Introduction
Glioma is the most common intracranial primary tumor
and domestically accounts for 35–60% of brain tumors (1). As malignant glioma is an invasive
growth, it is difficult to completely remove the tumor cells from
within the surrounding brain tissue by surgery, which ultimately
leads to poor prognoses (2). In
addition, as the blood-brain barrier can limit the entry of
anti-cancer drugs into the central nervous system, surgery combined
with comprehensive chemotherapy or radiotherapy treatment is not
sufficiently effective, and the median survival of patients with
highly malignant glioblastoma remains <1 year (3). With the gradual elucidation of the
effects of long-chain non-coding RNA (lncRNA) and microRNA (miRNA
or miR) and the invasion mechanism of glioma, lncRNA and miRNA were
demonstrated to serve a regulatory role in the glioma invasion
process (4). Therefore, a novel
method to clinically inhibit malignant glioma cell migration and
invasion is required to identify lncRNA and miRNAs and explore the
molecular mechanism by which they regulate glioma cell migration
and invasion.
According to previous studies, lncRNA-PVT1 is highly
expressed in breast cancer, prostate cancer, ovarian cancer,
gastric cancer and lung cancer (4–7).
However, the high expression of lncRNA-PVT1 in tumor cells remains
unclear. In recent years, numerous studies have confirmed that
miR-200 family expression disorders exist in multiple tumor tissues
and participate in the regulation of biological characteristics
such as tumor cell proliferation, migration and invasion (8–10). The
miR-200 family inhibits epithelial-mesenchymal transition (EMT) by
directly targeting the expression of zinc finger E-box binding
homeobox 1/2 (ZEB1/ZEB2) and then inhibits the migration and
invasion of tumor cells (11,12).
miR-200a can inhibit the migration and invasion of cluster of
differentiation 133/1+ ovarian cancer stem cells (13) and nasopharyngeal carcinoma cells
(14) by downregulating ZEB2. In
previous studies, miRNA arrays were used to screen miRNA expression
profiles closely associated with their invasion and migration in
high-grade and low-grade glioma cells (15,16). The
miR-200 family was demonstrated to be more notably expressed in
high-grade glioma cells than in low-grade glioma cells (16). The prominent feature of high-grade
glioma cells compared with low-grade glioma cells is their high
invasiveness (16). Therefore, it
was speculate that the miR-200 family may participate in the
regulation of glioma cell migration and invasion (17). In the present study, the association
between lncRNA-PVT1 was investigated and miR-200a and their role in
the migration and invasion of glioma cells was explored in order to
reveal the key miRNAs and molecular mechanisms that regulate the
migration of glioma cells and to identify novel methods for the
clinical inhibition of glioma cell migration and invasion.
Materials and methods
Specimens and cell culture
Paired glioma samples (grade II–IV) and adjacent
non-tumor tissues were obtained from 48 patients who had undergone
gastrointestinal surgery between 2008 and 2013 at the First
Hospital of Lanzhou University (Lanzhou, China). The patients' age
range was from 30–70 years old. There were 23 female and 28 male
patients recruited. All specimens were immediately frozen in liquid
nitrogen and stored at 80°C until total RNA extraction. Written
informed consent was obtained from all patients. No patient
received chemotherapy or radiotherapy before surgery. The protocol
was approved by the Research Ethics Committee of First Hospital of
Lanzhou University (Gansu, China).
The human glioma cell lines U-87MG, U373MG, U251,
SHG44, T98G and CHG-5, and human astrocyte normal cells HEB were
purchased from the Cellular Resource Center of Shanghai Institutes
for Biological Sciences, Chinese Academy of Sciences (Shanghai,
China). Cells were cultured at 37°C in high-glucose Dulbecco's
modified Eagle's medium containing 10% fetal bovine serum (both
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and were
dissociated with 0.25% trypsin. In addition, the U-87 cell line
used is derived from a glioblastoma of unknown origin and has been
authenticated as the U-87MG strain by STR profiling. Furthermore,
the U-373-MG cell line used is known to be a derivative of the
human U-251-MG astrocytoma cell line, and has been authenticated as
the U-373-MG strain by STR profiling.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
RNA from glioma tissue and cells was extracted using
TRIzol reagent (Life Technologies; Thermo Fisher Scientific, Inc.)
and the RNA was measured using a NanoDrop-2000 spectrophotometer.
The cDNA was then synthesized using PrimeScript RT Master Mix
System (Takara Biotechnology Co., Ltd., Dalian, China) following
the manufacturer's protocol. The PCR solution was prepared on ice
as follows: 1 µl cDNA, 1 µl primers and 10 µl of SYBR Green qRT-PCR
Master Mix (5 µl; Invitrogen; Thermo Fisher Scientific, Inc.). The
final volume was then adjusted to 20 µl using RNase-free water. All
reactions were carried out in an ABI FAST 7500 system. The relative
expression level of each gene was calculated using the
2−ΔΔCq method (9). Each
reaction was repeated three times. Primers for PCR amplification
were as follows: PVT1 forward, 5′-AAAACGGCAGCAGGAAATGT-3′ and
reverse, 5′-ATTCCCATAGAAGGGGCAGG-3′; miR-200a forward,
5′-CGTAACACTGTCTGGTAACGATGT-3′; and U6 forward,
5′-CTCGCTTCGGCAGCACA-3′. The reverse primers for miR-200a and the
internal reference U6 are universal primers from the EXPRESS
SYBR® GreenER™ miRNA qRT-PCR kit (Invitrogen; Thermo
Fisher Scientific, Inc.). GAPDH (forward,
5′-CGGAGTCAACGGATTTGGTCGTAT-3′ and reverse,
5-AGCCTTCTCCATGGTGGTGAAGAC-3) was used as a reference protein. The
PCR conditions included an initial denaturation step of 94°C for 2
min, followed by 30 cycles of 94°C for 30 sec, 59°C for 30 sec and
72°C for 2 min, and a final elongation step at 72°C for 10 min.
