Introduction
Paroxysmal nocturnal hemoglobinuria (PNH) is a
non-malignant clonogenic disease originating from hematopoietic
stem cells that is caused by somatic gene mutations (1,2). The
pathological defect of PNH is the abnormal synthesis of glycosyl
phosphatidyl inositol (GPI) caused by phosphatidylinositol glycan
anchor biosynthesis class A (PIG-A) gene mutations on the X
chromosome, which leads to the reduction or deletion of
GPI-anchored proteins that are connected to the membrane of blood
cells [including cluster of differentiation (CD)55, CD59 and CD56]
and eventually causes activation of the complement system, leading
to symptoms of intravascular hemolysis (3,4). The
main clinical manifestations of PNH are chronic intravascular
hemolysis, bone marrow failure, high-risk complications of
thrombosis, renal failure and pulmonary hypertension (5–7).
Our previous study (8) sequenced the whole genome exon of
CD59− cells derived from 13 patients with PNH and
identified additional mutant genes such as cut-like homeobox 1
(CUX1), mixed-lineage leukemia 2 (MLL2), SUZ12, RB transcriptional
corepressor 1 (RB1), mucin 4 (MUC4) and tet methylcytosine
dioxygenase 2 (TET2), although not PIG-A. The functions of these
genes are associated with the regulation of cell proliferation,
differentiation, resistance to apoptosis, invasion and progression
of tumor cells (9–11). This suggests that, although PNH is a
benign clonal proliferative disease, its clonal composition has
similar biological characteristics to leukemia and other tumors.
The proliferation advantage of PNH clones may require the
participation of gene defects other than PIG-A gene mutations,
suggesting that there may be additional gene mutations in patients
with PNH (12–15). Therefore, the present study performed
500-depth whole exome sequencing (WES) for 13 patients with PNH.
The mutation rate of the PIG-A gene was 30.77% (4/13) in 100-depth
sequencing, whereas recombinant signal binding protein of
immunoglobulin κJ region (RBPJ) gene mutation was detected in 6
patients (46.15%). However, in 500-depth sequencing, the mutation
rate of PIG-A was higher than in 100-depth sequencing (61.54%,
8/13), which was consistent with literature reports, and 10
patients (76.92%) exhibited RBPJ gene mutations. In-depth analysis
and screening of the WES results were conducted. The present study
screened out the RBPJ gene with high expression and first mutation
level. It has been reported that RBPJ serves a key role in
promoting the occurrence and development of various malignant
tumors. Thus, inhibiting the expression of RBPJ may impair or
promote the growth of potential tumors (16,17).
However, to the best of our knowledge, there have been no previous
studies on the application of RNA interference to knock down the
expression of RBPJ in patients with PNH.
The current study evaluated the regulation of PNH
primary cells by RBPJ depletion. Small interfering RNA (siRNA) was
used to efficiently knock down the RBPJ gene in PNH clones of 5
patients in vitro, and cell proliferation, apoptosis rate
and cell cycle changes were observed to validate its functions.
Materials and methods
Patients
The clinical characteristics of the initial 13
patients with PNH, who were recruited from January-December 2015
and underwent WES, are presented in Table I. The results of WES were analyzed,
13 target genes were screened, and an additional 30 patients with
PNH were recruited from January 2016-December 2017 for PCR
experiments. The clinical characteristics of the additional 30
patients are presented in Table II.
An Automatic hematology Instrument (LH 750; Beckman Coulter, Inc.)
and an Automatic biochemical Instrument (C8000; Abbott
Pharmaceutical Co., Ltd.) were utilized to detect blood cell counts
[white blood cell (WBC), hemoglobinuria (Hb), platelet and
reticulocyte (Ret)] and hemolysis indices [lactate dehydrogenase
(LDH), total bilirubin and direct bilirubin), respectively. All
patients were diagnosed with PNH and were admitted to the
Department of Hematology of Tianjin Medical University General
Hospital (Tianjin, China). The diagnostic criteria refer to the
international PNH Research Group criteria (18). The control group consisted of 30
healthy donors, which were recruited from the Physical Examination
Center of General Hospital of Tianjin Medical University from
January 2017 to December 2017. Healthy controls (19 males and 11
females) were matched by sex and age, with an average age of 42
years (age range, 19–68 years). The Ethics Committee of Tianjin
Medical University approved the present study (approval no.
IRB2018-YX-042), and all the enrolled cases provided written
informed consent.
 | Table I.Initial clinical characteristics of
13 PNH patients. |
Table I.
Initial clinical characteristics of
13 PNH patients.
| Clinical
feature | Measurement |
|---|
| Clinical
classification of PNH, n (%) |
|
|
Classical PNH | 7 (53.85) |
|
PNH-AA | 6 (46.15) |
|
Subclinical-PNH | 0 |
| Sex, n (%) |
|
|
Male | 6 (46.15) |
|
Female | 7 (53.85) |
| Age
(years), median age (range) | 31 (21–73) |
| Blood
examination |
|
| RET
% | 7.98±4.57 |
| RBC
(*1012/l) | 2.44±0.63 |
| WBC
(*109/l) | 3.94±2.14 |
| HGB
(g/l) | 82.08±20.24 |
| PLT
(*109/l) | 73.46±66.66 |
| D-Dimer
(ug/ml) |
1,672.00±1,404.00 |
| LDH
(U/l) |
1,058.00±663.30 |
| TBIL
(µmol/l) | 24.48±14.69 |
| DBIL
(µmol/l) | 7.18±3.59 |
|
Granulocyte CD59−
(%) | 78.58±17.02 |
|
Erythrocyte CD59−
(%) | 52.53±30.28 |
|
Flaer−/CD14−
(%) | 81.52±12.63 |
|
Flaer−/CD24−
(%) | 85.52±13.29 |
| Table II.Initial clinical characteristics of
30 PNH patients. |
Table II.
Initial clinical characteristics of
30 PNH patients.
| Characteristic | Patients | Controls | P-value |
|---|
| Total, n | 30 | 30 |
|
| Gender,
male/female | 19/11 | 19/11 |
|
| Age (years), median
(range) | 38 (18–75) | 42 (19–68) |
|
| Clinical
classification, n (%) |
| 0 |
|
|
Classical PNH | 23 (76.67) |
|
|
|
PNH-AA | 6
(20.00) |
|
|
|
Subclinical-PNH | 1 (3.33) |
|
|
| History
of thrombosis, n (%) | 4
(13.33) | 0 |
|
| Parameters at
baseline |
|
|
|
| HGB
(g/l) |
78.71±20.94 | 127.30±9.64 | <0.001 |
| Ret
(%) | 9.85±5.24 |
1.34±0.33 | 0.0017 |
| WBC
(×109/l) |
5.89±3.55 |
6.11±1.51 | 0.778 |
| PLT
(×109/l) |
100.40±65.77 |
171.80±56.06 | 0.003 |
| TBIL
(µmol/l) |
31.79±15.07 |
10.75±6.11 | 0.016 |
| DBIL
(µmol/l) |
8.34±3.54 |
3.77±2.17 | 0.045 |
| LDH
(U/l) |
1,578.00±931.70 |
191.50±228.70 | <0.001 |
| Cr
(µmol/l) |
60.19±27.47 |
53.35±21.67 | 0.531 |
|
Granulocyte CD59−
(%) |
80.44±17.30 |
1.84±1.05 | <0.001 |
|
Erythrocyte CD59−
(%) |
52.38±30.25 |
2.69±1.27 | 0.006 |
|
Flaer−/CD14−
(%) |
77.30±21.28 | Undetected |
|
|
Flaer−/CD24−
(%) |
82.65±17.62 | Undetected |
|
Exome sequencing, reference genome and
target area information
A total of 13 blood samples and paired normal
fingernail samples were subjected to WES. In order to enrich the
coding regions as much as possible, Sure Select Human All Exon 50M
(Agilent Technologies, Inc.) was used for blood samples and Sure
Select Human All Exon V4 (Agilent Technologies, Inc.) for nail
samples. High-throughput sequencing of blood and nail samples was
performed using an Illumina HiSeq2500 instrument (Illumina, Inc.).
