Introduction
Fracture repair is a multi-staged and complex
process, the end goal of which is the recovery of the functional
and biomechanical state (1). A
variety of biological factors influencing the outcome of fracture
healing have been identified in recent decades (2,3). Bone
fracture may immediately sever local vasculature and result in the
formation of a hematoma, which leads to low oxygen tension and
regional hypoxia (4). As a result,
hypoxia-inducible factor-1α (HIF-1α) may be upregulated at the
fracture site. Recently, numerous studies have focused on the
function of HIF-1α in fracture healing (5,6).
Previous work by our group identified that HIF-1α
could be induced by hypoxia in the early stages of bone
regeneration (7). Vascular
endothelial growth factor (VEGF) induced by HIF-1α promotes
angiogenesis, which contributes to intramembranous ossification
adjacent to the fracture site, endochondral ossification at the
fracture site, and the ensuing remodeling phase of fracture repair
(8). Autophagy, as a cell
degradation process, targets dysfunctional cytosolic
macromolecules, membranes and organelles, which serve an important
role in energy and nutrient regulation (9). A previous study has demonstrated that
activation of autophagy may protect organs and tissues against
infarction injury, including neurodegeneration, cardiomyopathies
and abnormal skeletal development (10). It has been demonstrated that
fractures may impair cellular homeostasis and place bone cells
under considerable stress, which may lead to the activation of
autophagy and promote fracture healing in turn (11,12).
An improved understanding of HIF-1α and autophagy in
fracture healing may aid the development of novel potential
therapies for fracture healing. Therefore, the aim of the present
study was to investigate whether autophagy is involved in fracture
healing and the potential role of HIF-1α in autophagy during
fracture healing.
Materials and methods
Animal care and experiments
All experimental procedures involving rats were
approved by the Ethics Committee on Animal Experimentation of
Capital Medical University (Beijing, China; approval no.
AEEI-2017-098). A total of 64 male Sprague-Dawley rats (6 weeks;
220±25 g) were obtained from the Laboratory Animal Center of
Capital Medical University. Upon arrival at the central animal
facility, rats were allowed 1 week to acclimatize prior to random
assignment into two groups: A control group treated with dimethyl
sulfoxide (DMSO; n=32); and an experimental group treated with 0.05
mg/kg echinomycin (n=32). Each group was treated intraperitoneally
every other day for a total of 42 days. Subsequent to peritoneal
injection of 1% sodium pentobarbital (40 mg/kg body weight; cat.
no. P3761; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), all rats
underwent fracture of the left tibia, which was performed with a
blunt guillotine apparatus driven by a drop weight, as previously
described (12). To achieve
intramedullary fixation, a 0.8 mm Kirschner wire (K-wire) was
inserted through the intercondylar notch until it was seated in the
distal cortex. Radiographs were obtained immediately to confirm
K-wire placement as well as the extent of fractures. Rats were
sacrificed at 7, 14, 28 and 42 days after fracture (n=8/group at
each time point) by cervical dislocation following anesthesia by
peritoneal injection of 1% sodium pentobarbital (40 mg/kg body
weight).
Micro-computed tomography (micro-CT)
analysis
K-wires in the tibia were removed carefully in order
to protect the fracture site, and then an Inveon micro-CT (Siemens
AG, Munich, Germany) was used to scan the dissected tibia. The beam
protocol was set as follows: 15-mm isometric voxel size, 800 mA and
80 kV. The proximal and distal bone tissue that was 2.5 mm far away
from the fracture line was selected as the region of interest
(ROI). The callus perimeter was determined by a semi-automated
contouring method. Contours were drawn to reveal the periosteal
surface of the ROI in the tibia. The tibiae were reconstructed
using Mimics software version 20.0 (Materialise NV, Leuven,
Belgium), and bone histomorphometric analysis, including of bone
mineral density (BMD) and bone volume fraction [bone volume/total
volume (BV/TV)], was performed using the built-in software in the
micro-CT system (data not shown) (13).
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (qPCR)
Total RNA was isolated from the callus using TRIzol
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
according to the manufacturer's protocol. Then, cDNA was reverse
transcribed using a ReverTra Ace kit (Toyobo Life Science, Osaka,
Japan). qPCR was performed to measure mRNA levels relative to GAPDH
expression. The sequences of the primers used were as follows: For
runt-related transcription factor 2 (Runx2), forward,
5′-CCCACGAATGCACTATCCAG-3′ and reverse, 5′-GGCTTCCATCAGCGTCAACA-3′;
for alkaline phosphatase (ALP), forward, 5′-GGACGGTGAACGGGAGAAC-3′
and reverse, 5′-CCCTCAGAACAGGGTGCGTAG-3′; for
microtubule-associated protein 1 light chain 3 (LC3 II) forward,
5′-AAACGCATTTGCCATCACA-3 and reverse, 5′-CCCTCAGAACAGGGTGCGTAG-3′;
for P62, forward, 5′-GGGGACTTGGTTGCCTTTT-3′ and reverse,
5′-CAGCCATCGCAGATCACATT-3′; and for Unc-51-like autophagy
activating kinase 1 (ULK1), forward, 5′-TCGAGTTCTCCCGCAAGG-3′ and
reverse, 5′-CGTCTGAGACTTGGCGAGGT-3′. The PCR conditions were set as
follows: 3 min at 95°C, followed by 40 cycles at 95°C for 15 sec,
55–60°C for 30 sec and 72°C for 1 min, then 72°C for 10 min as a
final extension. The expression values were normalized against
GAPDH using the 2−∆∆Cq method (14). In order to minimize confounding
variance, two independent samples were analyzed in technical
triplicates. Technical replicates were averaged prior to all
software analysis.
