Introduction
Asthma is a common airway inflammatory disease that
affects more than 300 million people worldwide (1,2) and this
figure is estimated to increase to 400 million by 2025 (1). The World Health Organization (WHO)
placed asthma as 14th most critical disorder worldwide (1–3). Asthma
is characterized by airway inflammation, airway
hyper-responsiveness (AHR) and reversible airflow obstruction
(4). In general, asthma is caused by
T helper 2 (Th2)-driven inflammatory responses that induce airway
eosinophilia and mucus production in the lungs. The Th2-mediated
eosinophilic disease is commonly associated with atopy and is
characterized by an increase expression of Th2 cytokines, including
interleukin (IL)-4, IL-5 and IL-13 (5,6). On the
other hand, the non-Th2-subtype is characterized by the lack of Th2
inflammation and frequently associated with neutrophilic or
paucigranulocytic inflammation within the airways (5,7,8).
Current medications for the management of asthma
include short acting β-agonists, long acting β2-adrenergic
agonists, and inhaled corticosteroids, which could alleviate the
asthma attacks by relaxing the a smooth muscle in the airway.
However, certain adverse effects including tachycardia, anxiety,
osteoporosis, stunting of growth in children and cataract formation
could be seen following prolonged used (8,9).
Therefore, a safer alternative for the management of asthma is
needed and the use of natural products seems to be a promising
approach.
Lignosus rhinocerotis (Cooke) Ryvarden (Tiger
Milk mushroom) or locally known as “cendawan susu rimau” has a long
history of use as natural remedies for various diseases by the
local and indigenous communities in Malaysia (10). A previous study had demonstrated that
L. rhinocerotis sclerotia exhibited anti-proliferative
(11) and immunomodulatory
properties (12). Furthermore, Lee
et al (10) reported that
sclerotial powder of L. rhinocerotis also demonstrated
anti-inflammatory properties in carrageenan-induced paw oedema
model in rats. The first report on the anti-asthmatic properties of
L. rhinocerotis was reported by Johnathan et al
(13) which demonstrated that the
oral administration of L. rhinocerotis extract significantly
reduced the level of Th2 cytokines in the bronchoalveolar lavage
fluid (BALF), IgE level in the serum and attenuated the number of
leukocyte infiltrating into the lung tissues. In the present study,
the effectiveness of intranasal administration of hot water extract
of sclerotial powder of the L. rhinocerotis in the
ovalbumin-induced allergic asthma mouse model was examined.
Materials and methods
Preparation of L. rhinocerotis by hot
water extraction
Sclerotia of L. rhinocerotis cultivar TM02
was obtained from Ligno Biotech Sdn. Bhd. (Selangor, Malaysia) in
dried powdered form. To prepare the extract, 50 g of L.
rhinocerotis sclerotium powder was immersed in 600 ml of
purified distilled water and subjected to hot water extraction
using a Soxhlet extraction machine (14) for 5 days. The extract was then
subjected to a rotary evaporator (Ilshin BioBase, Gyeonggi-do,
South Korea) for freeze-drying into lyophilized powder form. A
total of ~5 g of L. rhinocerotis extract (LRE) could be
obtained from 50 g of sclerotial powder.
Animals
Ethical approval was obtained from the Animal Ethics
Committee of the Universiti Science Malaysia (Kelantan, Malaysia;
Animal Ethics Approval/2016/799). A total of 36 female Balb/c mice,
aged 6–8 weeks (weight, 20–30 g), were used in this experiment. The
mice (n=36) were maintained in polystyrene cages in an
air-controlled room at 25±1°C with a 12 h light/dark cycle and they
were given food pellet and water ad libitum. The animals
were acclimatized to the experimental environment prior to the
commencement of the study.
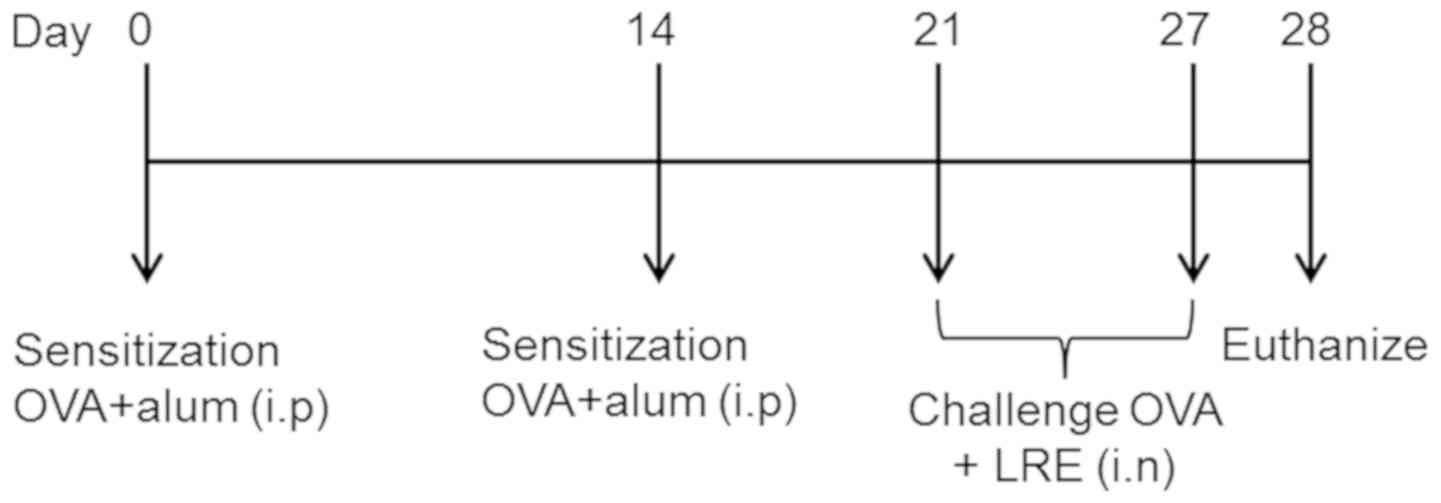
Sensitization, challenge and
treatment
The mice were randomly divided into six groups with
six mice per group (n=6): i) Normal group (as negative control),
ii) sensitization plus OVA challenge (as positive OVA control),
iii) sensitization plus OVA challenge/intranasal LRE (125 mg/kg per
body weight), iv) sensitization plus OVA challenge/intranasal LRE
(250 mg/kg per body weight), v) sensitization plus OVA
challenge/intranasal LRE (500 mg/kg per body weight) and vi)
sensitization plus OVA challenge/intraperitoneal (i.p)
dexamethasone (3 mg/kg per body weight; Nacalai Tesque, Inc.,
Kyoto, Japan). On day 0 and day 14, the mice were sensitized with
20 µg ovalbumin (OVA) and 4 mg aluminium hydroxide (alum) (both
Nacalai Tesque, Inc.) in 100 µl PBS (15). Starting on day 21, the mice were
challenged with 1% OVA aerosol for 20 min/day for 7 consecutive
days by using an ultrasonic nebulizer (Mabist mist; Mabist DMI
Healthcare, Illinois, CA, USA) as described in Fig. 1. The animals in the normal group were
sensitized and challenged with normal saline at the same time
intervals. Finally, 24 h following the final challenge, the mice
were euthanized with i.p pentobarbital (200 mg/kg) and samples were
collected; bronchoalveolar lavage fluid (BALF) for inflammatory
cell counts and cytokine determination, blood (0.5 ml/mice) for
total immunoglobulin (Ig) E level, lung draining lymph nodes (LN)
for cell subset populations and lungs for histopathological
analysis.