LncRNA profiling
RNA purity and integrity were analyzed using an
Agilent Bioanalyzer 2100 for lncRNA microarray. Qualified RNA was
purified using RNase-free DNase and RNeasy mini kit (Qiagen, Inc.,
Valencia, CA, USA). Total RNA was then amplified and labeled using
a Low Input Quick Amp Labeling kit, One-Color (Agilent
Technologies, Inc., Santa Clara, CA, USA) following the
manufacturer's protocol. The RNeasy mini kit was then used to
purify labeled complementary (c)RNA. Each slide was hybridized at
65°C with 600 ng Cy3-labeled cRNA using a Gene Expression
Hybridization kit (Agilent Technologies, Inc.) in a hybridization
oven (Agilent Technologies, Inc.). Following 17 h hybridization,
slides were washed in staining dishes (Thermo Fisher Scientific,
Inc.) with a Gene Expression Wash Buffer kit (Agilent Technologies,
Inc.). Slides were scanned with an Agilent microarray scanner
(Agilent Technologies, Inc.) using default settings and dye
channel. Finally, data were extracted with Feature Extraction
Software 10.7 (Agilent Technologies, Inc.). Raw data were
normalized using a quantile algorithm with Gene Spring software
11.0 (Agilent Technologies, Inc.).
Subcellular fractionation
The nuclear and cytosolic fractions of U87 cells
were separated with a PARIS kit (Life Technologies; Thermo Fisher
Scientific, Inc.). RNA was isolated immediately from the fractions
using TRI reagent (Molecular Research Center, Inc., Cincinnati, OH,
USA). qPCR was conducted using SYBR Green Master mix system
(Invitrogen; Thermo Fisher Scientific, Inc.) in a final reaction
volume of 20 µl in a Mx3000 Stratagene PCR amplifier (Agilent
Technologies, Inc.). The primers were used at a final concentration
of 0.5 µM. The sequences for the primers used for qPCR
amplification were as follows: PVT1 forward,
5′-AAAACGGCAGCAGGAAATGT-3′ and reverse, 5′-ATTCCCATAGAAGGGGCAGG-3′.
After the reactions, the cycle threshold (CT) values were
determined using fixed threshold settings. The miRNA expression in
the cytosol and nuclei was normalized to U6 snRNA and mRNA
expression in the cells was normalized to GAPDH. Primer sequences,
thermocycling conditions and the quantification method used as the
same as previously described.
Cell transfection
The short hairpin RNA (shRNA) sequence targeting
lncRNA-PVT1 was ligated into the pLKO.1-Puro vector (Invitrogen;
Thermo Fisher Scientific, Inc.). The lentivirus was packaged in 293
cells (Cellular Resource Center of Shanghai Institutes for
Biological Sciences, Chinese Academy of Sciences), and lentiviruses
were collected from the supernatant. U87 cells (1×105
cells/well) were seeded on 6-well plates and incubated at 37°C for
a further 24 h prior to transfection. The cells were then
transduced lentiviruses using Lipofectamine 3000 (Thermo Fisher
Scientific, Inc.) at 37°C for 24 h. The medium was then replaced
with 100 µl complete culture medium at 37°C. Following 48 h of
transfection, the cells were used for subsequent experiments. A
total of 100 nM small interfering (si)RNA duplexes (si-PVT#1,
si-PVT#2 and si-PVT#3), miR-200a mimics, a miR-200a inhibitor and
negative control (NC) RNA duplexes was synthesized by Shanghai
Jieli Biotechnology Co., Ltd., (Shanghai, China) for use in
transient transfection assays. siRNA sequences were as follows:
si-PVT#1 5′-GUGAUUUACCAGUCAGUGAAU-3′, si-PVT#2
5′-GUGAUUAACAGUCCCUGAAU-3′ and si-PVT#3
5′-GUGAAAUGCCAGUAUAUGAAU-3′. miRNA sequences were as follows:
miR-200a mimics forward, 5′-UGGCAGUCUCUUAGAUGGUGG-3′ and reverse,
5′-GCCAUGCAAGACACUGCCAGG-3′; miR-200a inhibitor forward,
5′-GAGGGCAGAATCATCACGAAGT-3′ and reverse,
5′-TGAGAGATCTGGTTCCCGAAAC-3′ negative control forward,
5′-GCTCGTGGCTTAGGAGATTG-3′ and reverse,
5′-CTGGCAAAGCATGAGGAACT-3′.
Cell growth assay
Cell growth was measured using MTT (5 mg/ml;
Sigma-Aldrich; Merck KGaA). Cells were transfected in 96-well
plates with siRNA or vector and incubated at 37°C for 48 h,
followed by the addition of 20 µl Cell Titer 96 AQueous One
solution (Thermo Fisher Scientific, Inc.) and further incubation at
37°C for 1–4 h. Absorbance was recorded at 490 nm. Six replicates
were conducted in the assay.
Wound healing assay
Cells were cultured in 6-well plates with DMEM
supplemented with 10% FBS. When the cells in the six-well plate
reached 80% confluence, the cell layer was scratched with a sterile
10-ml pipette tip, washed with medium, and incubated at 37°C with
DMEM containing 1% FBS for 48 h. To prevent cell proliferation, the
cells were preincubated with mitomycin C (10 µg/ml; Adooq
Bioscience LLC, Irvine, CA, USA) for 1 h at 37°C. Images were
captured at different time points (0 and 48 h) using a light
microscope at a magnification of ×200.
Transwell assay
U87 cells were adjusted to a concentration of
2.0×105 with serum-free medium following routine
digestion by Trypsin-EDTA solution (Invitrogen; Thermo Fisher
Scientific, Inc.). Then, 200 µl cell suspension was transferred to
the upper chamber of a Transwell plate. The chamber was implanted
in 24-well plates. Then, 500 µl complete medium containing 10% FBS
was added to the lower chamber, and the plates were incubated at
37°C for 24 h. Following removing the Transwell chamber, the
remaining cells were removed from the upper chambers with cotton
swabs. Then, the cells were fixed at 10 min at room temperature
with 3.7% formaldehyde and stained at room temperature for 1 min
with crystal violet. The migrated cells were counted under using a
light microscope (magnification, ×200) and the mean number of
migrated cells per field of view (of five random fields) were
counted, which was considered an indicator of cell invasion
ability.
