Introduction
Multiple myeloma (MM), the second most common
hematologic malignant tumor, is characterized by the clonal
proliferation and accumulation of plasma cells (PCs) in bone marrow
(1). The morbidity of MM is
0.5–1/100,000 in Asia, whereas the morbidity in Africa and America
is 10–12/100,000 (2). Currently,
immunomodulatory medicine, proteasome inhibitors and autologous
stem cell transplantation are primary therapeutic strategies for
patients with MM (3). The techniques
used for the diagnosis and therapy of patients with MM have
recently progressed; however, the clinical outcomes of patients
with MM remain poor with a 5-year survival rate of only 30–40%
remaining (4). Further research is
therefore required to understand the mechanisms underlying MM
pathogenesis and to develop novel treatment options for patients
with this disease.
microRNAs (miRNAs) are endogenous, non-coding, short
RNA molecules that are ~22 nucleotides in length (5). miRNAs are able to silence gene
expression by directly interacting with complementary sites within
the 3′-untranslated regions (3′-UTRs) of their target gene to cause
mRNA degradation and/or translational suppression (6). Each miRNA may regulate hundreds of
different genes. Thus implicating the regulation of a wide range of
biological processes, including cell proliferation, cycle,
apoptosis, invasion, epithelial-mesenchymal transition, metastasis
and drug resistance (7–9). Previous studies have disclosed the
importance of miRNAs in MM formation and progression (10–12). A
variety of miRNAs have been revealed to be dysregulated in MM
(13–15). For example, miR-338-3p (16), miR-324-5p (17) and miR-320c (18) are downregulated in MM; whereas
miR-19a (19), miR-32 (20), and miR-210 (21) are upregulated in MM. Dysregulated
miRNAs may serve as oncogenes or tumor suppressors, thus
contributing to MM malignant progression (22,23).
Therefore, further clarification concerning the expression pattern,
roles and underlying molecular mechanisms of miRNAs in MM would
provide novel clinical intervention tools for patients with this
fatal malignancy.
miR-765 is reportedly abnormally expressed and
contributes to the tumorigenesis of many different types of human
cancer, such as esophageal squamous cell carcinoma (24), osteosarcoma (25) and hepatocellular carcinoma (26). However, the expression pattern,
specific roles and underlying mechanism of miR-765 in MM remain
largely unknown. The expression of miR-765 in MM cell lines and MM
patient plasma was detected in the present study. In addition, the
detailed roles and associated mechanisms of miR-765 in MM were
examined. Data obtained in the present study may aid the
elucidation of the functional roles of miR-765 in MM carcinogenesis
and progression.
Materials and methods
Clinical specimens
Bone marrow aspirates were collected from 27 MM
patients (15 males, 12 females; age range, 23–57 years) and 11
healthy individuals (7 males, 4 females; age range, 36–52 years) at
The First Affiliated Hospital of Nanchang University, Nanchang,
P.R. China between June 2015 and August 2017. Patients treated with
radiotherapy, chemotherapy, immunomodulatory medicine, proteasome
inhibitors and autologous stem cell transplantation were excluded
from the current study. Plasma cells were purified from bone marrow
aspirates using CD138 MicroBeads (cat. no. 130-051-301; Miltenyi
Biotec GmbH) in accordance with manufacturer's protocol. Plasma
cells were quickly frozen in liquid nitrogen and then maintained at
−80°C. The current study was approved by the Ethics Committee of
The First Affiliated Hospital of Nanchang University (Nanchang,
China). All participants provided written informed consent prior to
enrollment.
Cell lines
Three human MM cell lines (U266, MM1S and RPMI-8226)
were purchased from the American Type Culture Collection. RPMI-1640
medium supplemented with 10% fetal bovine serum and 1%
penicillin/streptomycin (all, Gibco; Thermo Fisher Scientific,
Inc.) was used to culture all MM cell lines. Cells were maintained
at 37°C in a humidified chamber supplied with 5%
CO2.
Transfection assay
A miR-765 inhibitor (5′CAUCACCUUCCUUCUCCUCCA3′), a
corresponding negative control miRNA inhibitor (NC inhibitor;
5′ACUACUGAGUGACAGUAGA3′), a small interfering RNA (siRNA) targeting
SOX6 expression (SOX6 siRNA; 5′GCAGGAAUUUGGACACCUU3′) and a
negative control siRNA (NC siRNA; 5′UUCUCCGAACGUGUCACGUTT3′) were
ordered from Shanghai GenePharma Co., Ltd. Cells were plated into
six-well plates at a density of 5×105 cell per well and
cultured overnight at 37°C. Cell transfection was performed using
Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) in accordance with the manufacturer's protocol. Reverse
transcription-quantitative (RT-q) PCR and flow cytometric analysis
of cell apoptosis were performed 48 h following transfection. A
Cell counting kit-8 (CCK-8) assay and western blot analysis were
conducted at 24 and 72 h post-transfection, respectively.