Eosinophil and inflammatory cell
counts
Following sacrifice, BALF was obtained using an
endotracheal tube by instilling and aspirating from the trachea
with 0.4 ml of 1% fetal calf serum (Capricon Scientific GmbH,
Ebsdorfergrund, Germany) in phosphate buffer saline (PBS) for three
times. BALF were centrifuged at (350 × g for 5 min at 4°C) and the
cell pellet was re-suspended with 1XPBS (50–200 µl, depending on
cell numbers) and centrifuged (350 × g for 5 min at 4°C) at room
temperature. Slides for differential cell counts were air dried,
fixed with methanol for 1–2 min and stained with Giemsa-stain for 8
min (Merck KGaA, Darmstadt, Germany) at room temperature. The
absolute numbers of each cell types (neutrophil, eosinophil and
lymphocyte) were identified using a hemocytometer, depending on the
standard morphology criteria of cells using a fluorescence
microscope at ×1,000 magnification and under an oil immersion
lens.
Cytokines quantification and IgE
determination
The total IgE level (BD Bioscience, San Jose, CA,
USA) in serum and Th2 cells secreting IL-4 (cat. no. 431105), IL-5
(catalog no. 431205; both Biolegend, Inc., San Diego, CA, USA) and
IL-13 (cat. no. 900-K207; Peprotech, Inc., Rocky Hill, NJ, USA) in
BALF were measured using ELISA kits according to the manufacturer's
protocol.
Cell surface staining
All fluorochrome-labelled monoclonal antibodies used
in surface staining were titrated prior to use in order to
determine the optimal antibody dilution for the surface marker
detection by flow cytometry (data not shown). Lung draining lymph
nodes (LN) cells were stained and analyzed with the combinations of
the monoclonal antibodies: Cluster of differentiation
(CD)3-PerCP-Cy™5.5 rat anti-mouse (1:200; cat. no.
561609) and CD4-fluorescein isothiocyanate (FITC; 1:200; cat. no.
553650; both BD Pharmingen™; BD Biosciences). The single
cell suspensions were centrifuged (300 × g for 10 min at 4°C) and
re-suspended in FACS staining buffer [PBS + 0.5% bovine serum
albumin (Thermo Fisher Scientific, Inc. Waltham, MA, USA) + 0.05%
sodium azide]. Following staining, the samples were scanned using a
flow cytometer (BD FACS Canto II™) and analyzed using
FCS Express 6 Flow Research Edition (De Novo Software, Glendale,
CA, USA).
Histopathological analysis
Following BALF collection, mice lungs were perfused
with PBS solution, removed and fixed in 10% neutral-buffered
formalin overnight at room temperature. A conventional tissue
processing method was applied in which the lung tissues were
embedded in paraffin and cut into 5-µm thickness sections, followed
by Harris haematoxylin staining for 20 min and eosin staining for 3
min at room temperature (both Sigma-Aldrich; Merck KGaA,
Dramstradt, Germany) for analysis of leukocyte infiltration. Lungs
were stained with periodic acid-schiff staining (Sigma-Aldrich;
Merck KGaA) for 20 min at room temperature for the analysis of
goblet cell hyperplasia. The tissue was subsequently mounted and
cover-slipped with di-n-butyl phthalate in xylene (DPX) mounting
medium. Morphometric histological analyses were performed under a
fluorescence microscope (Olympus Corporation, Tokyo, Japan). The
cell infiltration intensity at the peribronchiolar and the
perivascular region was graded on the hot spot area using the
following scores; 0: No inflammatory cells, 1: A few inflammatory
cells, 2: A ring of inflammatory cells (1 cell layer deep), 3: A
ring of inflammation cells (2–4 cells layer deep), 4: A ring of
inflammation cells (>4 cells layer deep). For the analysis of
mucus, the scoring was done according to the following method; 0:
No goblet cells, 1: <25% of epithelium, 2: 25–50% of epithelium,
3: 50–75% of epithelium, 4: >75% of epithelium (16,17).
Statistical analysis
Data was expressed as mean ± standard error of the
mean and experiments were performed in triplicate. Statistical
significance was determined by one-way analysis of variance,
followed by Bonferroni's post hoc test to determine the significant
difference between the treatment groups by using GraphPad Prism
software version 6.01 (Graphpad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Eosinophil and inflammatory cell
counts
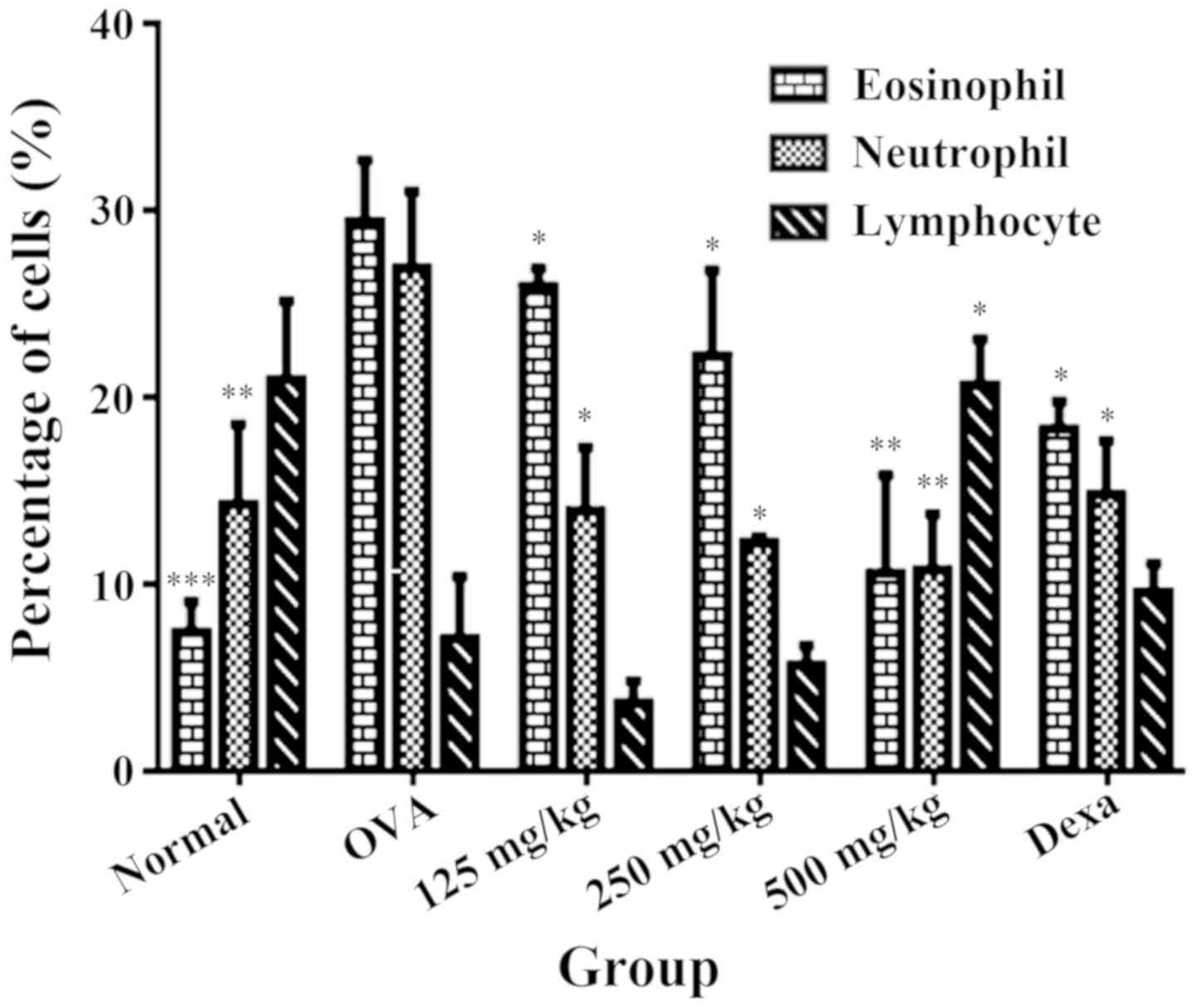
The number of eosinophils, neutrophils and
lymphocytes present in BALF from the normal group were 9.0±1.64,
13.0±2.15, 18.3± and 3.18 respectively. Sensitization and challenge
with OVA stimulated a significant increase of the eosinophil and
neutrophil counts (36.17±1.74 and 26.83±2.24 respectively) compared
with the normal group, indicating a satisfactory level of
sensitization towards the allergens. However, OVA sensitization and
challenged demonstrated a reduction in lymphocyte count with
7.17±1.92 compared with the normal group. LRE treatment (125, 250
and 500 mg/kg) and dexamethasone significantly attenuated the
eosinophil and neutrophil infiltration, reaching its highest level
at 500 mg/kg (P<0.01; Fig. 2).
