Introduction
As mesenchymal stem cells have the ability for
differentiation into osteoclasts, they have become important
multifunctional seeds for bone tissue engineering research
(1). When the ability of this
directional differentiation is strengthened or suppressed, it
disrupts the balance of bone-related metabolism in the body, which
is a main cause of osteoporosis (OP) development (2). However, the search for effective
strategies for preventing OP remains an active and challenging area
of research.
OP is a metabolic bone disease characterized by a
reduction of bone mass, mainly due to the augmentation of bone
absorption (3). The occurrence of OP
is directly associated with the activity of osteoblasts and
osteoclasts, particularly the process of osteoclastogenesis. OP is
associated with an increased risk of fracture and mortality,
representing a classic aging-associated disease (4–6); the
lifetime risk of an osteoporotic fracture in men over the age of 50
is 30% (7). Vitamin D3 is the
traditional treatment for OP, but these effects are insufficient to
completely explain the anti-OP mechanism (8). Thus, novel anti-OP drugs are required,
warranting further research to identify novel drug targets. Recent
studies (9) have demonstrated a
close correlation between adiponectin and bone metabolism,
suggesting a possible candidate for the OP pathogenic mechanism and
treatment target.
Adiponectin is mainly secreted by adipocytes and
serves an important role in preventing obesity, diabetes and
atherosclerosis (10–12). Adiponectin was demonstrated to
inhibit receptor activator of nuclear factor (NF)-κB ligand
(RANKL)-induced osteoclast differentiation, and showed a
significant negative correlation with bone mineral density (BMD)
(13,14). RANK can be combined with its ligand
RANKL or other members of the tumor necrosis factor (TNF) ligand
family, and osteoclast precursor cells can continuously generate
RANK. Once the content of RANK in the body is reduced, osteoclast
progenitor cell proliferation is rapidly promoted through the
negative feedback system, and osteoclastogenesis is simultaneously
stimulated to markedly enhance bone absorption ability,
contributing to OP (15,16). However, previous studies have
demonstrated that adiponectin knockout in mice causes a significant
increase in the number of mature osteoclasts compared to that in
wild type mice, suggesting that adiponectin blocks the activity of
osteoblasts to directly or indirectly stimulate osteoclast
differentiation and osteoclastogenesis, resulting in obvious bone
absorption and bone loss leading to OP (17,18).
However, a role of adiponectin in promoting osteoclasts has only
been reported by a few studies (13,19) and
thus it is possible that this effect only occurs under certain
conditions, and not universally.
Monocytes/macrophages and neutrophils can produce a
large amount of adiponectin in response to inflammation (i.e.,
acute host defense against microorganisms, arthritis, and endotoxin
shock), and adiponectin can regulate the bone metabolism effects in
many types of cells, including osteoblasts, chondrocytes and
osteoclasts (20–24). However, to the best of our knowledge,
the effects of adiponectin on bone marrow-derived monocytes (BMMs)
have not been demonstrated to date. Therefore, the aim of the
present study was to investigate the effects of adiponectin
treatment in mouse BMMs and determine its overall influence on
osteoclastogenesis.
Materials and methods
Ethics and BMM culture
All animal experiments were conducted following the
approval of the Institutional Review Board (Medical Ethics
Committee) of the First Affiliated Hospital of Nanchang University
(Nanchang, China). Primary bone marrow samples were obtained from
the long bones (i.e., the femur and tibia) of 4–6-week-old C57BL/6
male mice, as previously described (25,26). A
total of 15 C57BL/6 male mice were purchased from the Department of
Animal Science (Nanchang University, Nanchang, China). Animals were
housed at 18–22°C with 50–60% humidity using 12-h light/dark
cycles. Animals were fed in regular intervals: 3–7 g/day/animal;
3–4 times per week; water ad libitum. In brief, bone marrow
cells were cultured in a T75 flask in complete α-minimal essential
medium (α-MEM) supplemented with 10% fetal bovine serum (FBS) (both
from Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 1%
penicillin/streptomycin, and 50 ng/ml macrophage colony stimulating
factor (M-CSF; R&D Systems, Inc., Minneapolis, MN, USA) at 37°C
in a humidified atmosphere (5% CO2, 95% air) for 24 h.
Non-adherent cells were subsequently collected and reseeded in
another T75 flask to continue incubation. Following culture for 3–4
days, medium was replaced and the culture was washed gently with
phosphate-buffered saline (PBS) three times to remove impurities
and non-adherent cells; adherent cells were used as the BMMs. Cell
culture was continued for ~3 days until the cells reached 90%
confluence. BMMs were collected by trypsin digestion, and then the
cells were continuously cultured in fresh plates or prepared for
subsequent experiments.
