Introduction
Chronic liver diseases remain a major problem
throughout the world and lead to considerable morbidity and
mortality (1). The most common
pathological outcome and the most severe complication of chronic
liver diseases are liver fibrosis which are global health and
economic burdens (2). Liver fibrosis
characteristically show over-accumulation of extracellular matrix
(ECM), mainly type I fibrillar collagen (Collagen I) in response to
chronic liver damage (3). Various
etiological factors and stimuli can result in development of liver
fibrosis, including viral hepatitis (hepatitis C and B),
environmental carcinogens, excessive ethanol consumption, activated
hepatic stellate cells (HSCs) and non-alcoholic steatohepatitis
(NASH) (4,5). Liver fibrosis is generally asymptomatic
and is often overlooked by patients and their families. When
patients are diagnosed with liver fibrosis, the strategies for
treatment and therapeutic options are seriously limited (6). In addition, there are no vaccine or
effective anti-fibrotic drugs so far, and the patients with liver
fibrosis need to receive long-term and repeated drug treatments,
which is difficult to accept (7).
Therefore, it is necessary to search for a better therapy to
directly prevent liver fibrosis, and a better understanding of
pathogenesis underlying the development of liver fibrosis is
urgently needed.
In recent years, therapy based on cells has been
widely investigated in the area of tissue or organ protection
(8). Therefore, we focus on the
cells which are connected with the development of liver fibrosis.
HSCs have been considered as a kind of lipocytes, which exhibit a
key role in the process of liver fibrosis when the liver is damaged
(9,10). HSCs are regarded as the potential
aetiology because they are the key cell type which is in charge of
ECM formation during hepatic fibrogenesis (11). Activation of HSCs is essential to the
initiation and progression of liver fibrosis (12). When HSCs are activated, they become
fibrogenic myofibroblasts and exhibit the function of fibrogenic
myofibroblast-like cells, which can secrete α-smooth muscle actin
(α-SMA), TIMP-1 and inhibit HSC cell apoptosis and collagen-I
secretion (13–15). Excessive insoluble collagen I and
matrix components in the intracellular and perisinusoidal spaces
can bring severe compromise to the hepatic function (16,17).
MicroRNAs (miRNAs) have been found to be the most
abundant small non-coding RNAs (ncRNAs) (18). miRNAs are a type of 18 to 25
nucleotides (nt) in length, single-stranded and evolutionary
conserved endogenous RNAs (19).
miRNAs can regulate the expression of post-transcriptional genes by
incompletely binding with their target genes 3′-UTR. Accumulating
evidence suggests that miRNAs are related to the cell
proliferation, differentiation, apoptosis and development of
various diseases (20). Recent
studies have shown that miRNAs take part in the process of liver
diseases, such as liver injury, hepatic cirrhosis, hepatocellular
carcinoma and liver fibrosis (21–24).
Moreover, there is increasing evidence that miRNAs are involved in
various fibrotic diseases, including kidney, cardiac, cystic and
pulmonary fibrosis (25–28). miRNAs may serve as antifibrotic or
profibrotic genes during liver fibrosis. However, the mechanism
effects of miRNAs need further investigation.
let-7a has been found to be involved in the
development of chronic liver diseases (29). However, let-7a has not been indicated
to be related to liver fibrosis. Here, we investigated the
expression level of let-7a in patients with liver fibrosis to
reveal the molecular mechanism of let-7a regulating the
pathogenesis of liver fibrosis. Our study aimed to provide an
accurate target and vital evidence to better understand the
underlying pathogenesis for early diagnosis and treatment for liver
fibrosis.
Patients and methods
Human liver samples
Liver samples were collected from patients and
volunteers attending The Second Affiliated Hospital of Qiqihar
Medical University (Qiqihar, China) from January, 2016 to March,
2017. Fibrotic liver samples were taken from 12 non-tumorous
patients (6 males and 6 females; age range 20–40 years) who were
diagnosed to have liver fibrosis. The tissues of patients were
taken and after pathological examination the remaining tissues were
treated with RNAKeeper (Vazyme Biotech Co., Ltd., Nanjing, China)
and stored at −80°C (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). Normal liver tissues (n=12) were obtained from healthy liver
transplant donors undergoing hepatic resection and regarded as
controls in this study. All clinical samples were collected in The
Second Affiliated Hospital of Qiqihar Medical University.
This study was approved by the Clinical Ethics
Committee of The Second Affiliated Hospital of Qiqihar Medical
University. All participants agreed with the research, participated
in this study willingly and signed an informed consent before
sample collection.
The control tissues and liver fibrosis tissues were
stored at −80°C for further analysis. According to the pathological
results of liver biopsy, the cases of the patients with liver
fibrosis was 4 (S0), 4 (S1), 2 (S2), 1 (S3), 1 (S4). According to
the different stage, the value of acoustic radiation force impulse
(ARFI) of the patients with liver fibrosis was 0.98±0.10,
1.14±0.11, 1.38±0.40, 1.91±0.70 and 2.12±0.75 m/sec,
respectively.
Collection of human blood samples
Blood samples were obtained from patients and
volunteers in The Second Affiliated Hospital of Qiqihar Medical
University from January, 2016 to October, 2017. Human blood samples
were collected from 22 hospitalized patients who had liver fibrosis
(11 males and 11 females; age range 21–40 years) and 22 age- and
sex-matched healthy subjects according to a protocol approved by
the Institutional Review Board. The healthy volunteers who suffered
from known acute or chronic diseases, who took medications or
alcohol within 24 h or were aged less than 21 years were excluded.
All human experiments were conducted according to the clinical
Εthics Committee of The Second Affiliated Hospital of Qiqihar
Medical University.
RNA extraction and cDNA synthesis
The frozen liver samples were thawed and homogenized
at 4°C prior to RNA isolation. TRIzol reagent (Thermo Fisher
Scientific, Inc.) was used to isolate the total RNAs from the liver
samples, blood samples and cells according to the protocols. After
isolation, RNAs were quantified by Nanodrop machine (Thermo Fisher
Scientific, Inc.) and RNAs were used in further experiments when
the concentration was higher than 200 (ng/µl) and A260/280 ratio
was 1.7–2.0. Single-stranded cDNAs were synthesized using reverse
transcription kit (Vazyme Biotech Co., Ltd.). For cDNA synthesis, a
10 µl reaction volume composed of 500 ng RNAs, 4 µl reverse
transcriptase and double-distilled water (ddH2O) were
added into 200 µl EP tubes. The reactions were initially incubated
at 50°C for 15 min and then at 85°C for 5 sec.
qPCR
qPCR analysis was performed by using 10 µl SYBR
Green PCR Master Mix (Vazyme Biotech Co., Ltd.), 7 µl
ddH2O, 1 µl cDNA and 2 µl sequence-specific primers
under RT-qPCR machine (Roche Diagnostics GmbH, Mannheim, Germany).
