Introduction
Bladder cancer (BC) is the fourth most prevalent
solid tumor type in males and the seventh most prevalent in females
worldwide (1). It is accountable for
~3% of cancer-associated deaths (2).
Although certain therapeutic methods, including radiotherapy,
surgery and chemotherapy, have been developed for BC treatment, the
recurrence rate remains high (2,3) and the
prognosis of BC patients is poor (4). Therefore, it is urgently required to
explore the regulatory mechanisms underlying the occurrence of BC,
which will contribute to the identification of therapeutic targets
and the development of novel treatments.
MicroRNAs (miRNAs/miRs) are a class of non-coding
RNAs and post-transcriptionally regulate gene expression by
recognizing the complementary sequence in the 3′ untranslated
region (3′-UTR) of their target mRNAs (5,6). miRNAs
exert vital functions in a broad variety of biological processes,
including development, cell proliferation and apoptosis (7). Dysregulated expression of miRNAs is
usually observed in almost all cancer types, including colorectal
(8), liver (9) and bladder cancer (4). Increasing evidence indicates that
certain miRNAs may serve as oncogenes or tumor suppressors to
regulate BC development and progression (4,10). For
instance, Yuan et al (11)
reported that miR-124-3p inhibits the growth and metastasis of BC
by degrading the mRNA of aurora kinase A. Furthermore, Feng et
al (12) indicated that
miR-556-3p contributes to BC cell proliferation and invasiveness
through inhibiting DAB2 interacting protein expression. Another
previous study indicated that miR-758-3p inhibits hepatocellular
carcinoma progression (13).
miR-758-3p is also implicated in cervical cancer (14). However, the biological functions of
miR-758-3p in BC have not been previously reported. Due to the
significance of miR-758-3p in the abovementioned cancer types, the
present study sought to investigate the function and potential
mechanisms of miR-758-3p in BC.
In the present study, it was demonstrated that
miR-758-3p expression was downregulated in BC tissues and cell
lines. Furthermore, transfection with miR-758-3p mimics markedly
repressed the proliferation, migration and invasion of BC cells. It
was also revealed that Notch receptor 2 (NOTCH2) was a direct
target of miR-758-3p. In summary, the present study illustrated
that miR-758-3p inhibits BC progression via targeting NOTCH2,
suggesting that miR-758-3p may be a promising therapeutic target
for BC treatment.
Materials and methods
Human tissues
A total of 33 BC tissues (age range, 61±8.1 years;
female, n=4; male, n=29) and matched normal tissues (at least 3 cm
away from the tumor border and with no microscopic evidence of
tumor cells) were collected from patients diagnosed with BC at the
Xiangyang Central Hospital (Xiangyang, China) from January 2014 to
September 2016. All patients provided written informed consent.
Samples from patients who received radiotherapy or chemotherapy
prior to surgery were excluded. The tissues were stored in liquid
nitrogen at −80°C until use. The clinicopathological
characteristics of the 33 patients with BC were also recorded. The
present study was approved by the Ethics Committee of Xiangyang
Central Hospital (Xiangyang, China).
Cell culture and transfection
The J82, UMUC3, T24 and 5637 BC cell lines as well
as the SV-HUC-1 normal bladder cell line were obtained from the
Cell Bank of the Chinese Academy of Sciences (Shanghai, China).
Cells were maintained in RPMI-1640 medium (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) containing 10% fetal
bovine serum (FBS; Invitrogen; Thermo Fisher Scientific, Inc.) and
1% penicillin/streptomycin.
For cell transfection, the miR-758-3p mimics
(5′-UUUGUGACCUGGUCCACUAACC-3′), miR-758-3p inhibitor
(5′-GGUUAGUGGACCAGGUCACAAA-3′), inhibitor control
(5-GCGUAACUAAUACAUCGGAUUCGU-3) and mimic control
(5′-ACAUCUGCGUAAGAUUCGAGUCUA-3′) were purchased from GenePharma
(Shanghai, China). Cells were transfected with miR-758-3p mimics or
controls using Lipofectamine 2000™ (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. For
NOTCH2 overexpression, the sequence encoding the NOTCH2
intracellular segment was inserted into the pcDNA3 vector to
generate pcDNA3-NOTCH2. Then pcDNA3-NOTCH2 vector (1 µg) was
transfected into BCa cell lines using Lipofectamine 2000™
(Invitrogen; Thermo Fisher Scientific, Inc.). After 48 h, the
overexpression efficiency was evaluated and gene expression was
determined using reverse transcription-quantitative polymerase
chain reaction.