RT-q PCR
Total RNA was extracted from tissues or cells using
a TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. A NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc.) was used to
determine the concentration of total RNA. To detect miR-765
expression, cDNA was synthesized from total RNA using a TaqMan
MicroRNA Reverse Transcription kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The temperature protocol for reverse
transcription was as follows: 16°C for 30 min, 42°C for 30 min and
85°C for 5 min. qPCR was then performed using a TaqMan MicroRNA PCR
kit with an Applied Biosystems 7500 Sequence Detection system (each
from Applied Biosystems; Thermo Fisher Scientific, Inc.). The
temperature protocol for qPCR were as follows: 50°C for 2 min, 95°C
for 10 min; 40 cycles of denaturation at 95°C for 15 sec; and
annealing/extension at 60°C for 60 sec. For the measurement of SOX6
mRNA expression, reverse transcription was performed using a
PrimeScript RT Reagent kit (Takara Biotechnology Co., Ltd.). The
temperature protocol for reverse transcription was as follows: 37°C
for 15 min and 85°C for 5 sec. Synthesized cDNA was subsequently
used for qPCR using a SYBR Premix Ex Taq™ kit (Takara Biotechnology
Co., Ltd.). The temperature protocol for qPCR was as follows: 5 min
at 95°C, followed by 40 cycles of 95°C for 30 sec and 65°C for 45
sec. U6 small nuclear RNA and GAPDH served as an internal control
to normalize the relative expression level of miR-765 and SOX6
mRNA, respectively. Relative gene expression was calculated using
the 2−ΔΔCq method (27).
The following primers were utilized for qPCR: miR-765 forward,
5′-GUAGCCAAGGAATCCGAAGGA-3′ and reverse,
5′-GCGAGGAAGGAGGAGGAAGGT-3′; U6 forward 5′-CTCGCTTCGGCAGCACA-3′ and
reverse, 5′-AACGCTTCACGAATTTGCGT-3′; SOX6 forward,
5′-CCCGTACAGTTCATTCCGTC-3′ and reverse, 5′-AGCCTTGGGTTAATTTGTGG-3′;
GAPDH forward, 5′-CGGAGTCAACGGATTTGGTCGTAT-3′ and reverse,
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′.
Cell counting kit-8 (CCK-8) assay
Cellular proliferation was determined using a CCK-8
assay (Dojindo Molecular Technologies, Inc.) as per the
manufacturer's protocol. Transfected cells were collected at 24 h
post-transfection, suspended in RPMI-1640 medium and inoculated
into each well of a 96-well plate at a density of 3×103
cells. Cells were then incubated at 37°C with 5% CO2 at
different time points (0, 24, 48 and 72 h). A total of 10 µl CCK-8
solution was added into each well and incubated for 2 h at 37°C.
Absorbance at 450 nm was determined using a microplate reader
(Bio-Rad Laboratories, Inc.).
Flow cytometric analysis of cell
apoptosis
Following incubation for 48 h, transfected cells
were harvested and washed with PBS. The apoptosis rate of
transfected cells was examined using an FITC Apoptosis Detection
kit (Biolegend, Inc.) according to the manufacturer's protocol.
Transfected cells were resuspended in 100 µl of binding buffer
followed by staining with 5 µl of Annexin V–FITC and 5 µl of
propidium iodide. After 15 min incubation at room temperature in
the dark, stained cells were detected via flow cytometry FACScan
(BD Biosciences). CellQuest version 5.1 (BD Biosciences) software
was used to analyze data.
Bioinformatics analysis and luciferase
reporter assay
The putative targets of miR-765 were predicted using
two microRNA target prediction websites: TargetScan (http://targetscan.org/) and miRDB (http://mirdb.org/). Luciferase reporter plasmids,
including pMIR-SOX6-3′-UTR wild type (wt 1 and 2) and
pMIR-SOX6-3′-UTR mutant (mut 1 and 2), were chemically created by
Shanghai GenePharma Co., Ltd. For the reporter assay, cells were
inoculated into 24-well plates at a density of 1.0×105
cells/well 12 h prior to transfection. A miR-765 inhibitor or NC
inhibitor along with the pMIR-SOX6-3′-UTR wt or pMIR-SOX6-3′-UTR
mut was co-transfected into cells using the Lipofectamine 2000
reagent, based on the manufacturer's protocol. A dual-luciferase
reporter assay system (Promega Corporation) was applied to measure
the luciferase activity at 48 h post-transfection. The firefly
luciferase activity of each well was normalized to that of the
Renilla luciferase activity.
Western blot analysis
A Total Protein Extraction kit (Nanjing KeyGen
Biotech Co., Ltd.) was utilized to isolate total protein from
cultured cells. The concentration of total protein was determined
using a BCA Protein Assay kit (Pierce; Thermo Fisher Scientific,
Inc.). Equal quantities of protein were separated via 10% SDS-PAGE
and transferred to PVDF (Merck KGaA). Subsequent to 2 h blocking at
room temperature with 5% dried skimmed milk in TBST, membranes were
incubated with primary antibodies overnight at 4°C against SOX6
(1:1,000; cat. no. ab84880) or GAPDH (1:1,000; cat. no. ab9484;
each, Abcam). This was followed by further incubation with
horseradish peroxidase-conjugated secondary antibodies (1:6,000;
cat. no. ab6789; Abcam) for 2 h at room temperature. Protein
signals were detected using an enhanced chemiluminescent reagent
(GE Healthcare Life Sciences). GAPDH was used as a loading control.
Quantity One software version 4.62 (BioRad Laboratories, Inc.) was
utilized for the analysis of density.
Statistical analysis
SPSS 19 software package (IBM Corp.) was used for
statistical analysis. Differences between groups were analyzed
using a student's t-test or one-way ANOVA. A post-hoc
Student-Newman-Keuls test was then utilized. A Spearman's
correlation analysis was utilized to determine the correlation
between miR-765 and SOX6 mRNA levels in the plasma from patients
with MM. All data was presented as the mean ± standard deviation
from at least three repeats of each independent experiment.
P<0.05 was considered to indicate a statistical significant
difference.