Cell viability assay
To clarify the effects of adiponectin on the growth
and proliferation of BMMs, the Cell Counting Kit-8 (CCK-8; Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) assay was conducted
according to the manufacturer's instructions. In brief, the BMMs
were treated with various concentrations of adiponectin (0, 0.2, 1
and 5 µg/ml; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China)
in 96-well plates at a density of 5×103 cells/well and
cultured with α-MEM supplemented with 10% FBS and 50 ng/ml M-CSF at
37°C for 24 h, and 10 µl CCK-8 solution was added to each well and
incubated for another 2 h at 37°C. The absorbance was then detected
at a wavelength of 450 nm on an ELX800 HT spectrophotometer
(Bio-Tek Instruments Inc., Winooski, VT, USA). Cell viability was
calculated relative to that of the control group. The experiment
was repeated three times independently. The viability and growth of
BMMs were plotted and statistically compared using GraphPad Prism
5.0 software.
Osteoclastogenesis assay
The BMMs were plated in a 96-well plate at
5×103 cells/well. The cells were cultured in complete
α-MEM supplemented with 10% FBS, 1% penicillin/streptomycin, and 50
ng/ml M-CSF, along with 50 ng/ml RANKL (PeproTech, Inc., Rocky
Hill, NJ, USA) and different concentrations of adiponectin (0, 0.2,
1 and 5 µg/ml). The cell culture medium was changed every 2 days
until the mature osteoclasts formed (~1 week). The cells were then
washed with PBS three times. The cells were fixed in 4% buffered
paraformaldehyde for 30 min at 37°C and stained for
tartrate-resistant acid phosphatase (TRAP) at 37°C for 1 h. The
number and area of multinucleated (two or more nuclei)
TRAP-positive cells were counted in all wells under a light
microscope (magnification, ×100).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
RT-qPCR was used to determine the mRNA expression
level of adiponectin in the BMMs. Following three generations of
BMM culture, cells were seeded in 6-well plates in α-MEM
supplemented with 10% FBS, 1% penicillin/streptomycin and 50 ng/ml
M-CSF, with or without 1 µg/ml adiponectin, and incubated at 37°C
for 24 h. Total RNA was then extracted from each well using TRIzol
reagent (Life Technologies; Thermo Fisher Scientific, Inc.)
following the manufacturer's suggestion. Total RNA (1 mg) was used
to prepare cDNA using reverse transcriptase (Takara Bio, Inc.,
Otsu, Japan). Samples contained total RNA (2 µl), anchored oligo
(DT)18 primer (0.5 µg/µl; 1 µl), random primer (0.1
µg/µl; 1 µl), 2X ES reaction mix (10 µl), EasyScript RT/RI Enzyme
mix (1 µl), gDNA remover (1 µl) and water to 20 µl. The mixture was
incubated for 10 min at 25°C and reverse transcribed at 42°C for 15
min. cDNA samples (2 µl per 20 µl reaction) were then used as
templates for qPCR to detect the expression of the gene of interest
and reference gene (adiponectin and GAPDH, respectively). qPCR was
performed using SYBR Premix Ex Taq II (Tli RNaseH Plus; DRR820A;
Takara Bio, Inc.) with an ABI StepOnePlus system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) under the following
cycling conditions: Initial denaturation at 95°C for 5 min, 40
cycles of denaturation at 95°C for 10 sec, and amplification at
60°C for 30 sec, with a final extension for 30 sec at 72°C. All
expression levels were calculated using the 2−ΔΔCq
method as described previously (27). GAPDH was used as a control for
normalizing gene expression levels. Each sample was repeated three
times. The following primers were used: GAPDH, forward,
5′-TGACCTCAACTACATGGTCTACA-3′ and reverse,
5′-CTTCCCATTCTCGGCCTTG-3′; and adiponectin, forward,
5′CATGCCCATTCGCTTTACCA-3′ and reverse,
5′-GGAGGCCTGGTCCACATTAT-3′.
