Introduction
According to a recent epidemiological survey, the
rate of the loss of one or more teeth in individuals is 6.45%
(1). With the aging of the
population in China, the incidence of tooth loss exhibits yearly
increases. Severe alveolar bone loss is commonly observed in
patients with periodontal disease or in those who wear dentures for
an extended period, which may cause atrophy of the alveolar ridge
(2). The significant reduction in
the contact area of the mandibular tissue in patients with severe
atrophy of the alveolar ridge makes denture retention difficult and
the occlusal ability of these patients frequently remains poor
after repair (3). This has a severe
impact on the physical and mental wellbeing of the affected
patients, thereby impairing their quality of life. Therefore,
determining the specific pathophysiological mechanisms underlying
alveolar atrophy has become an issue of prime importance. A recent
study has revealed that the pathophysiological mechanisms
underlying alveolar ridge atrophy are associated with a condition
wherein the osteoclast metabolism is higher than the osteoblast
metabolism (4). Studying alveolar
bone-associated osteoclasts, their osteogenic metabolism and the
underlying regulatory mechanisms using molecular biology techniques
will help in the elucidation of the causes of alveolar ridge
atrophy.
MicroRNAs (miRNAs/miRs) are a class of small
non-coding RNAs of 19–25 nucleotides in length that are capable of
regulating gene expression post-transcriptionally by binding to the
3′-untranslated region of their target mRNAs (5). miRNAs regulate the expression of ~30%
of all human genes (6). Increasing
evidence has indicated that miRNAs have important roles in
physiological development and in pathogenesis (7). miRNA expression profiles are useful
tools to study the molecular mechanisms of diseases (8). For instance, miRNA expression profile
analysis has indicated that miR-148a-3p is markedly upregulated
during osteoclastogenesis of human CD14+ peripheral
blood mononuclear cells (PBMCs). Overexpression of Homo
sapiens (hsa)-miR-148a-3p in CD14+ PBMCs facilitated
osteoclastogenesis, whereas suppression of hsa-miR-148a-3p
expression inhibited it (9). Several
miRNAs that participate in osteoclast formation, differentiation,
apoptosis and resorption have been identified (9–11). For
instance, miR-148b directly targets the gene encoding noggin, which
is involved in the regulation of bone morphogenetic protein
2-induced osteogenic differentiation and bone formation (12). Photo-activated miR-148b nanoparticle
conjugates improve the closure of critical-sized calvarial defects
in mice (13). Yoshitaka et
al (14) have indicated that
miR-337-5p was significantly downregulated in patients with
chondrosarcoma as well as in chondrosarcoma cell lines. Although
miRNAs appear to be critical regulators of bone-associated
osteoclast metabolism and osteogenesis, few studies have examined
their roles in alveolar ridge atrophy.
Recently, stem cell-based tissue engineering has
been successful in reconstructing several tissue types, including
dental tissues (15). Adult bone
marrow mesenchymal stem cells (MMSCs) are not only easily obtained
and expanded in culture but also have the ability of multilineage
differentiation, i.e., these cells are able to differentiate to
form a variety of tissue types, including bone, cartilage, tendon
and adipose tissues (16). MMSCs
from orofacial bone (OMMSCs) are the ideal seeding cells for the
reconstruction of the alveolar ridge tissue (17). It is thus essential to explore the
role of miRNAs in OMMSCs.
The present study aimed to identify miRNAs that may
be associated with the mechanisms underlying alveolar ridge
atrophy. Hence, differentially expressed miRNAs between alveolar
ridge atrophy and normal tissue samples were identified by miRNA
microarray analysis. Various deregulated miRNAs in alveolar ridge
atrophy were then confirmed by reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis. Subsequent in
vitro functional assays indicated that miR-148b-3p, miR-337-5p
and miR-423-5p promote the proliferation of OMMSCs and inhibit
their apoptosis.
Materials and methods
Sample collection
Alveolar ridge atrophy tissue samples were obtained
from elderly male patients (aged >70 years), who required repair
of the alveolar ridge. Normal alveolar ridge tissue samples were
obtained from adult male patients (age, 25–35 years) who required
removal of wisdom teeth. All participants were recruited between
January 2013 and December 2013 from the Chinese PLA General
Hospital (Beijing, China). In total, 21 alveolar ridge atrophy
tissue samples from 21 different patients and 21 alveolar ridge
normal tissue samples from 21 different healthy volunteers were
obtained. All tissue samples were immediately snap-frozen in liquid
nitrogen and stored at −80°C until total RNA extraction. A total of
3 alveolar ridge atrophy tissue samples and 3 alveolar ridge normal
tissue samples were randomly selected for the miRNA microarray
analysis. The present study was performed according to principles
of the Helsinki Declaration from 1975 and its revision from 1983.
The use of the aforementioned samples in the present study was
approved by the Committees for Ethical Review of Research Involving
Human Subjects of the Chinese PLA General Hospital (Beijing,
China). Written informed consent was received from each tissue
donor.
RNA extraction
Total RNA was extracted from the frozen tissue using
the TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) according to the manufacturer's protocol. The
total RNA in the sample was quantified using a NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc.) and the RNA
integrity was evaluated by agarose gel electrophoresis.
miRNA microarray assay
The miRNA microarray assays were performed using an
Affymetrix miRNA array system (Affymetrix; Thermo Fisher
Scientific, Inc.) following the manufacturer's protocol in order to
screen for differential expression profiles of miRNA in the
alveolar ridge atrophy tissue samples vs. the normal tissue
samples. In brief, the total RNA was subjected to Poly A-tailing
and then labeled with biotin using a FlashTag Biotin HSR RNA
Labeling Kit (Affymetrix; Thermo Fisher Scientific, Inc.).
Hybridization was then performed using the Affymetrix GeneChip
miRNA 4.0 (Affymetrix; Thermo Fisher Scientific, Inc.). The
hybridization signals were produced using the Affymetrix Command
Console Software 3.0 (Affymetrix; Thermo Fisher Scientific, Inc.).
The raw data were normalized using the Robust Multichip Average
algorithm. The differentially expressed miRNAs were identified by
screening for fold changes in expression and by the P-value
(fold-change, >1.5; P<0.05). To evaluate the correlations
between the results of the individual experiments, a hierarchical
clustering analysis was performed using Cluster 3.0 software (open
source, http://bonsai.hgc.jp/~mdehoon/software/cluster/software.htm).