The mixtures were placed in a 96-well plate and the conditions
were: 95°C for 10 sec, 40 cycles at 95°C for 10 sec and 60°C for 30
sec. The relative expression levels of target genes were quantified
by a LightCycler 480 System (Roche Diagnostics GmbH) on the basis
of the threshold cycle (CT). U6 was regarded as a control gene for
the expression of miRNAs. The expression of GAPDH was used to
normalize the expression of other genes. All primers used in the
present study were designed and synthesized by GenePharma,
Shanghai, China. Primer sequences are listed in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Primers | Forward | Reverse |
|---|
| U6 |
GCTTCGGCAGCACATATACT |
TTCACGAATTTGCGTGTCAT |
| GAPDH |
TCCACTGGCGTCTTCACC |
GGCAGAGATGATGACCCTTTT |
| hsa-let-7a |
CGATTCAGTGAGGTAGTAGGTTGT |
TATGGTTGTTCTGCTCTCTGTCTC |
| let-7a |
GCGCCTGAGTAGTTG |
CAGGGGGGGTCCGAGGT |
| Col-1 |
GATTGAGAACATCCGCAGC |
CATCTTGAGGTCACGGCAT |
| Col-4 |
ATCTCTGCCAGGACCAAGTG |
CGGGCTGACATTCCACAAT |
| α-SMA |
GTCCCAGACATCAGGGAGTAA |
TCGGATACTTCAGCGTCAGGA |
Cell culture
The HSC cell line LX-2 cells (cat. no. GD-C624092)
were obtained from Changsha Yingrun Biotechnologies Co., Ltd.
(Changsha, China). The cells were cultured in DMEM medium (Hyclone;
GE Healthcare Life Sciences, Logan, MT, USA) containing 10% fetal
bovine serum (FBS; Thermo Fisher Scientific, Inc.), 100 µg/ml
penicillin-streptomycin antibiotics (cat. no. C0222; Beyotime
Institute of Biotechnology, Haimen, China) and 2 µM of L-glutamine
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The cells were
seeded in 25 cm2 flasks and grown in an incubator with
5% CO2 and 37°C (Thermo Fisher Scientific, Inc.). The
cells were plated at a density of 5×105 cells/well and
routinely subcultured using 0.25% trypsin when the confluence
reached approximately 80%. The method of quantification was the
2−ΔΔCq method (30).
Cell transfection
To overexpress or silence let-7a, let-7a mimics,
let-7a inhibitor, miR-NC and antagomir-NC were transfected into
LX-2 cells. Transient transfections were carried out using
transfection reagent X-treme (Vazyme Biotech Co., Ltd.) on the
basis of previous studies. On the day prior to cell transfection,
LX-2 cells were plated in 96-well or 6-well plates. Let-7a mimics,
let-7a inhibitor, miR-NC and antagomir-NC were designed and
synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China). The
concentration of let-7a-5p mimics and miR-NC was 100 nM. The
concentration of let-7a-5p inhibitor and antagomir-NC was 50 nM.
After 24 h transfection the cells were used for further
experiments.
Cell viability analysis
LX-2 cells were placed at a density of
2×103 cells/well in 96-well plates (Corning
Incorporated, Corning, NY, USA). After culture of 24 h, 20 µl
volume of MTT solution (5 mg/ml, BioSharp, Hefei, China) was added
and the cells were incubated for 4 h at 37°C in an incubator. Then
about 150 µl of dimethyl sulfoxide (DMSO) was added and the plates
were gently shaken. The optical density values (λ=490 nm) were
detected using the enzyme linked immunosorbent assay. All
experiments were repeated at least three times.
TUNEL staining
To measure the apoptosis levels of LX-2 cells,
1×105 LX-2 cells were grown in a small dish. The cells
transfected with let-7a mimics, let-7a inhibitor, miR-NC and
antagomir-NC were used for TUNEL staining. The culture medium in
the dish was discarded, and cells were fixed in 4% PFA for 30 min
at room temperature after being washed with PBS. After the closure
and penetration procedures, TUNEL staining solution Vial1 and Vial2
were mixed at a ratio of 1:9 according to the instructions of the
kit. Then, cells were incubated with the TUNEL reagent for 1 h at
room temperature in the dark. After being washed with PBS, cells
were incubated with Hoechst staining solution for 20 min at room
temperature, and kept in the dark. Finally, the cells were observed
under a fluorescence microscope (Nikon Corporation, Tokyo, Japan).
Ten images were randomly captured of every angle. The
quantification of TUNEL staining was performed by measuring the
percentage of TUNEL-positive cells.
Western blotting
Cells after transfection with let-7a were lysed in
500 µl volume of RIPA lysis buffer for 20 min in a 4°C freezer.
After extraction, the concentration of total proteins was measured
using BCA protein detection kit (Beyotime Institute of
Biotechnology). After heat denaturation at 100°C for 10 min, equal
amounts of extracted protein samples were separated by 8–10%
SDS-PAGE and transferred onto PVDF membrane (EMD Millipore,
Billerica, MA, USA). After using 5% bovine serum albumin
(Sigma-Aldrich; Merck KGaA) to block non-specific protein binding
at room temperature for 1 h, the membrane was incubated in
anti-TGF-β or anti-SMAD antibody with a gentle shaking overnight at
4°C. Then, the membrane was incubated with secondary antibody and
the signal was detected using an ECL system. The expression of
GAPDH was used as loading control to quantify the expression of
TGF-β and SMAD by using Image Studio software. For western blot
analysis, anti-TGF-β (anti-rabbit; cat. no. CST3711; 1:1,000)
anti-SMAD2 (anti-rabbit, cat. no. CST5339; 1:1,000), anti-SMAD3
(anti-rabbit, cat. no. 9513; 1:1,000) antibodies, the second
antibody (anti-rabbit; cat. no. CST7074; 1:1,000), or the second
antibody (anti-mouse; cat. no. 5571; 1:1,000) all from Cell
Signaling Technology, Inc., (Danvers, MA, USA) and GAPDH
(anti-mouse; cat. no. sc-47724; 1:500) from Santa Cruz
Biotechnology, Inc., (Dallas, TX, USA).