Immunohistochemistry (IHC)
Sections were prepared and processed using standard
techniques, as described previously (15). Briefly, tissues from the tibia were
decalcified and paraffin embedded in preparation for
tartrate-resistant acid phosphatase staining. Following antigen
retrieval and blocking of endogenous peroxide activity, the slides
were incubated with primary antibodies against ALP (cat. no.
ab84401; Abcam, Cambridge, MA, USA; dilution 1:100), Runx2 (cat.
no. H00000860-M04; Abnova, Taipei, Taiwan; dilution 1:100) and LC3
II (cat. no. ab48394; Abcam; dilution 1:100) in a humidified
chamber at 4°C overnight. Subsequently, sections were incubated
with goat anti-rabbit immunoglobulin G secondary antibody (cat. no.
ab6720; Abcam; dilution 1:100) for 10 min at 37°C. Following
washing with phosphate-buffered saline (PBS), the sections were
incubated with streptavidin-peroxidase conjugate (Rockland
Immunochemicals, Inc., Limerick, PA, USA) for 10 min at room
temperature. Final staining was performed using diaminobenzidine
tetrahydrochloride solution (Ventana Medical Systems, Inc., Tucson,
AZ, USA) and sections were counterstained with hematoxylin for 10
min at room temperature, and then mounted. Scores were assessed by
two independent observers and evaluated according to the percentage
of positively stained cells and intensity of staining.
Immunostaining intensity was rated as follows: 0, negative; 1,
weak; 2, moderate; 3, strong. The percentage of positively stained
cells was rated as follows: Grade 0, <25%; grade 1, <50%;
grade 2, <75%; and grade 3, >75%.
Western blot analysis
Dissected calluses from fractured bone for each time
point (7, 14, 28 and 48 days) were washed with PBS. Tissue lysates
were prepared with lysis buffer containing protease inhibitors
(Sigma-Aldrich; Merck KGaA). Protein quantification was performed
using a bicinchoninic acid protein assay kit (Thermo Fisher
Scientific, Inc.). Lysates (30 mg) were separated on 10%
SDS-polyacrylamide gels and electrotransferred onto polyvinylidene
difluoride membranes (EMD Millipore, Bedford, MA, USA). Membranes
were blocked with 5% non-fat dry milk and incubated overnight at
4°C with primary antibodies against HIF-1α (cat. no. ab463; Abcam;
dilution 1:1,000), ULK1 (cat. no. ab128859; Abcam; dilution
1:1,000), p62 (cat. no. ab96134; Abcam; dilution 1:1,000) and those
aforementioned against ALP (dilution 1:2,000), Runx2 (dilution
1:500) and LC3 II (dilution 1:2,000). Membranes were then incubated
with horseradish peroxidase-conjugated secondary antibodies (cat.
no. ab97200, Abcam; dilution 1:5,000) for 2 h at room temperature.
Bands were developed using chemiluminescence reagents (Thermo
Fisher Scientific, Inc.) and densitometric results were analyzed
with Image Quant LAS4000 (GE Healthcare Life Sciences, Little
Chalfont, UK). GAPDH (cat. no. G5262; Sigma-Aldrich; Merck KGaA;
dilution 1:5,000) served as a loading control.
Statistical analysis
Each experiment was repeated at least three times.