The average sequencing depths for blood and nail samples were 100×
(range, 47–107×) and 102× (range, 96–106×), respectively. The
sequencing depth of the above genes is presented in Fig. 1A. Broadband Wireless Access (BWA) was
used to perform the alignment with the default parameters and human
genome 19/the Genome Reference Consortium Human Genome Build 37
(hg19/GRCh37; ftp://ftp.ncbi.nlm.nih.gov/genomes/H_sapiens) served
as the reference genome. The relevant details were as follows:
Genome version number, hg19/GRCh37; genome size, 3095677412 bp;
capture chip, SeqCap EZ Human Exome Library NimbleGen v2.0; exon
sequencing genes >20,000 genes; and capture target area size,
44.1 Mb. The associated 1000G database was utilized to annotate and
report mutations in MAF (https://www.ncbi.nlm.nih.gov/variation/tools/1000genomes/).
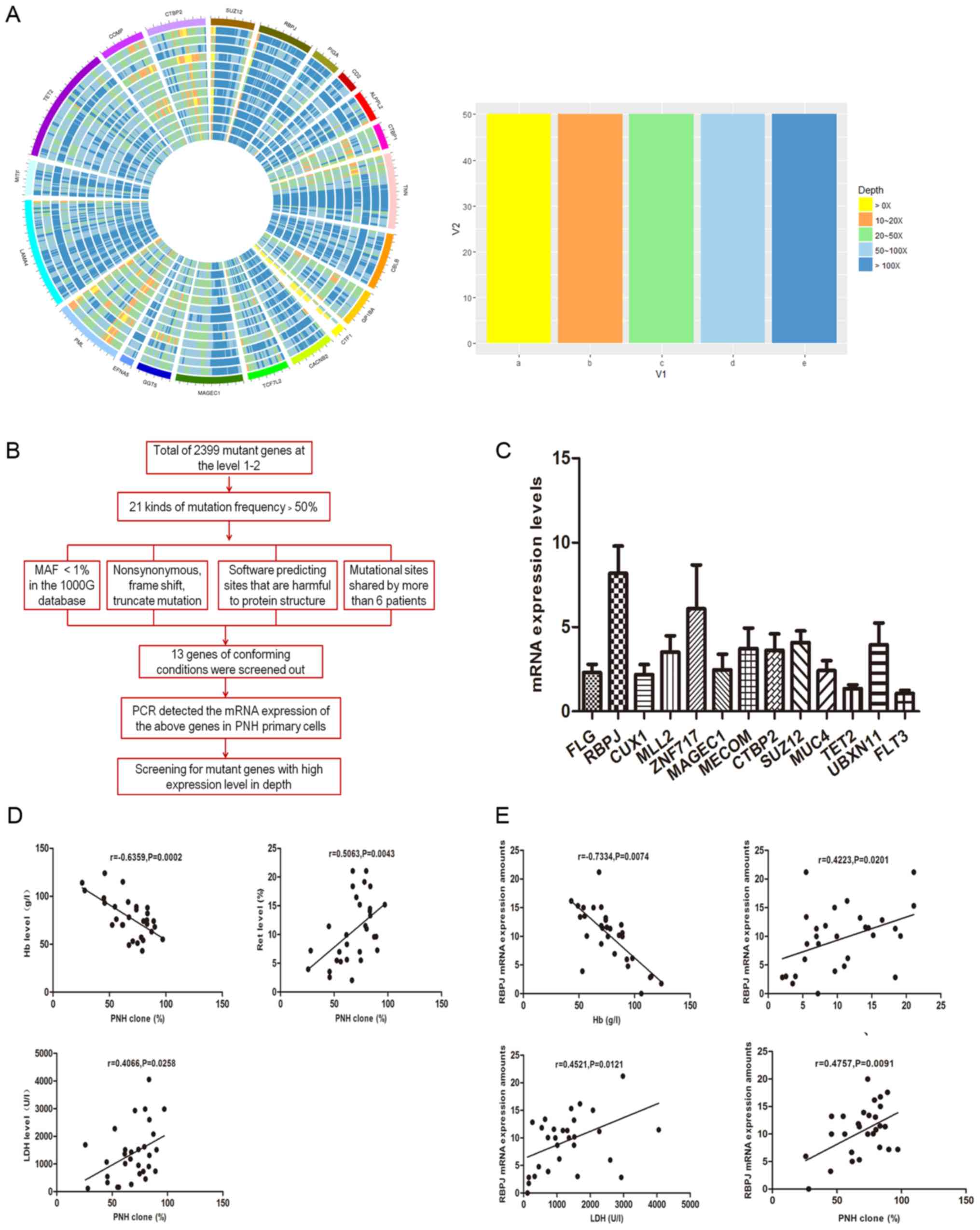 | Figure 1.(A) The outermost color represents
different genes, different colors represent different sequencing
depth, each gene width represents the length of the corresponding
gene coding region. (B) Flow chart of screening target genes. (C)
Comparison of relative expression levels of 13 target mutant genes
in 30 PNH patients which were screened by whole exome sequencing.
(D) Correlation analysis between PNH cloning proportion and
hemolysis index. (E) Correlation analysis between RBPJ mRNA
expression and clinical hemolysis index. The expression of RBPJ
mRNA was negatively correlated with the proportion of the level of
Hb, and positively correlated with the proportion of RET, LDH
levels and the proportion of PNH clones. PNH, paroxysmal nocturnal
hemoglobinuria; RPBJ, recombinant signal binding protein of
immunoglobulin κJ region; Hb, hemoglobinuria; RET, reticulocyte;
LDH, lactate dehydrogenase; FLG, filaggrin; CUX1, cut-like homeobox
1; MLL2, mixed-lineage leukemia 2; ZNF717, zinc finger protein 717;
MAGEC1, MAGE family member C1; MECOM, MDS1 and EVI1 complex locus;
CTBP2, C-terminal binding protein 2; MUC4, mucin 4; TET2, tet
methylcytosine dioxygenase 2; UBXN11, UBX domain protein 11; FLT3,
fms related tyrosine kinase 3; MAF, minor allele frequency; PCR,
polymerase chain reaction. |
Flow cytometry and cell sorting
To obtain CD59− and CD59+
neutrophils for RNA extraction, cell sorting was performed in
peripheral blood of patients with PNH and healthy controls on the
same day of blood extraction. Red blood cells were lyzed with 10 ml
erythrocytolysin solution (BD Biosciences) and then centrifuged at
150 × g for 5 min at room temperature. Cells were then washed twice
with PBS and resuspended in 300 µl PBS. Immunomagnetic cell
selection was performed using phycoerythrin (PE)
conjugated-anti-CD59 Phycoerythrin (1:5; cat. no. 555764; BD
Biosciences), followed by anti-PE microbeads (Miltenyi Biotec
GmbH). Samples were separated using MS Columns (Miltenyi Biotec
GmbH), and purity was verified in each fraction by flow cytometry
using an Aria II instrument (BD Biosciences). All results were
analyzed using CellQuest™ Pro Software 4.0.2 (BD Biosciences).