However, the lymphocyte counts significantly increased (P<0.01)
following treatment with 500 mg/kg of LRE compared with the OVA
group.
RE suppresses the level of Th2
cytokine and serum immunoglobulin E (IgE) production
Results indicated that LRE significantly (P<0.01)
attenuated the level of IL-4 and IL-5 at 500 mg/kg compared with
the OVA group (Fig. 3A and B) and
notably, dexamethasone significantly reduced IL-4 level in BALF
(P<0.05; Fig. 3A). In contrast,
LRE (250 mg/kg) significantly restored the IL-13 level compared
with the OVA-challenged group (P<0.01; Fig. 3C). These findings imply that LRE is
capable of modifying the Th2 predominant immune activity in
OVA-induced mouse model of asthma. Therefore, to further assess
whether LRE could modify the OVA-specific Th2 response, the serum
IgE level was determined using ELISAs.
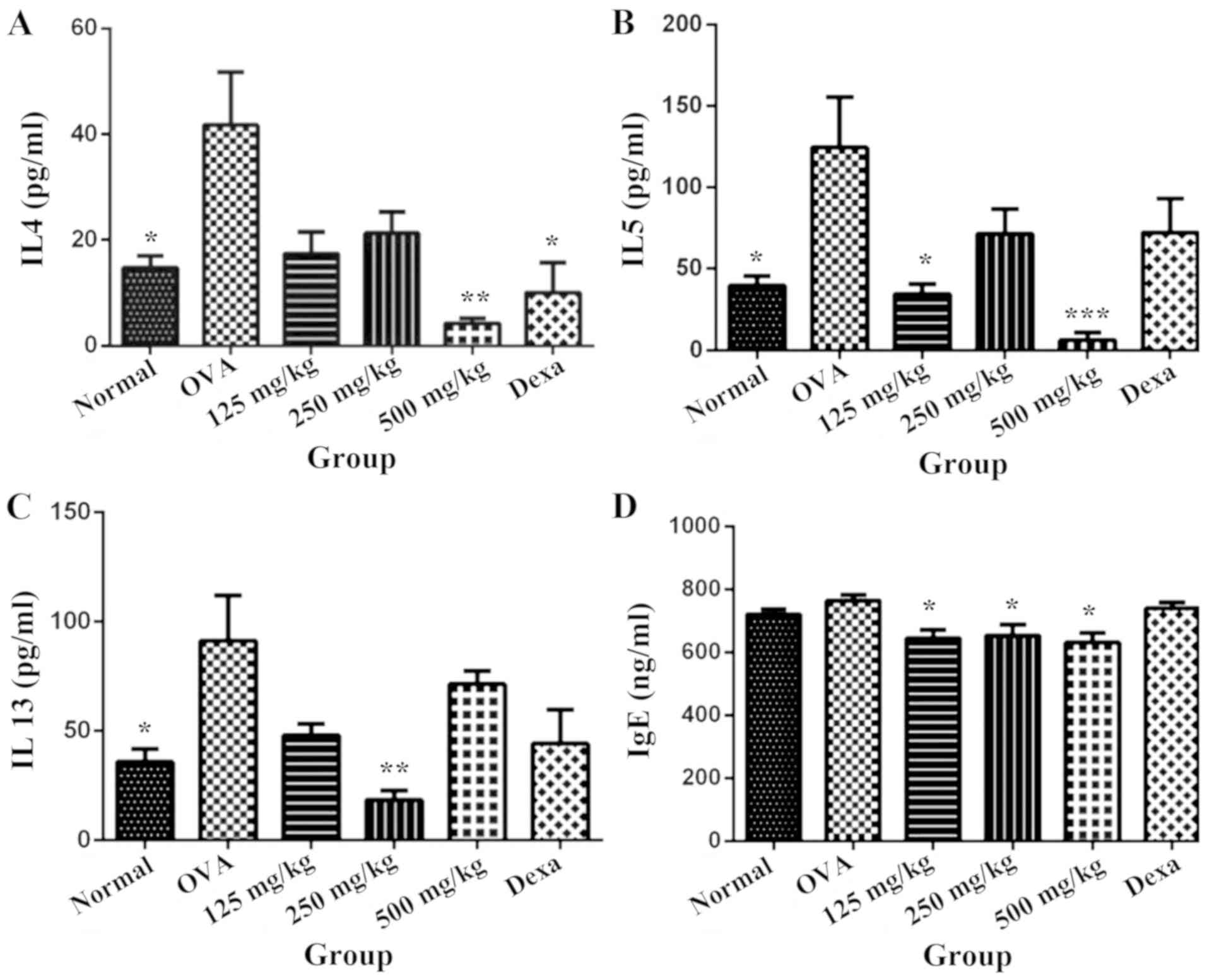 | Figure 3.LRE inhibits the production of helper
T2 cell-specific cytokines in BALF and IgE in blood serum. Effects
of intranasal administration of LRE (125, 250 and 500 mg/kg) and
administration of Dexa (i.p; 3 mg/kg) on the level of (A) IL-4, (B)
IL-5, (C) IL-13 in BALF and (D) IgE in serum are presented. Mice
were sensitized on day 0 and 14 with (OVA; 200 mg/ml) prepared in
alum and challenged with 1% OVA. The normal group was administered
(i.p) and challenged with saline instead of OVA. Values are
expressed as the mean ± standard deviation (n=6/group). *P<0.05,
**P<0.01 and ***P<0.001 indicates significant different from
OVA. BALF, bronchoalveolar lavage fluid; OVA, ovalbumin; alum,
aluminium hydroxide; i.p., intraperitoneal; Dexa, dexamethasone;
LRE, Lignosus rhinocerotis extract; Ig, immunoglobulin; IL,
interleukin. |
Taking this into account, the concentration of IgE
present in the blood serum of normal group was 720.8±15.66 ng/ml,
whereas sensitization and OVA challenge promoted a slight increase
to 764.6±18.74 ng/ml (Fig. 3D).