Results
miR-765 is upregulated in plasma from
patients with MM and MM cell lines
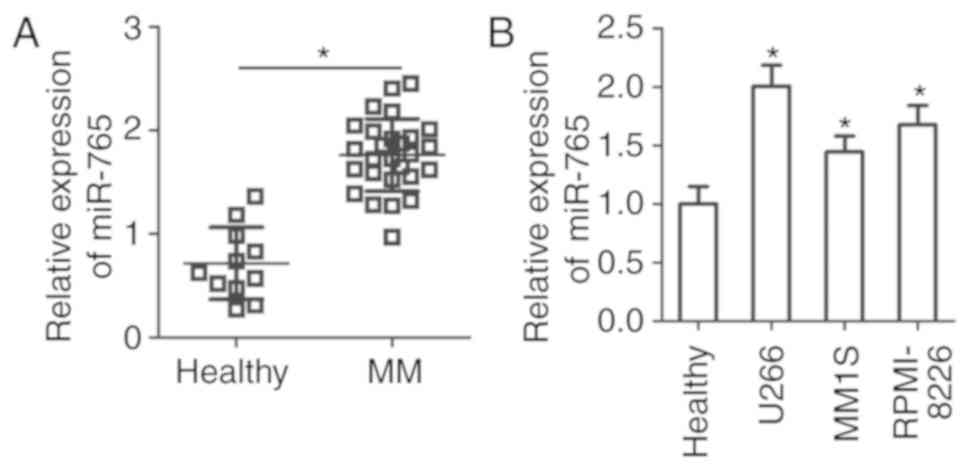
To assess the expression status of miR-765 in MM,
the expression of miR-765 was determined in the plasma cells of 27
patients with MM and 11 healthy individuals. The results of RT-qPCR
revealed that miR-765 was highly expressed in plasma samples from
patients with MM compared with healthy individuals (P<0.05;
Fig. 1A). miR-765 expression in
three human MM cell lines was then examined, including U266, MM1S
and RPMI-8226. Consistently, the expression of miR-765 was higher
in all three MM cell lines compared with that in the plasma samples
from healthy individuals (P<0.05; Fig. 1B). These results suggest that miR-765
is upregulated in MM, and this upregulation may serve an important
role in MM progression.
Inhibition of miR-765 attenuates cell
proliferation and promotes apoptosis in MM
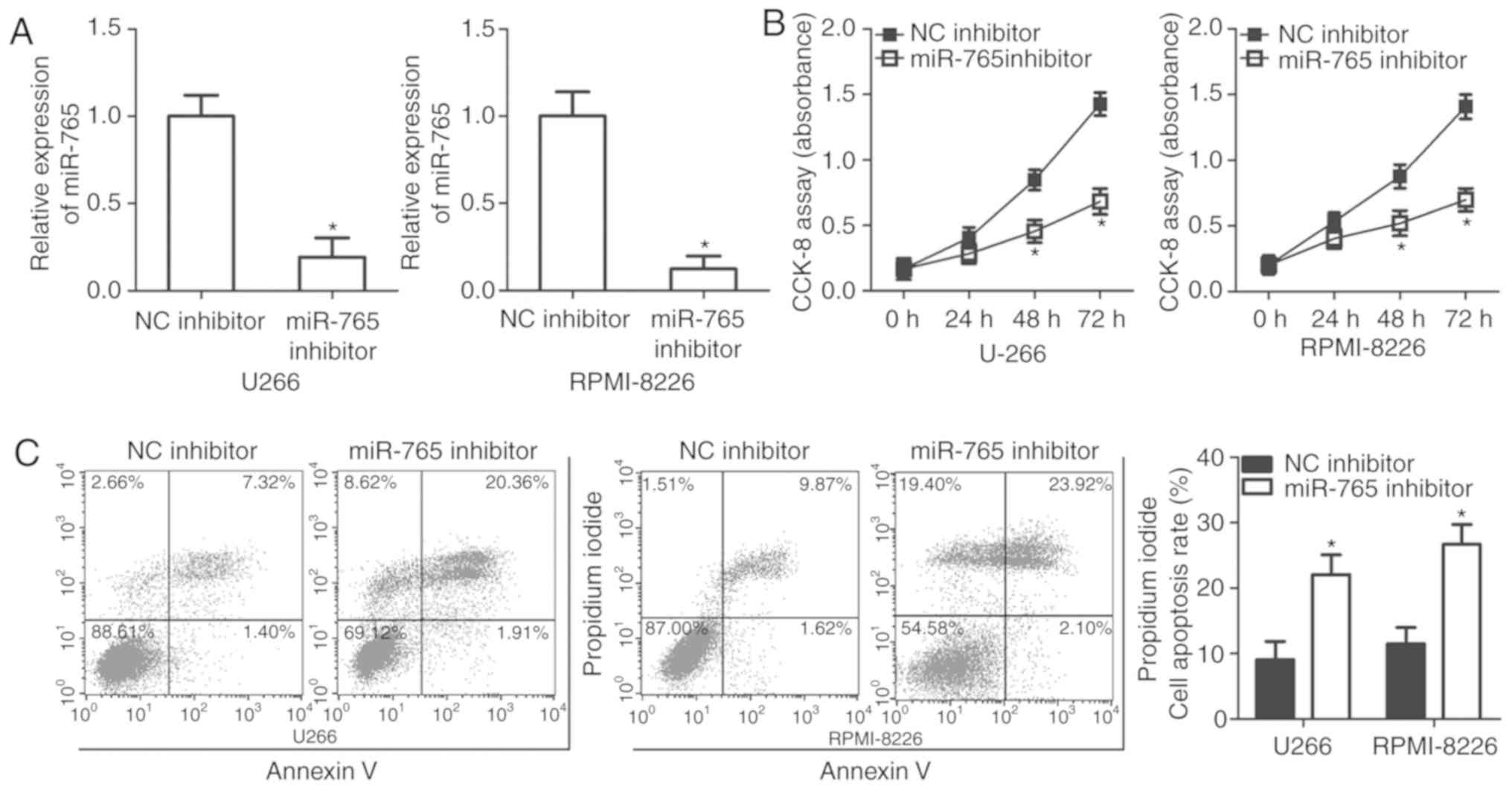
U266 and RPMI-8226 cell lines possessed a relatively
high miR-765 expression among the three MM cell lines and thus,
were selected for further functional experiments. To assess the
role of miR-765 in MM cells, miR-765 was significantly knocked down
in U266 and RPMI-8226 cells by transfection with a miR-765
inhibitor (P<0.05; Fig. 2A). A
CCK-8 assay was then performed at different time points to
determine the effect of miR-765 in MM cell proliferation. It was
observed that the downregulation of miR-765 significantly impaired