RT-qPCR
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to extract total RNA from cells. Total
RNA (1 µg) was reverse transcribed into cDNA using the PrimeScript™
RT reagent kit (Takara Biotechnology Co., Ltd., Dalian, China),
according to the manufacturer's protocol. qPCR was subsequently
performed using the SYBR Green I Supermix (Takara Biotechnology
Co., Ltd.), according to the manufacturer's protocol using an
iCycler IQ multicolor Detection System (Bio-Rad Laboratories,
Hercules, CA, USA). The following thermocycling conditions were
used for the qPCR: Initial denaturation at 95°C for 10 min; 40
cycles of 95°C for 15 sec and 60°C for 1 min. The primer pairs used
were as follows: miR-758-3p forward,
5′-ACACTCCAGCTGGGTTTGTGACCTGGTCCA-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGGTTAGTG-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′;
NOTCH2 forward, 5′-CAAGGAACCTGCTTTGATGACA-3′ and reverse,
5′-GGGGAACAGGGAGCCAATAC-3′; and GAPDH forward,
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse, 5′-TGGTGAAGACGCCAGTGGA-3′.
The mRNA levels were quantified using the 2−∆∆Cq method
and U6 was used as a normalization control (15).
Cell Counting Kit (CCK)-8
proliferation assay
Cell proliferation was measured using a CCK-8
proliferation assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Cells were seeded into 96-well plates at a
density of 2×103 cells/well) and cultured for the
indicated durations. Following the addition of 10 µl CCK-8 reagent,
the plates were incubated for 1 h at 37°C. Subsequently, the
absorbance at 450 nm was determined using a microplate reader
(Berthold Technologies GmbH, Bad Wildbad, Germany).
Colony formation assay
Cells were seeded into 6-well plates at
1×103 cells/well and cultured for 12 days. The colonies
were fixed using methanol for 15 min at room temperature, stained
using 0.5% crystal violet for 20 min at room temperature. The total
number of visible colonies was examined under an optical light
microscope (magnification, ×40; Olympus Corporation, Tokyo,
Japan).
Migration and invasion assays
Transwell chambers (BD Biosciences, Franklin Lakes,
NJ, USA) were used for Transwell migration and invasion assays.
Cells (5×104) in 100 µl serum-free medium were seeded
into the upper chamber [pre-coated with Matrigel® (1:6
dilution; BD Biosciences) for the invasion assay]. The lower
chamber was filled with 600 µl medium containing 10% FBS. Following
24-h incubation, cells that had migrated to the lower side of the
membrane were fixed with polyoxymethylene at room temperature for
30 min and stained with 0.5% crystal violet at room temperature for
30 min. Images of the cells were captured under an optical
microscope.
Western blot analysis
Total protein was extracted from cells using
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific,
Inc.). Total protein was quantified using a bicinchoninic acid
assay and 40 µg protein/lane was separated via SDS-PAGE on a 12%
gel. The separated proteins were transferred onto polyvinylidene
fluoride membranes (Thermo Fisher Scientific, Inc.) and blocked for
3 h at room temperature with 5% non-fat milk in PBS (Thermo Fisher
Scientific, Inc.) containing 0.1% Tween-20 (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). The membranes were incubated with the
following primary antibodies: Anti-NOTCH2 (1:1,500; cat. no. 5732)
and mouse anti-GAPDH (1:5,000; cat. no. 5174; both Cell Signaling
Technology, Danvers, MA, USA) overnight at 4°C. Subsequently, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibodies (1:5,000; cat. no. ab7090; Abcam, Cambridge,
MA, USA) for 1 h at room temperature. Protein bands were visualized
using the Pierce™ ECL Western Blotting Substrate (Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Protein expression was quantified using ImageJ software (version
1.41; National Institutes of Health, Bethesda, MD, USA).
Luciferase assay
The potential binding site of NOTCH2 3′-UTR for
miR-758-3p was predicted using the TargetScan7 tool (http://www.targetscan.org/vert_71/). The
sequences containing the wild-type (WT) or site-mutated (Mut)
region of NOTCH2 were synthesized by Sangon (Shanghai, China) and
inserted into the pGL3 vector (Promega Corporation, Madison, WI,
USA). For the luciferase reporter assay, miR-758-3p or NC mimics
and the respective reporter plasmids were transfected into BC cells
using Lipofectamine 2000™ (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. After 24 h, the
Renilla and firefly luciferase activity was determined using
the Dual-Luciferase Reporter Assay System (Promega Corp.) according
to the manufacturer's protocols and a luminometer (Infinite 200 PRO
NanoQuant; Tecan Group Ltd., Männedorf, Switzerland).
Statistical analysis
Statistical analysis was performed using SPSS 20.0
(IBM Corp., Armonk, NY, USA) or GraphPad Prism version 5 (GraphPad
Software, Inc., LA Jolla, CA, USA). The assays were performed as
three independent replicates. Values are expressed as the mean ±
standard deviation. P-values were calculated using Student's t-test
or one-way analysis of variance followed by Tukey's post hoc test.