Cell cycle distribution and apoptosis
analysis
A flow cytometry assay was performed in order to
detect the effect of downregulating lncRNA-PVT1 on cell cycle
distribution and apoptosis. For the cell cycle distribution
analysis, U87 cells were transfected with si-PVT1, collected 72 h
later, subjected to trypsinization, and fixed at 4°C for 18 h with
ice-cold 70% ethanol. The fixed cells were stained at room
temperature for 30 min with with 500 µl propidium iodide (PI)/RNase
buffer (BD Biosciences, San Jose, CA, USA) and analyzed using a
flow cytometer. For cell apoptosis analysis, U87 cells transfected
with si-PVT1 were collected 72 h later, stained at room temperature
for 30 min with Annexin V-fluorescein isothiocycanate
(FITC)/propidium iodide (PI) apoptosis detection kit (BD
Biosciences). The cells were analyzed using a flow cytometer and
Cell Quest acquisition software (version 2.9; BD Biosciences).
Three replicates were performed.
Immunohistochemistry assays
The tumors from the xenograpft mice were fixed using
10% formaldehyde for 2 h at room temperature, embedded in paraffin
and processed into 5-µm-thick sections. Antigen retrieval was
performed on the tumor sections using Lab Vision™ Tris-HCl buffer
for heat-induced epitope retrieval (cat no. AP-9005-050; Thermo
Fisher Scientific, Inc.) for 15 min at 65°C. Following cooling at
room temperature, the sections were washed with distilled water and
blocked in 10% normal goat serum (Thermo Fisher Scientific, Inc.)
at room temperature for 30 min. Paraffin sections were stained with
hematoxylin at room temperature for 5 min and eosin at room
temperature for 3 min. Ki-67 were conducted by using the
streptavidin-peroxidase method (Zhongshan Golden Bridge Biotech
Co., Ltd., Beijing, China). The tissue sections were deparaffinized
with xylene and rehydrated with decreasing concentrations of
ethanol. The sections were incubated overnight using a primary
rabbit polyclonal antibody against Ki-67 (1:500; cat. no. 16667;
Abcam, Cambridge, UK) in a humidified container at 4°C.
Subsequently, proteins were incubated with rabbit
horseradish-peroxidase-labeled immunoglobulin G (1:5,000; cat. no.
ab6728; Abcam) for 12 h at 4°C. Images were acquired using a Leica
IX71 light microscope (Olympus, Japan) at a magnification of
×200.
Luciferase assay
To examine interactions between PVT1 and miR-200a,
wild-type PVT and mutant PVT were combined with pmirGLO-promotor
vector (Shanghai GenePharma Co., Ltd., Shanghai, China) according
to the manufacturer's protocol. 293 cells were plated at a density
of 8×103 cells/well in 96-well plates and cultured at
37°C with 5% CO2 overnight. Cells were then
co-transfected with 100 ng wild-type or mutant luciferase reporter
plasmids, and 400 ng miR-200a mimic or NC miRNAs using
Lipofectamine 3000. After 48 h, luciferase activity was measured
using the Bright-Glo™ Luciferase assay system (Promega Corporation,
Madison, WI, USA) and normalized to that of Renilla luciferase
activity.
Western blot analysis
Protein samples from U87 cells were exrtracted and
homogenized using radioimmunoprecipitation assay lysis buffer
(Invitrogen; Thermo Fisher Scientific, Inc.) and quantified via the
standard BCA method. Equal amounts of protein from each sample (50
µg/lane) were separated by SDS-PAGE on a 10% gel and electroblotted
onto nitrocellulose membranes. Following blocking with TBST
supplemented with 5% skimmed milk at 4°C overnight, the membranes
were incubated with primary antibodies at 4°C overnight. The
primary antibodies used were as follows: anti-NCAM1 (1:1,000; cat.
no. 9272), anti-PTEN (1:1,000; cat. no. 79156) and anti-GAPDH
(1:2,000; cat. no. 8245; Abcam). GAPDH was used as the internal
control. The membranes were then incubated with horseradish
peroxidase-conjugated secondary anti-primary IgG antibodies
(1:5,000; cat. no. 7097; Abcam) for 1 h at room temperature.
Following a wash with TBS, immunoreactive bands on the membrane
were visualized using SuperSignal West Pico Chemiluminescent
Substrate Trial kit (Thermo Fisher Scientific, Inc.), the results
analyzed with Quantity One software (version v4.6; Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
SPSS 18.0 (SPSS, Inc., Chicago, IL, USA) was used
for statistical analysis. Data are presented as the mean ± standard
deviation. Statistical evaluation was performed using a paired
Student's t-test for lncRNA-PVT1 levels in glioma tissues, and
unpaired data were compared using unpaired Student's t-tests.
Comparisons of data between multiple groups were analyzed using one
way analysis of variance followed by Dunnett's test. P<0.05 was
considered to indicate a statistically significant difference. Each
experiment was repeated in triplicate.