RT-qPCR
First, the differential expression of 20 selected
miRNAs in the alveolar ridge atrophy vs. normal tissue samples
identified in the microarray assay was validated by RT-qPCR. The
relative expression levels of three miRNAs (miR-148b-3p, miR-337-5p
and miR-423-5p) in 18 pairs of alveolar ridge atrophy and normal
tissue samples, excluding the three pairs used for the microarray
assay, were measured by RT-qPCR. RT was performed using the
TaqMan® MicroRNA Reverse Transcription Kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Real-time PCR was performed using SYBR
Green (Takara Bio, Inc., Dalian, China) and a 7500 real-time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). A 20
µl total volume of the qPCR mixture was prepared with 5 µl
20×-dilution RT products, 500 nM of forward and reverse primers and
2× SYBR Green qPCR SuperMix (Invitrogen; Thermo Fisher Scientific,
Inc.). Each sample was tested in triplicate. The thermocycling
conditions were as follows: 50°C for 2 min, 95°C for 2 min,
followed by 45 cycles (at 95°C for 5 sec and 55°C for 20 sec). The
primers were synthesized by Takara Bio, Inc. and their sequences
are listed in Table I. U6 RNA served
as the endogenous control. Relative miRNA expression was calculated
using the 2−ΔΔCq method (18). All experiments were performed in
triplicate.
 | Table I.Sequences of primers used for
polymerase chain reaction, as well as miRNA mimics and
inhibitors. |
Table I.
Sequences of primers used for
polymerase chain reaction, as well as miRNA mimics and
inhibitors.
| Name | Sequence (5′-3′) |
|---|
| miRNA-RT |
GTTGGCTCTGGTGCAGGGTCCGAGGTATTCGCACCAGAGCCAACCTCAGG |
| miR-196a-3p-F |
GGACGGCAACAAGAAACTG |
| miR-188-3p-F |
TTTCTCCCACATGCAGGG |
| miR-335-5p-F |
GAGGTCAAGAGCAATAACGAA |
| miR-148b-3p-F |
GGGTCAGTGCATCACAGAA |
| miR-302a-3p-F |
CGGTAAGTGCTTCCATGTTT |
| miR-29b-3p-F |
GCAGTAGCACCATTTGAAATC |
| miR-183-3p-F |
AGGAGTGAATTACCGAAGGG |
| miR-183-5p-F |
GAGGTATGGCACTGGTAGAA |
| miR-15a-5p-F |
TTTGGTAGCAGCACATAATGG |
| miR-337-5p-F |
CCCGAACGGCTTCATACA |
| miR-144-5p-F |
CGGGCGGATATCATCATATAC |
| miR-133a-3p-F |
TCCTTTGGTCCCCTTCAAC |
| miR-144-3p-F |
GGCGGGTACAGTATAGATGA |
| miR-448-F |
CGGGTTGCATATGTAGGATG |
| miR-515-3p-F |
GTGGAGTGCCTTCTTTTGG |
| miR-489-3p-F |
GCGGGTGACATCACATATAC |
| miR-22-3p-F |
GGAAAGCTGCCAGTTGAAG |
| miR-590-3p-F |
CGGGGGGCTAATTTTATGTATAA |
| miR-496-F |
GTGCTGAGTATTACATGGCC |
| miR-18b-5p-F |
GTGTAAGGTGCATCTAGTGC |
| miR-423-5p-F |
GAGGGGCAGAGAGCGA |
| U6_F |
TTCCTCCGCAAGGATGACACGC |
| miR-148b-3p
mimic |
UCAGUGCAUCACAGAACUUUGU |
| miR-148b-3p
inhibitor |
ACAAAGUUCUGUGAUGCACACUGA |
| miR-337-5p
mimic |
GAACGGCUUCAUACAGGAGUU |
| miR-337-5p
inhibitor |
AACUCCUGUAUGAAGCCGUUC |
| miR-423-5p
mimic |
UGAGGGGCAGAGAGCGAGACUUU |
| miR-423-5p
inhibitor |
AAAGUCUCGCUCUCUCUGCCCCUCA |
| NC mimic |
UCACAACCUCCUAGAAAGAGUAGA |
| NC inhibitor |
UCUACUUCUUUCUAGGAGGUUGUGA |
Cell culture and transfection
Human OMMSCs were obtained from the Cell Bank of the
Chinese Academy of Sciences (Shanghai, China). The OMMSCs were
cultured in α-minimal essential medium (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.) and 1% penicillin-streptomycin
(Invitrogen; Thermo Fisher Scientific, Inc.) in a humidified
incubator with a 5% CO2 atmosphere at 37°C. The human
OMMSCs were grown in attached culture. The miRNA mimics, inhibitors
(antisense), and negative controls (NC) were purchased from RiboBio
(Guangzhou, China) and their sequences are listed in Table I. The OMMSCs were transfected using
Lipofectamine RNAiMAX (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. At 48 h after
transfection, the cell proliferation and apoptosis assays were
performed.
Cell proliferation assay
Cell proliferation was assessed using a Cell
Counting Kit (CCK)-8 assay kit (Beyotime Institute of
Biotechnology, Shanghai, China) following the manufacturer's
protocol. Following culture for 24 h, OMMSCs were placed in 96-well
plates at a density of 5×103 cells per well and
transfected with the miRNA mimics, inhibitors and NCs,
respectively, with each condition performed in triplicate. After
culturing for 0, 1, 2 or 3 days, 10 µl CCK-8 reagent was added to
each well followed by gentle agitation. The cells were then
cultured at 37°C in a 5% CO2 atmosphere for 4 h.
Subsequently, the absorbance was measured at 450 nm using an MK3
microplate reader (Thermo Fisher Scientific, Inc.).
Apoptosis assay using flow
cytometry
OMMSCs transfected with miRNA mimics, inhibitors and
NCs, respectively, were used in this experiment. Cell pellets
(5×103 cells) were re-suspended in 500 µl binding
buffer. Subsequently, 1.25 µl annexin V-fluorescein isothiocyanate
and 10 µl propidium iodide were added to the cell suspension (0.5
ml) and the cells were cultured in the dark for 15 min at 25°C.
Cell apoptosis was then assessed by flow cytometry (BD Biosciences,
San Jose, CA, USA). Each experiment was performed in
triplicate.