The association between miR-758-3p expression and the
clinicopathological characteristics of patients with BC was
analyzed using the Chi-square test. Spearman's rank correlation
analysis was performed to analyze the correlation between
miR-758-3p and NOTCH2 expression levels. P<0.05 was considered
to indicate statistical significance.
Results
miR-758-3p is downregulated in BC
tissues and cell lines
To investigate the function of miR-758-3p in BC, its
expression was analyzed in tumor tissues and adjacent normal
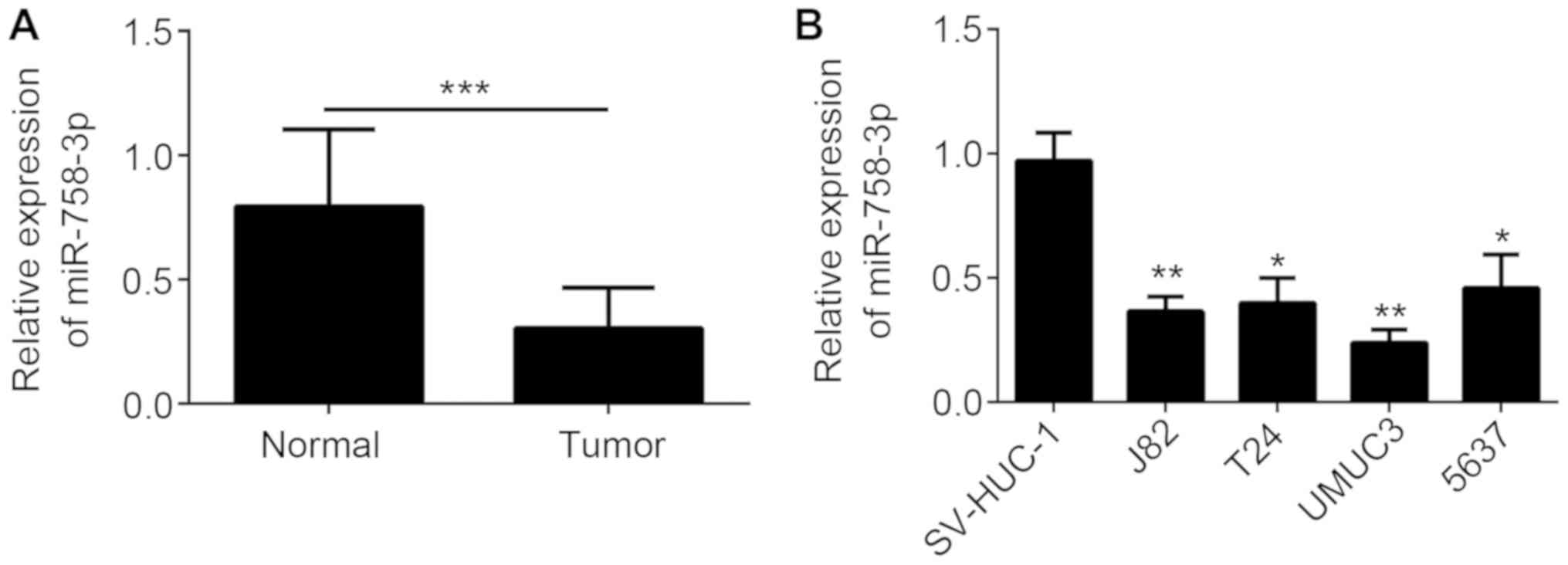
tissues of 33 BC patients. As presented Fig. 1A, miR-758-3p was downregulated in BC
tissues compared with that in the matched normal tissues. In
addition, miR-758-3p expression was downregulated in BC cell lines
compared with that in the SV-HUC-1 normal bladder cell line
(Fig. 1B). The association between
miR-758-3p expression and the clinicopathological characteristics
of patients with BC was examined (Table
I).
 | Table I.Association between miR-758-3p
expression and clinicopathological characteristics of patients with
bladder cancer (n=33). |
Table I.
Association between miR-758-3p
expression and clinicopathological characteristics of patients with
bladder cancer (n=33).