To ensure the viability of PNH cells in culture and
the transfection efficiency, CD59− of neutrophils from
15 patients with PNH were sorted by flow cytometry. Upon cell
sorting, the number of cells reached 2×107, and the
purity of the cells was 96–97%.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from CD59− and
CD59+ cells of patients with PNH and normal controls,
and from mononuclear cells, respectively, with RNeasy kit (Takara
Bio, Inc.). A Reverse transcription kit (Takara Bio, Inc.) was used
to synthesize cDNA from 1 µg total RNA and was purified using the
QIAquick PCR Purification Kit (Qiagen, Inc.). QuantiTect SYBR Green
PCR Kit (Tli RNaseH Plus; Takara Bio, Inc.) and Light Cycler 1.5
Real-Time PCR System (Roche Diagnostics, Indianapolis, IN, USA)
were used to perform RT-qPCR in duplicates. Specific primers
designed to amplify cross-exons of the RBPJ gene (158 bp) are
detailed in Table III. The
thermocycling conditions were as follows: Initial denaturation at
95°C for 30 sex; followed by 45 cycles of denaturation at 94°C for
5 sec, annealing at 60°C for 30 sec and extension at 70°C for 30
sec. A final extension was conducted at 72°C for 10 min. Bio-Rad
CFX Manager software 3.1 (Bio-Rad Laboratories, Inc.) was used to
analyze the melting and amplification curves (quantitative curve),
and the quantitative cycle (Cq) values of each group was
determined. The relative quantitative multiplier of each group
(relative fold) was expressed by 2−ΔΔCq value (19) and used for statistical analysis.
| Table III.Gene primer and siRNA sequences. |
Table III.
Gene primer and siRNA sequences.
| Name | Sequence
(5′-3′) | Length (bp) |
|---|
| RBPJ | Forward,
AGTCACTCCTGTGCCTGTGGTAG | 158 |
|
| Reverse,
CCATCTCCAACCTTCTCGGAATGC |
|
| siRNA-RBPJ1 | Forward,
GCACUCCCAAGAUU GAUAA | 21 |
|
| Reverse, UUAUCAA
UCUUGGGAGUGC |
|
| siRNA-RBPJ2 | Forward,
GCACUCCCAAGAUU GAUAA | 21 |
|
| Reverse,
UUAUCAAUCUUGGGAGUGC |
|
| siRNA-RBPJ3 | Forward,
CUGACUCAGACAAGCGAAA | 21 |
|
| Reverse,
UUUCGCUUGUCUGAGUCAG |
|
| siRNA-scr | Forward,
UUGAAGUUAUGUAUCCUCCUU | 21 |
|
| Reverse,
CUGAAGCUGCUGGGAGUAAUU |
|
| GAPDH | Forward,
GGAGCGAGATCCCTCCAAAAT | 137 |
|
| Reverse,
GGCTGTTGTCATACTTCTCATGG |
|
Cell culture
CD59− neutrophils were inoculated into
RPMI-1640 culture medium (Beijing Solarbio Science & Technology
Co., Ltd.) containing 10% (v/v) fetal bovine serum (Beijing
Solarbio Science & Technology Co., Ltd.), penicillin (100 U/ml)
and streptomycin (100 U/ml), and were suspended in a saturated
humidity chamber containing 5% CO2 at 37°C. The
centrifugal fluid exchange method (1.4 × g for 5 min at room
temperature) was used to replace the medium every 48 h and maintain
strict aseptic conditions. Logarithmic growth phase cells were used
for the experiments if their rejection rate of trypan blue (Beijing
Solarbio Science & Technology Co., Ltd.) staining was >95%.
The steps of trypan blue staining were as follows: Cell suspension
(0.1 ml) was added into 4 ml trypan blue solution (final dilution,
1:40) and placed at room temperature for 3–5 min. A drop of cell
suspension was taken and placed on a slide. The number of living
and dead cells in 1,000 cells was subsequently counted. From these
results, cell viability was calculated as follows: Cell viability
(%)=number of unstained cells/total number of observed cells ×100.
Following 72 h of cell culture, cell count reached 107
cells.
Preparation of RBPJ-siRNAs and
transfection
Specific siRNAs of the RBPJ gene (siRNA-RBPJ1,
siRNA-RBPJ2 and siRNA-RBPJ3) were purchased from Genomeditech Co.,
Ltd., whereas siRNA-scramble (scr) served as an internal reference
(Genomeditech Co., Ltd.). The siRNA sequences are provided in
Table III. All transfections with
siRNAs were performed using Lipofectamine™ 3000 (lipo3000; cat. no.
100022234; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
siRNA-RBPJ and lipo3000 were diluted with Opti-MEM (Invitrogen;
Thermo Fisher Scientific, Inc.). In order to achieve a high
transfection efficiency and gene blocking effect, siRNAs were
fluorescently labeled (Dextran, Cy7 labeled, Genomeditech Co.,
Ltd.), and concentration gradient experiments were conducted on
siRNAs (Genomeditech Co., Ltd.) and lipo3000 (cat. no. 100022234;
Thermo Fisher Scientific, Inc.). The experimental steps of the
concentration gradient were as follows: A 24-well plate
(4–8×105 cells/well) was used to perform concentration
gradient cell transfection experiments. The final concentration of
siRNA and lipofectamine™ 3000 was 30 nM (siRNA, 0.75 µl;
lipofectamine™ 3000, 1 µl), 50 nM (siRNA, 1.25 µl, lipofectamine™
3000, 1 µl) and 100 nM (siRNA, 2.5 µl, lipofectamine™ 3000, 1 µl).
The silencing efficiency of siRNA was detected by PCR 48 h
following transfection. When cells were co-transfected with
lipo3000 and RBPJ-specific siRNAs at 50 nM (lipo3000, 1 µl/well;
siRNA1, 1.25 µl/well; total volume of medium, 500 µl/well), the
siRNA was capable of efficiently blocking the expression of the
target gene (siRNA silencing efficiency was detected using RT-qPCR
48 h following transfection). The transfection efficiency of siRNA
to the cultured cells was revealed to be 75–80%. After the above
preliminary experiments, siRNA1 was used for subsequent experiments
following validation of transfection. A 24-well plate was used for
the cell transfection experiment. At 24 h prior to transfection,
2×105 cells were inoculated into 400 µl Opti-MEM to
ensure that the cell density reached 4–8×105 cells/well
at the time of transfection. The diluent of lipo3000 and siRNA was
gently mixed to guarantee a final concentration of 50 nM. The
mixture was incubated at room temperature for 20 min and then added
to the 24-well plate at 100 µl/well and mixed evenly. The plates
were incubated for 18–48 h at 37°C and 5% CO2. At 4–6 h
post-transfection, the culture medium was replaced with fresh
medium.