Construction model of liver
fibrosis
To generate the model of liver fibrosis, 20 female
C57BL/6 mice (approximately 18–22 g) were purchased. Mice were kept
at 25°C and raised for a week before experiments. Mice were
intraperitoneally injected with olive oil vehicle (Sigma-Aldrich;
Merck KGaA) and they served as the control. In addition, mouse
model with liver fibrosis was intraperitoneally injected with CCl4
(0.5 µl/g, Sigma-Aldrich; Merck KGaA) which was normally used as
hepatotoxic chemical twice weekly. After injections of 10 weeks,
the mice were sacrificed and the liver tissues and blood samples
were harvested.
Statistical analysis
The data were analyzed from three or more
independent experiments. Analysis was performed by using GraphPad
Prism 5 software (GraphPad Software, Inc., La Jolla, CA, USA).
Differences between the groups were analyzed by one-way ANOVA,
followed by a Tukey's test for multiple comparisons. The
Mann-Whitney U test was used for comparison between groups.
P<0.05, P<0.01 and P<0.001 was considered statistically
significant.
Results
Expression of let-7a in patients with
liver fibrosis
To determine whether let-7a is related to liver
fibrosis, the expression of let-7a was detected in the tissues and
blood samples from patients with liver fibrosis using RT-qPCR
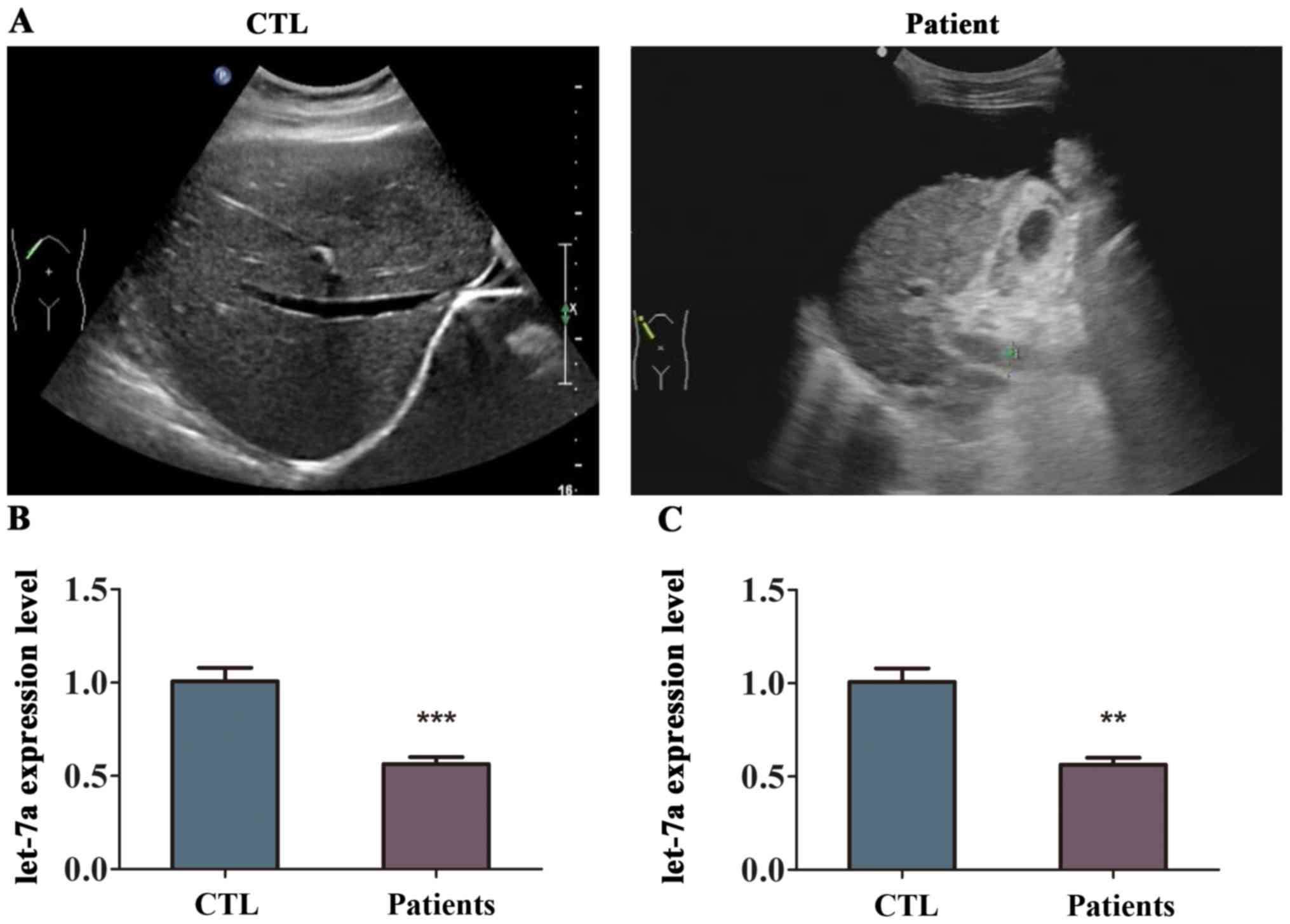
analysis. First, the analysis of supersonic inspection showed that
the liver volume of patient with liver fibrosis was decreased and
the tunica of liver was damaged compared with healthy volunteers
(Fig. 1A). The echo in the liver
from patient with liver fibrosis become deeper and weaker with
uneven distribution, and the analysis showed that the hepatic vein
was not clear (Fig. 1A). The level
of let-7a was differentially expressed in the liver fibrosis
tissues compared to normal liver tissues as indicated in Fig. 1B. The results of RT-qPCR revealed
that the expression of let-7a was lower in the liver tissues from
patients with liver fibrosis compared with that in healthy
volunteers, and the difference was statistically significant
(Fig. 1B). In addition, let-7a was
markedly downregulated in the blood samples of patients with liver
fibrosis (Fig. 1C). These results
suggested that let-7a was reduced in patients with liver
fibrosis.
