu J and Wang R: A novel N staging system
for NPC based on IMRT and RTOG guidelines for lymph node levels:
Results of a prospective multicentric clinical study. Oncol Lett.
16:308–316. 2018.PubMed/NCBI
|
|
33
|
Kang Y and Massague J:
Epithelial-mesenchymal transitions: Twist in development and
metastasis. Cell. 118:277–279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Medici D, Hay ED and Olsen BR: Snail and
Slug promote epithelial-mesenchymal transition through
beta-catenin-T-cell factor-4-dependent expression of transforming
growth factor-beta3. Mol Biol Cell. 19:4875–4887. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alves CC, Carneiro F, Hoefler H and Becker
KF: Role of the epithelial-mesenchymal transition regulator Slug in
primary human cancers. Front Biosci (Landmark Ed). 14:3035–3050.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kojc N, Zidar N, Gale N, Poljak M, Fujs
Komlos K, Cardesa A, Höfler H and Becker KF: Transcription factors
Snail, Slug, Twist, and SIP1 in spindle cell carcinoma of the head
and neck. Virchows Arch. 454:549–555. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Preca BT, Bajdak K, Mock K, Lehmann W,
Sundararajan V, Bronsert P, Matzge-Ogi A, Orian-Rousseau V,
Brabletz S, Brabletz T, et al: A novel ZEB1/HAS2 positive feedback
loop promotes EMT in breast cancer. Oncotarget. 8:11530–11543.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mahmoud MM, Kim HR, Xing R, Hsiao S,
Mammoto A, Chen J, Serbanovic-Canic J, Feng S, Bowden NP, Maguire
R, et al: Twist1 integrates endothelial responses to flow in
vascular dysfunction and atherosclerosis. Circ Res. 119:450–462.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pan B, Ye Y, Liu H, Zhen J, Zhou H, Li Y,
Qu L, Wu Y, Zeng C and Zhong W: URG11 regulates prostate cancer
cell proliferation, migration, and invasion. Biomed Res Int.
2018:40607282018. View Article : Google Scholar : PubMed/NCBI
|