Statistical analysis
Statistical analysis was performed using SPSS 19.0
software (IBM Corp., Armonk, NY, USA). Values are expressed as the
mean ± standard deviation. A moderated t-test was performed to
identify miRNAs that were differentially expressed between the
alveolar ridge atrophy and normal tissue samples. The statistically
significant differences of results obtained by RT-qPCR analysis and
the cell proliferation and apoptosis assays, were assessed by an
independent-samples t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression profiling of microRNA in
tissue samples
To identify whether miRNAs have a role in alveolar
ridge atrophy, a miRNA microarray analysis was performed to examine
the global miRNA expression in alveolar ridge atrophy and normal
tissue samples. To identify differentially expressed miRNAs,
moderated t-tests were performed, and 52 miRNAs whose expression
levels were considerably different between the alveolar ridge
atrophy and normal tissue samples were retrieved. A map of
unsupervised hierarchical clustering analysis of the differentially
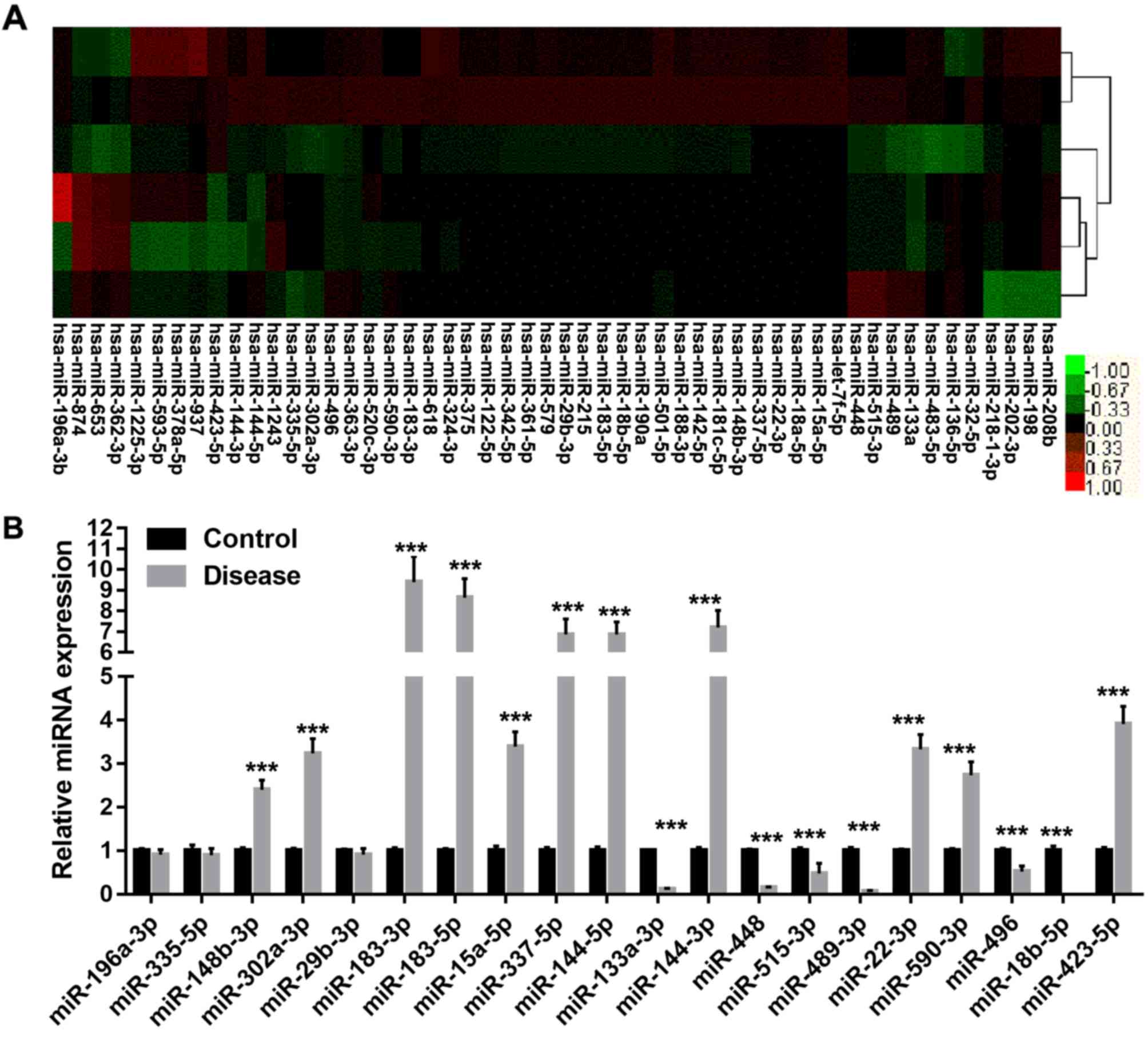
expressed miRNAs is presented in Fig.
1A. The results indicated that various miRNAs may be involved
in alveolar ridge atrophy. To validate the microarray data, RT-qPCR
was performed to detect the differential expression of 20 selected
miRNAs (fold change, >2) between the alveolar ridge atrophy and
normal tissue samples that were used in the microarray assay
(Fig. 1B). The expression of 11
miRNAs (miR-148b-3p, miR-302a-3p, miR-183-3p/5p, miR-15a-5p,
miR-337-5p, miR-144-3p/5p, miR-22-3p, miR-590-3p and miR-423-5p)
was significantly upregulated (fold change, >2) in the alveolar
ridge atrophy tissues samples compared with that in the normal
alveolar ridge tissue samples (P<0.05), while the expression of
five miRNAs (miR-133a-3p, miR-448, miR-515-3p, miR-489-3p and
miR-18b-5p) was significantly downregulated (fold change, <1/2;
P<0.05).