Western blotting
Generation-three BMMs were seeded in a
10-cm-diameter petri dish at a density of 5×105
cells/petri dish and cultured in α-MEM supplemented with 10% FBS,
1% penicillin/streptomycin, and with 0, 10, 30, or 50 ng/ml M-CSF
at 37°C. When the cells were fully confluent, they were washed with
PBS three times, and 5 µl protease inhibitor (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) and 495 µl radioimmunoprecipitation assay
buffer (Applygen Technologies, Inc., Beijing, China) were added to
each petri dish. The cells were lysed and mixed on an ice slab for
8 min, and then transferred to EP tubes. Lysates were centrifuged
at 12,000 × g at 4°C for 10 min, and the supernatants that
contained the proteins were collected. Protein concentrations were
measured using a bicinchoninic acid assay. A total of 25 µg of each
protein lysate was resolved using 10% SDS-PAGE, and
electrophoretically transferred to polyvinylidene difluoride
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
then blocked with 5% skimmed milk in TBS-Tween (0.05 M Tris, 0.15 M
NaCl pH 7.5 and 0.2% Tween-20) for 1 h at room temperature,
sequentially blotted with the primary antibodies anti-adiponectin
19F1 (1:1,000; cat. no. D8730) and anti-β-actin (1:2,000; cat. no.
D2764) for overnight at 4°C and secondary antibody (horseradish
peroxidase-conjugated goat anti-rabbit IgG; 1:1,000; cat. no.
P1402) (all from Abcam, Cambridge, UK) for 2 h at room temperature
and visualized using enhanced chemiluminescence (Sigma-Aldrich;
Merck KGaA). The intensity of each band was analyzed using Image
Lab 4.0.1 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
All statistical analyses were performed using SPSS
23.0 software (IBM Corporation, Armonk, NY, USA). The data are
expressed as the means ± standard deviation. One-way analysis of
variance (ANOVA) was performed for comparisons among multiple
groups, and Tukey's test was used for the post hoc test that
followed ANOVA. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effects of adiponectin on the growth
and proliferation of BMMs
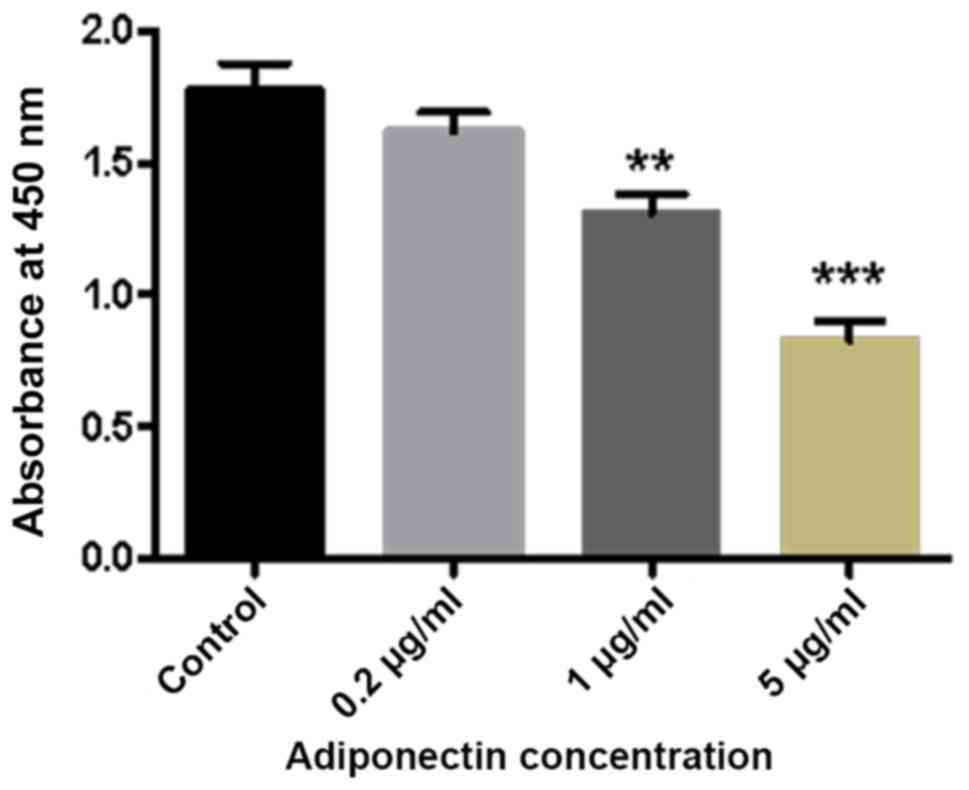
The results of the CCK-8 assay demonstrated a marked
decrease in the absorbance of BMMs with increasing adiponectin
concentration. Furthermore, compared with the control group, there
were significant differences in cell proliferation detected in the
1 and 5 µg/ml adiponectin groups (P<0.01 and P<0.001,
respectively; Fig. 1).