All statistical comparisons were performed with SPSS software,
version 19.0 (IBM Corp., Armonk, IL, USA). Data are presented as
the mean ± standard deviation. The differences between the groups
were analyzed by Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
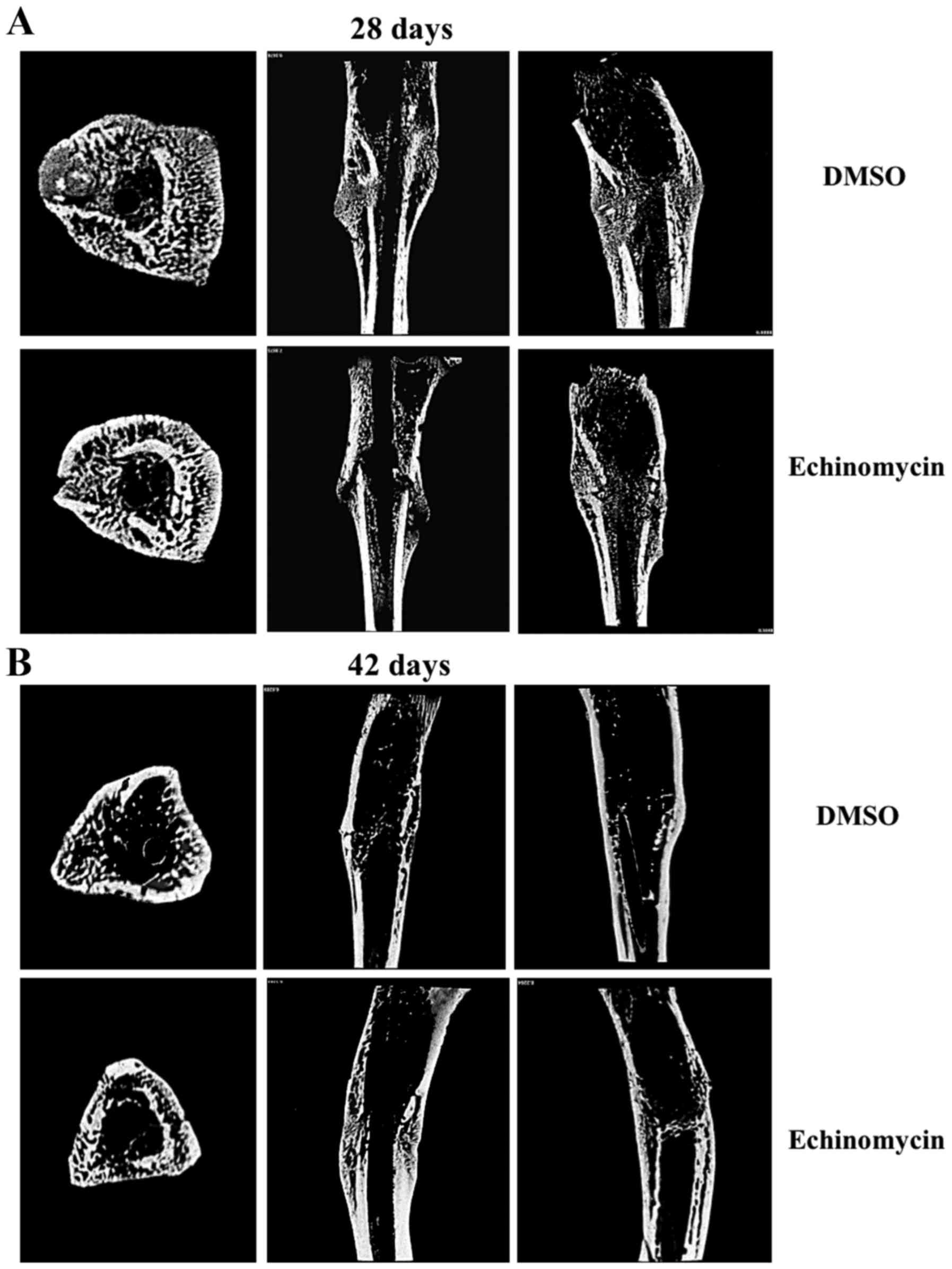
Echinomycin inhibits callus formation
and remodelling
First, the differences in callus formation and
remodeling between echinomycin and DMSO administration groups were
compared. Micro-CT was used to remodel the fracture calluses and to
quantify the degree of mineralization. The results demonstrated
that treatment with echinomycin could markedly inhibit new bone
formation and the subsequent remodeling process of fractured bone
in rats. As a result, fractured tibiae exhibited delayed repair in
echinomycin-treated rats compared with the control group. Notably,
the control group displayed near-complete healing of the tibia at
day 42 (Fig. 1). These results
suggested that loss of function of HIF-1α by echinomycin could
inhibit fracture healing.
Echinomycin modulates HIF-1α target
genes and autophagy markers at the mRNA level
Subsequently, the potential mechanism of inhibition
of HIF-1α in fracture healing was investigated. The mRNA levels of
HIF-1α, its target-genes (osteoblast markers) and autophagy markers
were assessed following echinomycin administration. It was
identified that compared with in the DMSO group, the levels of ALP
(Fig. 2A) and Runx2 (Fig. 2B) were significantly decreased
following echinomycin administration. Furthermore, the levels of
autophagy markers, LC3 II (Fig. 2C)
and ULK1 (Fig. 2D) were also reduced
in the echinomycin treatment group. The mRNA levels of P62 were
increased in the echinomycin treatment group (Fig. 2E), while HIF-1α exhibited no
significant difference in its expression at any time point between
the two groups (Fig. 2F). These
results indicated that loss of function of HIF-1α by echinomycin
may inhibit osteoblast formation and downregulate autophagy in
fracture healing.