Validation of let-7a expression in
mice with liver fibrosis
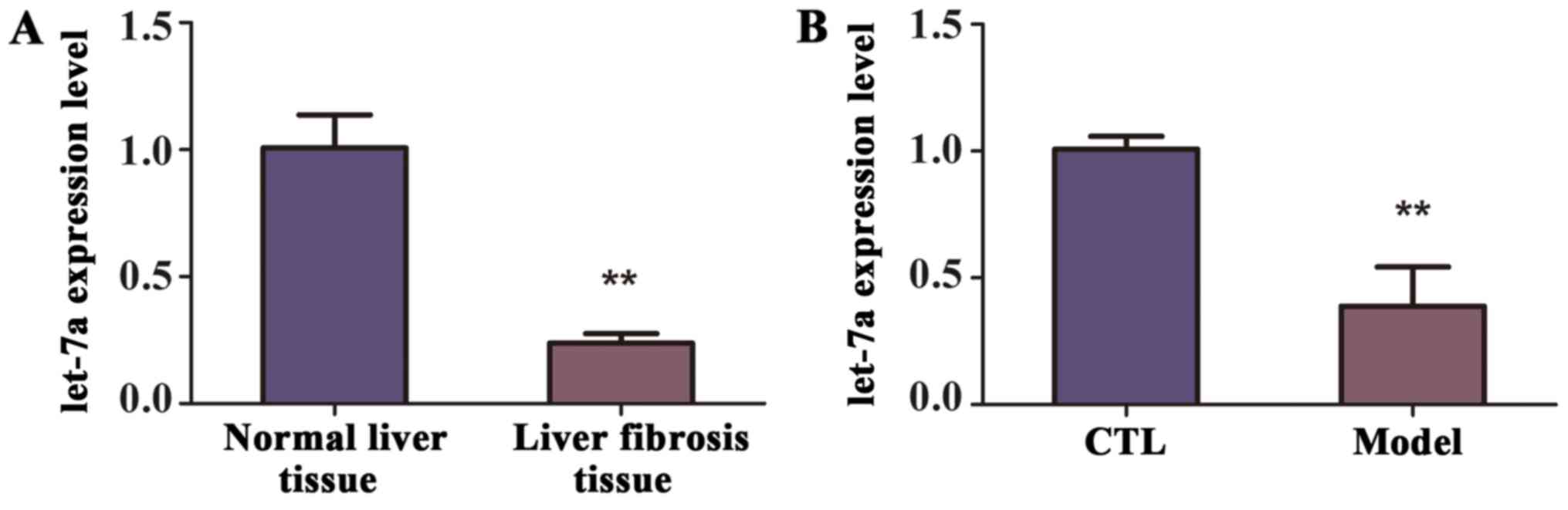
To identify the level of let-7a in the mice with
liver fibrosis, a model of liver fibrosis was generated. Briefly,
mice (n=10) were intraperitoneally injected CCl4 twice weekly for
10 weeks, while the control group (n=10) were treated with olive
oil. The decreased expression of let-7a in the liver tissues from
mice of liver fibrosis was revealed by RT-qPCR (Fig. 2A). Similarly, according to the
RT-qPCR analysis, the expression of let-7a was observably decreased
in the blood samples of mice with liver fibrosis compared with
control group, and the difference was statistically significant
(Fig. 2B). Therefore, we concluded
that let-7a was reduced in the liver tissues and blood samples of
mice with liver fibrosis.
The role of let-7a in the activation
of HSCs
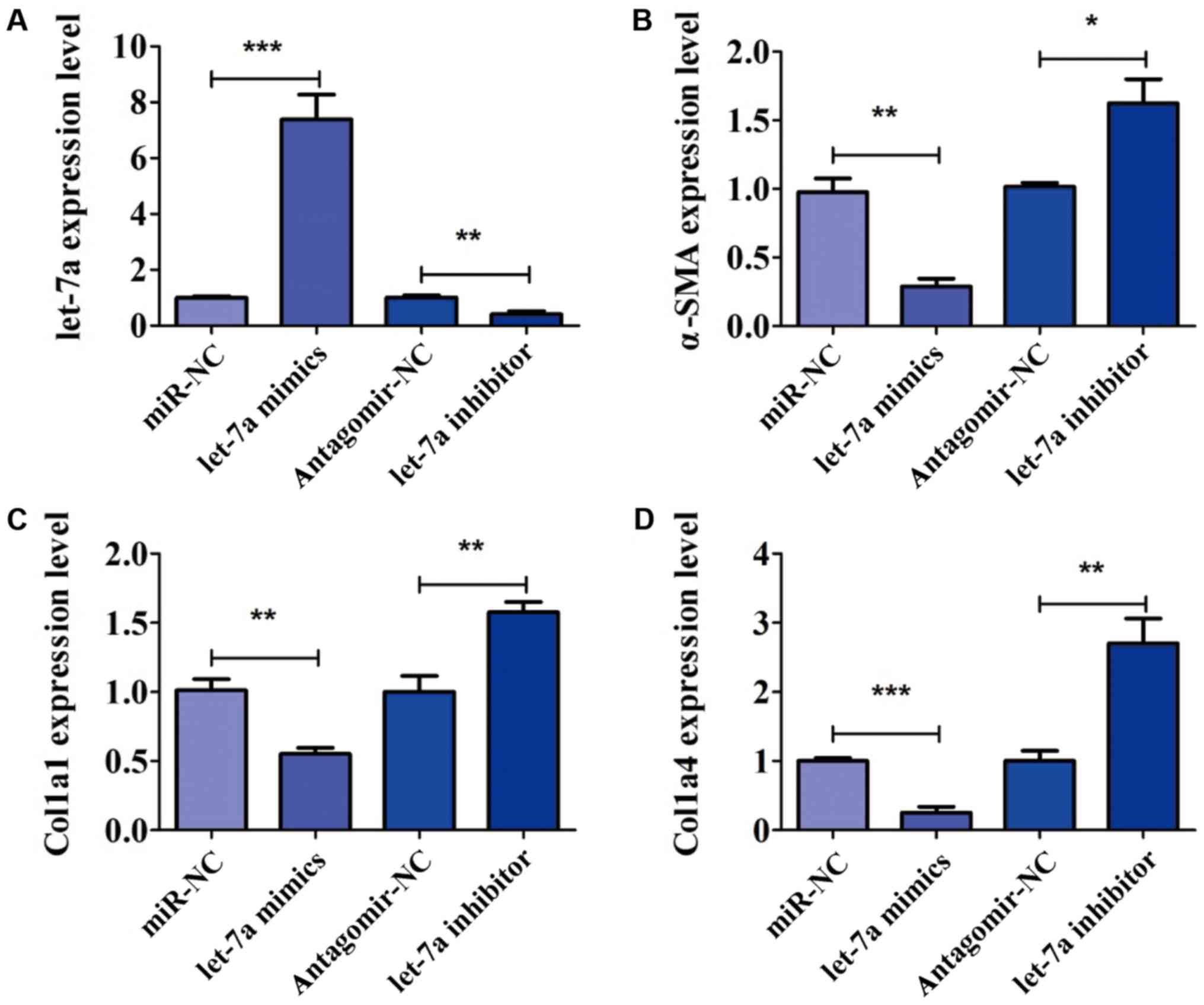
To further confirm the role of let-7a in the
activation of HSCs, LX-2 cells were transfected with let-7a mimics,
let-7a inhibitor, miR-NC and antagomir-NC for 24 h. The expression
of let-7a was detected by PCR analysis. The expression of let-7a
was significantly increased after transfection of let-7a mimics,
but decreased in the presence of let-7a inhibitor (Fig. 3A). After transfection, the expression
of markers of HSC activation, including α-SMA, Col1a1 and Col1a4
was analyzed. We found that overexpression of let-7a significantly
decreased the expression level of α-SMA, while the decrease in
let-7a elevated the level of α-SMA (Fig.