Effect of adiponectin on
osteoclastogenesis
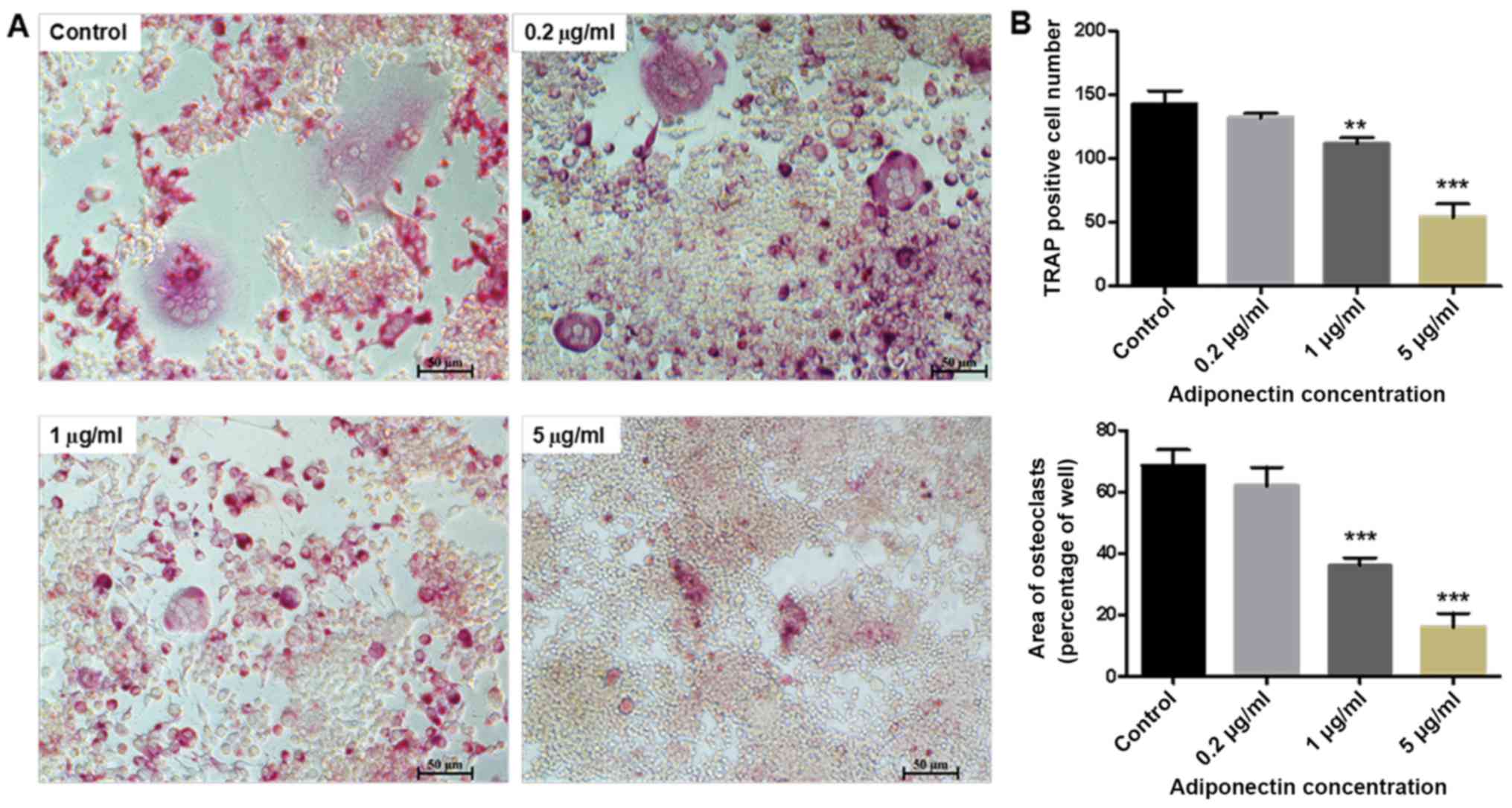
As presented in Fig.
2A, a large number of osteoclasts was observed in the control
group (0 µg/ml adiponectin). However, treatment with 5 µg/ml
adiponectin strongly inhibited the RANKL-induced formation of
osteoclast-like multinucleated cells from the BMM cells. Both the
number and area of multinucleated (two or more nuclei)
TRAP-positive cells were significantly higher in the control group
than in the 1 and 5 µg/ml adiponectin groups (P<0.01 and
P<0.001, respectively; Fig.
2B).