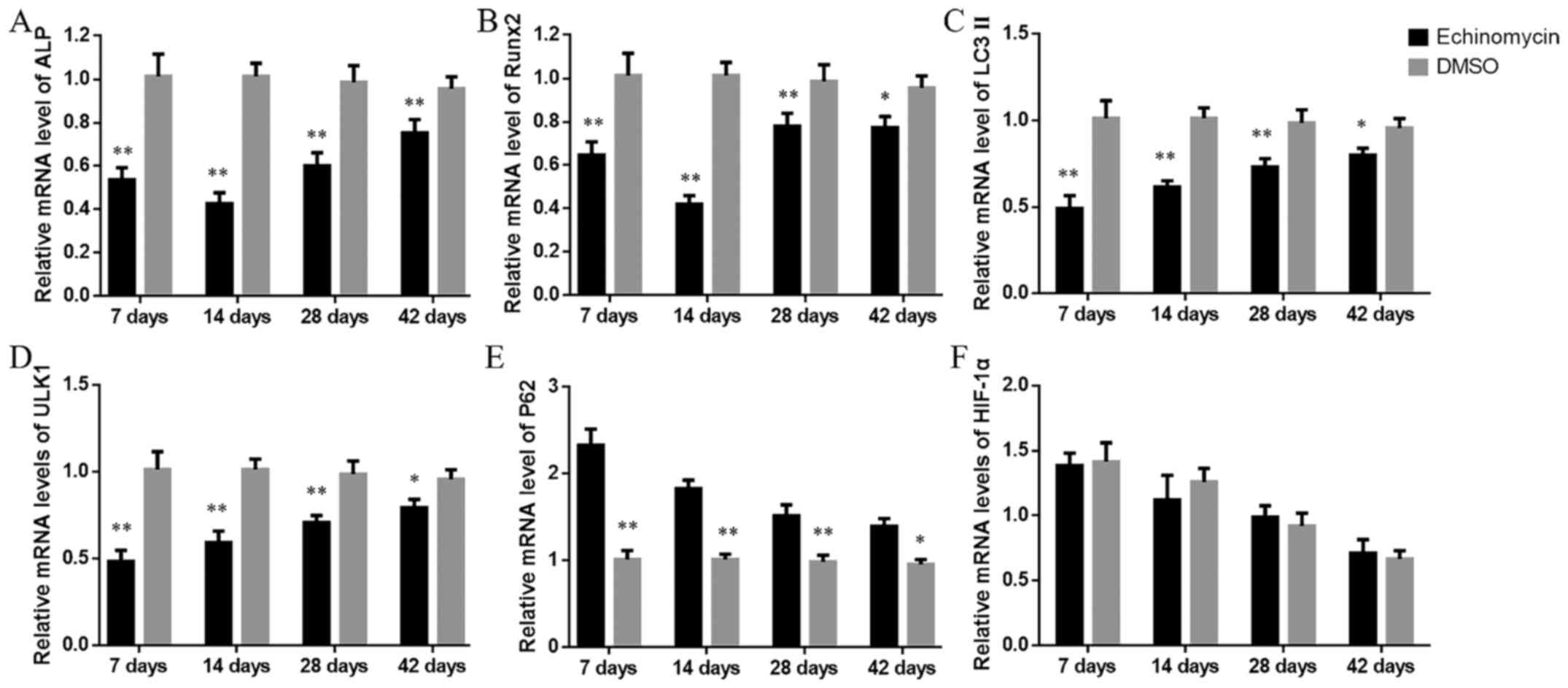 | Figure 2.mRNA levels of (A) ALP, (B) Runx2, (C)
LC3 II, (D) ULK1, (E) P62 and (F) HIF-1α at the indicated time
points. *P<0.05, **P<0.01 vs. DMSO-treated group. ALP,
alkaline phosphatase; Runx2, runt-related transcription factor 2;
LC3 II, microtubule-associated protein 1 light chain 3; ULK1,
Unc-51 like autophagy activating kinase 1; HIF-1α,
hypoxia-inducible factor-1α; DMSO, dimethyl sulfoxide. |
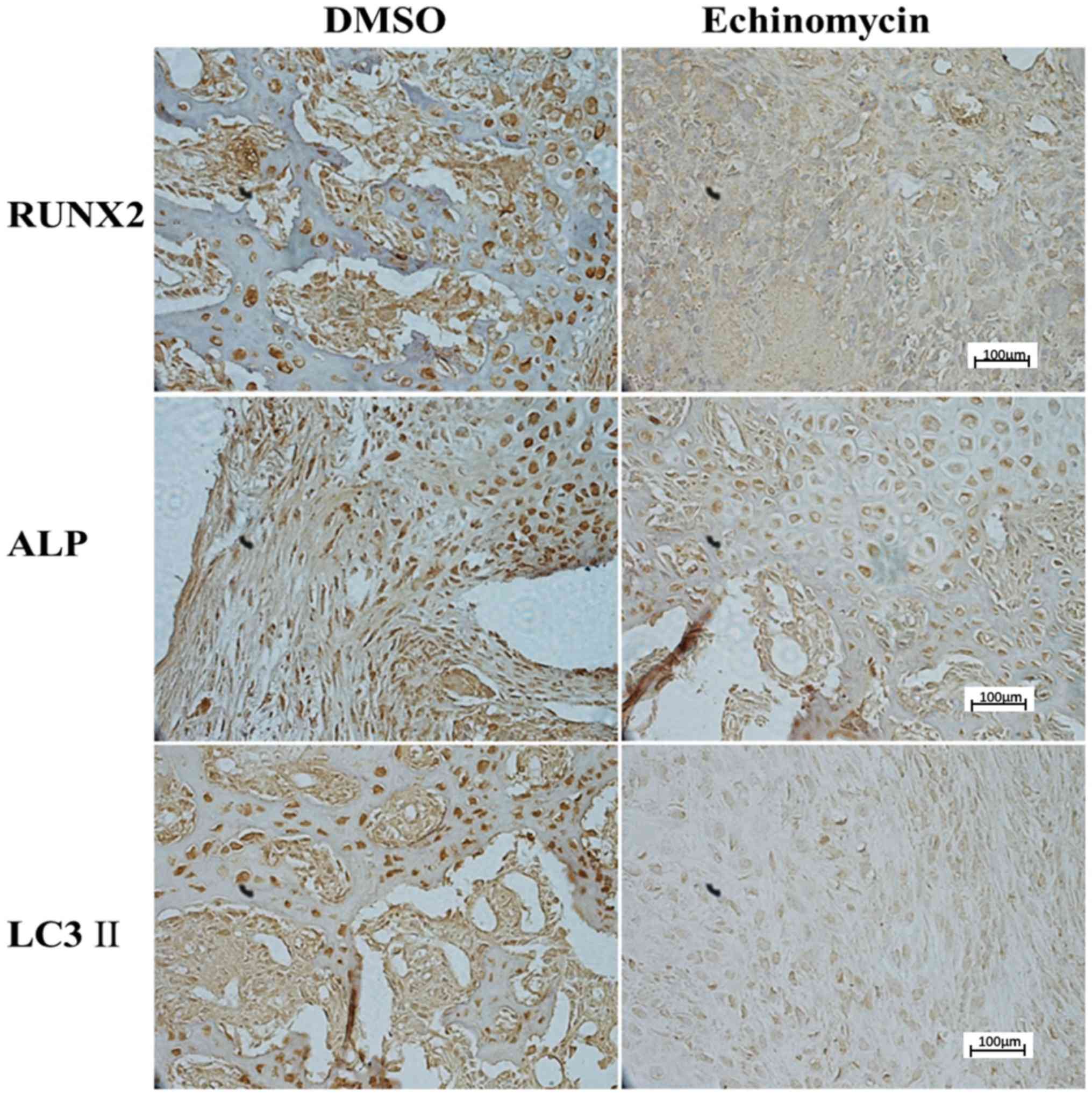
Echinomycin decreases the expression
of HIF-1α target genes and autophagy markers at the protein
level
Protein levels of HIF-1α target genes and molecular
markers of autophagy were further evaluated by IHC and western blot
analysis. As indicated in Fig. 3,
rats with echinomycin administration exhibited a significant
downregulation of Runx2, ALP and LC3 II expression at day 42 after
fracture. In addition, western blot analysis demonstrated that
echinomycin treatment led to the significantly decreased expression
of Runx2, ALP, ULK1 and slightly decreased the expression of LC3
II. The expression of P62 was also increased (Figs. 4 and 5). These results were consistent with those
at the mRNA level, which further suggested a potential role of
HIF-1α in fracture healing.
 | Figure 4.Western blot analysis of HIF-1α, ULK1,
P62, LC3 II, Runx2 and ALP at the indicated time points after
fracture. HIF-1α, hypoxia-inducible factor-1α; ULK1, Unc-51-like
autophagy activating kinase 1; LC3 II, microtubule-associated
protein 1 light chain 3; Runx2, runt-related transcription factor
2; ALP, alkaline phosphatase; DMSO, dimethyl sulfoxide. |
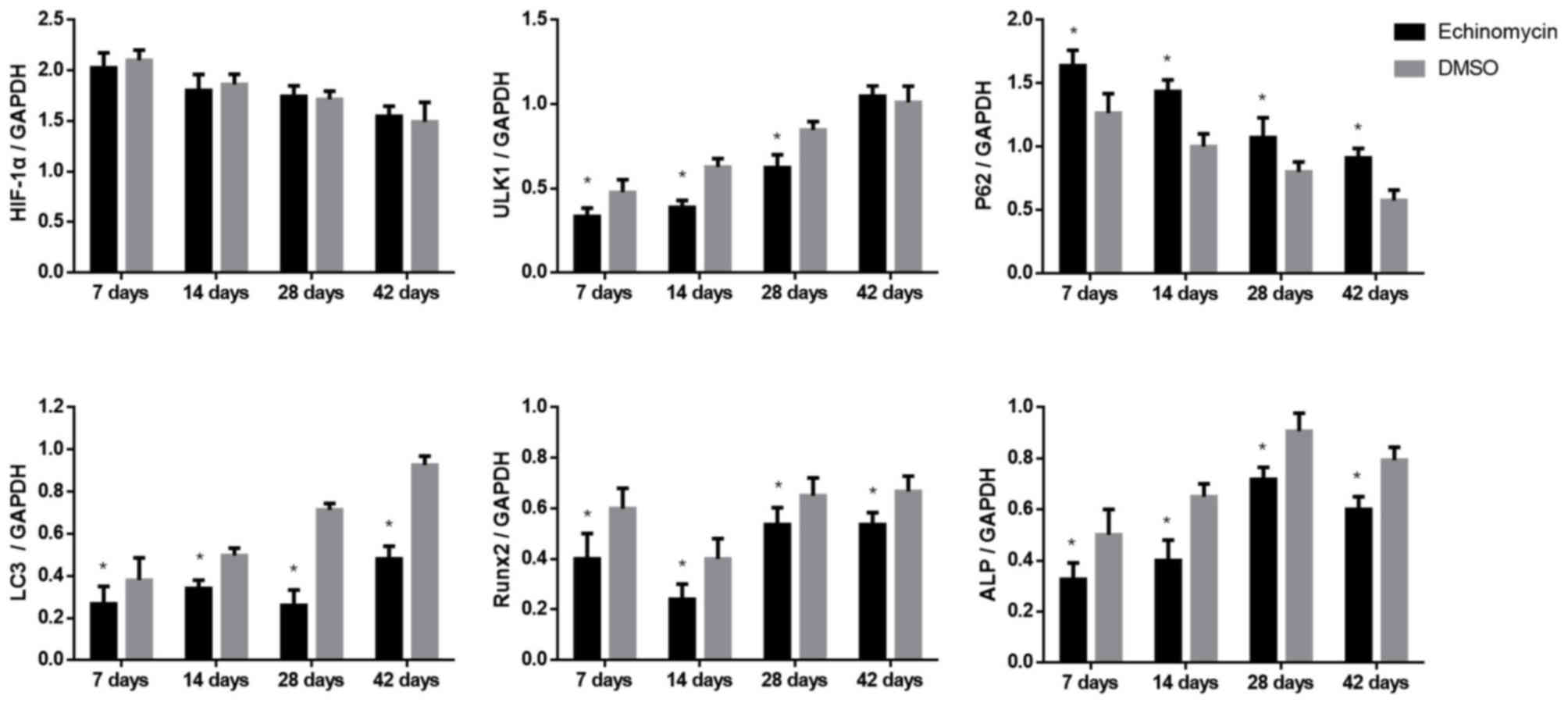 | Figure 5.Quantitative analysis of HIF-1α, ULK1,
P62, LC3 II, Runx2 and ALP protein expression at the indicated time
points after fracture. *P<0.01 vs. DMSO-treated group. HIF-1α,