Relative expression of miR-148b-3p,
miR-337-5p and miR-423-5p in tissue samples
Studies have indicated that certain miRNAs,
including miR-148b-3p, miR-337-5p and miR-423-5p, participate in
bone metabolism. In the present study, the expression levels of
these 3 miRNAs were significantly upregulated in the alveolar ridge
atrophy tissue samples compared with those in the normal alveolar
ridge tissue samples (P<0.05). Thus, miR-148b-3p, miR-337-5p and
miR-423-5p were selected for further experiments, including the
determination of their relative expression levels in alveolar ridge
atrophy or normal tissue samples and cellular physiological
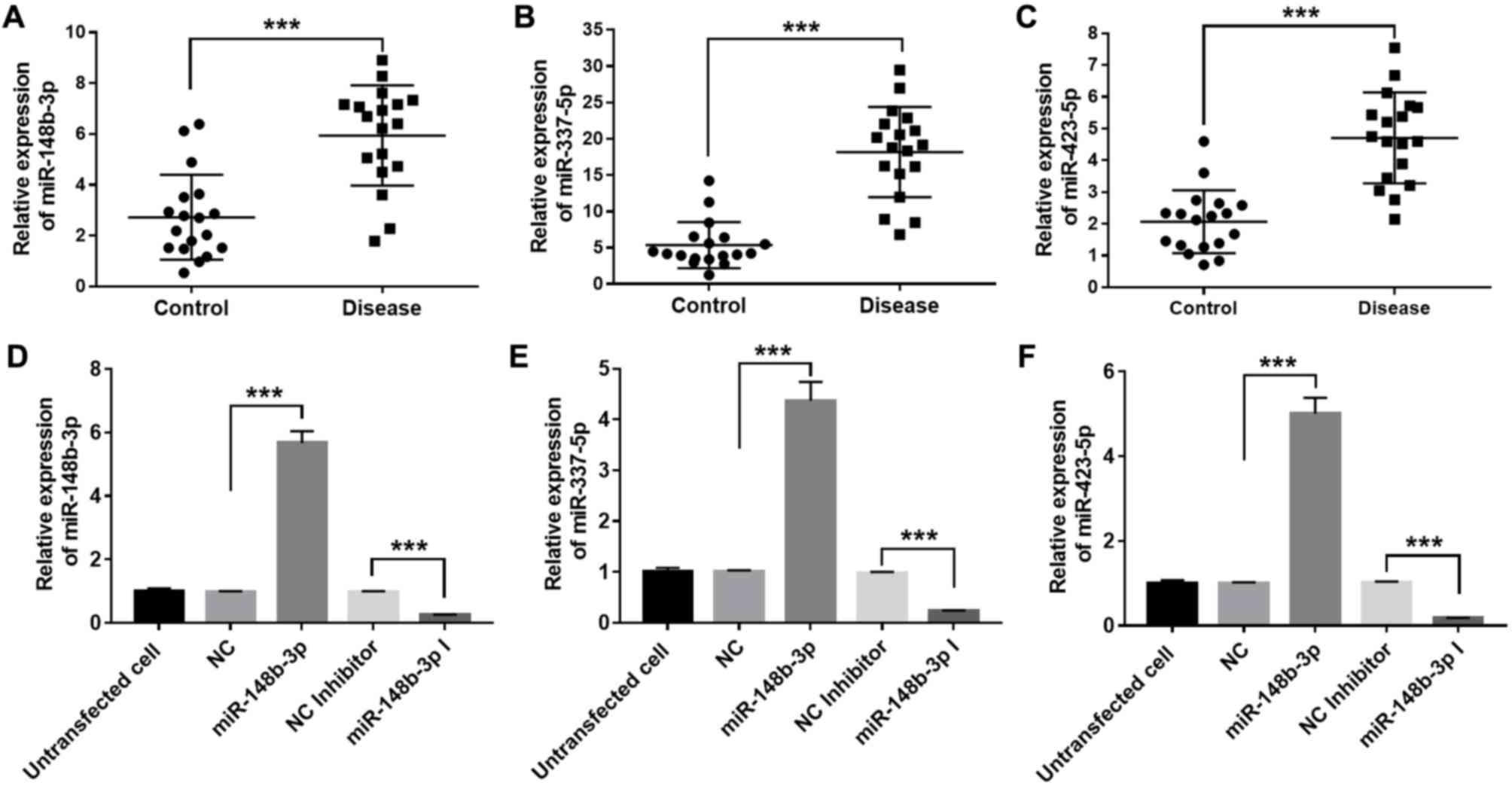
function assays. To confirm the expression levels of miR-148b-3p,
miR-337-5p and miR-423-5p in alveolar ridge atrophy and normal
alveolar ridge tissue samples, their relative expression levels in
18 pairs of alveolar ridge atrophy and normal tissue samples,
excluding the three pairs used in the microarray assay, were
measured using RT-qPCR. The results indicated that the relative
expression levels of miR-148b-3p, miR-337-5p and miR-423-5p in the
alveolar ridge atrophy tissue samples were higher than those in
alveolar ridge normal tissue samples (P<0.001), which is
consistent with the microarray data (Fig. 2A-C). To further assess the biological
functions of miR-148b-3p, miR-337-5p and miR-423-5p in the OMMSCs,
these miRNAs were ectopically overexpressed by transient
transfection with miR-148b-3p, miR-337-5p and miR-423-5p mimics,
respectively, and furthermore, knockdown of these miRNAs was
achieved via transient transfection with miR-148b-3p, miR-337-5p
and miR-423-5p inhibitors, respectively. It was experimentally
verified that the expression levels of miR-148b-3p, miR-337-5p and
miR-423-5p were significantly enhanced after transfection with
their respective mimics, while their expression levels were
significantly decreased after transfection with their respective
inhibitors (P<0.001; Fig.
2D-F).
miR-148b-3p, miR-337-5p and miR-423-5p
promote OMMSC proliferation
The proliferation of OMMSCs was determined by the
CCK-8 assay (Fig. 3). On the third
day after transfection with the mimics, the proliferative ability
of the OMMSCs was markedly enhanced compared with that in the
NC-transfected group (P<0.05). By contrast, on the third day
following transfection with the inhibitors, the proliferative
ability of the OMMSCs was markedly reduced compared with that in
the NC-transfected group (P<0.01). The results indicated that
miR-148b-3p, miR-337-5p and miR-423-5p promote the proliferation of
OMMSCs.