Cell Counting Kit-8 (CCK-8) assay for
cell proliferation
A hemocytometer was used to count the number of
cells in the prepared cell suspension, and then the cells were
inoculated in 96-well plate (100 µl/well). Following 4 h of cell
culture, CCK-8 reagent (Beijing Solarbio Science & Technology
Co., Ltd.) was added to the culture for a certain time (5–10 min at
room temperature) and then the optical density (OD) value at 450 nm
of was determined. A standard curve was created depicting the
number of cells (x-axis) vs. the OD value (y-axis). Subsequently,
the cell proliferation activity was determined by adding 10 µl
CCK-8 solution to the 96-well plates containing 100 µl
(5×104 cells/ml) cell suspension per well. Blank control
wells were prepared at the same time. Next, the cells were
incubated for 4 h under 5% CO2 and 37°C conditions. The
OD value at 450 nm was determined using a Microplate Reader (cat.
no. ELX800; BioTek Instruments, Inc.). The median value was
obtained by repeating the experiment 3 times and using the
following formula: Cell proliferation activity=OD value of the
experimental wells-OD value of the control wells; where the OD
value of the experimental wells represents the absorption of cells
and CCK-8 solution, while the OD value of the control wells
represents the absorption of the well with medium and CCK-8
solution.
Cell cycle analysis
A cell suspension (1 ml containing 1×106
cells/ml) was obtained following transfection for 72 h by washing
the cells 2 times with 1 ml PBS pre-cooled at 4°C, followed by
centrifugation (150 × g for 5 min at room temperature) of the
supernatant. PBS was added to the cell suspension and incubated
with 70% ethanol at 4°C for pre-cooling. Following 4 h of
incubation, all samples were separated by centrifugation (300 × g
for 5 min at room temperature). Next, 500 µl cell cycle dye (BD
Pharmingen; Becton, Dickinson and Company, Franklin Lakes, NJ, USA)
was added to the cell suspension and incubated for 15–20 min at
room temperature. Flow cytometry analysis (CellQuest™ Pro Software
4.0.2; BD Biosciences) was then used to detect the relative content
of DNA in the cells, and the results of DNA distribution of cell
proportion in the phase of cell cycle were analyzed by Modifit
(Verify Software House Inc, Topsham, ME, USA) with a propidium
iodide kit (PI, 20 µg/ml) containing RNase A (10 µg/ml; BD
Biosciences). The percentage of DNA in each phase of cell cycle was
analyzed to reflect the proliferation status of the cells and the
content of DNA in non-diploid (aneuploid) cells. The DNA content of
G2 and M-phase cells is typically double that exhibited by cells in
the G0 and G1 phases of the cell cycle, while the DNA content of
S-phase cells is intermediate between these values (20).
Apoptosis
Following 24, 48 or 72 h of culture, the cells were
washed twice with cold PBS and resuspended in 1X Binding Buffer (BD
Pharmingen; Becton, Dickinson and Company) at a concentration of
1×106 cells/ml. A total of 100 ml solution
(1×105 cells) was transferred to a 5-ml culture tube.
Then, 5 µl fluorescein isothiocyanate (FITC)-Annexin V (BD
Pharmingen; Becton, Dickinson and Company) and 5 µl propidium
iodide (PI; BD Pharmingen; Becton, Dickinson and Company) were
added to the cells, followed by gently mixing at room temperature
and subsequent incubation for 15 min in the dark. Next, 400 ml 1X
Binding Buffer was added to each test tube. Flow cytometry analysis
(CellQuest™ Pro Software 4.0.2 (BD Biosciences) was performed
within 1 h. Surviving cells were FITC-Annexin V and PI-negative;
early apoptotic cells were FITC-Annexin V-positive and PI-negative;
and late apoptotic or dead cells were FITC-Annexin V and
PI-positive.
Statistical analysis
SPSS 23.0 statistical software (IBM Corp., Armonk,
NY, USA) was used for statistical analysis of the data. Data are
presented as the mean ± standard deviation, and the standard
deviation was used to estimate the sample distribution.
Non-normally distributed data comparisons between groups were
performed via one-way analysis, and Tukey's honestly significant
difference was used as the post hoc test for multiple comparisons.
Spearman's correlation analysis was used to evaluate the
association between qualitative variables. Paired Student's t-test
was used for pairwise comparison between groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Screening of mutations in the target
gene RBPJ from WES sequencing results
A total of 13 patients with PNH patients were
sequenced by 500-depth, of which 61.54% (8/13) exhibited PIG-A gene
mutations. The mutation rate of the PIG-A gene was 61.54% (8/13) in
500-depth sequencing, which was significantly higher than that of
100-depth sequencing (30.77%; 4/13) and consistent with the
literature (21). The mutation types
mainly included frame shift, splicing, stop-gain and non-synonymous
mutations.
The 500-depth sequencing results revealed additional
somatic gene mutations other than those affecting PIG-A. According
to the risk of mutation, it can be divided into five grades, of
which 1–2 was the first mutation explored in the present study
(22). There were 2,399 mutant genes
with a mutation grade of 1 or 2, and 21 had mutation frequencies
>50% [namely, kinesin family member 24, MAGE family member C1
(MAGEC1), fms related tyrosine kinase 3 (FLT3), mediator of DNA
damage checkpoint 1, leukocyte immunoglobulin like receptor B3
(LILRB3), CUX1, zinc finger protein 717 (ZNF717), double homeobox 4
like 4, UBX domain protein 11 (UBXN11), teashirt zinc finger
homeobox 1, filaggrin (FLG), trichohyalin, SUZ12, RBPJ, C-terminal
binding protein 2 (CTBP2), MDS1 and EVI1 complex locus (MECOM),
MUC4, MLL2, POTE ankyrin domain family member H, TET2, and NBPF
member 1]. For the mutant genes mentioned above, the following
screening requirements were also set according to the mutation
forms and the degree of damage of the mutation sites to the
structure of the encoding protein: i) Their mutation rate in the
genome of 1,000 individuals in the 1000G database is <1%; ii)
they contain splicing mutations, frame shifts or non-synonymous
mutations; iii) the software predicts the harmful mutation sites in
the protein structure; and iv) there are mutual mutation sites in
>6 patients. A total of 13 mutant genes met the above
conditions, including FLG, RBPJ, CUX1, MLL2, ZNF717, MAGEC1, MECOM,
CTBP2, SUZ12, MUC4, TET2, UBXN11 and FLT3. Furthermore, 30 patients
with PNH were selected, from whom the mRNA of CD59−
cells was extracted, and the mRNA expression of the above 13 genes
was detected by RT-qPCR (Fig. 1B).
The correlation between PNH clones and clinical markers, relative
expression of mutant gene and hemolysis markers was also analyzed.
Ultimately, the target genes that were highly expressed and have a
significant correlation with PNH hemolysis index were selected to
carry out the following experiments.
The results were as follows: The RBPJ gene was
highly expressed, and the relative expression level of the RBPJ
gene was significantly correlated with the clinical index of PNH
(Table IV; Fig. 1C and D). Upon screening and analyzing
the sequencing results, the highly expressed and first-class
mutation gene RBPJ was evaluated in depth. In total, 69.23% (9/13)
of patients had RBPJ gene mutations, including non-synonymous,
splicing and frame-shift mutations. The mutation sites were as
follows: c.-57-2A>C (exon 2), c.163T>A (exon 12), c.174C>A
(exon 12) and c.177T>A (exon 12).
| Table IV.High expression of RBPJ mRNA in
comparison with controls. |
Table IV.