hypoxia-inducible factor-1α; ULK1, Unc-51-like autophagy activating
kinase 1; LC3 II, microtubule-associated protein 1 light chain 3;
Runx2, runt-related transcription factor 2; ALP, alkaline
phosphatase; DMSO, dimethyl sulfoxide. |
Discussion
Fracture repair is a highly complex but
well-orchestrated regenerative process involving specific gene
expression and cellular recruitment (16). Fractures lead to inadequate blood
supply and decreased O2 availability, which results in
the upregulation of HIF-1α (17).
HIF-1α, an oxygen-sensitive transcription factor, mediates cellular
responses to hypoxia during pathological processes in many
diseases. Previous studies have demonstrated that an increase in
HIF-1α and its downstream cytokines is apparent in the process of
fracture healing (7,17).
When HIF-1α interacts with the hypoxia response
element (HRE) DNA sequence of a target gene, numerous
hypoxia-sensitive genes are upregulated, including VEGF, heme
oxygenase 1 and inducible nitric oxide synthase (18). Of note, it has been reported that
HIF-1α could enhance the expression of Runx2, which participates in
the process of osteogenesis in vivo (7). Runx2 is a key regulator of bone
formation via the regulation of osteoclast and osteoblast
differentiation. By regulating the cell cycle of osteoblasts and
the expression of specific extracellular matrix protein genes in
osteoblasts, Runx2 can promote bone formation and inhibit bone
resorption (19). As a consequence,
HIF-1α is considered to aid in accelerating fracture healing
(7). Echinomycin is an established
inhibitor of HIF-1α, which acts by inhibiting its DNA-binding
activity (20). In the current
study, it was hypothesized that echinomycin may have an inhibitory
role on fracture healing by downregulating the target genes of
HIF-1α.
In this study, the expression of HIF-1α was detected
at the mRNA and protein levels in fracture calluses. It was