the proliferation of U266 and RPMI-8226 cells after 48 and 72 h
(P<0.05; Fig. 2B). Since cell
proliferation is closely associated with cell apoptosis, flow
cytometry analysis was performed to detect the proportion of
apoptotic U266 and RPMI-8226 cells after transfection with a
miR-765 inhibitor or an NC inhibitor. Apoptosis rate was determined
to be significantly increased in U266 and RPMI-8226 cells
transfected with the miR-765 inhibitor relative to that in cells
transfected with the NC inhibitor (P<0.05; Fig. 2C). The results indicate that miR-765
may serve an oncogenic role in the development of MM.
SOX6 is a direct target gene of
miR-765 in MM cells
It is generally accepted that miRNAs contribute to
cancer initiation and progression by regulating their target genes
(6). To determine the mechanisms
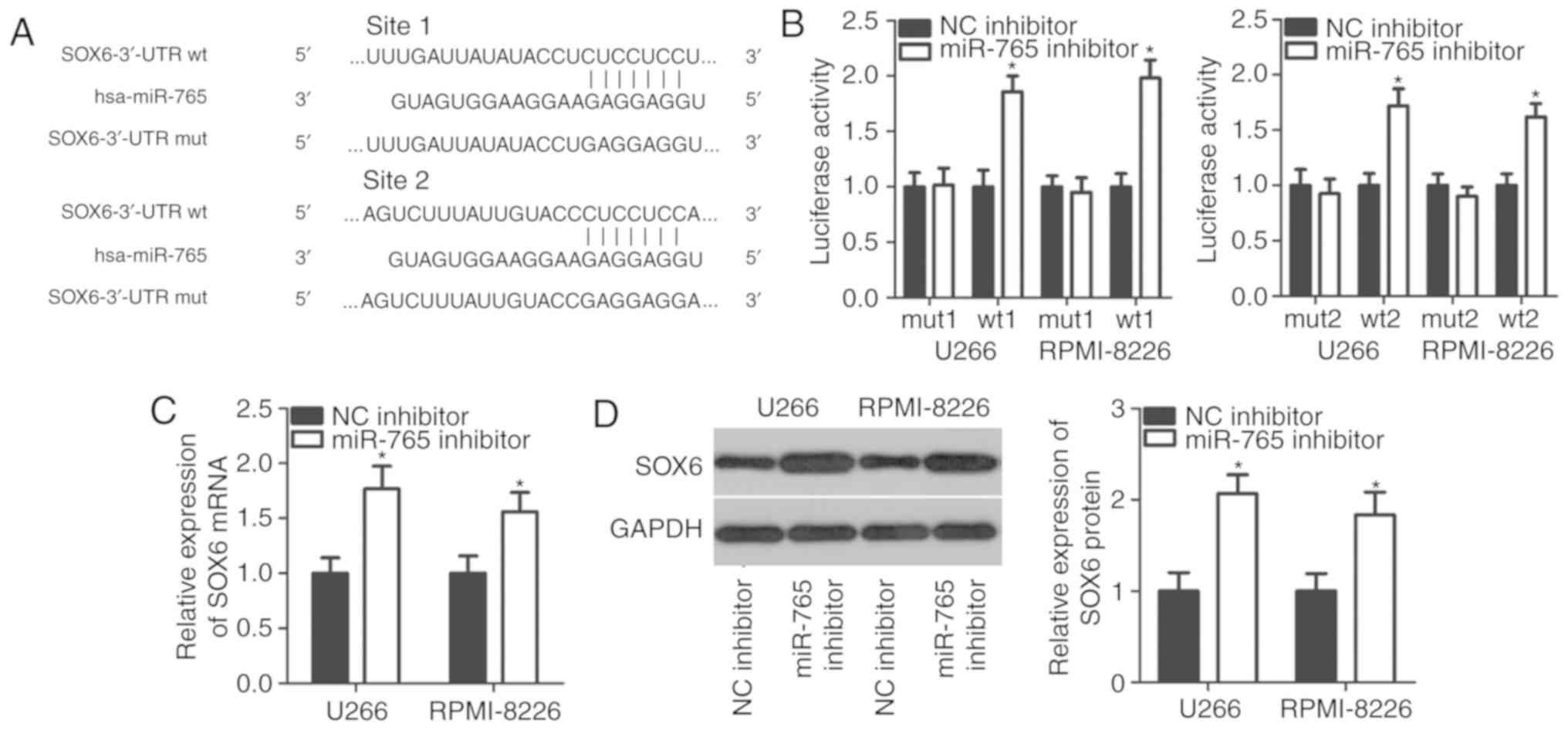
underlying miR-765 activity in MM cells, bioinformatics analysis
was performed to search for potential targets of miR-765. Two
putative miR-765-binding sites were located in the 3′-UTR of SOX6
(Fig. 3A). SOX6 was selected for
further experimental validation as SOX6 is involved in the
tumorigenesis and tumor development of multiple types of human
cancer (28–30). A luciferase reporter assay was
performed to confirm whether miR-765 directly targets the 3′-UTR of
SOX6. The results revealed that co-transfection with the
pMIR-SOX6-3′-UTR wt (1 and 2) and miR-765 inhibitor in U266 and
RPMI-8226 cells led to a significant increase in luciferase
activity (P<0.05; Fig. 3B).