Dexamethasone however did not demonstrate any significant change
(P>0.05) compared with the OVA-induced group.
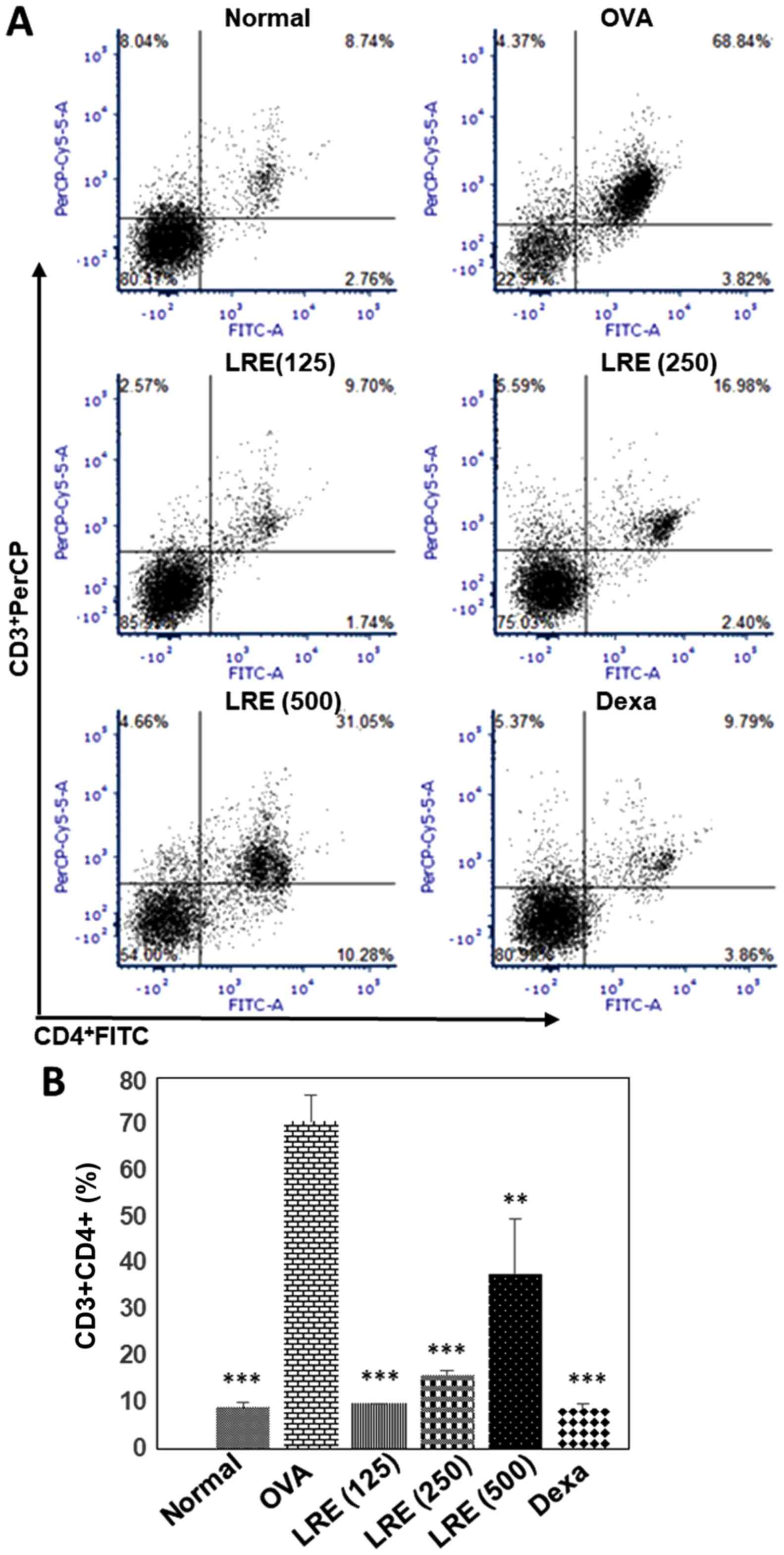
The effects of LRE on helper T cell
populations
Fig. 4A and B
presents the isolation of cells from lung-draining LN. The cells
were double-stained with CD3-PerCP and CD4-FITC and analyzed by
flow cytometry. The percentage of CD3+CD4+
cells in the normal group was 8.74±0.87%. On the other hand, the
stimulation with OVA increased the percentage of the cells by
68.84±4.82% (Fig. 4B). Treatment
with LRE indicated the reduction of helper T-lymphocytes percentage
at all dosages and reached its greater reduction at 125 mg/kg
(P<0.001). Similarly, dexamethasone significantly attenuated the
CD3+CD4+ cell population (P<0.001)
compared with the OVA group.