Results
lncRNA-PVT1 is upregulated in glioma
tissues and cells
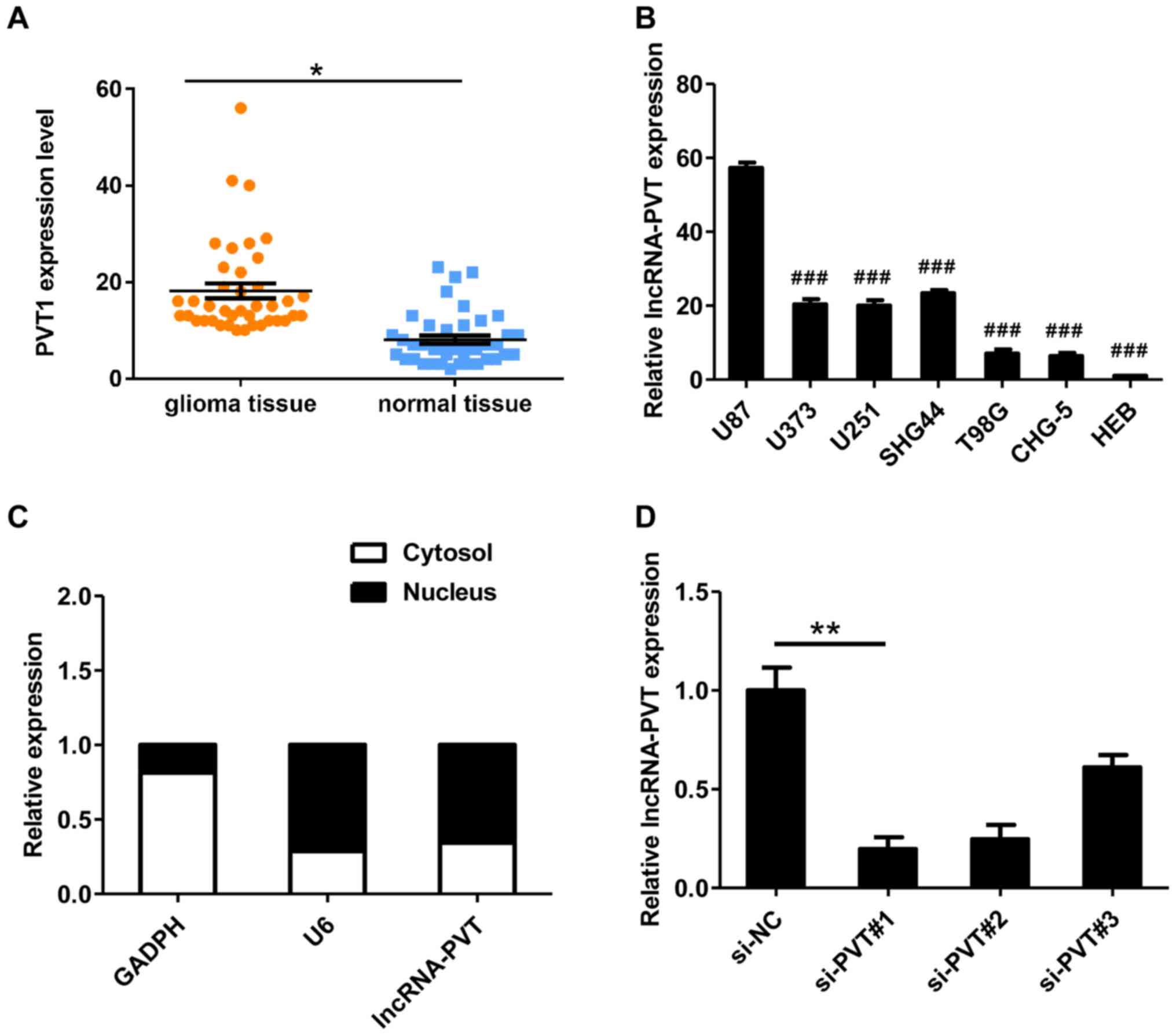
RT-qPCR was used to detect lncRNA-PVT1 levels in
glioma tissues from 48 patients. The expression of lncRNA-PVT in
glioma tissues was significantly higher compared with normal
tissues (Fig. 1A). It was also
demonstrated that lncRNA-PVT was upregulated in glioma cell lines
compared with HEB cells (Fig. 1B)
and preferentially located in the nucleus (Fig. 1C). In summary, these data suggest
that lncRNA-PVT is highly expressed in glioma.
To assess the possible role of lncRNA-PVT in glioma,
three different siRNAs targeting lncRNA-PVT1 were designed (named
si-PVT#1-3) for transfection into U87 cells. These three siRNAs
were effective at reducing the level of endogenous lncRNA-PVT
(Fig. 1D). To avoid the possibility
of an off-target, only si-PVT#1 was used for the follow-up
experiments.
Knockdown of lncRNA-PVT inhibits
glioma cell growth and invasion
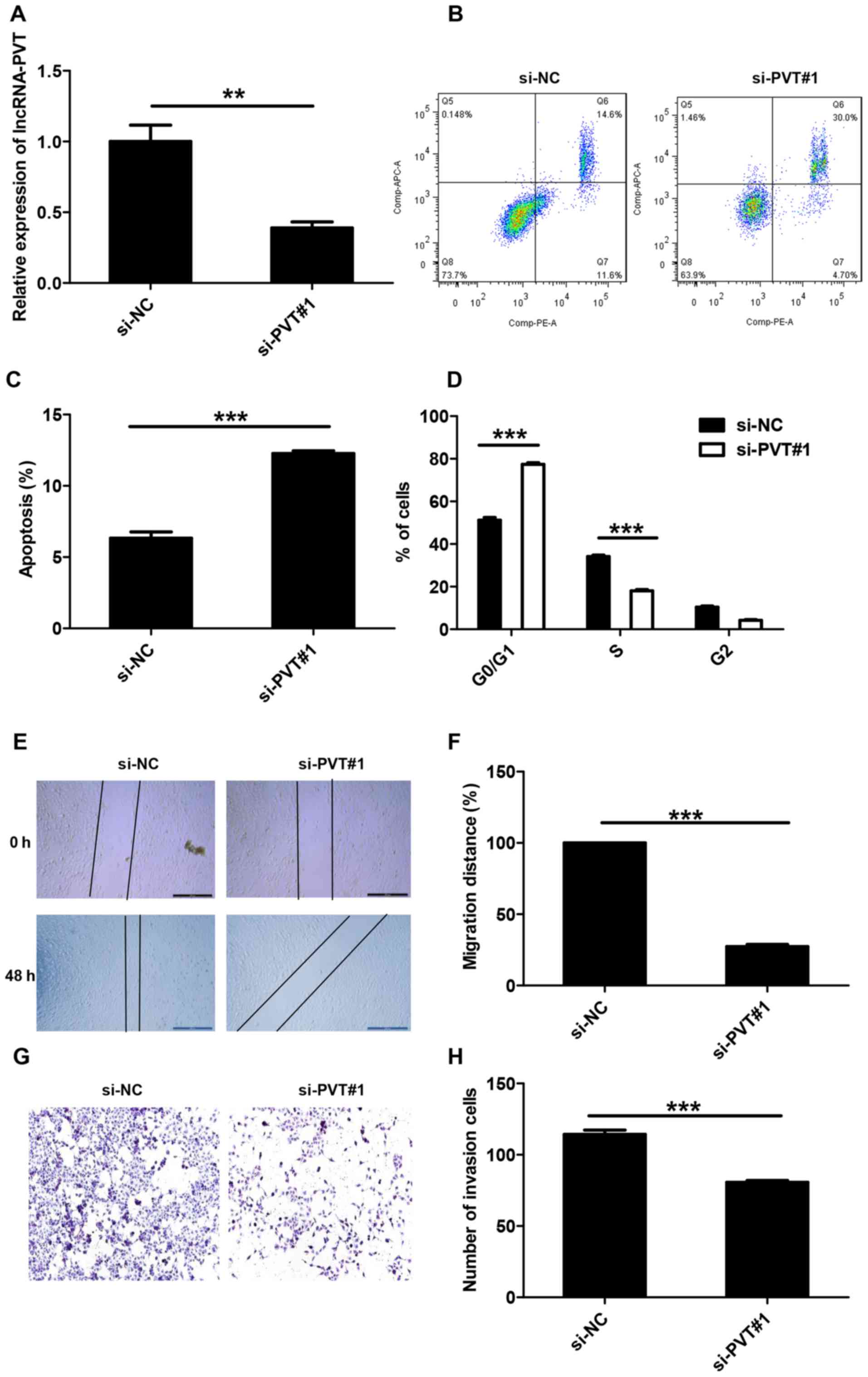
RT-qPCR was used to detect lncRNA-PVT1 levels and
the results demonstrated that lncRNA-PVT1 levels in the si-NC group
was 2.4 times higher compared with the si-PVT#1 group; therefore
the transfection was successful (Fig.