High expression of RBPJ mRNA in
comparison with controls.
| Gene |
CD59−cell group | Control group | P-value |
|---|
| FLG | 2.310±1.193 | 2.682±1.346 | 0.624 |
| RBPJ | 8.177±3.974 | 3.683±2.661 | 0.044 |
| CUX1 | 2.182±1.454 | 0.780±0.842 | 0.067 |
| MLL2 | 3.517±2.377 | 2.637±1.367 | 0.450 |
| ZNF717 | 6.083±6.377 | 8.137±7.183 | 0.629 |
| MAGEC1 | 2.470±2.276 | 2.803±2.788 | 0.825 |
| MECOM | 3.737±2.946 | 2.303±1.634 | 0.322 |
| CTBP2 | 3.620±2.384 | 6.453±4.262 | 0.186 |
| SUZ12 | 4.087±1.655 | 6.453±4.117 | 0.022 |
| MUC4 | 2.413±1.438 | 6.120±4.023 | 0.041 |
| TET2 | 1.353±0.552 | 5.067±4.675 | 0.002 |
| UBXN11 | 3.940±3.201 | 4.607±2.907 | 0.714 |
| FLT3 | 1.067±0.471 | 2.353±1.990 | 0.006 |
Expression level of RBPJ mRNA is
increased in 30 patients with PNH and significantly correlates with
clinical data
To obtain CD59−/CD59+
neutrophils from 30 PNH patients and CD59+ neutrophils
from 30 healthy controls, immunomagnetic beads cell sorting was
performed in peripheral blood of patients with PNH and healthy
controls on the same day of blood extraction. The RT-qPCR results
of CD59− and CD59+ neutrophils were as
follows: The RBPJ expression levels in the CD59− cell
group were markedly higher than that of the CD59+ cell
of the same patients with PNH and normal control groups (9.54±4.12
vs. 2.37±0.65 and 1.09±0.03, respectively; P1=0.0153 and
P2=0.0074), and the difference between the CD59+ cell
group and the normal group was not statistically significant
(P=0.1305; data not shown).
The correlation between the proportion of PNH clones
and clinical hemolysis indicators was analyzed, and the results
revealed that the PNH clone proportion was negatively correlated
with the levels of Hb (Fig. 1D;
r=−0.6359, P=0.0002), and positively correlated with Ret and LDH
levels (Fig. 1D; r=0.5063, P=0.0043
and r=0.4066, P=0.0258, respectively). This indicated that the
clone size of PNH is directly correlated with the clinical
hemolysis index.
The correlation between the expression of RBPJ mRNA
and hemolysis indicators was also analyzed. The results revealed
that the expression of RBPJ mRNA was negatively correlated with Hb
levels (r=−0.7334, P=0.0074); positively correlated with the
percentage of Ret (r=0.4223, P=0.0201); positively correlated with
LDH levels (r=0.4521, P=0.0121); and positively correlated with PNH
clones (r=0.4757, P=0.0091; Fig.
1E). These results indicate that the expression level of the
RBPJ gene is associated with the hemolytic index in patients with
PNH.
Proliferation of PNH clones is
inhibited upon RBPJ gene knockdown by siRNA constructs in
vitro
A total of 3 siRNA sequences (siRNA1, siRNA2 and
siRNA3) were designed to transfect PNH primary cells in conjunction
with lipo3000 to determine their potential capacity of silencing
the expression of RBPJ (Fig. 2). The
mRNA expression level of RBPJ in the transfected cells was detected
by RT-qPCR. The 3 siRNAs were able to significantly inhibit the
expression of RBPJ. Of them, RBPJ-siRNA1 was the most effective, as
it reduced the expression of RBPJ in PNH primary cells by >75%
(Fig. 2A). Therefore, siRBPJ-1 was
selected to inhibit the expression of RBPJ in PNH primary
cells.
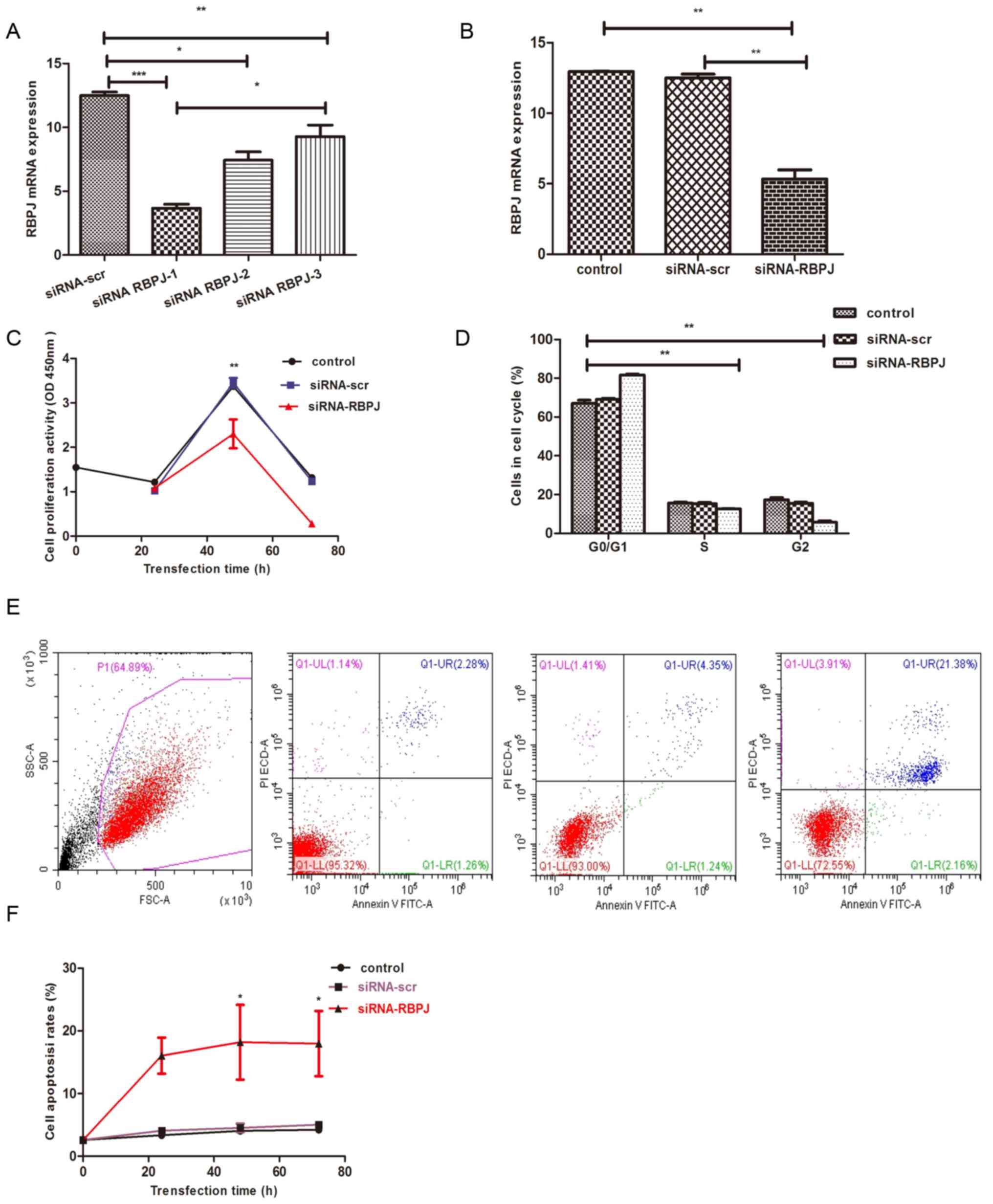 | Figure 2.(A) Evaluation of the inhibitory
effects of siRNAs on RBPJ expression. Quantified RT-qPCR data for
RBPJ in PNH primary cells transfected with siRNA-RBPJ. (B)
Inhibition of RBPJ by siRNA-RBPJ in stable PNH primary cells. PNH
primary cells were infected with RBPJ-siRNA and scr-siRNA to
generate stable clones which were then cultured in media for 72 h
before they were harvested. RBPJ levels were significantly reduced
in PNH primary cells. (C) Cell growth was examined via cell
counting kit-8 assay. Significantly reduced cell growth via RBPJ
inhibition was detected as early as 48 h after seeding, compared
with controls and scr-siRNA. (D) PNH primary cells were analyzed in
a cell cycle phase assay at 72 h after seeding. It was demonstrated
that the percentage of S-phase proliferating cells and G2-phase
post-replicating cells in RBPJ-depleted PNH primary cells was
significantly reduced, whereas the percentage of G0/G1-phase cells
in RBPJ-depleted cells was significantly increased, compared with
control cells. (E and F) Cell apoptosis rate was examined by flow
cytometry. Significantly increased cell apoptosis rate by RBPJ
inhibition was detected as early as 24 h after seeding, compared
with controls and scr-siRNA. *P<0.05, **P<0.01,
***P<0.001. siRNA, small interfering RNA; scr, scrambled
control; PNH, paroxysmal nocturnal hemoglobinuria; RPBJ,
recombinant signal binding protein of immunoglobulin κJ region; OD,
optical density. |
RBPJ-siRNA1 was transfected into PNH primary cells,
which were then cultured in medium and harvested 24–72 h later. The
levels of RBPJ were decreased in transfected PNH primary cells
according to the results of RT-qPCR (Fig. 2B). These data suggest that, upon