identified that the levels of HIF-1α did not differ significantly
between the echinomycin and control groups throughout the fracture
healing process, which was consistent with a previous study
(21). However, Runx2 and ALP were
significantly downregulated in the echinomycin group. A possible
interpretation is that an increase of HIF-1α was induced by the
condition of hypoxia caused by fracture, which was the same in both
groups, and echinomycin inhibited HIF-1α by affecting the
DNA-binding activity of HIF-1α with its target genes, and not via
affecting the expression and transcription of HIF-1α. Furthermore,
micro-CT of the tibia demonstrated that the density and the amount
of callus tissue between the echinomycin and control groups were
different. The volume of callus in the echinomycin group was
significantly increased compared with in the control group during
the callus formation stage, and BMD was notably increased in the
control group (data not shown). In summary, these results support
the proposal that administration with echinomycin may delay the
fracture healing by causing downregulating of the HIF-1α target
genes.
In recent years, the study of autophagy in bone
research has been an emerging field. Autophagy is a catabolic
process characterized by removal of defective organelles and
mobilization of nutrients in order to maintain energy balance
(22). Increasing evidence has
demonstrated that energy status and nutrient levels are key
regulatory factors of autophagy (23). Once the physiological balance is
disrupted, autophagy may be triggered in cells to enable a response
to the changing conditions (23,24).
Previous studies have demonstrated that autophagy is an
HIF-1α-dependent adaptive metabolic response to hypoxia (25–27).
However, it had thus far remained unclear how autophagy is altered
if HIF-1α is inhibited.