2A). To explore the potential mechanism by which lncRNA-PVT1
promotes the proliferation of glioma cells in vitro, flow
cytometry was used to differentiate the apoptotic population and
cell cycle distribution of lncRNA-PVT and control glioma cells.
Compared with the control group, the si-PVT#1 group exhibited a
significant increase in the percentage of early apoptotic cells
(Fig. 2B and C). At the same time,
significant G0/G1 phase arrest was observed in cells transfected
with si-PVT#1 (Fig. 2D).
To determine whether the expression level of
lncRNA-PVT is associated with glioma, wound-healing experiments and
Transwell experiments were used to analyze the effect of lncRNA-PVT
knockdown on U87 cell invasion. Compared with control treatment,
knockdown of lncRNA-PVT inhibited cell migration (Fig. 2E and F) and significantly reduced
cell invasion (Fig. 2G and H).
lncRNA-PVT is negatively associated
with the expression of miR-200a
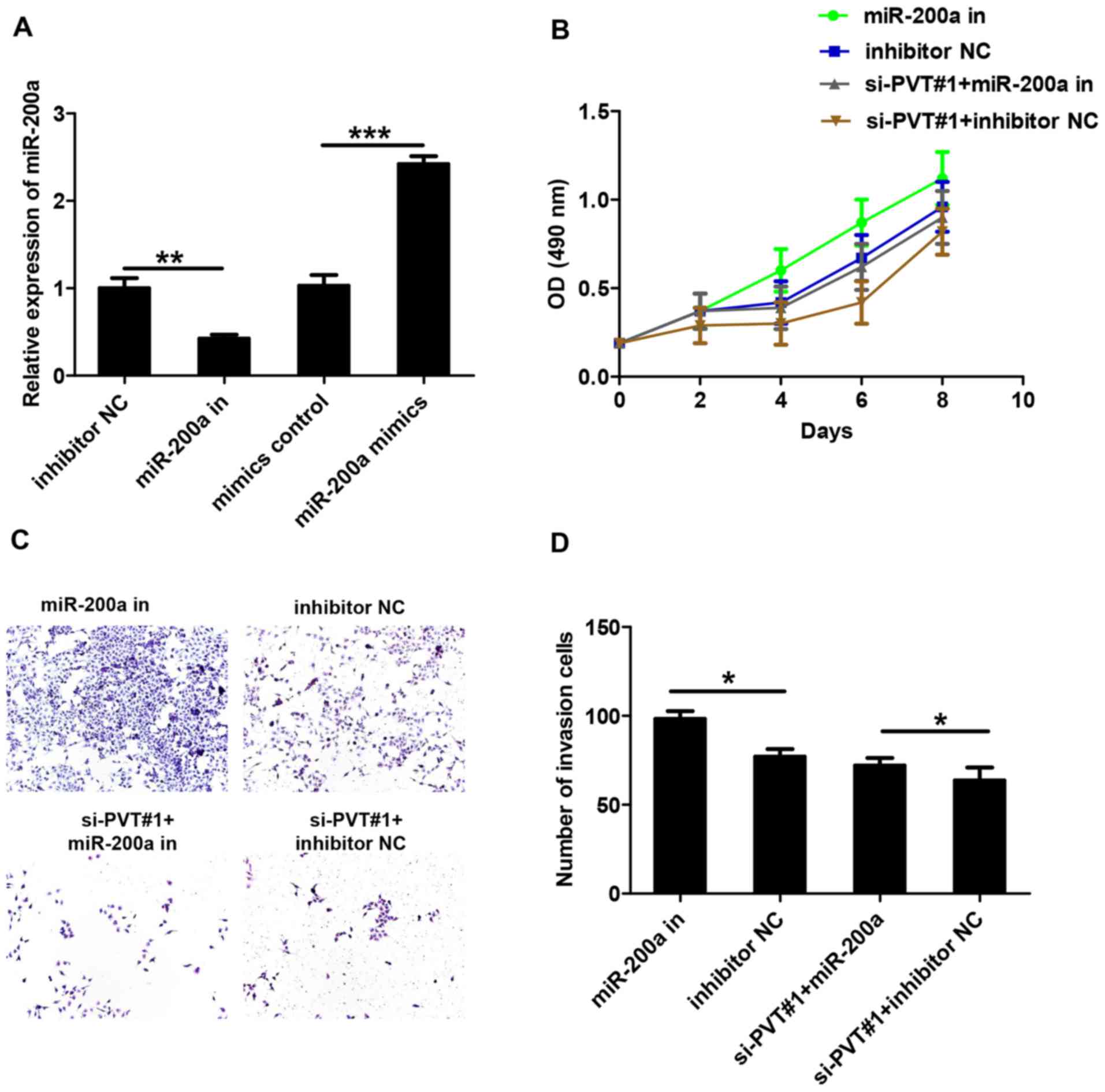
RT-qPCR was used to detect miR-200a levels, and the
results demonstrated transfection success (Fig. 3A). It has been reported previously
that miRNAs interact with lncRNAs to regulate their expression
levels (10,11). To verify the effect of miR-200a on
lncRNA-PVT, U87 cells were transfected with miR-200a inhibitors and
si-PVT#1. MTS proliferation assays demonstrated that miR-200a
inhibitors were able to eliminate the effects of si-PVT#1 on cell
viability (Fig. 3B). A Transwell
invasion assay demonstrated that miR-200a inhibitors enhanced the
invasion of U87 cells compared with the inhibitor NC group
(Fig. 3C and D). In addition, the
co-transfection of si-PVT#1 with miRNA-200a inhibitors eliminated
the inhibitory effect of si-PVT#1 on glioma cell invasion when
compared with the si-PVT#1 + inhibitor NC group. These data
indicate that miR-200a can inhibit the function of lncRNA-PVT.