|
| miR-758-3p
expression |
|
|---|
|
|
|
|
|---|
| Clinicopathological
characteristic | Low (n=18) | High (n=15) | P-value |
|---|
| Age (years) |
|
| 0.169 |
|
<60 | 5 | 8 |
|
| ≥60 | 13 | 7 |
|
| Sex |
|
| 0.607 |
|
Female | 3 | 1 |
|
| Male | 15 | 14 |
|
| TNM stage |
|
| 0.038 |
| I/II | 4 | 9 |
|
|
III/IV | 14 | 6 |
|
| Lymph node
metastasis |
|
| 0.034 |
| Yes | 9 | 13 |
|
| No | 9 | 2 |
|
miR-758-3p suppresses the
proliferation, migration and invasion of BC cells
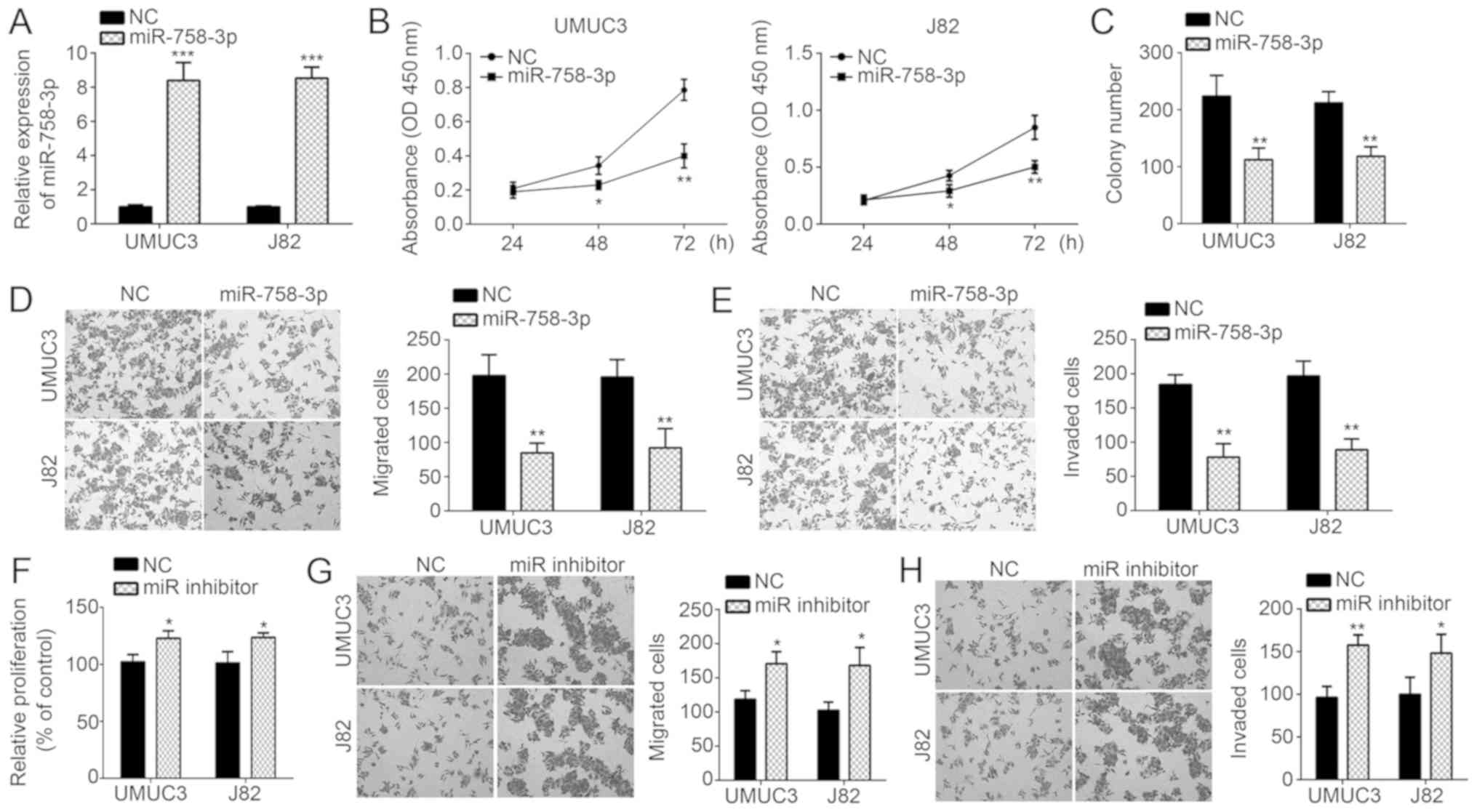
To explore the role of miR-758-3p in BC, miR-758-3p
mimics were transfected into UMUC3 and J82 cells. RT-qPCR analysis
confirmed that miR-758-3p levels were markedly increased in UMUC3
and J82 cells after transfection (Fig.
2A). CCK-8 and colony formation assays were then performed to
evaluate the cell proliferation ability. The results indicated that
miR-758-3p overexpression inhibited the proliferation and colony
formation of UMUC3 and J82 cells (Fig.
2B and C). Furthermore, as indicated by the Transwell assay,
transfection of miR-758-3p mimics into UMUC3 and J82 cells markedly
inhibited migration and invasion (Fig.
2D and E). In addition, to further validate the function of
miR-758-3p, UMUC3 and J82 cells were transduced with miR-758-3p
inhibitor. Through CCK-8 and Transwell assays, it was revealed that
miR-758-3p inhibition significantly promoted proliferation,
migration and invasion (Fig. 2F-H).
Taken together, miR-758-3p suppresses BC proliferation and
progression.