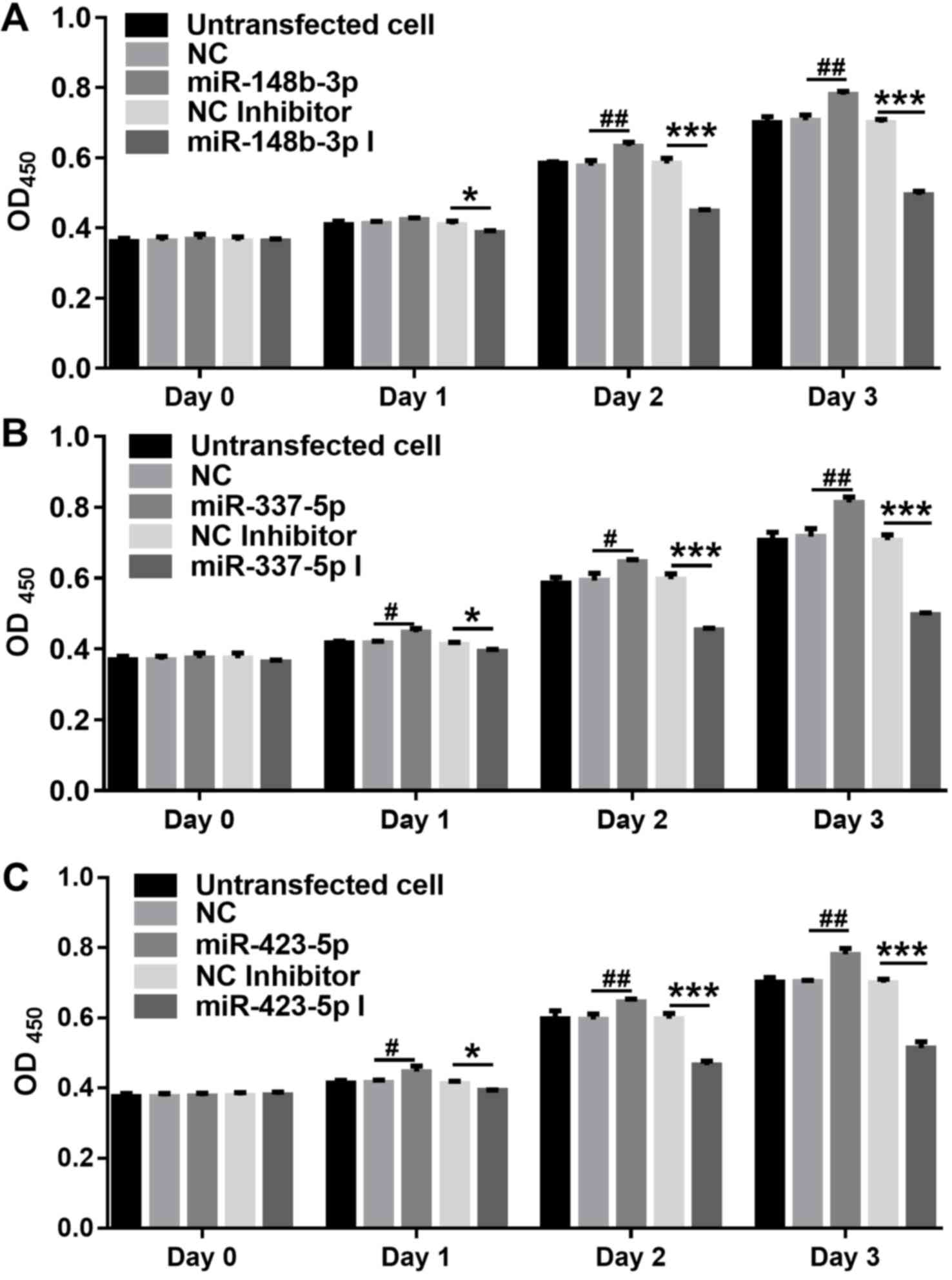 | Figure 3.Proliferation of OMMSCs analyzed by
the Cell Counting Kit-8 assay. (A) Proliferation of the OMMSCs
transfected with NC, miR-148b-3p mimics, NC inhibitor or
miR-148b-3p inhibitor. (B) Proliferation of the OMMSCs transfected
with NC, miR-337-5p mimics, NC inhibitor or miR-337-5p inhibitor.
(C) Proliferation of the OMMSCs transfected with NC, miR-423-5p
mimics, NC inhibitor or miR-423-5p inhibitor. Values are expressed
as the mean ± standard deviation. *P<0.05 and ***P<0.001 vs.
NC inhibitor; #P<0.05 and ##P<0.01 vs.
NC. OMMSCs, bone marrow mesenchymal stem cells from orofacial bone;
NC, negative control; miR, microRNA; OD450, optical
density at 450 nm; miR-148b-3p I, miR-148b-3p inhibitor. |
miR-148b-3p, miR-337-5p and miR-423-5p
inhibit OMMSC apoptosis
The effect of miR-148b-3p, miR-337-5p and miR-423-5p
overexpression on the apoptosis in OMMSCs was assessed by flow
cytometry. As presented in Figs.
4–6, the results indicated that
miR-148b-3p, miR-337-5p or miR-423-5p overexpression significantly
inhibited apoptosis in OMMSCs (P<0.05). By contrast, knockdown
of miR-148b-3p, miR-337-5p or miR-423-5p expression promoted the
apoptosis in OMMSCs (P<0.001). These results suggested that
miR-148b-3p, miR-337-5p and miR-423-5p promote the proliferation of
OMMSC and inhibit their apoptosis.