Expression of adiponectin mRNA and
protein in BMMs
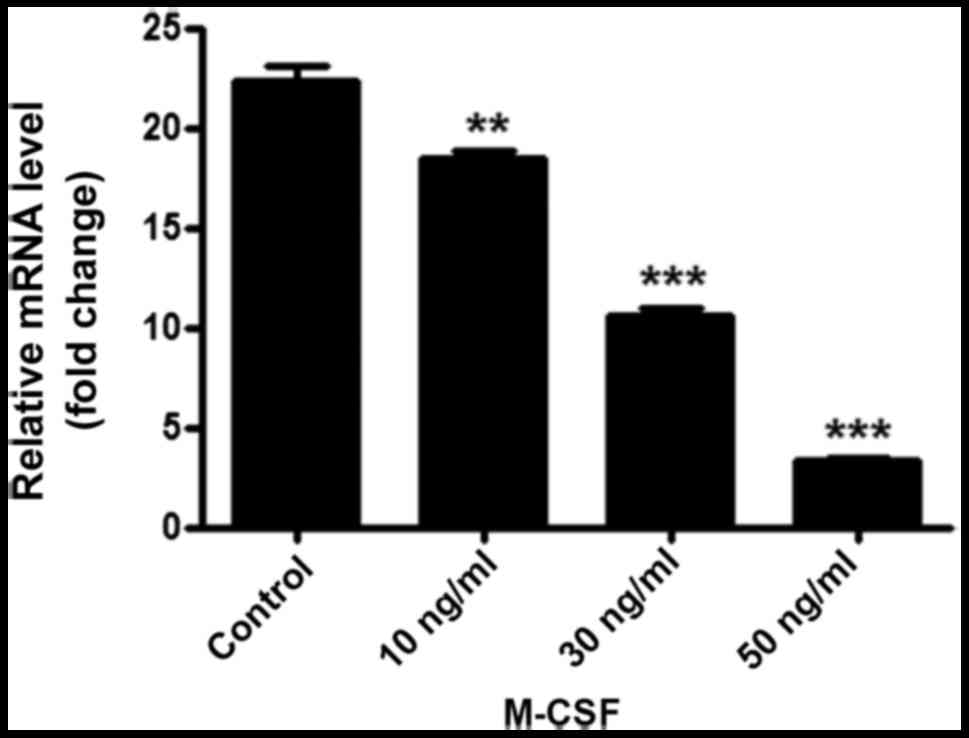
The qPCR results demonstrated that adiponectin mRNA
is expressed in BMMs. Furthermore, with an increasing concentration
of M-CSF, the adiponectin mRNA content gradually and significantly
decreased (P<0.01; Fig. 3).
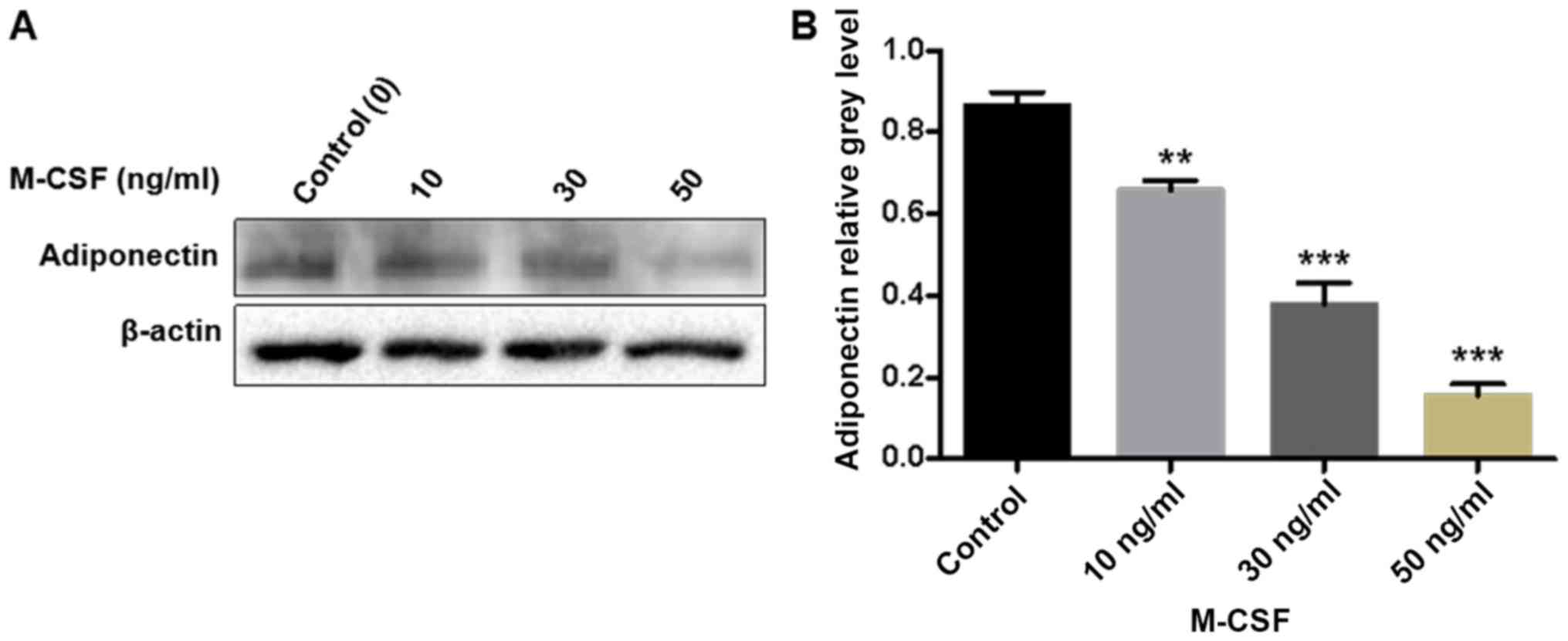
Similarly, the western blotting results demonstrated that an
increasing concentration of M-CSF gradually decreased the protein
expression of adiponectin in BMMs, with a particularly large
decrease detected in the group treated with 50 ng/ml M-CSF. Gray
value analysis of each band indicated a statistically significant
difference between the groups (P<0.001; Fig. 4).
Discussion
The present study confirms a direct effect of
adiponectin on BMMs, indicating an important regulatory role in
bone metabolism. First, adiponectin inhibited the growth and
proliferation of BMMs in a concentration-dependent manner, which is
consistent with previous findings (28). Further, adiponectin strongly
inhibited RANKL-induced osteoclasts formation. Thus, the present
study suggests that adiponectin serves an important role as a
potent negative regulator of osteoclastogenesis in BMMs, confirming
speculations of the relevant literature.
Osteoclast differentiation and formation is a
complex process that is mediated by many factors, including
important cytokines and signaling pathways such as M-CSF and RANKL,
two key cytokines that contribute to inducing the maturation of
osteoclasts (29–31). RANKL expressed on the surface of
osteoblasts/interstitial cells combines with the RANK on the
surface of the osteoclast precursor cells, which greatly induces
differentiation into mature osteoclasts and further promotes cell
proliferation (32). RANKL-RANK
signaling thus serves a pivotal role in osteoclastogenesis and
osteoclast function (33). Yamaguchi
et al (34) previously
proposed that adiponectin acts as a potent regulator of bone
resorption and osteoclastogenesis. The mechanism likely comprises
the suppression of the TNF-α/RANKL-induced differentiation of
osteoclasts by interfering with TNF receptor-associated factor 6