transfection of siRNA targeting the RBPJ1 gene into PNH primary
cells, the RBPJ gene was silenced, and its mRNA expression level
was reduced by >75%.
To clarify whether the growth of PNH primary cells
was affected by RBPJ inhibition, CCK-8 assay was used to analyze
the growth ability of PNH primary cells (Table V). Compared with the controls, RBPJ
inhibition was detected 48 h following siRNA transfection, which
significantly reduced cell growth (Fig.
2C). Cell apoptosis rate was analyzed at 24–72 h after
transfection (Table V), and the rate
of cell apoptosis increased following siRNA-RBPJ transfection,
gradually increasing with transfection time (Fig. 2E and F). These results further
suggested that inhibition of RBPJ may prevent the growth of PNH
primary cells by inhibiting cell proliferation.
| Table V.Comparison of proliferative activity
and apoptosis rates of 24, 48 and 72 h before and after siRNA RBPJ
transfection in paroxysmal nocturnal hemoglobinuria primary
cells. |
Table V.
Comparison of proliferative activity
and apoptosis rates of 24, 48 and 72 h before and after siRNA RBPJ
transfection in paroxysmal nocturnal hemoglobinuria primary
cells.
|
| Proliferative
activity (%) | Apoptosis rate
(%) |
|---|
|
|
|
|
|---|
| Time, h | Control | siRNA-scr | siRNA-RBPJ | Control | siRNA-scr | siRNA-RBPJ |
|---|
| 0 | 1.55±0.12 | 0 | 0 | 2.58±0.21 | 0 | 0 |
| 24 | 1.60±0.06 | 1.64±0.06 | 1.48±0.14 | 3.32±0.21 | 3.74±0.43 |
9.55±1.65c |
| 48 | 3.37±0.06 | 3.45±0.18 |
2.30±0.56a | 4.04±0.61 | 4.51±1.16 |
16.93±1.41d |
| 72 | 1.96±0.21 | 1.26±0.39 |
0.49±0.09b | 4.27±0.39 | 5.00±0.52 |
25.71±3.95e |
The cell cycle phases were analyzed 72 h following
inoculation (Table VI). The results
revealed that, compared with the control and scr-transfected cells,
the percentage of S-phase proliferating cells in RBPJ knockdown PNH
primary cells decreased markedly, while the percentage of
G0/G1-phase cells increased (Fig.
2D). These results suggest that inhibition of RBPJ may prevent
the growth of PNH primary cells by inhibiting cell
proliferation.
| Table VI.Comparison of paroxysmal nocturnal
hemoglobinuria primary cell cycle distribution before and after
transfection of siRNA RBPJ. |
Table VI.
Comparison of paroxysmal nocturnal
hemoglobinuria primary cell cycle distribution before and after
transfection of siRNA RBPJ.
| Phase | Control (%) | siRNA-scr (%) | siRNA-RBPJ (%) |
|---|
| G0/G1 | 66.95±6.42 | 66.67±4.89 |
81.66±4.04a |
| S | 16.45±2.08 | 16.92±1.53 |
6.707±1.53b |
| G2 | 16.60±8.18 | 16.40±6.21 |
11.64±2.52c |
Discussion
PNH is a benign clonal disease of hematopoietic stem
cells, the pathogenesis of which remains unclear. PNH clones have
immune-escape characteristics (23,24) and
anti-apoptotic properties (25,26), and
second gene mutations (27–32) may be involved in the amplification of
PNH clones. Next-generation sequencing (NGS) achieves genetic
heterogeneity due to the combination of somatic mutations and
complex clonal structure, reflecting the sequence of genetic
defects, and it is currently widely used in leukemia and other
malignancies (33–36). This sequencing technology has the
following advantages: i) The vast majority of disease variations in
all exome regions of the human genome can be detected; and ii) it
can detect common and low-frequency (i.e., mutation frequency
<5%) mutations that cannot be detected by Sanger sequencing.
Previous sequencing results revealed that genetic
mutations such as TET2, SUZ12, CUX1, RBPJ, MAGEC1 and MLL2 were
identified as the key driving factor for the evolution of myeloid
tumors or cancer clones (10,11,37–40).
The selection and evolution of genetic clones serves a role in
malignant tumors and benign hematological diseases. The mutational
events identified in the present study are not unique to PNH, but
significantly overlap with the spectrum of mutations observed in
myeloid neoplasms and other solid tumors.
In vitro experiments on the highly expressed
and first-class mutated RBPJ gene revealed that the expression of
this gene in CD59− cells was significantly higher than
that in CD59+ cells and controls. Furthermore, its
expression was negatively correlated with Hb levels, and positively
correlated with the percentage of Ret, LDH levels and PNH clones.
These findings suggest that the abnormal expression and mutation of
the RBPJ gene may be closely correlated with PNH clones. As the key
transcription factor of the Notch signaling transduction pathway,
RBPJ can mediate the transcriptional activation of the 4 Notch
receptors in the nucleus and serve a pivotal role in the
integration of Notch signaling pathways, thus being important in
the regulation of the Notch signaling pathway (41,42). The
Notch-RBPJ signaling pathway serves a pivotal role in the
differentiation of T cells in the thymus. The elimination of RBPJ
in early thymus cells can decrease the differentiation αβ T cells
and increase the differentiation of γδ T cells (43,44).