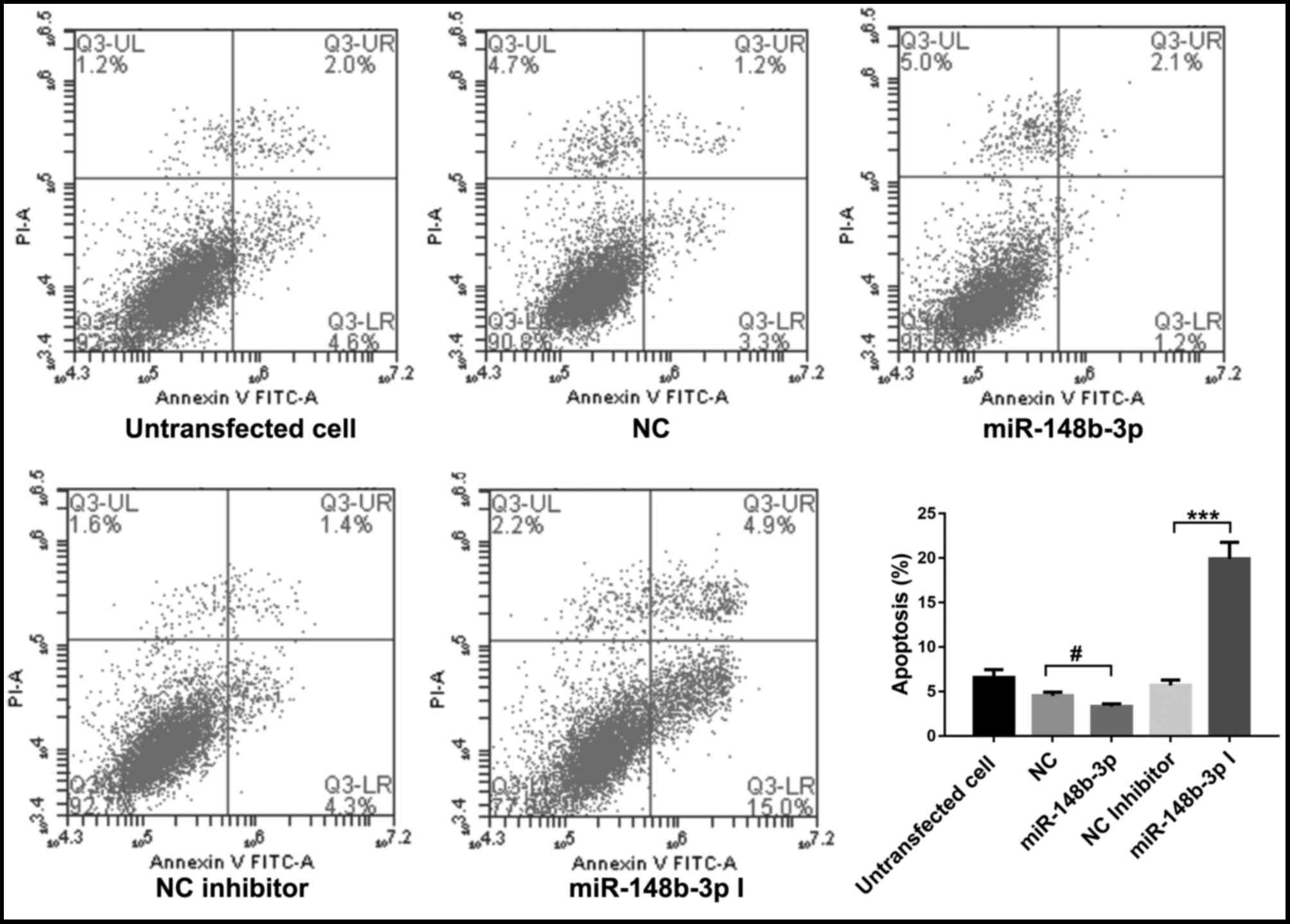 | Figure 4.Analysis of the apoptosis of bone
marrow mesenchymal stem cells from orofacial bone transfected with
NC, miR-148b-3p mimics, NC inhibitor or miR-148b-3p inhibitor.
Values are expressed as the mean ± standard deviation.
***P<0.001 vs. NC inhibitor; #P<0.05 vs. NC. PI,
propidium iodide; FITC, fluorescein isothiocyanate; NC, negative
control; miR, microRNA; miR-148b-3p I, miR-148b-3p inhibitor; Q,
quadrant; UL, upper left; LR, lower right. |
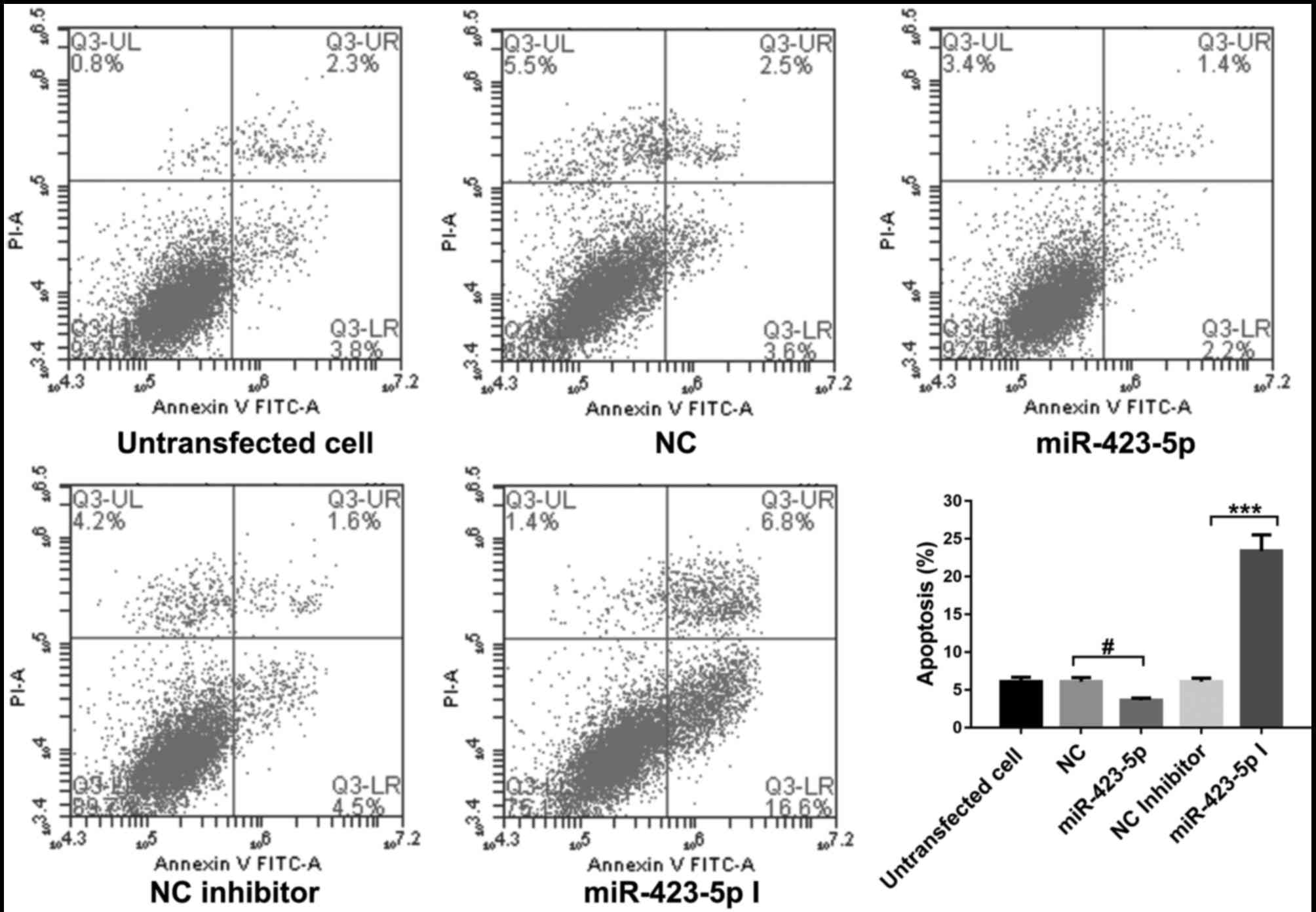 | Figure 6.Analysis of the apoptosis of the bone
marrow mesenchymal stem cells from orofacial bone transfected with
NC, miR-423-5p mimics, NC inhibitor or miR-423-5p inhibitor. Values
are expressed as the mean ± standard deviation. ***P<0.001 vs.