production and calcium signaling. As NF-κB regulated by RANKL has
an important role in the formation and function of osteoclasts
(35), this suggests that
adiponectin may be closely associated with NF-κB signaling
pathways. Further, adiponectin may inhibit the expression of a key
protein in the process of the induction of BMMs into mature
osteoclasts, ultimately resulting in a decrease in osteoclast
formation and bone resorption. As BMMs treated with a high
concentration of M-CSF exhibited significantly suppressed
expression of adiponectin at both the mRNA and protein levels, it
is likely that BMMs can secrete a certain amount of adiponectin
when cultured in M-CSF. It can be speculated that among the BMMs
secreting endogenous adiponectin by autocrine, paracrine and
endocrine mechanisms, the generated adiponectin will constantly
reduce its own expression in BMMs stimulated with a high
concentration of M-CSF via negative feedback regulation (36).
Nevertheless, there are several limitations of the
present study that should be considered. First, it was not feasible
to fully explore the molecular mechanism by which adiponectin
exerts its effects on BMMs, including which signaling pathway is
affected, which will be a focus of subsequent research. Second,
these results are only based on in vitro experiments in
BMMs; therefore, an in vivo experiment with an animal model
will be required to verify the results and determine the potential
clinical relevance of adiponectin in OP.
In summary, the present study is a clarification of
direct effects of adiponectin on bone cells. Specifically,
stimulation of osteoblast proliferation by adiponectin was
demonstrated, together with inhibition of osteoclastogenesis. These
findings have demonstrated and validated this via cell viability
assay, osteoclastogenesis assay and detection of relevant proteins
and genes by western blotting and RT-qPCR. Furthermore, primary
cells were used as the research basis, which yielded more reliable
results. This is due to commercial cell lines exhibiting
biochemical alteration, which leads to variations in the cells.