Interaction between lncRNA-PVT1 and
miR-200a
To evaluate the association between lncRNA-PVT1 and
miR-200a, lncRNA-PVT1 or lncRNA-PVT1 containing a miR-200a binding
site mutation were ligated into the psiCHECK vector and named
PVT-WT and PVT-MUT, respectively. Following transfection with
miR-200a mimic and PVT-WT or PVT-MUT, expression vectors were
co-transfected into cells, and the luciferase activity was measured
by a dual-luciferase assay, and the results demonstrated a
significant decrease in enzyme activity, compared with controls
(Fig. 4A). In addition,
overexpression of lncRNA-PVT1 significantly inhibited the
expression of miR-200a, whereas the silencing of lncRNA-PVT1
increased the expression of miR-200a (Fig. 4B and C). However, inhibition of
miR-200a expression enhanced the expression of lincRNA-PVT1. The
expression of lncRNA-PVT1 was attenuated following the
co-transfection of miR-200a inhibitor with si-PVT#1 into cells
compared with transfection with the NC control (Fig. 4D). Previous studies have reported
that miR-200a can regulate the expression of phosphatase and tensin
homolog and neural cell adhesion molecule 1 (18). Western blot analysis demonstrated
that overexpression of miR-200a inhibited the expression of these
genes (Fig. 4E). The inhibition of
the expression of these genes was also observed in U87 cells
transfected with si-PVT#1 (Fig. 4F).
In addition, the expression of these genes was inhibited when the
miR-200a inhibitor and si-PVT#1 were co-transfected into U87 cells
(Fig. 4G). These results suggest
that miR-200a is targeted by lncRNA PVT1.
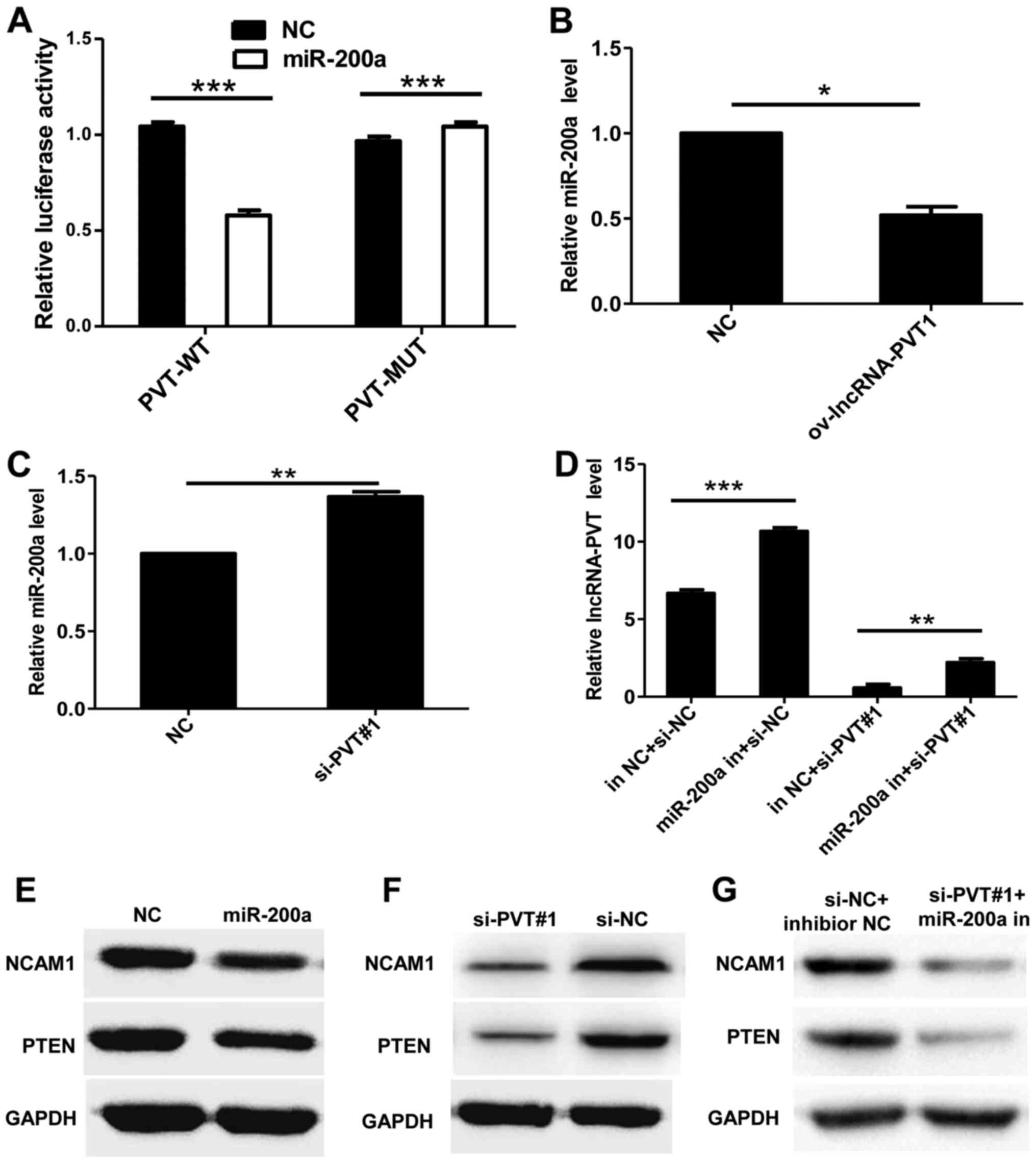 | Figure 4.Interaction between lncRNA-PVT1 and
miR-200a. (A) Luciferase activity of PVT1 mRNA 3′-untranslated
region (WT or MUT) in the presence or the absence of miR-200a by
dual-luciferase reporter system. (B) Overexpression of lncRNA-PVT1
significantly inhibited the expression of miR-200a, (C) whereas
silencing lncRNA-PVT1 significantly increased the expression of
miR-200a. (D) The expression of lncRNA-PVT1 was attenuated
following co-transfection of miR-200a inhibitors with si-PVT#1 into
cells compared with the transfection of the NC. (E) Western blot
analysis demonstrated that overexpression of miR-200a inhibited the
expression of these genes. (F) The inhibition of gene expression
was also observed in U87 cells transfected with si-PVT#1. (G) The
expression of these genes was also inhibited when the miR-200a
inhibitor and si-PVT#1 were co-transfected into U87 cells. Data are
presented as the mean ± standard deviation of triplicate
experiments. *P<0.05, **P<0.01, ***P<0.001 as indicated.