3B). RT-qPCR assay indicated that LX-2 cells transfected with
let-7a mimics decreased level of Col1a1, and the expression of
Col1a1 was increased by transfection of let-7a inhibitor (Fig. 3C). In addition, the expression of
Col1a4 was inhibited by let-7a mimics in comparison with miR-NC
(Fig. 3D). Furthermore, the
expression level of Col1a4 was higher in cells treated with let-7a
inhibitor than that in group antagomir-NC (Fig. 3D). The above results confirmed that
let-7a led to a marked decrease of HSC activation.
The effect of let-7a on cell viability
and apoptosis of HSCs
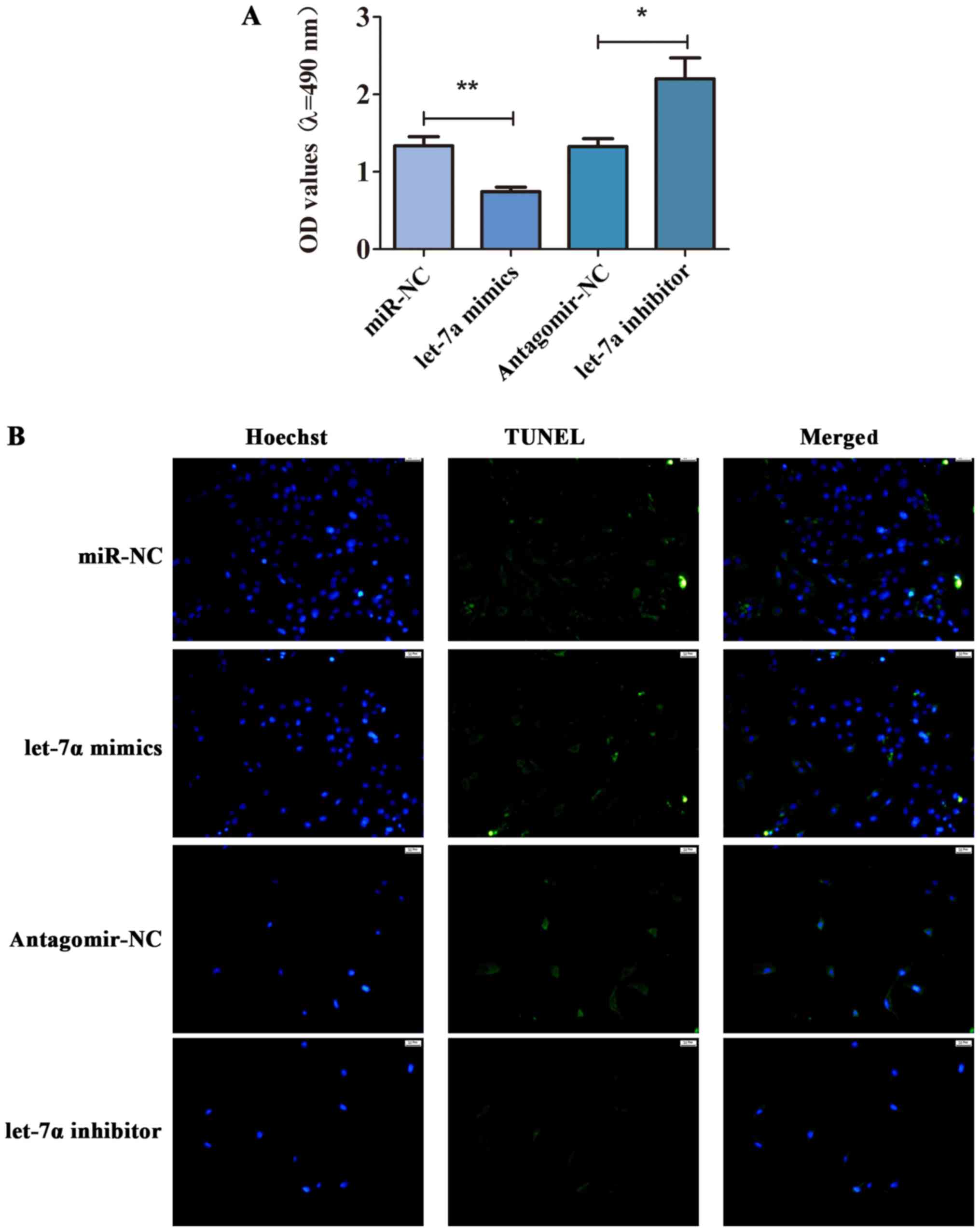
To further study the role of let-7a in the cell
viability and apoptosis of HSCs, MTT assay and TUNEL staining were
performed. As shown in Fig. 4A,
let-7a was overexpressed, and the cell viability was inhibited.