Accordingly, these findings support that adiponectin may be
developed as a synthetic drug targeting the activity of osteoblasts
or osteoclasts in the future, which may ultimately have utility for
the treatment of OP.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 00019509) and the
Gan-Po Talents Project 555 of Jiangxi Province.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW conceived and designed the study, performed the
experiments, analyzed the data, prepared reviewed and edited the
manuscript. LH performed the experiments and analyzed the data. JL
conceived and designed the study and revised the manuscript. All
authors read and approved the manuscript.
Ethics approval and consent to
participate
All animal experiments were conducted following the
approval of the Institutional Review Board (Medical Ethics
Committee) of the First Affiliated Hospital of Nanchang University
(Nanchang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BMMs
|
bone marrow-derived monocytes
|
|
M-CSF
|
macrophage colony stimulating
factor
|
|
OP
|
osteoporosis
|
|
TNF
|
tumor necrosis factor
|
|
TRAP
|
tartrate resistant acid
phosphatase
|
|
CCK-8
|
cell counting kit-8
|
|
RANKL
|
receptor activator of nuclear
factor-κB ligand
|
|
FBS
|
fetal bovine serum
|
|
ANOVA
|
analysis of variance
|
References
|
1
|
Vats A, Tolley NS, Polak JM and Buttery
LD: Stem cells: Sources and applications. Clin Otolaryngol Allied
Sci. 27:227–232. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Poudyal H and Brown L: Osteoporosis and
its association with non-gonadal hormones involved in hypertension,
adiposity and hyperglycaemia. Curr Drug Targets. 14:1694–1706.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Consensus development conference:
Diagnosis, prophylaxis, and treatment of osteoporosis. Am J Med.
94:646–650. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
NIH Consensus Development Panel on
Osteoporosis Prevention, Diagnosis, and Therapy: Osteoporosis
prevention, diagnosis, and therapy. JAMA. 285:785–795. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harvey N, Dennison E and Cooper C:
Osteoporosis: Impact on health and economics. Nat Rev Rheumatol.
6:99–105. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kanis JA, Odén A, McCloskey EV, Johansson
H, Wahl DA and Cooper C: IOF Working Group on Epidemiology and
Quality of Life: A systematic review of hip fracture incidence and
probability of fracture worldwide. Osteoporos Int. 23:2239–2256.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Melton LJ III, Chrischilles EA, Cooper C,
Lane AW and Riggs BL: Perspective. How many women have
osteoporosis? J Bone Miner Res. 7:1005–1010. 1992.PubMed/NCBI
|
|
8
|
South-Paul JE: Osteoporosis: Part I.
Evaluation and assessment. Am Fam Physician. 63:897–904, 908.
2001.PubMed/NCBI
|
|
9
|
Kanazawa I: Adiponectin in metabolic bone
disease. Curr Med Chem. 19:5481–5492. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kubota N, Terauchi Y, Yamauchi T, Kubota
T, Moroi M, Matsui J, Eto K, Yamashita T, Kamon J, Satoh H, et al:
Disruption of adiponectin causes insulin resistance and neointimal
formation. J Biol Chem. 277:25863–25866. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamauchi T, Kamon J, Minokoshi Y, Ito Y,
Waki H, Uchida S, Yamashita S, Noda M, Kita S, Ueki K, et al:
Adiponectin stimulates glucose utilization and fatty-acid oxidation
by activating AMP-activated protein kinase. Nat Med. 8:1288–1295.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hotta K, Funahashi T, Arita Y, Takahashi
M, Matsuda M, Okamoto Y, Iwahashi H, Kuriyama H, Ouchi N, Maeda K,
et al: Plasma concentrations of a novel, adipose-specific protein,
adiponectin, in type 2 diabetic patients. Arterioscler Thromb Vasc
Biol. 20:1595–1599. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo XH, Guo LJ, Xie H, Yuan LQ, Wu XP,
Zhou HD and Liao EY: Adiponectin stimulates RANKL and inhibits OPG
expression in human osteoblasts through the MAPK signaling pathway.