NOTCH2 is a target of miR-758-3p in BC
cells
To further determine the mechanisms of miR-758-3p in
BC, the downstream target genes of miR-758-3p were searched with
TargetScan software. Among all candidates of predicted potential
targets of miR-758-3p, NOTCH2 ranked high and was previously
reported to promote BC progression (16). Thus, NOTCH2 was selected for further
investigation. There was a complementary sequence of miR-758-3p in
the 3′-UTR region of NOTCH2 mRNA (Fig.
3A). To confirm the direct binding interaction in vitro,
WT and Mut luciferase reporter plasmids were constructed and used
in a luciferase reporter assay. The results demonstrated that
miR-758-3p mimics inhibited the luciferase intensity of the
NOTCH2-WT reporter plasmid in UMUC3 and J82 cells, while mutation
of the complementary binding site abrogated this effect (Fig. 3B). In a further experiment,
miR-758-3p mimics markedly decreased NOTCH2 expression in UMUC3 and
J82 cells (Fig. 3C and D). In
addition, the expression of NOTCH2 was examined in BC tissues,
revealing an inverse association between the expression of
miR-758-3p and NOTCH2 (Fig. 3E).
Furthermore, NOTCH2 levels were determined in BC tissues by RT-qPCR
and western blot analysis, demonstrating that NOTCH2 expression was
significantly upregulated in BC tissues compared with that in
adjacent normal tissues (Fig. 3F and
G).
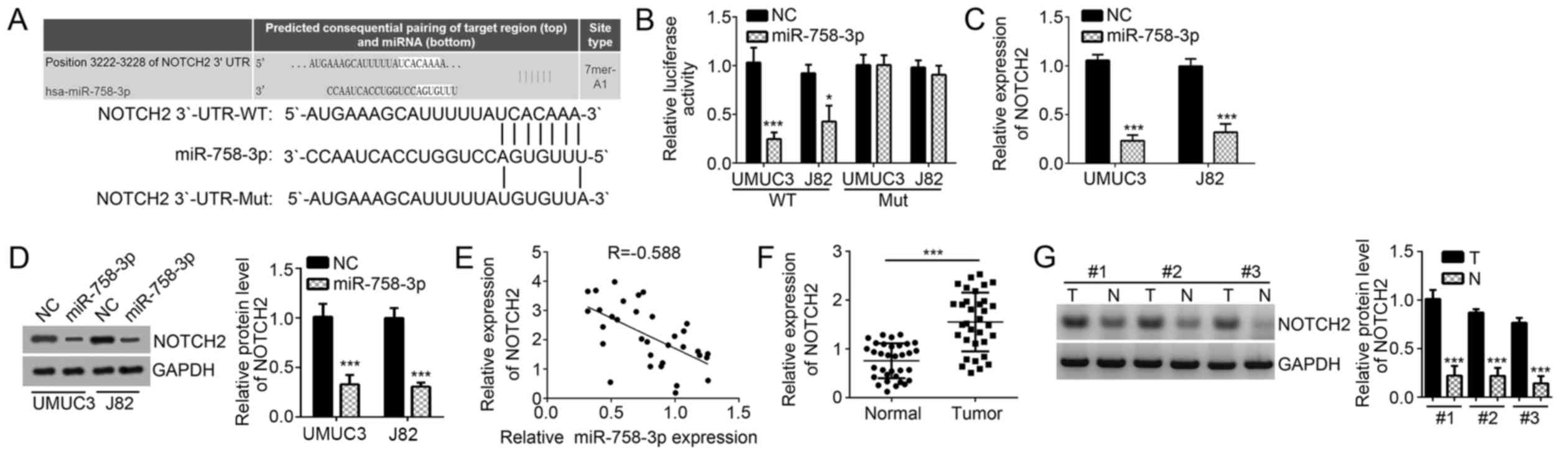 | Figure 3.NOTCH2 is a target of miR-758-3p in BC
cells. (A) The predicted complementary site in the 3′-UTR of NOTCH2
with miR-758-3p. (B) A luciferase reporter assay in UMUC3 and J82
cells indicated that miR-758-3p mimics inhibited the luciferase
activity of the reporter plasmid carrying the WT fragment from the
3′-UTR of NOTCH2 but not the mutant 3′-UTR fragment. (C and D)
miR-758-3p mimics reduced the mRNA and protein levels of NOTCH2 in
UMUC3 and J82 cells, as indicated by RT-qPCR and western blot
analysis, respectively. (E) The correlation between NOTCH2 mRNA and
miR-758-3p expression in BC tissues from 33 cases was determined by
Spearman's correlation analysis. (F and G) mRNA and protein levels
of NOTCH2 in BC tissues and adjacent normal tissues were measured