However, the luciferase activity in cells transfected with the
pMIR-SOX6-3′-UTR mut (1 and 2) and miR-765 inhibitor was not of
significance. Furthermore, the mRNA and protein levels of SOX6 were
evaluated in U266 and RPMI-8226 cells upon miR-765 downregulation.
The results revealed that the downregulation of miR-765
significantly increased SOX6 expression in U266 and RPMI-8226 cells
at mRNA (P<0.05; Fig. 3C) and
protein levels (P<0.05; Fig. 3D).
These results demonstrate that SOX6 is a direct target gene of
miR-765 in MM cells.
Downregulation of SOX6 is negatively
correlated with miR-765 expression in the plasma from patients with
MM
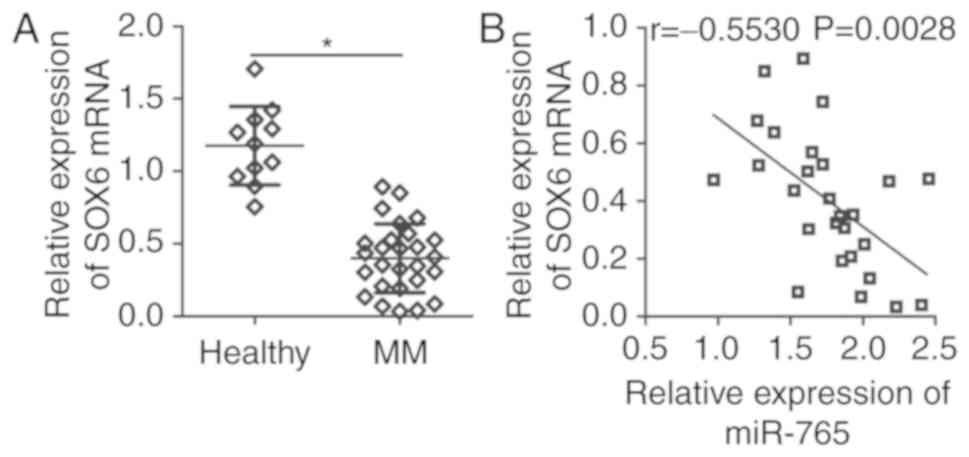
To further determine the association between miR-765
and SOX6 in MM, SOX6 expression was assessed in the plasma of 27
patients with MM patients and 11 healthy individuals. RT-qPCR
analysis revealed that SOX6 mRNA expression was significantly lower
in the plasma of patients with MM (P<0.05; (Fig. 4A). In addition, a significant
negative correlation was identified between miR-765 and SOX6 mRNA
expression in the plasma of patients with MM (r=−0.5530; P=0.0028;
Fig. 4B), as determined via
Spearmans correlation analysis. These results suggest that the
downregulation of SOX6 in MM may be, at least in part, caused by
miR-765 upregulation.