NC inhibitor; #P<0.05 vs. NC. PI, propidium iodide;
FITC, fluorescein isothiocyanate; NC, negative control; miR,
microRNA; miR-423-5p I, miR-423-5p inhibitor; Q, quadrant; UL,
upper left; LR, lower right. |
Discussion
Patients with periodontal disease or individuals who
wear dentures for an extended period are often observed to exhibit
atrophy of the alveolar ridge. Bone regeneration contributes to
reestablishing the proper contour of the alveolar ridge to allow
for implant placement (19). Studies
have indicated that miRNAs have important roles in the occurrence
and development of various types of disease and may function as a
novel targets for clinical therapy (20). In the present study, a miRNA
microarray analysis was performed to identify miRNAs that are
differentially expressed in alveolar ridge atrophy and normal
tissue samples. A total of 52 miRNAs that were differentially
expressed between the two sample groups were retrieved. The
expression of 20 selected differentially expressed miRNAs was
analyzed by RT-qPCR; 11 were identified to be upregulated and 5
were downregulated. Among these 16 miRNAs, only 3 (miR-148b-3p,
miR-337-5p and miR-423-5p) have been indicated to be involved in
bone metabolism by previous studies (13,14,21).
Schoolmeesters et al (20) have revealed a critical role of
miR-148b levels in inducing differentiation in the absence of
external cues and stimulating osteogenesis in over-propagated human
mesenchymal stem cells. Song et al (22) have indicated that miR-148b-laden
titanium implants promote osteogenic differentiation of rat bone
marrow mesenchymal stem cells. Qureshi et al (23) reported that miR-148b-nanoparticle
conjugates promote light-mediated osteogenesis of human adipose
stromal stem cells. miR-337-5p targeting the 3¢-untranslated region
of the gene encoding nucleophosmin 1 has been associated with
adverse outcomes in patients with acute myeloid leukemia (24). Huang et al (25) have indicated that miR-337
overexpression promotes the proliferation of chondrocytes and
inhibits their apoptosis. miR-337 is associated with chondrogenesis
by regulating the expression of transforming growth factor β
receptor 2 (26). miR-423-5p is
downregulated in osteosarcoma cells expressing CD117 and STRO-1
(21). The miR-423-5p levels were
considerably elevated and correlated with the prognosis of patients
with hormone-induced non-traumatic osteonecrosis of the femoral
head (27). These studies indicate
that miR-148b-3p, miR-337-5p and miR-423-5p participate in
osteoclast metabolism. In the present study, the expression levels
of miR-148b-3p, miR-337-5p and miR-423-5p were further validated in
18 pairs of alveolar ridge atrophy and normal tissue samples,
indicating that the levels were consistent with the microarray and
RT-qPCR results obtained with a small number of samples.
OMMSCs are multipotent non-hematopoietic adult stem
cells that are highly self-renewable and have a multidirectional
differentiation potential. The characteristics of OMMSCs, including
low immunogenicity (28),
hematopoietic ability (29), as well
as and immune reconstitution and immunomodulation (30), render them ideal seed cells for
transplantation, particularly in bone tissue engineering. In the
present study, the effects of miR-148b-3p, miR-337-5p and
miR-423-5p on OMMSCs were further investigated by transient
transfection with their respective mimics and inhibitors. The
results indicated that the proliferative ability of OMMSCs was
promoted after transfection with the miR-148b-3p, miR-337-5p and
miR-423-5p mimics, and repressed after transfection with the
miR-148b-3p, miR-337-5p and miR-423-5p inhibitors, compared with
that in the NC-transfected groups. The results of the apoptosis
assay indicated that miR-148b-3p, miR-337-5p and miR-423-5p inhibit
OMMSC apoptosis.
Of note, the present study had certain limitations.
First, the association between miR-148b-3p, miR-337-5p and
miR-423-5p and their target genes was not investigated.
Furthermore, the specific mechanisms underlying miR-148b-3p-,
miR-337-5p- and miR-423-5p-mediated inhibition of OMMSC apoptosis
were not investigated. Additionally, only 3 tissue samples were
utilized for each group in the microarray assay. Furthermore, the
donors of the normal and atrophic samples were not age-matched.
In conclusion, the present study identified 11
miRNAs that were differentially expressed between alveolar ridge
atrophy and normal tissue samples, of which miR-148b-3p-3p,
miR-337-5p and miR-423-5p were previously reported to be associated
with bone metabolism. Loss- and gain-of-function assays indicated
that miR-148b-3p, miR-337-5p and miR-423-5p promote the
proliferation of OMMSC and inhibit their apoptosis. These results
support the hypothesis that certain miRNAs are involved in the
pathogenesis of alveolar ridge atrophy and provide a novel
perspective for understanding the mechanisms underlying this
condition.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
National Natural Science Foundation of China (grant no.
81371183).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors collected the samples. SY, JG and LZ
performed the cell culture, flow cytometric analysis and CCK-8
assay. HX performed the RNA extraction and RT-qPCR. SY designed the
study and CD performed the statistical analyses. SY wrote the
manuscript and JG assisted with drafting the manuscript. All
authors have read and approved the final manuscript.