by RT-qPCR and western blot analysis, respectively. *P<0.05;
***P<0.001 vs. control group. UTR, untranslated region; WT,
wild-type; miR, microRNA; Mut, mutated; NC, negative control; Hsa,
Homo sapiens; T, tumor tissue; N, normal tissue; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction; BC,
bladder cancer; NOTCH2, Notch receptor 2. |
miR-758-3p suppresses BC cell
proliferation, migration and invasion through targeting NOTCH2
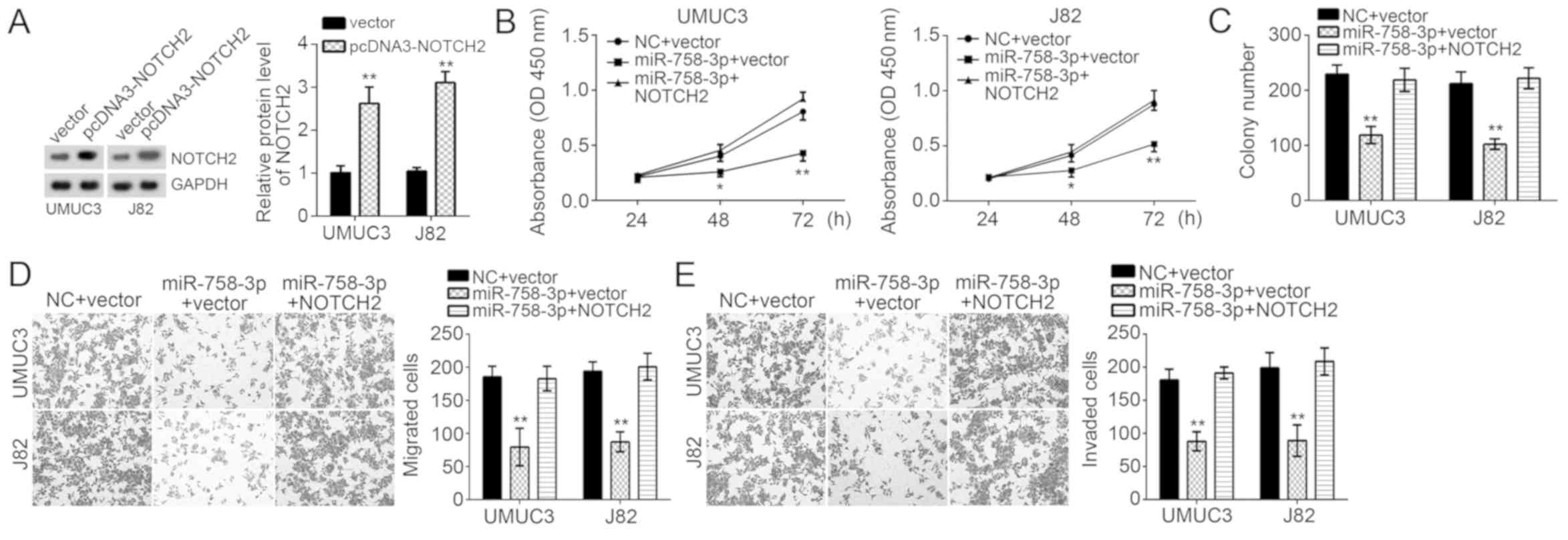
To determine whether suppression of cell
proliferation, migration and invasion by miR-758-3p relies on
NOTCH2, UMUC3 and J82 cells transduced with miR-758-3p mimics were
subjected to ectopic overexpression of NOTCH2. Western blot
analysis confirmed that the levels of NOTCH2, which were decreased
by miR-758-3p mimics, were restored by co-transfection with NOTCH2
overexpression vector (Fig. 4A).
Functional experiments indicated that restoration of NOTCH2
promoted the proliferation and colony formation ability of UMUC3
and J82 cells transfected with miR-758-3p mimics (Fig. 4B and C). Furthermore, overexpression
of NOTCH2 also rescued the migration and invasion of UMUC3 and J82
cells transfected with miR-758-3p mimics (Fig. 4D and E). In conclusion, miR-758-3p
inhibited BC cell proliferation, migration and invasion at least in
part through targeting the mRNA of NOTCH2 and promoting its
degradation.
Discussion
BC has become the most common malignancy of the
urinary tract, originating from bladder mucosa, worldwide (17). Each year, there are large numbers of
BC cases and increasing BC-related mortality rates (17). Thus, it is vital to reveal the
underlying mechanisms of the genesis and progression of BC and
develop effective therapeutic methods. Accumulating evidence
indicates that miRNAs are potential biomarkers for diagnosis and
prognosis in numerous cancer types (18,19). For
instance, miR-122 and miR-224 have been reported to serve as
biomarkers for early diagnosis of hepatocellular carcinoma
(20). The present study
demonstrated that miR-758-3p has a tumor suppressor function in BC
and therefore miR-758-3p may be a promising therapeutic target.