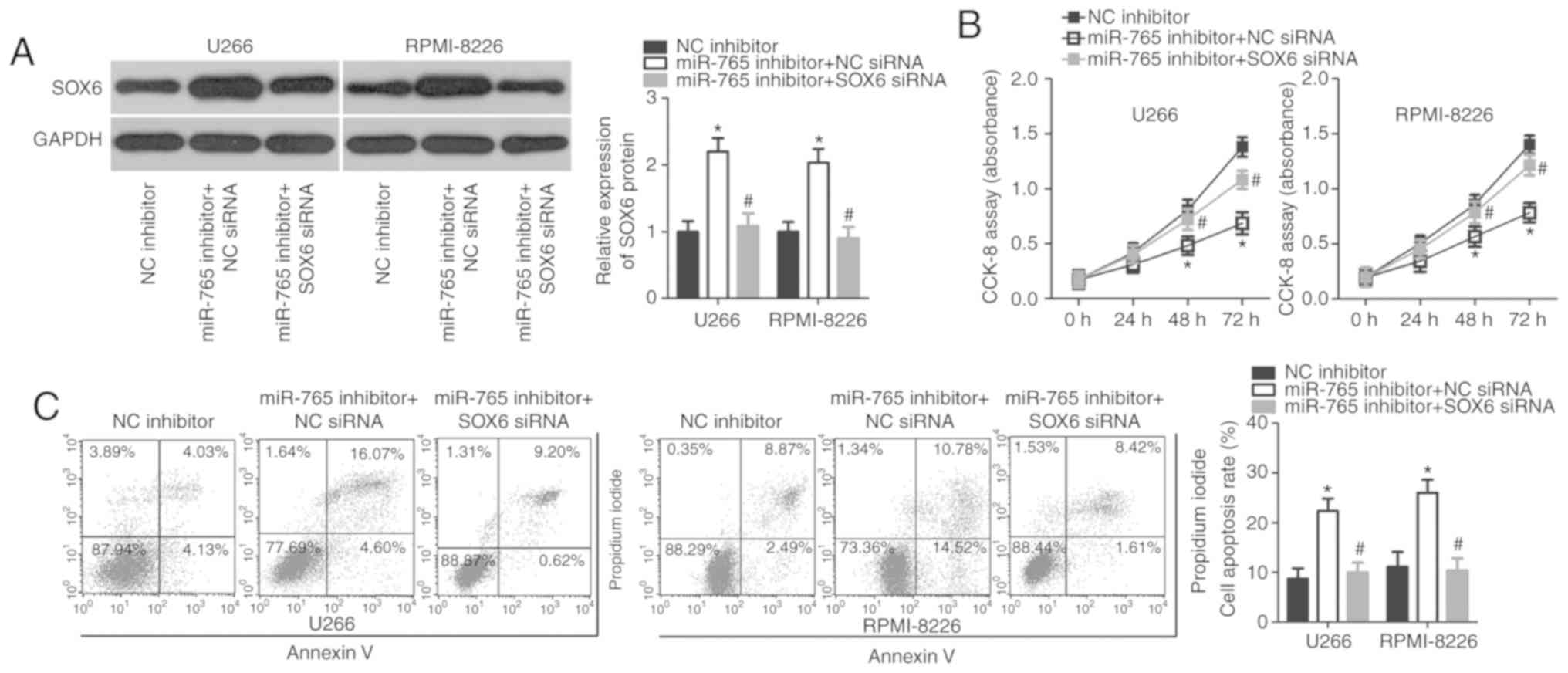
SOX6 is required for miR-765-induced
phenotypes in MM cells
As SOX6 was validated as a direct target of miR-765,
rescue experiments were subsequently performed to investigate
whether SOX6 was essential for miR-765 inhibitor-induced
proliferation suppression in MM cells. A miR-766 inhibitor was
co-transfected with SOX6 siRNA or NC siRNA into U266 and RPMI-8226
cells. Western blot analysis revealed that the upregulation of SOX6
caused by miR-765 downregulation was recovered in U266 and
RPMI-8226 cells after co-transfection with SOX6 siRNA (P<0.05;
Fig. 5A). Functional assays also
revealed that recovered SOX6 expression reversed the effects of
miR-765 downregulation in the proliferation (P<0.05; Fig. 5B) and apoptosis (P<0.05; Fig. 5C) of U266 and RPMI-8226 cells. These
data confirm that miR-765 may serve as an oncogene in MM cells, at
least partly, through the regulation of SOX6 expression.
Discussion
Previous studies have demonstrated that miRNAs are
closely associated with the occurrence and development of MM via
the regulation of cell proliferation, cycle, apoptosis,
angiogenesis and metastasis (31–33).
miRNAs have been proposed to be novel diagnostic biomarkers and
effective therapeutic targets for anticancer therapy (34). Accordingly, an in-depth understanding
of the biological functions of miRNAs in MM may be helpful to
identify promising therapeutic techniques for patients with MM. In
the present study, a series of experiments were performed to
determine the expression status of miR-765 in MM and to assess its
role in MM development. The molecular mechanisms underlying the
action of miR-765 in MM were also explored. The results revealed
that miR-765 may serve as a diagnostic biomarker and antitumor
therapeutic agent for MM.
miR-765 is aberrantly upregulated in esophageal
squamous cell carcinoma tissues and cell lines (24). The upregulation of miR-765 is
strongly associated with tumor stage, lymph node metastasis and
clinical stage of patients with esophageal squamous cell carcinoma
(24). Patients with esophageal
squamous cell carcinoma and high miR-765 levels exhibit poorer
overall survival and disease-free survival rates than patients with
low miR-765 levels (24).
Multivariate analysis has previously identified miR-765 as an
independent biomarker for the prediction of overall survival and
disease-free survival for patients with esophageal squamous cell
carcinoma (24). miR-765 expression
is also increased in osteosarcoma (25) and hepatocellular carcinoma (26). Expression levels of miR-765 are
significantly correlated with the better prognosis of osteosarcoma
patients (25). In contrast, miR-765
is downregulated in tongue squamous cell carcinoma (35). However, the expression pattern of
miR-765 in MM remains unclear. In the current study, RT-qPCR was
performed to measure the expression of miR-765 in the plasma of
patients with MM and cell lines. The results revealed that miR-765
was highly expressed in the plasma from patients with MM and MM
cell lines. These inconsistent results suggest a cancer-specific
expression pattern of miR-765 in human malignant tumors.
miR-765 is an oncogene in hepatocellular carcinoma
(26). The upregulation of miR-765
induces the proliferation and tumorigenesis of hepatocellular
carcinoma cells by regulating polyphosphate 4-phosphatase type II
(26). Inversely, miR-765 serves a
role in tumor suppression in tongue squamous cell carcinoma.
miR-765 directly targets laminin subunit gamma 2 to inhibit tongue
squamous cell carcinoma cell proliferation and invasion to increase
cell cycle arrest (35). However,
the exact roles of miR-765 in MM remain poorly understood. To the
best of our knowledge, the present study revealed that the
downregulation of miR-765 restricts cell proliferation and promotes
cell apoptosis in MM. These results suggest that miR-765 may serve
as a novel therapeutic target for patients with these specific
types of cancer.