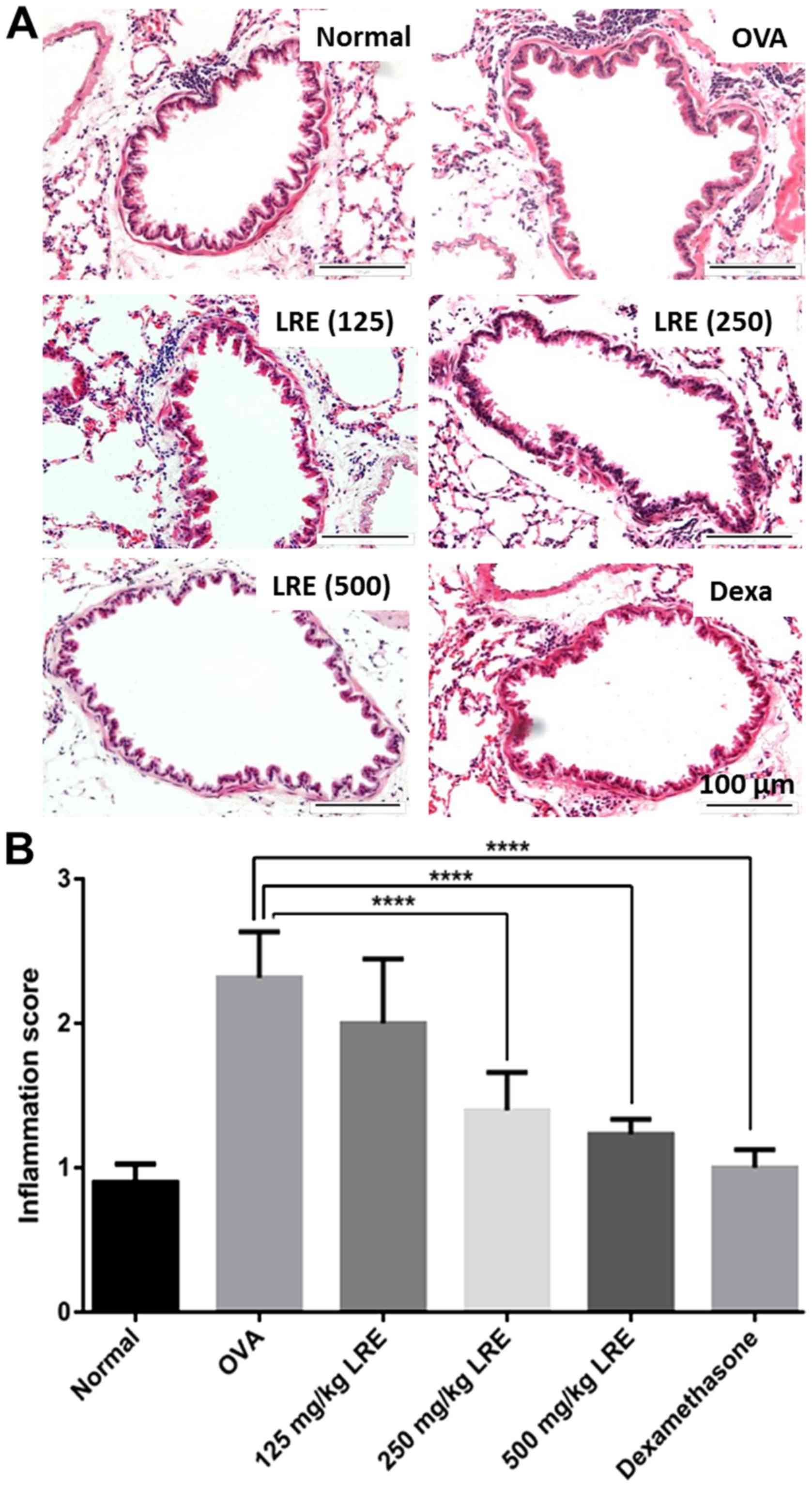
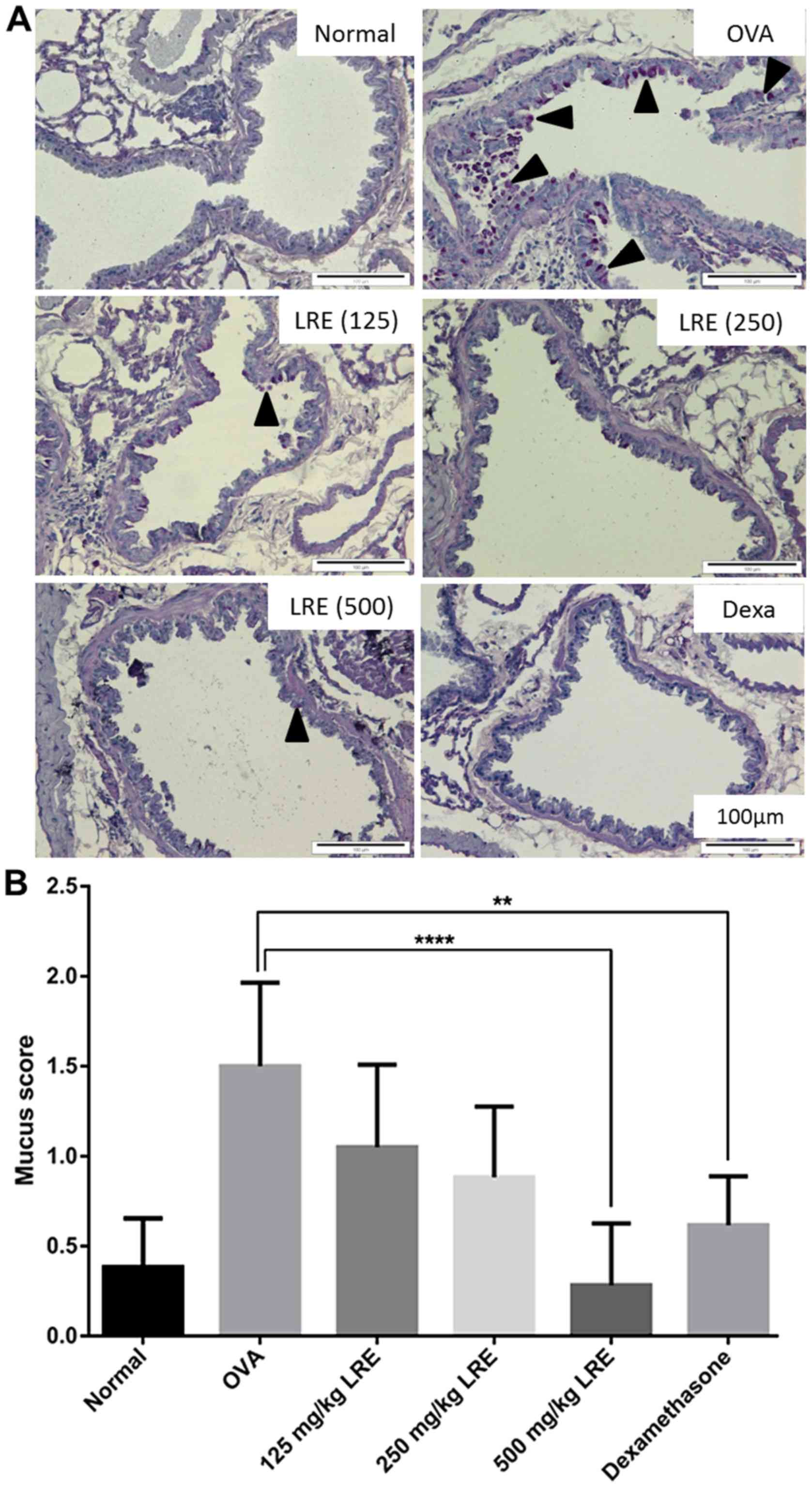
LRE suppresses leukocytes infiltration
and mucus production in lungs
LRE (250 and 500 mg/kg) and dexamethasone treatment
significantly diminished the eosinophil-rich leukocyte infiltration
compared with the OVA challenge group (P<0.0001; Fig. 5), while mucus hypersecretion in
OVA-induced mice was significantly attenuated by LRE treatment at
500 mg/kg (P<0.0001) and dexamethasone (P<0.01; Fig. 6).
Discussion
Asthma is a complex inflammatory airway disease that
results from the activation of various inflammatory and structural
cells, leading to airway inflammation, airway obstruction, mucus
hypersecretion and acute hyperresponsiveness (18). The anti-asthmatic activity of L.
rhinocerotis was investigated through a mouse model induced by
OVA sensitization. Generally, animals demonstrated similar
hallmarks to acute human allergic asthma characterized by
pathophysiological alterations in airways, mucus secretion,
production of allergen specific IgE and the increase of Th2
cytokines level (19). The mechanism
of asthma is not fully elucidated, but in general it occurs due to
an imbalance in Th1/Th2 components and other factors associated
with disease occurrence. It is characterized by the increasing
level of Th2 cytokines (IL-4, IL-5 and IL-13) and the reduction in
Th1 cytokine levels (interferon-gand IL-2) (20,21).