In the past decades, miRNAs have attracted wide
attention, and a vast number of studies have demonstrated their
essential and general functions in a diversity of biological
processes, including cell migration, proliferation and invasion
(21,22). For instance, Ding et al
(23) reported that miR-367
suppresses clear-cell renal cell cancer progression. Wu et
al (24) reported that miR-21
contributes to colorectal cancer progression via targeting
phosphatase and tensin homolog. Another previous study indicated
that miR-758-3p suppresses liver cancer development by suppressing
MDM2 and mammalian target of rapamycin (13). In cervical cancer patients, the
levels of miR-758 were reported to be decreased in the tumor
tissues, blood and cervical exfoliated cells, and it was indicated
that miR-758 may regulate the infiltration and invasion of cervical
cancer by targeting matrix extracellular phosphoglycoprotein
(14). These studies suggest a tumor
suppressor role for miR-758-3p. However, the effect of miR-758-3p
in BC has remained elusive. The present results indicated that
miR-758-3p was downregulated in BC tissues compared with that in
matched normal tissues. Furthermore, CCK-8, colony formation and
Transwell assays suggested that transfection with miR-758-3p mimics
markedly inhibited the malignant behavior of BC cells. In addition,
NOTCH2 was identified as a direct target of miR-758-3p in BC
cells.
NOTCH2, a member of the NOTCH family, has a role in
developmental processes. NOTHC2 signaling is evolutionarily
conserved and is involved in cell fate decisions (25). Increasing evidence has indicated that
NOTCH signaling is involved in the development and progression of
numerous human cancer types, including BC (16,26).
Furthermore, a recent review also indicated that NOTCH2 acts as an
oncogene that promotes cell proliferation and metastasis through
epithelial-to-mesenchymal transition, cell cycle progression and
maintenance of stem cells in BC (27). In the present study, NOTCH2 was
identified to be downregulated by miR-758-3p in BC cells.
Furthermore, the expression of NOTCH2 was negatively correlated
with that of miR-758-3p in BC tissues. Notably, restoration of
NOTCH2 reversed the effects of miR-758-3p mimics on BC cell
proliferation, migration and invasion.
In conclusion, the present study indicated a tumor
suppressive role of miR-758-3p in BC, as indicated by its
inhibitory effect on cell proliferation, migration and invasion
through repression of NOTCH2 expression.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW, BC and XS designed the study, analyzed and
interpreted the results and prepared the manuscript. HS, JZ, FZ and
JC performed the experiments. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The protocol used in the present study was approved
by the Institutional Ethics Committee of Xiangyang Central Hospital
(Xiangyang, China). All patients provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tan M, Mu X, Liu Z, Tao L, Wang J, Ge J
and Qiu J: microRNA-495 promotes bladder cancer cell growth and
invasion by targeting phosphatase and tensin homolog. Biochem
Biophys Res Commun. 483:867–873. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jiang Z, Zhang Y, Cao R, Li L, Zhong K,
Chen Q and Xiao J: miR-5195-3p inhibits proliferation and invasion
of human bladder cancer cells by directly targeting Oncogene KLF5.
Oncol Res. 25:1081–1087. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang H, Li Q, Niu X, Wang G, Zheng S, Fu G
and Wang Z: miR-143 inhibits bladder cancer cell proliferation and
enhances their sensitivity to gemcitabine by repressing IGF-1R
signaling. Oncol Lett. 13:435–440. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li P, Yang X, Cheng Y, Zhang X, Yang C,
Deng X, Li P, Tao J, Yang H, Wei J, et al: MicroRNA-218 increases
the sensitivity of bladder cancer to cisplatin by targeting Glut1.
Cell Physiol Biochem. 41:921–932. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guo YW, Ying L, Tian Y, Yang PQ, Zhu YC,
Wang ZP, Qiu F and Lin J: miR-144 downregulation increases bladder
cancer cell proliferation by targeting EZH2 and regulating Wnt
signaling. FEBS J. 280:4531–4538. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xu X, Chen H, Lin YW, Hu ZH, Mao YQ, Wu J,
Xu XL, Zhu Y, Li SQ, Zheng XY and Xie LP: MicroRNA-409-3p inhibits
migration and invasion of bladder cancer cells via targeting c-Met.
Mol Cells. 36:62–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang SL, Gao YL and Chen XB: MicroRNA-214
targets PCBP2 to suppress the proliferation and growth of glioma
cells. Int J Clin Exp Pathol. 8:12571–12576. 2015.PubMed/NCBI
|
|
8
|
Zhao J, Xu J and Zhang R: MicroRNA-411
inhibits malignant biological behaviours of colorectal cancer cells
by directly targeting PIK3R3. Oncol Rep. 39:633–642.
2018.PubMed/NCBI
|
|
9
|
Yu Z, Lin X, Tian M and Chang W:
microRNA196b promotes cell migration and invasion by targeting
FOXP2 in hepatocellular carcinoma. Oncol Rep. 39:731–738.