The present study provided experimental data to
suggest that HIF-1α may serve a critical role in the regulation of
autophagy during the process of fracture healing. LC3 serves a
critical role in the formation and elongation of the autophagosome
and is among the most studied proteins in monitoring autophagic
activity (28). In the current
study, the levels of LC3 II were slightly downregulated in the
echinomycin-treated group compared with in the control group. As
previously described, ULK1 is enzymatically activated in response
to nutrient deprivation and may phosphorylate autophagy-related
protein 13 to initiate the process of autophagy (29). A disruption of blood supply by
fracture may generate hypoxic conditions around the damaged area
and promote anaerobic metabolism, which may give rise to
insufficient nutrients and energy (2). Therefore, it is inferred that autophagy
following fracture may be an energy-dependent process and regulated
by the HIF-1α pathway (23). P62, a
multifunctional ubiquitin-binding protein, has been proposed to be
involved in the degradation of target proteins and cytoplasmic
bodies by lysosomal enzymes (30).
When also considering the expression data on P62, which were the
opposite to that of LC3 II and ULK1, the current results strongly
indicated that autophagy was inhibited during the process of
fracture healing in the echinomycin-treated group. The results were
consistent with the view that autophagy is an important process
utilized by HIF-1α to maintain cell homeostasis (22). Combined with other recent studies
(26,31), these results support the conclusion
that HIF-1α and autophagy are linked. Although the association
between HIF-1α and autophagy requires further investigation, this
suggests that maintenance of HIF-1α and autophagy may serve a
functional role in bone regeneration in fracture healing.
In conclusion, the present results demonstrate that
HIF-1α, which serves an important role in fracture healing, may
serve as a target to improve bone healing. Furthermore, the
autophagy induced by fracture may be HIF-1α-dependent and HIF-1α
may also affect the healing process by regulating autophagy. These
findings highlight the role of HIF-1α and provide a potential
strategy for the regeneration of bone tissue. However, determining
the factor(s) utilized by HIF-1α in the process of fracture healing
remains a topic of further study. Future studies should investigate
the regulatory mechanisms associated with HIF-1α and autophagy in
more depth, in order to facilitate the development of treatments
aimed at improving fracture healing.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81541135) and
Capital's Funds for Health Improvement and Research (grant no.
2018-2-2012).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JQ performed the majority of the experiments and
prepared the manuscript. JH performed the polymerase chain reaction
and western blot analyses. MZ analyzed the data. GC and HS designed
the experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee on Animal Experimentation of Capital Medical University,
Beijing, China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HIF-1α
|
hypoxia-inducible factor-1α
|
|
DMSO
|
dimethyl sulfoxide
|
|
ROI
|
region of interest
|
|
BV/TV
|
bone volume/total volume
|
|
BMD
|
bone mineral density
|
|
ALP
|
alkaline phosphatase
|
|
Runx2
|
runt-related transcription factor
2
|
|
VEGF
|
vascular endothelial growth factor
|
|
LC3 II
|
microtubule-associated protein 1 light
chain 3
|
|
ULK1
|
Unc-51-like autophagy activating
kinase 1
|
|
micro-CT
|
micro-computed tomography
|
|
IHC
|
immunohistochemistry
|
|
qPCR
|
quantitative polymerase chain
reaction
|
|
PBS
|
phosphate-buffered saline
|
References
|
1
|
Einhorn TA and Gerstenfeld LC: Fracture
healing: Mechanisms and interventions. Nat Rev Rheumatol. 11:45–54.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Giannoudis PV, Jones E and Einhorn TA:
Fracture healing and bone repair. Injury. 42:549–550. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oryan A, Monazzah S and Bigham-Sadegh A:
Bone injury and fracture healing biology. Biomed Environ Sci.
28:57–71. 2015.PubMed/NCBI
|
|
4
|
Sang X, Wang Z, Qin T and Li Y: Elevated
concentrations of hypoxia-inducible factor-1α in patients with
fracture and concomitant traumatic brain injury. Ann Clin Biochem.
54:584–592. 2017.PubMed/NCBI
|
|
5
|
Liu P, Liu J, Xia K, Chen L and Wu X:
Effect of leptin combined with CoCl2 on healing in Sprague Dawley
Rat fracture model. Sci Rep. 6:307542016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun G and Peng H: HIF-1α-induced
microRNA-210 reduces hypoxia-induced osteoblast MG-63 cell
apoptosis. Biosci Biotechnol Biochem. 79:1232–1239. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang J, Liu L, Feng M, An S, Zhou M, Li
Z, Qi J and Shen H: Effect of CoCl(2) on fracture repair in a rat
model of bone fracture. Mol Med Rep. 12:5951–5956. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schorn L, Sproll C, Ommerborn M, Naujoks
C, Kubler NR and Depprich R: Vertical bone regeneration using
rhBMP-2 and VEGF. Head Face Med. 13:112017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pierrefite-Carle V, Santucci-Darmanin S,
Breuil V, Camuzard O and Carle GF: Autophagy in bone: Self-eating
to stay in balance. Ageing Res Rev. 24:206–217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Singh SR, Zech ATL, Geertz B,
Reischmann-Dusener S, Osinska H, Prondzynski M, Kramer E, Meng Q,
Redwood C, van der Velden J, et al: Activation of autophagy
ameliorates cardiomyopathy in mybpc3-targeted knockin mice. Circ
Heart Fail. 10:e0041402017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang GE, Duan X, Lin D, Li T, Luo D, Wang
L and Lian K: Rapamycin-induced autophagy activity promotes bone
fracture healing in rats. Exp Ther Med. 10:1327–1333. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Q, Luo D, Li T, Liu Z, Zou W, Wang L,
Lin D and Lian K: Bone fracture in a rat femoral fracture model is
associated with the activation of autophagy. Exp Ther Med.