Identifying the direct target genes of miR-765 in MM
may facilitate the development of promising therapeutic targets.
SOX6, a member of the Sox transcription-factor family (36), is demonstrated to be a direct target
gene of miR-765 in MM cells. SOX6 is aberrantly upregulated in
numerous types of cancer, including pancreatic cancer (37), prostate cancer (38), ovarian cancer (29), colorectal cancer (39) and hepatocellular carcinoma (40). SOX6 may contribute to the occurrence
and development of tumorigenesis and tumor development through the
regulation of various biological features, including cell
proliferation, cycle, apoptosis, epithelial-mesenchymal transition
and metastasis (28–30). The current study demonstrated that
miR-765 directly targets SOX6 to implicate the progression of MM.
The restoration of SOX6 using miR-765 based targeted therapy may
therefore be considered a novel and promising therapeutic
opportunity for the treatment of patients with this aggressive
cancer.
In conclusion, the present study revealed that
miR-765 is upregulated in the plasma of patients with MM and MM
cell lines. In addition, miR-765 may serve oncogenic roles in MM by
regulating cell proliferation and apoptosis in vitro.
Furthermore SOX6 was identified as a direct target gene of miR-765
in MM cells. The results indicate that the inhibition of miR-765
may have great potential in inhibiting the rapid proliferation and
accumulation of MM cells, confirming that miR-765 is a promising
target for MM prevention and treatment. However, the effects of
SOX6 in MM cells were not examined in the current study. This
limitation however, should be resolved in future research.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
GC and Shifeng L designed the present study and
wrote the manuscript. RT-qPCR, western blot analysis and the CCK-8
assay were performed by Shifeng L, and Shengping L. HH and GC
performed flow cytometry analysis, the luciferase reporter assay
and statistical analysis. All authors have read and approved the
final draft of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Nanchang University
(Nanchang, China), and was performed in accordance with the
Declaration of Helsinki and the guidelines of the Ethics Committee
of The First Affiliated Hospital of Nanchang University. Written
informed consent was obtained from all patients for the use of
their clinical tissues.
Patient consent for publication
Patient consent for publication was obtained.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Anderson KC and Carrasco RD: Pathogenesis
of myeloma. Annu Rev Pathol. 6:249–274. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pruneri G, Cinieri S, Peccatori F and
Viale G: Unusual cases in multiple myeloma and a dramatic response
in metastatic lung cancer: Case 2. Plasma cell myeloma coexisting
with metastatic breast carcinoma in the bone marrow. J Clin Oncol.
23:232–233. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Naymagon L and Abdul-Hay M: Novel agents
in the treatment of multiple myeloma: A review about the future. J
Hematol Oncol. 9:522016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kumar SK, Lee JH, Lahuerta JJ, Morgan G,
Richardson PG, Crowley J, Haessler J, Feather J, Hoering A, Moreau
P, et al: Risk of progression and survival in multiple myeloma
relapsing after therapy with IMiDs and bortezomib: A multicenter
international myeloma working group study. Leukemia. 26:149–157.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang S, Hu C, Liu P and Lu M:
Tumor-derived exosomes in cancer metastasis risk diagnosis and
metastasis therapy. Clin Transl Oncol. 21:152–159. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan HL, Wang T and Zhang KH: MicroRNAs as
potential biomarkers for diagnosis, therapy and prognosis of
gastric cancer. Onco Targets Ther. 11:3891–3900. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sharma N and Baruah MM: The microRNA
signatures: Aberrantly expressed miRNAs in prostate cancer. Clin
Transl Oncol. 21:126–144. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu B, Ju S, Chu H, Shen X, Zhang Y, Luo X
and Cong H: The potential function of microRNAs as biomarkers and
therapeutic targets in multiple myeloma. Oncol Lett. 15:6094–6106.
2018.PubMed/NCBI
|
|
11
|
Nobili L, Ronchetti D, Agnelli L, Taiana
E, Vinci C and Neri A: Long Non-Coding RNAs in multiple myeloma.
Genes (Basel). 9(pii): E692018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rastgoo N, Abdi J, Hou J and Chang H: Role
of epigenetics-microRNA axis in drug resistance of multiple
myeloma. J Hematol Oncol. 10:1212017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang YZ, Zhang XY, Wan Q and Li J: Role of
Exosomal miRNA in multiple myeloma progression and its possible
mechanism-review. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 25:301–305.
2017.(In Chinese). PubMed/NCBI
|
|
14
|
Rossi M, Tagliaferri P and Tassone P:
MicroRNAs in multiple myeloma and related bone disease. Ann Transl
Med. 3:3342015.PubMed/NCBI
|
|
15
|
Bi C and Chng WJ: MicroRNA: Important
player in the pathobiology of multiple myeloma. Biomed Res Int.
2014:5215862014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cao Y, Shi X, Liu Y, Xu R and Ai Q:
MicroRNA-338-3p inhibits proliferation and promotes apoptosis of
multiple myeloma cells through targeting Cyclin-dependent kinase 4.
Oncol Res. 27:117–124. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tang B, Xu A, Xu J, Huang H, Chen L, Su Y,
Zhang L, Li J, Fan F, Deng J, et al: MicroRNA-324-5p regulates
stemness, pathogenesis and sensitivity to bortezomib in multiple
myeloma cells by targeting hedgehog signaling. Int J Cancer.