lncRNA, long non-coding RNA; miR, microRNA; WT, wild-type; MUT,
mutant; si, small interfering RNA; NC, negative control; in,
inhibitor; NCAM1, neural cell adhesion molecule 1; PTEN,
phosphatase and tensin homolog. |
Discussion
Although thousands of lncRNAs have been studied, the
functional studies of lncRNA have just begun. Previous functional
studies have demonstrated that lncRNA may function as an oncogene
in the development and progression of human cancer (2,3).
lncRNAs form a class of non-coding RNAs with a
transcript length of >200 nt (19). As lncRNAs lack a coding sequence
region, they cannot encode a protein, and they work only in the
form of RNA (19). Compared with
miRNA, lncRNA is longer, its structure is more complex, and its
mechanisms of action are more diverse (19). The transcripts produced by 4–9% of
the mammalian genome sequence are lncRNA (the corresponding ratio
of protein-coding RNA is 1%) (19).
Therefore, lncRNA regulation of life processes may be more
extensive (20). Previous studies
have demonstrated that lncRNA is associated with important
physiological processes such as chromatin silencing and genomic
imprinting (21,22). In recent years, lncRNA has been
demonstrated to be closely associated with the development of
various tumors (19). Certain
lncRNAs serve an important role in the development of tumors
(23). Certain abnormal expression
patterns of lncRNAs in tumors have an important role in the cancer
diagnosis, treatment and prognosis (24). lncRNA is gradually receiving greater
focus in tumor research. For example, lncRNA-HOTAIR can promote
breast cancer metastasis by interacting with the polycomb
repressive complex (25). Yang et
al (26,27) have also demonstrated that the lncRNAs
highly expressed in hepatocellular carcinoma and low expression in
tumor have important effects on the development of hepatocellular
carcinoma.
The PVT1 gene is often heterotopic in the tumor and
is expressed with other genes, as Nagoshi demonstrated in the
expression of fusion genes of PVT1-neurobeachin and WW domain
containing oxidoreductase in multiple myeloma (28). According to the published literature,
lncRNA-PVT1 is highly expressed in breast cancer, prostate cancer,
ovarian cancer, gastric cancer and lung cancer (6,7,28). However, the role of high lncRNA-PVT1
expression in tumor cells is not very clear. In the present study,
it was demonstrated that lncRNA-PVT1 promotes the proliferation and
invasion of tumor cells.
Previous studies have demonstrated that exogenous
overexpression of miR-200a can promote the invasion of U87 cells
and that inhibition of miR-200a expression can inhibit the invasion
of U87 cells (16,29). Thus, it was speculated that miR-200a
serves an important role in glioma invasion and may be one of the
key miRNAs associated with the regulation of glioma cell
migration.
In the present study, it was demonstrated that
lncRNA-PVT1 is frequently overexpressed in glioma tissue,
indicating the carcinogenic activity of lncRNA-PVT1. The discovery
of the function of lncRNA-PVT1 was also demonstrated by inhibiting
the expression of lncRNA-PVT1 in glioma cells. In recent years, a
number of studies have confirmed miR-200 family expression
disorders in a variety of tumor tissues, wherein they participate
in tumor cell proliferation, migration, and invasion and the
regulation of biological processes (8–10,29).
Although miR-200a has been demonstrated to target many
protein-coding genes, the present study suggested that miR-200a
also targets lncRNA-PVT1. It was demonstrated that the expression
of lncRNA-PVT1 and miR-200a in clinical glioma tissue was
negatively associated. This suggests that the expression of
miR-200a in glioma tissue can inhibit the expression of
lncRNA-PVT1.
In conclusion, the present data suggest that
lncRNA-PVT1 is upregulated in human glioma tissue and can be used
as a negative prognostic factor in patients with glioma. Decreasing
the expression of lncRNA-PVT1 inhibits the proliferation and
invasion of glioma cells and induces apoptosis. lncRNA-PVT1 acts as
an oncogene by upregulating miR-200a. The present study reveals the
overexpression of PVT1 in glioma tissue and cells and the oncogenic
role of PVT1 in gliomagenesis via sponging of miR-200a, thus
providing a potential biomarker for the early detection and
prognostic prediction of glioma. It has been reported that miR-200a
can directly target the mRNA of β-catenin (CTNNB1 transcription).
The downregulation of miR-200a may affect the tumor development by
activating the wnt pathway and causing epithelial-mesenchymal
transition. In further research, we will continue to explore the
possible mRNA and biologic process associated with glioma
genesis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Project of Lanzhou City (grant no. 2015-2-67) and the
Health Industry Research Plan of Gansu Province (grant no.
GWGL2014-49).
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and GY designed the present study and analyzed
the data. YL and GY performed the experiments.
Ethics approval and consent to
participate
Written informed consent was obtained from all
patients. The protocol was approved by the Research Ethics
Committee of First Hospital of Lanzhou University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tsuzuki T, Izumoto S, Ohnishi T, Hiraga S,
Arita N and Hayakawa T: Neural cell adhesion molecule L1 in
gliomas: Correlation with TGF-beta and p53. J Clin Pathol.
51:13–17. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kleber S, Sancho-Martinez I, Wiestler B,
Beisel A, Gieffers C, Hill O, Thiemann M, Mueller W, Sykora J, Kuhn
A, et al: Yes and PI3K bind CD95 to signal invasion of
glioblastoma. Cancer Cell. 13:235–248. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dolecek TA, Propp JM, Stroup NE and
Kruchko C: CBTRUS statistical report: Primary brain and central
nervous system tumors diagnosed in the United States in 2005–2009.