A mouse model of asthma was established using OVA by
sensitization and challenge to mimic the pathological alterations
in asthma patients. Generally, following allergens exposure, T
cells will be activated by the dendritic cells, which lead to Th2
response, resulting in the production of cytokines (IL-4, IL-5 and
IL-13). CD4+ cells secrete IL-4 and IL-13, that are
involved in the production of IgE by mast cells, transforming
growth factor β (TGF-β) and causing direct effects on fibroblasts,
epithelial cells, and airway smooth muscle that leads to airway
narrowing, AHR and structural changes (22). Furthermore, IL-5 is crucial in
development and activation of eosinophils that directly influence
the airway narrowing through the release of TGF-β, IL-4, and IL-13.
In the present study, Balb/c mice were used as a model for
induction of airway inflammation as they could develop an efficient
Th2-biased immunological response (23,24). In
this study, the animals (except for normal group) were sensitized
twice on day 0 and day 14, to ensure the eosinophils did not return
to the baseline level and to prevent the development of tolerance
(25). Dexamethasone was used as
positive control because it is an effective inhibitor of airway
inflammation and airway remodeling in animal models (26).
Lymphocytes coordinate the immune response and serve
a central role in cell mediated immunity. Lymphocyte subsets may
include helper T cells (CD4+ T-cells), cytotoxic T cells
(CD8+ T-cells), memory T cells and regulatory T cells
(Treg cells) (27). Flow cytometry
is used to provide absolute counts, percentages and/or ratios of
these lymphocyte subsets (27).
CD4+ T cells become activated and proliferate rapidly
upon encounter with antigens, secreting cytokines that sends
signals and maintain an active immune response (27). The CD4+ count provides a
picture of immune system competence, with higher counts typically
signifying healthier immune systems. Activation of T cells could
influence the severity of asthma, including the degree of airway
narrowing and bronchial eosinophil response (28). Apart from the involvement of Th1 and
Th2 cells in the pathogenesis of asthma, it was reported that Treg
cells also serve a crucial role in asthma (29,30).
Foxp3+ Treg cells are a distinct subset of
CD4+ T cells which can suppress effector
CD4+T cells responses (31) and have been demonstrated to serve a
crucial role in allergic diseases, including asthma (29,31,32).
Foxp3+ Treg cells can attenuate the Th2 and Th17
cell-mediated inflammation and prevent airway inflammation as well
as bronchial hyper-responsiveness in asthmatic patients and in
animal models (32,33). In the current study, OVA-sensitized
and -challenged animals had CD4+T cells activated and
therefore the inflammatory responses were induced, the cytokines
level was altered and the production of leukocyte infiltration in
the lungs regions increased. Notably, intranasal administration of
LRE modulated the percentages of CD3+CD4+
cells; similar observation could be observed in dexamethasone
treated-animals.
Allergic asthma is associated with eosinophilic
inflammation in the airways (34).
Generally, eosinophils are developed and distinguished under the
influence of IL-5, which enters the site of inflammation or
infection (35). Activation of
eosinophils leads to the release of pro-inflammatory mediators
including major basic protein, cationic protein, leukotriene C4,
prostaglandin E2 and thromboxane. They are also capable of
synthesizing and releasing interleukins (IL-3, IL-4, IL-5, IL-8,
IL-10, 1L-12 and IL-13), chemokines (CCL5/RANTES and
CCL11/eotaxin-1), tumor necrosis factor-α and TGF-β (36). A number of studies have pointed out
that the neutralization of IL-5 could diminish pulmonary
eosinophilia in response to allergens (26,35).
Furthermore, neutrophils are known to be one of the first
inflammatory cells to be recruited to the site of allergens
exposure. It produces metalloproteases and elastase, which are
vital in vascular permeability, mucus secretion and
bronchoconstriction (37). Nabe
et al (38), demonstrated an
increase of neutrophils level in the airways of mouse
OVA-challenged model. This study indicated that OVA-sensitized and
-challenged group escalated the number of eosinophil and neutrophil
cells and these cells were attenuated with the treatment of LRE and
dexamethasone. Furthermore, the suppression of the inflammatory
cells in BALF was also validated by lung tissue histology.
In allergen-sensitized model with atopic asthma,
re-exposure to the allergen leads to IgE mediated inflammatory
cascade in the airways. Airway resident cells (e.g., mast cells and
macrophages), newly mobilized immune cells (e.g., eosinophil and
neutrophil) and epithelial cells are vital in this inflammatory
cascade (39). In allergic
inflammation, there seems to be an imbalance between Th1/Th2
cytokines with dominance towards Th2 cytokines (IL-4, IL-5 and
IL-13) (40). IL-4 is known to be
the main cytokine involved in the pathogenesis of the allergic
response. It has vital functions in airway remodeling by
stimulating the mucus production and fibroblasts, inducing B-cells
to produce IgE and upregulation of molecules that enable the
migration of leukocytes to the airways (4). It has been extensively reported that
IL-5 is reliable in an eosinophil's development in the bone marrow
and their recruitment into interstitial mucosa and lungs due to
chemokine's production. Furthermore, IL-13 is reported to be the
most effective inducer of eotaxin expression by airway epithelial
cells on the respiratory tract and promotes mucus production in
lung tissues (41). IL-13 however,
is also reported to have independent roles from IL-4, IL-5,
eosinophil and IgE (42). IL-13 has
been demonstrated to independently elicit a number of key
pathological features of asthma including migration and
upregulation of adhesion molecules, goblet cell hyperplasia and
stimulation of airway hyperresponsiveness (43). It is crucial to know that there are
few therapies for asthma that are targeting Th2 cytokines, for
examples recombinant soluble IL-4 receptor antagonist
(Altrakincept®, Aerovance Incorporation; Pitrakinra and
Amgen), IL-4 receptor α-chain antibody (Dupilumab®,
Regeneron Pharmaceuticals), IL-13 blockade
(Lebrikizumab®, Genentech) and antibody to the IL-5
receptor (Benralizumab®, AstraZeneca/MedImmune)
(6,44,45).
According to Holgate (46), an increase level of IgE in asthmatic
patients is strongly associated with the increase of Th2 cytokines,
therefore worsening the clinical symptoms of the disease. IL-4
directly engages in the differentiation of B lymphocytes towards
IgE synthesis. This highlights the vital function of receptor
(FcεRI) that is present in mast cells, basophils and dendritic
cells that induce cellular activation, inflammation, and production
of mediators including Th2 cytokines (41,47). In
the present study, LRE (125, 250 and 500 mg/kg) reduced the IgE
level compared with the OVA group. Upon allergen exposure, the
IgE-coated mast cells identify the allergen deposited prior to
degranulating. Activation of mast cells leads to the production of
histamine, cysteinyl-leukotrienes and prostaglandin D2, which in
turn lead to the additional recruitment of Th2 cells, eosinophils
and basophils to the tissue (48).
This process promotes immediate bronchoconstriction, airway
inflammation and airway construction, congestion and systemic
reflexes (49).