142:109–120. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Alzrigat M and Jernberg-Wiklund H: The
miR-125a and miR-320c are potential tumor suppressor microRNAs
epigenetically silenced by the polycomb repressive complex 2 in
multiple myeloma. RNA Dis. 4(pii): e15292017.PubMed/NCBI
|
|
19
|
Zhang X, Chen Y, Zhao P, Zang L, Zhang Z
and Wang X: MicroRNA-19a functions as an oncogene by regulating
PTEN/AKT/pAKT pathway in myeloma. Leuk Lymphoma. 58:932–940. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hua J, Ding T and Yang L: Dysfunction of
microRNA-32 regulates ubiquitin ligase FBXW7 in multiple myeloma
disease. Onco Targets Ther. 9:6573–6579. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ikeda S, Kitadate A, Abe F, Saitoh H,
Michishita Y, Hatano Y, Kawabata Y, Kitabayashi A, Teshima K, Kume
M, et al: Hypoxia-inducible microRNA-210 regulates the DIMT1-IRF4
oncogenic axis in multiple myeloma. Cancer Sci. 108:641–652. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Z, Zhang G, Yu W, Gao N and Peng J:
miR-186 inhibits cell proliferation in multiple myeloma by
repressing Jagged1. Biochem Biophys Res Commun. 469:692–697. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saha MN, Abdi J, Yang Y and Chang H:
MiRNA-29a as a tumor suppressor mediates PRIMA-1Met-induced
anti-myeloma activity by targeting c-Myc. Oncotarget. 7:7149–7160.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jiang B, Xu G, Lv HQ, Huang M and Li Z:
Up-regulation of miR-765 predicts a poor prognosis in patients with
esophageal squamous cell carcinoma. Eur Rev Med Pharmacol Sci.
22:3789–3794. 2018.PubMed/NCBI
|
|
25
|
Liang W, Wei X, Li Q, Dai N, Li CY, Deng
Y, Jiang X, Tan XR, Dai XY, Li MX, et al: MicroRNA-765 enhances the
anti-angiogenic effect of CDDP via APE1 in Osteosarcoma. J Cancer.
8:1542–1551. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xie BH, He X, Hua RX, Zhang B, Tan GS,
Xiong SQ, Liu LS, Chen W, Yang JY, Wang XN and Li HP: Mir-765
promotes cell proliferation by downregulating INPP4B expression in
human hepatocellular carcinoma. Cancer Biomark. 16:405–413. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Z, Li J, Li K and Xu J: SOX6 is
downregulated in osteosarcoma and suppresses the migration,
invasion and epithelial-mesenchymal transition via TWIST1
regulation. Mol Med Rep. 17:6803–6811. 2018.PubMed/NCBI
|
|
29
|
Li Y, Xiao M and Guo F: The role of Sox6
and Netrin-1 in ovarian cancer cell growth, invasiveness, and
angiogenesis. Tumour Biol. 39:10104283177055082017.PubMed/NCBI
|
|
30
|
Zhu Y, Xia Y, Niu H and Chen Y: MiR-16
induced the suppression of cell apoptosis while promote
proliferation in esophageal squamous cell carcinoma. Cell Physiol
Biochem. 33:1340–1348. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang Y, Li F, Saha MN, Abdi J, Qiu L and
Chang H: miR-137 and miR-197 induce apoptosis and suppress
tumorigenicity by targeting MCL-1 in multiple myeloma. Clin Cancer
Res. 21:2399–2411. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qu X, Zhao M, Wu S, Yu W, Xu J, Xu J, Li J
and Chen L: Circulating microRNA 483-5p as a novel biomarker for
diagnosis survival prediction in multiple myeloma. Med Oncol.
31:2192014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma J, Liu S and Wang Y: MicroRNA-21 and
multiple myeloma: Small molecule and big function. Med Oncol.
31:942014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu Y, Wang H, Chen E, Xu Z, Chen B and Lu
G: Candidate microRNAs as biomarkers of thyroid carcinoma: A
systematic review, meta-analysis, and experimental validation.
Cancer Med. 5:2602–2614. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ding J, Yang C and Yang S: LINC00511
interacts with miR-765 and modulates tongue squamous cell carcinoma
progression by targeting LAMC2. J Oral Pathol Med. 47:468–476.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ohta S, Misawa A, Lefebvre V, Okano H,
Kawakami Y and Toda M: Sox6 up-regulation by macrophage migration
inhibitory factor promotes survival and maintenance of mouse neural
stem/progenitor cells. PLoS One. 8:e743152013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang W, Yuan Q, Jiang Y, Huang L, Chen C,
Hu G, Wan R, Wang X and Yang L: Identification of Sox6 as a
regulator of pancreatic cancer development. J Cell Mol Med.
22:1864–1872. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yu Y, Wang Z, Sun D, Zhou X, Wei X, Hou W,
Ding Y, Ma Y and Hou Y: miR-671 promotes prostate cancer cell
proliferation by targeting tumor suppressor SOX6. Eur J Pharmacol.
823:65–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li YC, Li CF, Chen LB, Li DD, Yang L, Jin
JP and Zhang B: MicroRNA-766 targeting regulation of SOX6
expression promoted cell proliferation of human colorectal cancer.
Onco Targets Ther. 8:2981–2988. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Guo X, Yang M, Gu H, Zhao J and Zou L:
Decreased expression of SOX6 confers a poor prognosis in
hepatocellular carcinoma. Cancer Epidemiol. 37:732–736. 2013.
View Article : Google Scholar : PubMed/NCBI
|