Neuro Oncol. 14 Suppl 5:v1–v49. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nagoshi H, Taki T, Hanamura I, Nitta M,
Otsuki T, Nishida K, Okuda K, Sakamoto N, Kobayashi S,
Yamamoto-Sugitani M, et al: Frequent PVT1 rearrangement and novel
chimeric genes PVT1-NBEA and PVT1-WWOX occur in multiple myeloma
with 8q24 abnormality. Cancer Res. 72:4954–4962. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu C, Wang Y, Li G, She L, Zhang D, Chen
X, Zhang X, Qin Z, Cao H and Liu Y: LncRNA PVT1 promotes malignant
progression in squamous cell carcinoma of the head and neck. J
Cancer. 9:3593–3602. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wan B, Wu HY, Lv DJ, Zhou XM, Zhong LR,
Lei B, Zhang SB and Mao XM: Downregulation of lncRNA PVT1
expression inhibits proliferation and migration by regulating p38
expression in prostate cancer. Oncol Lett. 16:5160–5166.
2018.PubMed/NCBI
|
|
7
|
Chen Y, Du H, Bao L and Liu W: LncRNA PVT1
promotes ovarian cancer progression by silencing miR-214. Cancer
Biol Med. 15:238–250. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen L, Wang X, Zhu Y, Zhu J and Lai Q:
miR-200b-3p inhibits proliferation and induces apoptosis in
colorectal cancer by targeting Wnt1. Mol Med Rep. 18:2571–2580.
2018.PubMed/NCBI
|
|
9
|
Ren W, Gao L, Qiang C, Li S, Zheng J, Wang
Q, Zhi Y, Cai G, Kong X, Zhou M, et al: Kindlin-2-mediated
upregulation of ZEB2 facilitates migration and invasion of oral
squamous cell carcinoma in a miR-200b-dependent manner. Am J Transl
Res. 10:2529–2541. 2018.PubMed/NCBI
|
|
10
|
Pillman KA, Phillips CA, Roslan S, Toubia
J, Dredge BK, Bert AG, Lumb R, Neumann DP, Li X, Conn SJ, et al:
miR-200/375 control epithelial plasticity-associated alternative
splicing by repressing the RNA-binding protein Quaking. EMBO J.
37(pii): e990162018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Meng YB, He X, Huang YF, Wu QN, Zhou YC
and Hao DJ: Long Noncoding RNA CRNDE Promotes Multiple Myeloma Cell
Growth by Suppressing miR-451. Oncol Res. 25:1207–1214. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xia H, Ng SS, Jiang S, Cheung WK, Sze J,
Bian XW, Kung HF and Lin MC: miR-200a-mediated downregulation of
ZEB2 and CTNNB1 differentially inhibits nasopharyngeal carcinoma
cell growth, migration and invasion. Biochem Biophys Res Commun.
391:535–541. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang C, Kang L, Wang X, Liu Y and Zhao X:
Expression of miR-200a and chemotherapeutic treatment efficacy of
glioma. Oncol Lett. 15:5767–5771. 2018.PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chiyomaru T, Fukuhara S, Saini S, Majid S,
Deng G, Shahryari V, Chang I, Tanaka Y, Enokida H, Nakagawa M, et
al: Long non-coding RNA HOTAIR is targeted and regulated by miR-141
in human cancer cells. J Biol Chem. 289:12550–12565. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu J, Rodova M, Nanta R, Meeker D, Van
Veldhuizen PJ, Srivastava RK and Shankar S: NPV-LDE-225
(Erismodegib) inhibits epithelial mesenchymal transition and
self-renewal of glioblastoma initiating cells by regulating miR-21,
miR-128 and miR-200. Neuro Oncol. 15:691–706. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J, Liu X, Wu H, Ni P, Gu Z, Qiao Y,
Chen N, Sun F and Fan Q: CREB up-regulates long non-coding RNA HULC
expression through interaction with microRNA-372 in liver cancer.
Nucleic Acids Res. 38:5366–5383. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Palazzo AF and Lee ES: Sequence
determinants for nuclear retention and cytoplasmic export of mRNAs
and lncRNAs. Front Genet. 9:4402018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Serviss JT, Johnsson P and Grander D: An
emerging role for long non-coding RNAs in cancer metastasis. Front
Genet. 5:2342014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen LL: Linking long noncoding RNA
localization and function. Trends Biochem Sci. 41:761–772. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang L, Peng D, Sood AK, Dang CV and
Zhong X: Shedding light on the dark cancer genomes: Long noncoding
RNAs as novel biomarkers and potential therapeutic targets for
cancer. Mol Cancer Ther. 17:1816–1823. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Helsmoortel H, Everaert C, Lumen N, Ost P
and Vandesompele J: Detecting long non-coding RNA biomarkers in
prostate cancer liquid biopsies: Hype or hope? Noncoding RNA Res.
3:64–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chisholm KM, Wan Y, Li R, Montgomery KD,
Chang HY and West RB: Detection of long non-coding RNA in archival
tissue: Correlation with polycomb protein expression in primary and
metastatic breast carcinoma. PLoS One. 7:e479982012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang F, Huo XS, Yuan SX, Zhang L, Zhou WP,
Wang F and Sun SH: Repression of the long noncoding RNA-LET by
histone deacetylase 3 contributes to hypoxia-mediated metastasis.
Mol Cell. 49:1083–1096. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang F, Zhang L, Huo XS, Yuan JH, Xu D,
Yuan SX, Zhu N, Zhou WP, Yang GS, Wang YZ, et al: Long noncoding
RNA high expression in hepatocellular carcinoma facilitates tumor
growth through enhancer of zeste homolog 2 in humans. Hepatology.
54:1679–1689. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nagoshi H, Taki T, Hanamura I, Nitta M,
Otsuki T, Nishida K, Okuda K, Sakamoto N, Kobayashi S,
Yamamoto-Sugitani M, et al: PVT1 rearrangement and novel chimeric
genes PVT1-NBEA and PVT1-WWOX occur in multiple myeloma with 8q24
abnormality. Cancer Res. 72:4954–4962. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu F, Zheng Y, Hong W, Chen B, Dong P and
Zheng J: MicroRNA-200a suppresses epithelial-to-mesenchymal
transition in rat hepatic stellate cells via GLI family zinc finger
2. Mol Med Rep. 12:8121–8128. 2015. View Article : Google Scholar : PubMed/NCBI
|