Compared with antagomir-NC, the cell viability of HSCs transfected
with let-7a inhibitor was enhanced (Fig.
4A). Besides, TUNEL staining displayed that let-7a promoted
apoptosis of HSCs, while let-7a inhibitor decreased the apoptotic
cells (Fig. 4B). These observations
implied that let-7a inhibited the cell viability but promoted
apoptosis in HSCs.
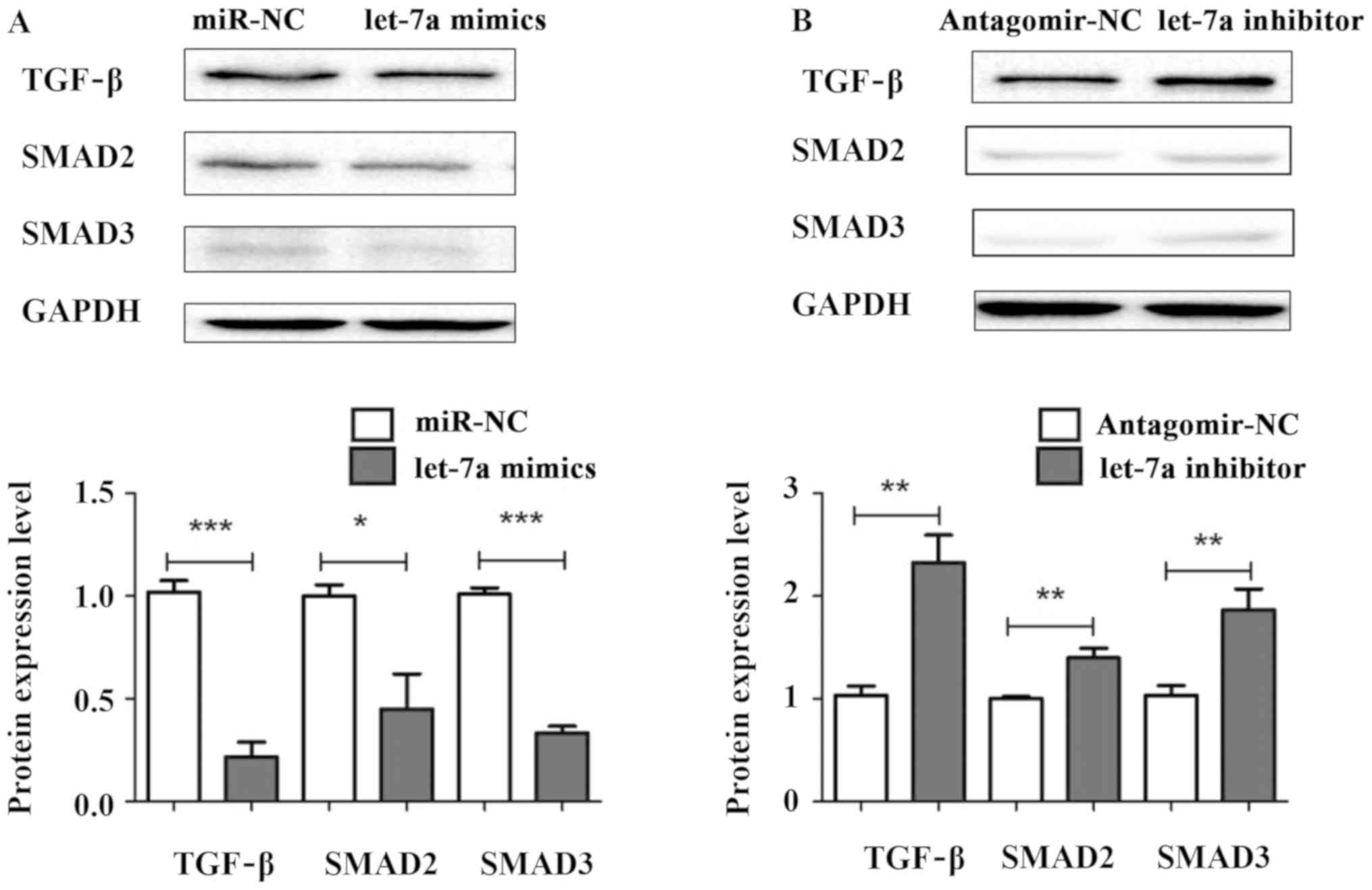
Let-7a suppresses liver fibrosis
through TGF-β/SMAD signaling pathway
To explore the underlying mechanism of let-7a
regulating liver fibrosis, let-7a mimics, let-7a inhibitor, miR-NC
and antagomir-NC were transiently transfected into HSCs. The
protein expression levels of TGF-β, SMAD2 and SMAD3 were measured
by western blotting. The results showed that the expression levels
of TGF-β, SMAD2 and SMAD3 were decreased in HSCs transfected with
let-7a mimics compared with miR-NC (Fig.
5A). Conversely, the protein levels of TGF-β, SMAD2 and SMAD3
were significantly elevated in cells after transfection with let-7a
inhibitor (Fig. 5B). Overall, our
results indicate that let-7a may regulate the activation, cell
viability and apoptosis of HSCs through TGF-β/SMAD signaling
pathway.
Discussion
Fibrosis is a complicated process involving HSC
activation, deposition of extracellular matrix and genetic and
epigenetic changes including miRNA. Liver fibrosis is caused by
viral hepatitis and alcoholic steatohepatitis, which leads to
cirrhosis and hepatocellular carcinoma (31,32).
Emerging studies show that HSC is pivotal event in liver fibrosis
(33,34). Therefore, in this study, our purpose
is to investigate the role of let-7a in hepatic stellate cells and
whether let-7a is dysregulated in the normal liver tissues and
liver fibrosis.
MicroRNAs (miRNAs) are a type of endogenous small
non-coding RNAs, and microRNAs are related to the development of
various diseases (35,36). miRNAs have been found to play a
pro-fibrotic or anti-fibrotic role in HSC activation and be in
connection with liver fibrosis. For example, miR-30a was
downregulated in fibrotic liver tissues and isolated HSCs, and
miR-30a suppresses HSC activation including cell proliferation,
expression of α-SMA and collagen by targeting Snai1 protein
(37). A study has shown that
miR-185 suppressed fibrogenic activation of hepatic stellate cells
by suppressing HSC activation through targeting RHEB and RICTOR.
miR-212 promotes liver fibrosis via activating HSCs and TGF-β
pathway through targeting SMAD7, which highlights that miR-212 can
be regarded as a key biomarker or therapeutic target for liver
fibrosis (38).