In the resent study, the expression of the RBPJ gene
was validated in patients with PNH and controls, and this gene was
confirmed to be highly expressed in PNH clones. This increased
expression may be an epiphenomenon or even the consequence (rather
than the cause) of PIG-A mutations. The expression of the RBPJ gene
may be positively or negatively regulated by a GPI-linked protein,
which may be upregulated or downregulated. Therefore, RBPJ gene
silencing was conducted in PNH primary cells in vitro, and
the changes in the apoptosis rate and proliferation index were
evaluated to explore the role of the RBPJ gene in PNH.
Previous studies on the Notch-RBPJ signaling pathway
in mice (42,45) revealed that mutation of the Notch
receptor and its downstream key transcription factor RBPJ may be
teratogenic to mouse embryos or lead to early mortality due to
failure of nerve tube closure. The results also revealed that the
Notch/RBPJ signaling transduction pathway was involved in the
growth and development of embryos. Fetal development is one of the
key and pivotal factors regulating the differentiation and
proliferation of neural progenitor cells. The expression of the
RBPJ gene in lung and prostate cancer cells was previously silenced
by Lv et al (46) and Xue
et al (47), and the results
suggested that the downregulation of RBPJ expression leads to a
significant decrease in the growth of cancer cells, which suggests
that RBPJ serves a downstream role in the Notch signaling pathway
and appears to be essential for the functional activation of Notch
signaling.
In the present study, siRNA technology was used to
silence the RBPJ gene in PNH primary cells. Subsequently, the
proportion of cells in the G0/G1 phase increased; the proportion of
S-phase cells decreased; cells were arrested in G0/G1 stage; the
apoptosis rate increased; and the cell proliferation ability
decreased. This suggested that high expression of the RBPJ gene may
be correlated with the proliferation of abnormal PNH clones.
In conclusion, besides mutations in PIG-A, the RBPJ
gene is a highly expressed and first-order mutant gene screened by
NGS, and its abnormal expression may contribute to the
proliferation of PNH clones.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from
National Natural Science Foundation of China (grant nos. 81770110,
81570106 and 81600093), the Tianjin Municipal Natural Science
Foundation (grant nos. 18JCYBJC27200 and 15JCYBJC24300) and Tianjin
Health and Family Planning Commission (grant no.
16ZXMJSY00180).
Availability of data and materials
All data generated or analyzed during our study are
included in this article.
Authors' contributions
RF designed the research and revised the manuscript.
LiyL performed the experiments, analyzed the data and wrote the
manuscript. HL, HW, ZL, YC, LijL, JS and GW contributed to the
experimental work and the collection of patient data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures performed in the present study were
in accordance with the 1964 Helsinki declaration and its later
amendments or comparable ethical standards, and were approved by
the Ethics Committee of Tianjin Medical University (Tianjin,
China). Informed consent was obtained from all individual
participants included in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rotoli B and Luzzatto L: Paroxysmal
nocturnal hemoglobinuria. Semin Hematol. 26:201–207.
1989.PubMed/NCBI
|
|
2
|
Luzzatto L: Recent advances in the
pathogenesis and treatment of paroxysmal nocturnal hemoglobinuria.
F1000Res. 5:2092016. View Article : Google Scholar
|
|
3
|
Parker CJ: The pathophysiology of
paroxysmal nocturnal hemoglobinuria. Exp Hematol. 35:523–533. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hill A, DeZern AE, Kinoshita T and Brodsky
RA: Paroxysmal nocturnal haemoglobinuria. Nat Rev Dis Primers.
3:170282017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schrezenmeier H, Muus P, Socié G, Szer J,
Urbano-Ispizua A, Maciejewski JP, Brodsky RA, Bessler M, Kanakura
Y, Rosse W, et al: Baseline characteristics and disease burden in
patients in the international Paroxysmal Nocturnal Hemoglobinuria
registry. Haematologica. 99:922–929. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Socié G, Mary JY, de Gramont A, Rio B,
Leporrier M, Rose C, Heudier P, Rochant H, Cahn JY and Gluckman E:
Paroxysmal nocturnal hemoglobinuria: Long-term follow-up and
prognostic factors. Lancet. 348:573–577. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
de Latour RP, Mary JY, Salanoubat C,
Terriou L, Etienne G, Mohty M, Roth S, de Guibert S, Maury S, Cahn
JY, et al: Paroxysmal nocturnal hemoglobinuria: Natural history of
disease subcategories. Blood. 112:3099–3106. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li LY, Liu ZY, Liu H, Liu CY, Shao ZH and
Fu R: Deep Sequencing of whole genome exon inParoxysmal Nocturnal
Hemoglobinuria. Am J Hematol. 92:E51–E53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ramdzan ZM and Nepveu A: CUX1, a
haploinsufficient tumour suppressor gene over-expressed in advanced
cancers. Nat Rev Cancer. 14:673–682. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kridel R, Sehn LH and Gascoyne RD:
Pathogenesis of follicular lymphoma. J Clin Invest. 122:3424–3431.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Montagner S, Leoni C, Emming S, Della
Chiara G, Balestrieri C, Barozzi I, Piccolo V, Togher S, Ko M, Rao
A, et al: TET2 regulates mast cell differentiation and
proliferation through catalytic and non-catalytic activities. Cell
Rep. 15:1566–1579. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sugimori C, Padron E, Caceres G, Shain K,
Sokol L, Zhang L, Tiu R, O'Keefe CL, Afable M, Clemente M, et al:
Paroxysmal nocturnal hemoglobinuria and concurrent JAK2(V617F)
mutation. Blood Cancer J. 2:e632012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fraiman YS, Cuka N, Batista D, Vuica-Ross
M and Moliterno AR: Development of paroxysmal nocturnal
hemoglobinuria in CALR-positive myeloproliferative neoplasm. J
Blood Med. 30:107–110. 2016.
|
|
14
|
Chen Y, Tao S, Deng Y, Song L and Yu L:
Chronic myeloid leukemia transformation in a patient with
paroxysmal nocturnal hemoglobinuria: A rare case report with
literature review. Int J Clin Exp Med. 15:8226–8229. 2015.