2018.PubMed/NCBI
|
|
10
|
Shang A, Yang M, Shen F, Wang J, Wei J,
Wang W, Lu W and Wang C and Wang C: miR-1-3p suppresses the
proliferation, invasion and migration of bladder cancer cells by
up-regulating SFRP1 expression. Cell Physiol Biochem. 41:1179–1188.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yuan Q, Sun T, Ye F, Kong W and Jin H:
MicroRNA-124-3p affects proliferation, migration and apoptosis of
bladder cancer cells through targeting AURKA. Cancer Biomark.
19:93–101. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng C, Sun P, Hu J, Feng H, Li M, Liu G,
Pan Y and Feng Y, Xu Y, Feng K and Feng Y: miRNA-556-3p promotes
human bladder cancer proliferation, migration and invasion by
negatively regulating DAB2IP expression. Int J Oncol. 50:2101–2112.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang D, Cho W, Li Z, Xu X, Qu Y, Jiang Z,
Guo L and Xu G: miR-758-3p suppresses proliferation, migration and
invasion of hepatocellular carcinoma cells via targeting MDM2 and
mTOR. Biomed Pharmacother. 96:535–544. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meng X, Zhao Y, Wang J, Gao Z, Geng Q and
Liu X: Regulatory roles of miRNA-758 and matrix extracellular
phosphoglycoprotein in cervical cancer. Exp Ther Med. 14:2789–2794.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hayashi T, Gust KM, Wyatt AW, Goriki A,
Jäger W, Awrey S, Li N, Oo HZ, Altamirano-Dimas M, Buttyan R, et
al: Not all NOTCH Is created equal: The oncogenic role of NOTCH2 in
bladder cancer and its implications for targeted therapy. Clin
Cancer Res. 22:2981–2992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He X, Ping J and Wen D: MicroRNA-186
regulates the invasion and metastasis of bladder cancer via
vascular endothelial growth factor C. Exp Ther Med. 14:3253–3258.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Souza MF, Kuasne H, Barros-Filho MC,
Cilião HL, Marchi FA, Fuganti PE, Paschoal AR, Rogatto SR and Cólus
IMS: Circulating mRNAs and miRNAs as candidate markers for the
diagnosis and prognosis of prostate cancer. PLoS One.
12:e01840942017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Masuda T, Hayashi N, Kuroda Y, Ito S,
Eguchi H and Mimori K: MicroRNAs as Biomarkers in Colorectal
Cancer. Cancers (Basel). 9:1242017. View Article : Google Scholar
|
|
20
|
Amr KS, Elmawgoud Atia HA, Elazeem
Elbnhawy RA and Ezzat WM: Early diagnostic evaluation of miR-122
and miR-224 as biomarkers for hepatocellular carcinoma. Genes Dis.
4:215–221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shao LP, Shen ZJ, Qian H, Zhou SF and Chen
YG: Knockdown of miR-629 inhibits ovarian cancer malignant
behaviors by targeting testis-specific Y-like protein 5. DNA Cell
Biol. 36:1108–1116. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zheng YG, Lu XW, Xu LP, Chen Z, Li QX and
Yuan J: MicroRNA-675 promotes glioma cell proliferation and
motility by negatively regulating retinoblastoma 1. Hum Pathol.
69:63–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ding D, Zhang Y, Wen L, Fu J, Bai X, Fan
Y, Lin Y, Dai H, Li Q, Zhang Y and An R: miR-367 regulates cell
proliferation and metastasis by targeting metastasis-associated
protein 3 (MTA3) in clear-cell renal cell carcinoma. Oncotarget.
8:63084–63095. 2017.PubMed/NCBI
|
|
24
|
Wu YY, Song Y, Xiong Y, Wang X, Xu K, Han
B, Bai Y, Li L, Zhang Y and Zhou L: MicroRNA-21 (Mir-21) promotes
cell growth and invasion by repressing tumor suppressor PTEN in
colorectal cancer. Cell Physiol Biochem. 43:945–958. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang L, Lin TS, Xu CC, Hu SK, Pan YY and
Jin R: miR-124 interacts with the Notch1 signalling pathway and has
therapeutic potential against gastric cancer. J Cell Mol Med.
20:313–322. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen X, Xiao W, Chen W, Liu X, Wu M, Bo Q,
Luo Y, Ye S, Cao Y and Liu Y: MicroRNA-26a and −26b inhibit lens
fibrosis and cataract by negatively regulating Jagged-1/Notch
signaling pathway. Cell Death Differ. 24:19902017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Goriki A, Seiler R, Wyatt AW,
Contreras-Sanz A, Bhat A, Matsubara A, Hayashi T and Black PC:
Unravelling disparate roles of NOTCH in bladder cancer. Nat Rev
Urol. 15:345–357. 2018. View Article : Google Scholar : PubMed/NCBI
|