Histological examination was performed on the lung
tissues to analyze the effect of LRE on the histological features
of asthma and to examine the structural changes or pathological
responses in the lung tissue. The development of asthma can be
characterized by the mobilization of migrated cells, especially
eosinophils and neutrophils into the peribronchiolar and
perivascular regions (50). In the
present study, histopathological analysis of lungs tissue sections
demonstrated that the sensitization and OVA challenge promoted
inflammation indicated by the presence of leukocyte infiltration
and markedly increased goblet cell hyperplasia and mucus
hypersecretion in the bronchi compared with the normal group. This
study demonstrated that LRE could effectively inhibit the
inflammatory response in the lung of the OVA-challenged mice by
reducing the number of leukocytes infiltration and mucus
production. A study by Jin et al (51) stated that Astragalus
membranaceus remarkably attenuated the airway inflammation in
OVA-sensitized animals. Similarly, ethanolic extract from
Erythrina mulungu Benth significantly decreased the cellular
inflammatory infiltration in the lung tissue (52). These results were in agreement with
the previous study which demonstrated that oral administration of
L. rhinocerotis effectively reduced the leukocyte migration
in the lung tissues (13). In
particular, studies on transbronchial biopsies of patients with
nocturnal asthma demonstrated that a decrease in lung functions was
correlated with the increasing number of CD4+ T
lymphocytes and eosinophils in the alveolar walls (53–55).
In this study the attenuation of asthma-associated
parameters i.e. Th2 cytokines, IgE, percentage of CD4+ T
cell population as well as leukocytes and mucus infiltration in the
lungs were demonstrated. Intranasal administration of LRE exhibited
protective effects against OVA-induced asthma; the results of the
present study suggest the potential of L. rhinocerotis as an
alternative for the management of allergic airway inflammation.
Acknowledgements
The authors would like to thank Mr. Jamaruddin Mat
Asan from the Immunology Department, School of Medical Sciences and
staff from Animal Research and Service Centre USM for their
assistance throughout the study.
Funding
The present study was funded by the Research
University Grants (grant no: 1001/PPSK/813065 and
1001/PPSK/812180).
Availability of data and materials
All data generated or analysed during the present
study are included in this published article.
Authors' contributions
SAM, NSM and NDAI performed the experiments and
manuscript preparation. SS was involved in the analysis and
validation of histology slides. RM intranasally administered of LRE
into the animals. AAN was involved in planning of and supervising
the project. All authors discussed the results and contributed to
the final manuscript.
Ethics approval and consent to
participate
The experimental protocols and animal care were
approved by the Animal Ethics Committee, Universiti Sains Malaysia
(Animal Ethics Approval/2016/(799).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Network GA: The global asthma report 2014.
(Auckland, New Zealand). Global Asthma Network, 2014.
2017.PubMed/NCBI
|
|
2
|
Murray CJ, Vos T, Lozano R, Naghavi M,
Flaxman AD, Michaud C, Ezzati M, Shibuya K, Salomon JA, Abdallah S,
et al: Disability-adjusted life years (DALYs) for 291 diseases and
injuries in 21 regions, 1990–2010: A systematic analysis for the
Global Burden of disease study 2010. Lancet. 380:2197–2223. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
To T, Stanojevic S, Moores G, Gershon AS,
Bateman ED, Cruz AA and Boulet LP: Global asthma prevalence in
adults: Findings from the cross-sectional world health survey. BMC
Public Health. 12:2042012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Steinke JW and Borish L: Th2 cytokines and
asthma. Interleukin-4: Its role in the pathogenesis of asthma, and
targeting it for asthma treatment with interleukin-4 receptor
antagonists. Respi Res. 2:66–70. 2001. View
Article : Google Scholar
|
|
5
|
Anto JM, Bousquet J, Akdis M, Auffray C,
Keil T, Momas I, Postma DS, Valenta R, Wickman M, Cambon-Thomsen A,
et al: Mechanisms of the development of allergy (MeDALL):
Introducing novel concepts in allergy phenotypes. J Allergy Clin
Immunol. 139:388–399. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fajt ML and Wenzel SE: Development of new
therapies for severe asthma. Allergy Asthma Immunol Res. 9:3–14.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kaur J, Alvarez A, Hanna HW, Krishnan AC,
Senador D, Machado TM, Altamimi YH, Lovelace AT, Dombrowski MD,
Spranger MD, et al: Interaction between the muscle metaboreflex and
the arterial baroreflex in control of arterial pressure and
skeletal muscle blood flow. Am J Physiol Heart Circ Physiol.
311:H1268–H1276. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Newnham JP: Is prenatal glucocorticoid
administration another origin of adult disease? Clin Exp Pharmacol
Physiol. 28:957–961. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Barnes PJ: Reduced histone deacetylase in
COPD: Clinical implications. Chest. 129:151–155. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee SS, Tan NH, Fung SY, Sim SM, Tan CS
and Ng ST: Anti-inflammatory effect of the sclerotium of Lignosus
rhinocerotis (Cooke) Ryvarden, the Tiger Milk mushroom. BMC
Complement and Altern Med. 14:3592014. View Article : Google Scholar
|
|
11
|
Yap YH, Tan N, Fung S, Aziz AA, Tan C and
Ng S: Nutrient composition, antioxidant properties, and
anti-proliferative activity of Lignosus rhinocerus Cooke
sclerotium. J Sci Food Agric. 93:2945–2952. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wong KH, Lai CK and Cheung PC:
Immunomodulatory activities of mushroom sclerotial polysaccharides.
Food Hydrocol. 25:150–158. 2011. View Article : Google Scholar
|
|
13
|
Johnathan M, Gan SH, Ezumi MF, Faezahtul
AH and Nurul AA: Phytochemical profiles and inhibitory effects of
Tiger Milk mushroom (Lignosus rhinocerus) extract on
ovalbumin-induced airway inflammation in a rodent model of asthma.
BMC Complement Altern Med. 16:1672016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jonathan G, Loveth K and Elijah O:
Antagonistic effect of extracts of some Nigerian higher fungi
against selected pathogenic microorganisms. Am Eur J Agric Environ
Sci. 2:364–368. 2007.
|
|
15
|
Jeon WY, Shin IS, Shin HK, Jin SE and Lee
MY: Aqueous extract of gumiganghwal-tang, a traditional herbal
medicine, reduces pulmonary fibrosis by transforming growth
Factor-β1/Smad signaling pathway in murine model of chronic Asthma.
PLoS One. 11:e01648332016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Myou S, Leff AR, Myo S, Boetticher E, Tong
J, Meliton AY, Liu J, Munoz NM and Zhu X: Blockade of inflammation
and airway hyperresponsiveness in immune-sensitized mice by
dominant-negative phosphoinositide 3-kinase-TAT. J Exp Med.
198:1573–1582. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee MY, Ahn KS, Kwon OK, Kim MJ, Kim MK,
Lee IY, Oh SR and Lee HK: Anti-inflammatory and anti-allergic
effects of kefir in a mouse asthma model. Immunobiol. 212:647–654.
2007. View Article : Google Scholar
|
|
18
|
Prado CM, Leick-Maldonado EA, Kasahara DI,
Capelozzi VL, Martins MA and Tibério IF: Effects of acute and
chronic nitric oxide inhibition in an experimental model of chronic
pulmonary allergic inflammation in guinea pigs. American Am J
Physiol Lung Cell Mol Physiol. 289:L677–L683. 2005. View Article : Google Scholar
|
|
19
|
Kumar RK, Herbert C and Foster PS: The
‘classical’ ovalbumin challenge model of asthma in mice. Curr Drug
Targets. 9:485–494. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kang JH, Kim BS, Uhm TG, Lee SH, Lee GR,
Park CS and Chung IY: Gamma-secretase inhibitor reduces allergic
pulmonary inflammation by modulating Th1 and Th2 responses. Am J
Respi Critical Care Med. 179:875–882. 2009. View Article : Google Scholar
|
|
21
|
Zhou M, Cui ZL, Guo XJ, Ren LP, Yang M,
Fan ZW, Han RC and Xu WG: Blockade of Notch signalling by
γ-secretase inhibitor in lung T cells of asthmatic mice affects T
cell differentiation and pulmonary inflammation. Inflammation.