Ethical approval and consent to
participate
The use of tissue samples in the present study was
approved by the Committees for Ethical Review of Research Involving
Human Subjects of the Chinese PLA General Hospital (Beijing,
China). Written informed consent was received from each tissue
donor.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Silva-Junior MF, Batista MJ and de Sousa
MDLR: Incidence of tooth loss in adults: A 4-year population-based
prospective cohort study. Int J Dent. 2017:60747032017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gül EB, Yolcu Ü, Atala MH and Eşer B:
Simple treatment plan for severe atrophic alveolar ridges: 2 case
reports. Int J Dent Sci Res. 3:96–101. 2015.
|
|
3
|
Liu ZP and Stomatology DO: Observation on
the effect of lingual flange base method for repairing alveolar
ridge severe atrophic mandibular dentition lack of 36 cases. J
Qiqihar Uni Med. 3500–3501. 2015.(In Chinese).
|
|
4
|
Ji X, Chen X and Yu X: MicroRNAs in
osteoclastogenesis and function: Potential therapeutic targets for
osteoporosis. Int J Mol Sci. 17:3492016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Erson AE and Petty EM: MicroRNAs in
development and disease. Clin Genet. 74:296–306. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chang TC and Mendell JT: microRNAs in
vertebrate physiology and human disease. Annu Rev Genomics Hum
Genet. 8:215–239. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng P, Chen C, He HB, Hu R, Zhou HD, Xie
H, Zhu W, Dai RC, Wu XP, Liao EY and Luo XH: miR-148a regulates
osteoclastogenesis by targeting V-maf musculoaponeurotic
fibrosarcoma oncogene homolog B. J Bone Miner Res. 28:1180–1190.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu J, Dang L, Li D, Liang C, He X, Wu H,
Qian A, Yang Z, Au DW, Chiang MW, et al: A delivery system
specifically approaching bone resorption surfaces to facilitate
therapeutic modulation of microRNAs in osteoclasts. Biomaterials.
52:148–160. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ke K, Sul OJ, Rajasekaran M and Choi HS:
MicroRNA-183 increases osteoclastogenesis by repressing heme
oxygenase-1. Bone. 81:237–246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li KC, Lo SC, Sung LY, Liao YH, Chang YH
and Hu YC: Improved calvarial bone repair by hASCs engineered with
Cre/loxP-based baculovirus conferring prolonged BMP-2 and MiR-148b
co-expression. J Tissue Eng Regen Med. 11:3068–3077. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qureshi AT, Doyle A, Chen C, Coulon D,
Dasa V, Del Piero F, Levi B, Monroe WT, Gimble JM and Hayes DJ:
Photoactivated miR-148b-nanoparticle conjugates improve closure of
critical size mouse calvarial defects. Acta Biomater. 12:166–173.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoshitaka T, Kawai A, Miyaki S, Numoto K,
Kikuta K, Ozaki T, Lotz M and Asahara H: Analysis of microRNAs
expressions in chondrosarcoma. J Orthop Res. 31:1992–1998. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Iwata T, Washio K, Yoshida T, Ishikawa I,
Ando T, Yamato M and Okano T: Cell sheet engineering and its
application for periodontal regeneration. J Tissue Eng Regen Med.
9:343–356. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Orbay H, Tobita M and Mizuno H:
Mesenchymal stem cells isolated from adipose and other tissues:
Basic biological properties and clinical applications. Stem Cells
Int. 2012:4617182012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Niibe K, Zhang M, Nakazawa K, Morikawa S,
Nakagawa T, Matsuzaki Y and Egusa H: The potential of enriched
mesenchymal stem cells with neural crest cell phenotypes as a cell
source for regenerative dentistry. Jpn Dent Sci Rev. 53:25–33.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maiorana C, Beretta M, Rancitelli D,
Grossi GB, Cicciù M and Herford AS: Histological features and
biocompatibility of bone and soft tissue substitutes in the
atrophic alveolar ridge reconstruction. Case Rep Dent.
2016:36086022016.PubMed/NCBI
|
|
20
|
Schoolmeesters A, Eklund T, Leake D,
Vermeulen A, Smith Q, Force Aldred S and Fedorov Y: Functional
profiling reveals critical role for miRNA in differentiation of
human mesenchymal stem cells. PLoS One. 4:e56052009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao F, Lv J, Gan H, Li Y, Wang R, Zhang
H, Wu Q and Chen Y: MiRNA profile of osteosarcoma with CD117 and
stro-1 expression: miR-1247 functions as an onco-miRNA by targeting
MAP3K9. Int J Clin Exp Pathol. 8:1451–1458. 2015.PubMed/NCBI
|
|
22
|
Song W, Wu K, Yan J, Zhang Y and Zhao L:
MiR-148b laden titanium implant promoting osteogenic
differentiation of rat bone marrow mesenchymal stem cells. RSC Adv.
3:11292–11300. 2013. View Article : Google Scholar
|
|
23
|
Qureshi AT, Monroe WT, Dasa V, Gimble JM
and Hayes DJ: miR-148b-nanoparticle conjugates for light mediated
osteogenesis of human adipose stromal/stem cells. Biomaterials.
34:7799–7810. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng CK, Kwan TK, Cheung CY, Ng K, Liang
P, Cheng SH, Chan NP, Ip RK, Wong RS, Lee V, et al: A polymorphism
in the 3′-untranslated region of the NPM1 gene causes illegitimate
regulation by microRNA-337-5p and correlates with adverse outcome
in acute myeloid leukemia. Haematologica. 98:913–917. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang Z, Zhang N, Ma W, Dai X and Liu J:
MiR-337-3p promotes chondrocytes proliferation and inhibits
apoptosis by regulating PTEN/AKT axis in osteoarthritis. Biomed
Pharmacother. 95:1194–1200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhong N, Sun J, Min Z, Zhao W, Zhang R,
Wang W, Tian J, Tian L, Ma J, Li D, et al: MicroRNA-337 is
associated with chondrogenesis through regulating TGFBR2
expression. Osteoarthritis Cartilage. 20:593–602. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei B and Wei W: Identification of
aberrantly expressed of serum microRNAs in patients with
hormone-induced non-traumatic osteonecrosis of the femoral head.
Biomed Pharmacother. 75:191–195. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu J, Wang Q, Fu X, Wu X, Gu C, Bi J, Xie
F, Kang N, Liu X, Yan L, et al: Influence of immunogenicity of
allogeneic bone marrow mesenchymal stem cells on bone tissue
engineering. Cell Transplant. 25:229–242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chiba Y, Kuroda S, Osanai T, Shichinohe H,
Houkin K and Iwasaki Y: Impact of ageing on biological features of
bone marrow stromal cells (BMSC) in cell transplantation therapy
for CNS disorders: Functional enhancement by granulocyte-colony
stimulating factor (G-CSF). Neuropathology. 32:139–148. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang L, Su XS, Ye JS, Wang YY, Guan Z and
Yin YF: Bone marrow mesenchymal stem cells suppress metastatic
tumor development in mouse by modulating immune system. Stem Cell
Res Ther. 6:452015. View Article : Google Scholar : PubMed/NCBI
|