|
|
15
|
Tominaga R, Katagiri T, Kataoka K, Kataoka
K, Wee RK, Maeda A, Gomyo H, Mizuno I, Murayama T, Ogawa S and
Nakao S: Paroxysmal nocturnal hemoglobinuria induced by the
occurrence of BCR-ABL in a PIGA mutant hematopoietic progenitor
cell. Leukemia. 30:1208–1210. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Orent W, Mchenry AR, Rao DA, White C,
Klein HU, Bassil R, Srivastava G, Replogle JM, Raj T, Frangieh M,
et al: Rheumatoid arthritis-associated RBPJ polymorphism alters
memory CD4+ T cells. Hum Mol Genet. 25:404–417. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nagao H, Setoguchi T, Kitamoto S, Ishidou
Y, Nagano S, Yokouchi M, Abematsu M, Kawabata N, Maeda S, Yonezawa
S and Komiya S: RBPJ is a novel target for rhabdomyosar coma
therapy. PLoS One. 7:e392682012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Parker C, Omine M, Richards S, Nishimura
J, Bessler M, Ware R, Hillmen P, Luzzatto L, Young N, Kinoshita T,
et al: Diagnosis and management of paroxysmal nocturnal
hemoglobinuria. Blood. 106:3699–3709. 2006. View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kalejta RF, Shenk T and Beavis AJ: Use of
a membrane-localized green fluorescent protein allows simultaneous
identification of transfected cells and cell cycle analysis by flow
cytometry. Cytometry. 29:286–291. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shen W, Clemente MJ, Hosono N, Yoshida K,
Przychodzen B, Yoshizato T, Shiraishi Y, Miyano S, Ogawa S,
Maciejewski JP and Makishima H: Deep sequencing reveals stepwise
mutation acquisition in paroxysmal nocturnal hemoglobinuria. J Clin
Invest. 124:4529–4538. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Richards S, Aziz N, Bale S, Bick D, Das S,
Gastier-Foster J, Grody WW, Hegde M, Lyon E, Spector E, et al:
Standards and guidelines for the interpretation of sequence
variants: A joint consensus recommendation of the American College
of Medical Genetics and Genomics and the Association for Molecular
Pathology. Genet Med. 17:405–424. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Karadimitris A and Luzzatto L: The
cellular pathogenesis of paroxysmal noctural hemogolbinuria.
Leukemia. 15:1148–1152. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dacie JV: Paroxysmal nocturnal
haemoglobinuria. Proc R Soc Med. 56:587–596. 1963.PubMed/NCBI
|
|
25
|
Brodsky RA, Vala MS, Barber JP, Medof ME
and Jones RJ: Resistance to apoposis caused by PIG-A mutations in
paroxysmal noctural hemogolbinuria. Proc Natl Acad Sci USA.
94:8756–8760. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Horikawa K, Nakakuma H, Kawaguchi T,
Iwamoto N, Nagakura S, Kagimoto T and Takatsuki K: Apoptosis
resistance of blood cells from patients with paroxysmal nocturnal
hemoglobinuria, aplastic anemia, and myelodysplastic syndrome.
Blood. 90:2716–2722. 1997.PubMed/NCBI
|
|
27
|
Hansen NE and Killmann SA: Paroxysmal
nocturnal hemoglobinuria in myelofibrosis. Blood. 36:428–431.
1970.PubMed/NCBI
|
|
28
|
Luzzatto L, Familusi JB, Williams CK,
Junaid TA, Rotoli B and Alfinito F: The PNH abnormality in
myeloproliferative disorders: Association of PNH and acute
erythremic myelosis in two children. Haematologica. 64:13–30.
1979.PubMed/NCBI
|
|
29
|
Nishimura JI, Inoue N, Azenishi Y, Hirota
T, Akaogi T, Shibano M, Kawagoe K, Ueda E, Machii T and Takeda J:
Analysis of PIG-A gene in a patient who developed reciprocal
translocation of chromosome 12 and paroxysmal nocturnal
hemoglobinuria during follow-up of aplastic anemia. Am J Hematol.
51:229–233. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Inoue N, Izui-Sarumaru T, Murakami Y, Endo
Y, Nishimura J, Kurokawa K, Kuwayama M, Shime H, Machii T, Kanakura
Y, et al: Molecular basis of clonal expansion of hematopoiesis in 2
patients with Paroxysmal nocturnal hemoglobinuria. Blood.
108:4232–4236. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brodsky RA: Paroxysmal nocturnal
hemoglobinuria: Stem cells and clonality. Hematology Am Soc Hematol
Educ Program. 111–115. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Katagiri T, Tominaga R, Kataoka K, Maeda
A, Gomyo H, Mizuno I, Murayama T, Ogawa S and Nakao S: A cure for
paroxysmal nocturnal hemoglobinuria using molecular targeted
therapy specific to a driver mutation. Blood. 126:12152015.
|
|
33
|
Cancer Genome Atlas Research Network, ;
Ley TJ, Miller C, Ding L, Raphael BJ, Mungall AJ, Robertson A,
Hoadley K, Triche TJ Jr, Laird PW, et al: Genomic and epigenomic
landscapes of adult de novo acute myeloid leukemia. N Engl J Med.
368:2059–2074. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Leproust E: Target-enrichment strategies
for next generation sequencing. MLO Med Lab Obs. 44:26–27.
2012.PubMed/NCBI
|
|
35
|
Ng SB, Buckingham KJ, Lee C, Bigham AW,
Tabor HK, Dent KM, Huff CD, Shannon PT, Jabs EW, Nickerson DA, et
al: Exome sequencing identifies the cause of a mendelian disorder.
Nat Genet. 42:30–35. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Warr A, Robert C, Hume D, Archibald A,
Deeb N and Watson M: Exome sequencing: Current and future
perspectives. G3 (Bethesda). 5:1543–1550. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu C, Shi X, Wang L, Wu Y, Jin F, Bai C
and Song Y: SUZ12 is involved in progression of non-small cell lung
cancer by promoting cell proliferation and metastasis. Tumour Biol.
35:6073–6082. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kulic I, Robertson G, Chang L, Baker JH,
Lockwood WW, Mok W, Fuller M, Fournier M, Wong N, Chou V, et al:
Loss of the Notch effector RBPJ promotes tumorigenesis. J Exp Med.
212:37–52. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wong CC, Martincorena I, Rust AG, Rashid
M, Alifrangis C, Alexandrov LB, Tiffen JC, Kober C; Chronic Myeloid
Disorders Working Group of the International Cancer Genome
Consortium, ; Green AR, et al: Inactivating CUX1 mutations promote
tumorigenesis. Nat Genet. 46:33–38. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wienand K and Shires K: The use of MAGE C1
and flow cytometry to determine the malignant cell type in multiple
myeloma. PLoS One. 10:e01207342015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mugnaini E and Floris A: The unipolar
brush cell: A neglected neuron of the mammalian cerebellar cortex.
J Comp Neurol. 339:174–180. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nunzi MG, Birnstiel S, Bhattacharyya BJ,
Slater NT and Mugnaini E: Unipolar brush cells from a glutamatergic
projection system within the mouse cerebellar cortex. J Comp
Neurol. 434:329–341. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sotelo C and Changeux JP: Bergmann fibers
and granular cell migration in the cerebellum of homozygous weaver
mutant mouse. Brain Res. 77:484–491. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gerhardt DM, Pajcini KV, D'altri T, Tu L,
Jain R, Xu L, Chen MJ, Rentschler S, Shestova O, Wertheim GB, et
al: The Notch1 transcriptional activation domain is required for
development and reveals a novel role for Notch1 signaling in fetal
hematopoietic stem cells. Gene Dev. 28:576–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Maillard I, Weng AP, Carpenter AC,
Rodriguez CG, Sai H, Xu L, Allman D, Aster JC and Pear WS:
Mastermind critically regulates Notch-mediated lymphoid cell fate
decisions. Blood. 104:1696–1702. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lv Q, Shen R and Wang J: RBPJ inhibition
impairs the growth of lung cancer. Tumour Biol. 36:3751–3756. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xue L, Li H, Chen Q, Wang Z, Zhang P, Chen
H, Wang Z and Chong T: Inhibition of recombining binding protein
suppressor of hairless (RBPJ) Impairs the growth of prostate
cancer. Cell Physiol Biochem. 36:1982–1990. 2015. View Article : Google Scholar : PubMed/NCBI
|