In this study, the results indicated that let-7a was
markedly reduced in the liver tissues and blood samples from
patients with liver fibrosis compared with healthy volunteers in
clinic. In addition, the level of let-7a was decreased in liver
tissues and blood samples in mice with liver fibrosis which were
constructed by intraperitoneal injected with CCl4. Further analysis
revealed that overexpression of let-7a decreased the mRNA level of
markers of HSC activation, such as α-SMA, Col1a1 and Col1a4,
suggesting that let-7a inhibited the activation level of HSCs.
let-7a inhibitor increased the expression levels of these key
genes, which were measured by RT-qPCR analysis. Besides,
transfection of let-7a reduced cell viability and promoted
apoptosis of HSCs. In contrast, let-7a inhibitor increased e cell
viability and inhibited apoptosis of HSCs, which were in accordance
with our hypothesis. Moreover, western blot analysis showed that
let-7a might inhibit HSCs through TGFβ/SMAD signaling pathway. The
present study provides new insights into the mechanisms behind the
anti-fibrotic effect of let-7a. This study provided a potentially
accurate target and vital evidence to better understand the
underlying pathogenesis for early diagnosis and treatment of liver
fibrosis.
Acknowledgements
Not applicable.
Funding
Funding was received by Education Department of
Heilongjiang Province General Program (12531788).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YinghuiZ, YingboZ, JG, YL and KJ drafted the
manuscript. YinghuiZ collected human blood samples. JG and YL
helpedwith RNA extraction and cDNA synthesis. YinghuiZ and KJ
performed PCR. YingboZ contributed to cell viability analysis.
YinghuiZ and YingboZ wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Clinical Ethics
Committee of The Second Affiliated Hospital of Qiqihar Medical
University (Qiqihar, China). All participants agreed with the
research, participated in this study willingly and signed an
informed consent before sample collection.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
He X, Sun Y, Lei N, Fan X, Zhang C, Wang
Y, Zheng K, Zhang D and Pan W: MicroRNA-351 promotes
schistosomiasis-induced Liver fibrosis by targeting the vitamin D
receptor. Proc Natl Acad Sci USA. 115:180–185. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cao Q, Zhu X, Zhai X, Ji L, Cheng F, Zhu
Y, Yu P and Zhou Y: Leptin suppresses microRNA-122 promoter
activity by phosphorylation of foxO1 in hepatic stellate cell
contributing to leptin promotion of mouse liver fibrosis. Toxicol
Appl Pharmacol. 339:143–150. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chandel R, Saxena R, Das A and Kaur J:
Association of rno-miR-183-96-182 cluster with diethyinitrosamine
induced liver fibrosis in Wistar rats. J Cell Biochem.
119:4072–4084. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Afonso MB, Rodrigues PM, Simão AL, Gaspar
MM, Carvalho T, Borralho P, Bañales JM, Castro RE and Rodrigues
CMP: miRNA-21 ablation protects against liver injury and
necroptosis in cholestasis. Cell Death Differ. 25:857–872. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen Y, Ou Y, Dong J, Yang G, Zeng Z, Liu
Y, Liu B, Li W, He X and Lan T: Osteopontin promotes collagen I
synthesis in hepatic stellate cells by miRNA-129-5p inhibition. Exp
Cell Res. 362:343–348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wan Y, McDaniel K, Wu N, Ramos-Lorenzo S,
Glaser T, Venter J, Francis H, Kennedy L, Sato K, Zhou T, et al:
Regulation of cellular senescence by miR-34a in alcoholic liver
injury. Am J Pathol. 187:2788–2798. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brea R, Motiño O, Francés D, García-Monzón
C, Vargas J, Fernández-Velasco M, Boscá L, Casado M, Martín-Sanz P
and Agra N: PGE2 induces apoptosis of hepatic stellate cells and
attenuates liver fibrosis in mice by downregulating miR-23a-5p and
miR-28a-5p. Biochim Biophys Acta Mol Basis Dis. 1864:325–337. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cai Y, Huang G, Ma L, Dong L, Chen S, Shen
X and Zhang S, Xue R, Sun D and Zhang S: Smurf2, an E3 ubiquitin
ligase, interacts with PDE4B and attenuates liver fibrosis through
miR-132 mediated CTGF inhibition. Biochim Biophys Acta Mol Cell
Res. 1865:297–308. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Caviglia JM, Yan J, Jang MK, Gwak GY, Affo
S, Yu L, Olinga P, Friedman RA, Chen X and Schwabe RF: MicroRNA-21
and Dicer are dispensable for hepatic stellate cell activation and
the development of liver fibrosis. Hepatology. 67:2414–2429. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen Y, Yang S, Peng Y and Yang Z: The
regulatory role of IL-6R in hepatitis B-associated fibrosis and
cirrhosis. Braz J Med Biol Res. 50:e62462017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tang N, Wu Y, Cao W, Liang Y, Gao Y, Hu L,
Yang Q, Zhou Y, Tang F and Xiao J: Lentivirus-mediated
over-expression of let-7b microRNA suppresses Liver fibrosis in the
mouse infected with Schistosoma japonicum. Exp Parasitol.
182:45–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen W, Zhao W, Yang A, Xu A, Wang H, Cong
M, Liu T, Wang P and You H: Integrated analysis of microRNA and
gene expression profiles reveals a functional regulatory module
associated with liver fibrosis. Gene. 636:87–95. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma X, Luo Q, Zhu H, Liu X, Dong Z, Zhang
K, Zou Y, Wu J, Ge J and Sun A: Aldehyde dehydrogenase 2 activation
ameliorates CCl4-induced chronic liver fibrosis in mice by
up-regulating Nrf2/HO-1 antioxidant pathway. J Cell Mol Med.