J Bone Miner Res. 21:1648–1656. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jürimäe J and Jürimäe T: Plasma
adiponectin concentration in healthy pre- and postmenopausal women:
Relationship with body composition, bone mineral, and metabolic
variables. Am J Physiol Endocrinol Metab. 293:E42–E47. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lewiecki EM: RANK ligand inhibition with
denosumab for the management of osteoporosis. Expert Opin Biol
Ther. 6:1041–1050. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schett G, Hayer S, Zwerina J, Redlich K
and Smolen JS: Mechanisms of Disease: The link between RANKL and
arthritic bone disease. Nat Clin Pract Rheumatol. 1:47–54. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Szulc P and Delmas PD: Biochemical markers
of bone turnover: Potential use in the investigation and management
of postmenopausal osteoporosis. Osteoporos Int. 19:1683–1704. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamauchi T and Kadowaki T: Physiological
and pathophysiological roles of adiponectin and adiponectin
receptors in the integrated regulation of metabolic and
cardiovascular diseases. Int J Obes. 32 Suppl 7:S13–S18. 2008.
View Article : Google Scholar
|
|
19
|
Shinoda Y, Yamaguchi M, Ogata N, Akune T,
Kubota N, Yamauchi T, Terauchi Y, Kadowaki T, Takeuchi Y, Fukumoto
S, et al: Regulation of bone formation by adiponectin through
autocrine/paracrine and endocrine pathways. J Cell Biochem.
99:196–208. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Belaaouaj A, Kim KS and Shapiro SD:
Degradation of outer membrane protein A in Escherichia coli
killing by neutrophil elastase. Science. 289:1185–1188. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Adkison AM, Raptis SZ, Kelley DG and Pham
CT: Dipeptidyl peptidase I activates neutrophil-derived serine
proteases and regulates the development of acute experimental
arthritis. J Clin Invest. 109:363–371. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tkalcevic J, Novelli M, Phylactides M,
Iredale JP, Segal AW and Roes J: Impaired immunity and enhanced
resistance to endotoxin in the absence of neutrophil elastase and
cathepsin G. Immunity. 12:201–210. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ouchi N, Kihara S, Arita Y, Okamoto Y,
Maeda K, Kuriyama H, Hotta K, Nishida M, Takahashi M, Muraguchi M,
et al: Adiponectin, an adipocyte-derived plasma protein, inhibits
endothelial NF-kappaB signaling through a cAMP-dependent pathway.
Circulation. 102:1296–1301. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamaguchi N, Argueta JG, Masuhiro Y,
Kagishita M, Nonaka K, Saito T, Hanazawa S and Yamashita Y:
Adiponectin inhibits Toll-like receptor family-induced signaling.
FEBS Lett. 579:6821–6826. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Koga T, Inui M, Inoue K, Kim S, Suematsu
A, Kobayashi E, Iwata T, Ohnishi H, Matozaki T, Kodama T, et al:
Costimulatory signals mediated by the ITAM motif cooperate with
RANKL for bone homeostasis. Nature. 428:758–763. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Burgess TL, Qian Y, Kaufman S, Ring BD,
Van G, Capparelli C, Kelley M, Hsu H, Boyle WJ, Dunstan CR, et al:
The ligand for osteoprotegerin (OPGL) directly activates mature
osteoclasts. J Cell Biol. 145:527–538. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei X, Li G, Yang X, Ba K, Fu Y, Fu N, Cai
X, Li G, Chen Q, Wang M, et al: Effects of bone morphogenetic
protein-4 (BMP-4) on adipocyte differentiation from mouse
adipose-derived stem cells. Cell Prolif. 46:416–424. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chambers TJ: Regulation of the
differentiation and function of osteoclasts. J Pathol. 192:4–13.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Glantschnig H, Fisher JE, Wesolowski G,
Rodan GA and Reszka AA: M-CSF, TNFalpha and RANK ligand promote
osteoclast survival by signaling through mTOR/S6 kinase. Cell Death
Differ. 10:1165–1177. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stanley ER, Berg KL, Einstein DB, Lee PS,
Pixley FJ, Wang Y and Yeung YG: Biology and action of colony -
stimulating factor-1. Mol Reprod Dev. 46:4–10. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang QP, Li XP, Wang M, Zhao LL, Li H, Xie
H and Lu ZY: Adiponectin exerts its negative effect on bone
metabolism via OPG/RANKL pathway: An in vivo study. Endocrine.
47:845–853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yamaguchi N, Kukita T, Li YJ, Kamio N,
Fukumoto S, Nonaka K, Ninomiya Y, Hanazawa S and Yamashita Y:
Adiponectin inhibits induction of TNF-alpha/RANKL-stimulated NFATc1
via the AMPK signaling. FEBS Lett. 582:451–456. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Abu-Amer Y: NF-κB signaling and bone
resorption. Osteoporos Int. 24:2377–2386. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dimitri P and Rosen C: The central nervous
system and bone metabolism: An Evolving Story. Calcif Tissue Int.
100:476–485. 2017. View Article : Google Scholar : PubMed/NCBI
|