10:1675–1680. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bouxsein ML, Boyd SK, Christiansen BA,
Guldberg RE, Jepsen KJ and Müller R: Guidelines for assessment of
bone microstructure in rodents using micro-computed tomography. J
Bone Miner Res. 25:1468–1486. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gerstenfeld LC, Wronski TJ, Hollinger JO
and Einhorn TA: Application of histomorphometric methods to the
study of bone repair. J Bone Miner Res. 20:1715–1722. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Loi F, Córdova LA, Pajarinen J, Lin TH,
Yao Z and Goodman SB: Inflammation, fracture and bone repair. Bone.
86:119–130. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li W, Wang K, Liu Z and Ding W: HIF-1α
change in serum and callus during fracture healing in
ovariectomized mice. Int J Clin Exp Pathol. 8:117–126.
2015.PubMed/NCBI
|
|
18
|
Komatsu DE and Hadjiargyrou M: Activation
of the transcription factor HIF-1 and its target genes, VEGF, HO-1,
iNOS, during fracture repair. Bone. 34:680–688. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Perez-Campo FM, Santurtun A,
Garcia-Ibarbia C, Pascual MA, Valero C, Garces C, Sanudo C,
Zarrabeitia MT and Riancho JA: Osterix and RUNX2 are
transcriptional regulators of sclerostin in human bone. Calcif
Tissue Int. 99:302–309. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kong D, Park EJ, Stephen AG, Calvani M,
Cardellina JH, Monks A, Fisher RJ, Shoemaker RH and Melillo G:
Echinomycin, a small-molecule inhibitor of hypoxia-inducible
factor-1 DNA-binding activity. Cancer Res. 65:9047–9055. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Z, Zhang Z, Wu Y, Chen L, Luo Q,
Zhang J, Chen J, Luo Z, Huang X and Cheng Y: Effects of echinomycin
on endothelin-2 expression and ovulation in immature rats primed
with gonadotropins. Exp Mol Med. 44:615–621. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang H, Bosch-Marce M, Shimoda LA, Tan
YS, Baek JH, Wesley JB, Gonzalez FJ and Semenza GL: Mitochondrial
autophagy is an HIF-1-dependent adaptive metabolic response to
hypoxia. J Biol Chem. 283:10892–10903. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Parzych KR and Klionsky DJ: An overview of
autophagy: Morphology, mechanism, and regulation. Antioxid Redox
Signal. 20:460–473. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kroemer G: Autophagy: A druggable process
that is deregulated in aging and human disease. J Clin Invest.
125:1–4. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tsuzuki T, Okada H, Cho H, Tsuji S,
Nishigaki A, Yasuda K and Kanzaki H: Hypoxic stress simultaneously
stimulates vascular endothelial growth factor via hypoxia-inducible
factor-1 and inhibits stromal cell-derived factor-1 in human
endometrial stromal cells. Hum Reprod. 27:523–530. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu H, Huang S, Chen Z, Liu W, Zhou X and
Zhang D: Hypoxia-induced autophagy contributes to the invasion of
salivary adenoid cystic carcinoma through the HIF-1α/BNIP3
signaling pathway. Mol Med Rep. 12:6467–6474. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shelby SJ, Angadi PS, Zheng QD, Yao J, Jia
L and Zacks DN: Hypoxia inducible factor 1α contributes to
regulation of autophagy in retinal detachment. Exp Eye Res.
137:84–93. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang R, Xu Y, Wan W, Shou X, Qian J, You
Z, Liu B, Chang C, Zhou T, Lippincott-Schwartz J and Liu W:
Deacetylation of nuclear LC3 drives autophagy initiation under
starvation. Mol Cell. 57:456–466. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nazio F, Strappazzon F, Antonioli M,
Bielli P, Cianfanelli V, Bordi M, Gretzmeier C, Dengjel J,
Piacentini M, Fimia GM and Cecconi F: mTOR inhibits autophagy by
controlling ULK1 ubiquitylation, self-association and function
through AMBRA1 and TRAF6. Nat Cell Biol. 15:406–416. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lin X, Li S, Zhao Y, Ma X, Zhang K, He X
and Wang Z: Interaction domains of p62: A bridge between p62 and
selective autophagy. DNA Cell Biol. 32:220–227. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu H, Zhang Z, Xiong W, Zhang L, Xiong Y,
Li N, He H, Du Y and Liu Y: Hypoxia-inducible factor-1α promotes
endometrial stromal cells migration and invasion by upregulating
autophagy in endometriosis. Reproduction. 153:809–820. 2017.
View Article : Google Scholar : PubMed/NCBI
|