22:3965–3978. 2018. View Article : Google Scholar
|
|
14
|
Wang XC, Gusdon AM, Liu H and Qu S:
Effects of glucagon-like peptide-1 receptor agonists on
non-alcoholic fatty liver disease and inflammation. World J
Gastroenterol. 20:14821–14830. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Patel V, Joharapurkar A, Kshirsagar S,
Sutariya B, Patel M, Patel H, Pandey D, Patel D, Ranvir R, Kadam S,
et al: Coagonist of GLP-1 and glucagon receptor ameliorates
development of non-alcoholic fatty liver disease. Cardiovasc
Hematol Agents Med Chem. 16:35–43. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ye Y, Li Z, Feng Q, Chen Z, Wu Z, Wang J,
Ye X, Zhang D, Liu L, Gao W, et al: Downregulation of microRNA-145
may contribute to liver fibrosis in biliary atresia by targeting
ADD3. PLoS One. 12:e01808962017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Feng GX, Li J, Yang Z, Zhang SQ, Liu YX,
Zhang WY, Ye LH and Zhang XD: Hepatitis B virus X protein promotes
the development of liver fibrosis and hepatoma through
downregulation of miR-30e targeting P4HA2 mRNA. Oncogene.
36:6895–6905. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tao R, Fan XX, Yu HJ, Ai G, Zhang HY, Kong
HY, Song QQ, Huang Y, Huang JQ and Ning Q: MicroRNA-29b-3p prevents
Schistosoma japonicum-induced liver fibrosis by targeting COL1A1
and COL3A1. J Cell Biochem. 119:3199–3209. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen J, Yu Y, Li S, Liu Y, Zhou S, Cao S,
Yin J and Li G: MicroRNA-30a ameliorates Liver fibrosis by
inhibiting Beclin1-mediated autophagy. J Cell Mol Med.
21:3679–3692. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun J, Zhang H, Li L, Yu L and Fu L:
MicroRNA-9 limits Liver fibrosis by suppressing the activation and
proliferation of hepatic stellate cells by directly targeting
MRP1/ABCC1. Oncol Rep. 37:1698–1706. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schueller F, Roy S, Loosen SH, Alder J,
Koppe C, Schneider AT, Wandrer F, Bantel H, Vucur M, Mi QS, et al:
miR-223 represents a biomarker in acute and chronic liver injury.
Clin Sci (Lond). 131:1971–1987. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Demerdash HM, Hussien HM, Hassouna E and
Arida EA: Detection of microRNA in hepatic cirrhosis and
hepatocellular carcinoma in hepatitis C genotype-4 in Egyptian
patients. BioMed Res Int. 2017:18060692017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fittipaldi S, Vasuri F, Bonora S,
Degiovanni A, Santandrea G, Cucchetti A, Gramantieri L, Bolondi L
and D'Errico A: miRNA signature of hepatocellular carcinoma
vascularization: How the controls can influence the signature. Dig
Dis Sci. 62:2397–2407. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu N, Meng F, Zhou T, Han Y, Kennedy L,
Venter J, Francis H, DeMorrow S, Onori P, Invernizzi P, et al:
Prolonged darkness reduces liver fibrosis in a mouse model of
primary sclerosing cholangitis by miR-200b down-regulation. FASEB
J. 31:4305–4324. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sene LB, Rizzi VHG, Gontijo JAR and Boer
PA: Gestational low-protein intake enhances whole-kidney miR-192
and miR-200 family expression and epithelial-to-mesenchymal
transition in rat adult male offspring. J Exp Biol. 221:2212018.
View Article : Google Scholar
|
|
26
|
Wu Y, Liu Y, Pan Y, Lu C, Xu H, Wang X,
Liu T, Feng K and Tang Y: MicroRNA-135a inhibits cardiac fibrosis
induced by isoproterenol via TRPM7 channel. Biomed Pharmacother.
104:252–260. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Glasgow AMA, De Santi C and Greene CM:
Non-coding RNA in cystic fibrosis. Biochem Soc Trans. 46:619–630.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Miao C, Xiong Y, Zhang G and Chang J:
MicroRNAs in idiopathic pulmonary fibrosis, new research progress
and their pathophysiological implication. Exp Lung Res. 44:178–190.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lai CY, Lin CY, Hsu CC, Yeh KY and Her GM:
Liver-directed microRNA-7a depletion induces nonalcoholic fatty
liver disease by stabilizing YY1-mediated lipogenic pathways in
zebrafish. Biochim Biophys Acta Mol Cell Biol Lipids. 1863:844–856.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Watany MM, Hagag RY and Okda HI:
Circulating miR-21, miR-210 and miR-146a as potential biomarkers to
differentiate acute tubular necrosis from hepatorenal syndrome in
patients with liver cirrhosis: A pilot study. Clin Chem Lab Med.
56:739–747. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shi SQ, Ke JJ, Xu QS, Wu WQ and Wan YY:
Integrated network analysis to identify the key genes,
transcription factors, and microRNAs involved in hepatocellular
carcinoma. Neoplasma. 65:66–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schueller F, Roy S, Vucur M, Trautwein C,
Luedde T and Roderburg C: The role of miRNAs in the pathophysiology
of liver diseases and toxicity. Int J Mol Sci. 19:192018.
View Article : Google Scholar
|
|
34
|
Lin YC, Wang FS, Yang YL, Chuang YT and
Huang YH: MicroRNA-29a mitigation of toll-like receptor 2 and 4
signaling and alleviation of obstructive jaundice-induced fibrosis
in mice. Biochem Biophys Res Commun. 496:880–886. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Austermann C, Schierwagen R, Mohr R,
Anadol E, Klein S, Pohlmann A, Jansen C, Strassburg CP,
Schwarze-Zander C, Boesecke C, et al: microRNA-200a: A
stage-dependent biomarker and predictor of steatosis and liver cell
injury in human immunodeficiency virus patients. Hepatol Commun.
1:36–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu W, Wang Y, Zhang D, Yu X and Leng X:
MiR-7-5p functions as a tumor suppressor by targeting SOX18 in
pancreatic ductal adenocarcinoma. Biochem Biophys Res Commun.
497:963–970. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zheng J, Wang W, Yu F, Dong P, Chen B and
Zhou MT: MicroRNA-30a suppresses the activation of hepatic stellate
cells by inhibiting epithelial-to-mesenchymal transition. Cell
Physiol Biochem. 46:82–92. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhu J, Zhang Z, Zhang Y, Li W, Zheng W, Yu
J, Wang B, Chen L, Zhuo Q, Chen L, et al: MicroRNA-212 activates
hepatic stellate cells and promotes liver fibrosis via targeting
SMAD7. Biochem Biophys Res Commun. 496:176–183. 2018. View Article : Google Scholar : PubMed/NCBI
|