38:1281–1288. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shin YS, Takeda K and Gelfand EW:
Understanding asthma using animal models. Allergy Asthma Immunol
Res. 1:10–18. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Trifilieff A, El-Hashim A and Bertrand C:
Time course of inflammatory and remodeling events in a murine model
of asthma: Effect of steroid treatment. Am J Physiol Lung Cell Mol
Physiol. 279:L1120–L1128. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bosire EM, Nyamache AK, Gicheru MM,
Khamadi SA, Lihana RW and Okoth V: Population specific reference
ranges of CD3, CD4 and CD8 lymphocyte subsets among healthy
Kenyans. AIDS Res Ther. 10:242013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Larché M, Robinson DS and Kay AB: The role
of T lymphocytes in the pathogenesis of asthma. J Allergy Clin
Immunol. 111:450–464. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Barnes PJ: Immunology of asthma and
chronic obstructive pulmonary disease. Nat Rev Immunol. 8:183–192.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Massoud AH, Charbonnier LM, Lopez D,
Pellegrini M, Phipatanakul W and Chatila TA: An asthma-associated
IL4R variant exacerbates airway inflammation by promoting
conversion of regulatory T cells to TH17-like cells. Nat Med.
22:1013–1022. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tao B, Ruan G, Wang D, Li Y, Wang Z and
Yin G: Imbalance of peripheral Th17 and regulatory T cells in
children with allergic rhinitis AND bronchial asthma. Iran J
Allergy Asthma Immunol. 14:273–279. 2015.PubMed/NCBI
|
|
29
|
Tian M, Wang Y, Lu Y, Jiang YH and Zhao
DY: Effects of sublingual immunotherapy for dermatophagoides
farinae on Th17 cells and CD4(+) CD25(+) regulatory T cells in
peripheral blood of children with allergic asthma. Int Forum
Allergy Rhinol. 4:371–375. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma C, Ma Z, Fu Q and Ma S: Curcumin
attenuates allergic airway inflammation by regulation of CD4+ CD25+
regulatory T cells (Tregs)/Th17 balance in ovalbumin-sensitized
mice. Fitoter. 87:57–64. 2013. View Article : Google Scholar
|
|
31
|
Boyce JA and Austen KF: No audible
wheezing: Nuggets and conundrums from mouse asthma models. J Exp
Med. 201:1869–1873. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nials AT and Uddin S: Mouse models of
allergic asthma: Acute and chronic allergen challenge. Dis Models
Mech. 1:213–220. 2008. View Article : Google Scholar
|
|
33
|
Ming M, Luo Z, Lv S, Sun Q and Li C:
Inactivated Mycobacterium phlei inhalation ameliorates allergic
asthma through modulating the balance of CD4+CD25+ regulatory T and
Th17 cells in mice. Iran J Basic Med Sci. 19:953–959.
2016.PubMed/NCBI
|
|
34
|
Deckers J, Branco Madeira F and Hammad H:
Innate immune cells in asthma. Trends Immunol. 34:540–547. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stone KD, Prussin C and Metcalfe DD: IgE,
mast cells, basophils, and eosinophils. J Allergy Clin Immunol. 125
(2 Suppl 2):S73–S80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bandeira-Melo C, Bozza PT and Weller PF:
The cellular biology of eosinophil eicosanoid formation and
function. J Allergy Clin Immunol. 109:393–400. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Monteseirín J: Neutrophils and asthma. J
Investig Allergol Clin Immunol. 19:340–354. 2009.PubMed/NCBI
|
|
38
|
Nabe T, Hosokawa F, Matsuya K, Morishita
T, Ikedo A, Fujii M, Mizutani N, Yoshino S and Chaplin DD:
Important role of neutrophils in the late asthmatic response in
mice. Life Sci. 88:1127–1135. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Busse WW and Lemanske RF Jr: Asthma. New
Eng J Med. 344:350–362. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hwang SS, Kim YU, Lee S, Jang SW, Kim MK,
Koh BH, Lee W, Kim J, Souabni A, Busslinger M and Lee GR:
Transcription factor YY1 is essential for regulation of the Th2
cytokine locus and for Th2 cell differentiation. Proc Natl Acad Sci
USA. 110:276–281. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lambrecht BN and Hammad H: The immunology
of asthma. Nat Immunol. 16:45–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wills-Karp M: Interleukin-13 in asthma
pathogenesis. Immunol Rev. 202:175–190. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Brightling C, Saha S and Hollins F:
Interleukin-13: Prospects for new treatments. Clin Exp Allergy.
40:42–49. 2010.PubMed/NCBI
|
|
44
|
Desai D and Brightling C: Cytokine and
anti-cytokine therapy in asthma: Ready for the clinic? Clin Exp
Immunol. 158:10–19. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fatemi F, Sadroddiny E, Gheibi A,
Mohammadi Farsani T and Kardar GA: Biomolecular markers in
assessment and treatment of asthma. Respirology. 19:514–523. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Holgate ST: Innate and adaptive immune
responses in asthma. Nat Med. 18:673–683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gould HJ and Sutton BJ: IgE in allergy and
asthma today. Nat Rev Immunol. 8:205–217. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Barnes PJ: The cytokine network in asthma
and chronic obstructive pulmonary disease. J Clin Invest.
118:3546–3556. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Galli SJ, Grimbaldeston M and Tsai M:
Immunomodulatory mast cells: Negative, as well as positive,
regulators of immunity. Nat Rev Immunol. 8:478–486. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Curtis JL, Warnock ML, Arraj SM and
Kaltreider HB: Histologic analysis of an immune response in the
lung parenchyma of mice. Angiopathy accompanies inflammatory cell
influx. Am J Pathol. 137:689–699. 1990.PubMed/NCBI
|
|
51
|
Jin H, Luo Q, Zheng Y, Nurahmat M, Wu J,
Li B, Lv Y, Wang G, Duan X and Dong J: CD4+CD25+Foxp3+ T cells
contribute to the antiasthmatic effects of Astragalus membranaceus
extract in a rat model of asthma. Int Immunopharmacol. 15:42–49.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Amorim J, de Calvalho Borges M, Fabro AT,
Contini SHT, Valdevite M, Pereira AMS and Carmona F: The ethanolic
extract from Erythrina mulungu Benth. flowers attenuates
allergic airway inflammation and hyperresponsiveness in a murine
model of asthma. J Ethnopharmacol. Aug 10–2018.(Epub ahead of
print). View Article : Google Scholar
|
|
53
|
Kraft M: The distal airways: Are they
important in asthma? Eur Respir J. 14:1403–1417. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kraft M, Martin RJ, Wilson S, Djukanovic R
and Holgate ST: Lymphocyte and eosinophil influx into alveolar
tissue in nocturnal asthma. Am J Respir Crit Care Med. 159:228–234.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kelly EA, Houtman JJ and Jarjour NN:
Inflammatory changes associated with circadian variation in
pulmonary function in subjects with mild asthma. Clin Exp Allergy.
34:227–233. 2004. View Article : Google Scholar : PubMed/NCBI
|