Introduction
Stroke is the leading cause of mortality and
disability in China and the second most common cause of mortality
worldwide (1,2). It has previously been reported that
homocysteine (Hcy) is an important risk factor for stroke (3,4). High
levels of Hcy are automatically oxidized to produce reactive oxygen
species (ROS), which may damage the cardiovascular and immune
systems (5). Hcy is recognized as an
independent risk factor for cerebrovascular disease, but its role
in pathogenesis remains unclear.
The blood-brain barrier (BBB) is the structural
basis that maintains the internal environment of the central
nervous system (6). BBB destruction
is an important pathological characteristic of many neurological
diseases (6). Lominadze et al
(7) reported that cerebrovascular
disease was associated with BBB damage, while Kamath et al
(8) reported that high Hcy
concentrations inhibit tight junction (TJ) protein expression in
cerebrovascular endothelial cells, which results in damage to the
TJ structure. Lee's study revealed that high Hcy levels damaged the
BBB structure and increased BBB permeability (9). Tyagi et al (10) suggested that Hcy activates matrix
metalloproteinases (MMPs), whose overexpression reduces the ability
of cells to connect and alters the basement membrane, resulting in
destruction of the blood-brain barrier.
Scutellarin (Scu) is an active component of
Erigerontis Herba, which exhibits anti-inflammatory, antioxidative
and protective effect on endothelial cells (11). Erigerontis Herba has been widely used
in the treatment of cardiovascular and cerebrovascular diseases
(12). Yuan et al (12) reported that Scu inhibited microglial
activation and alleviated the symptoms of neuroinflammation, which
contributed to the clinical treatment of cerebral ischemia. Du
et al (13) demonstrated that
Scu activated the endothelial cGMP activated protein kinase G (PKG)
pathway to protect cerebral vascular endothelial cells.
Furthermore, Scu has been revealed to prevent learning and memory
defects in a rat model of Alzheimer's disease via reducing
oxidative stress and inflammation (14).
Exosomes are cell membrane-derived vesicles that
influence cellular activity by transporting proteins, lipids and
nucleic acids to target cells (15).
It has been reported that exosomes are able to promote functional
recovery and neurovascular plasticity following stroke (16), as well as serving an important role
in the pathogenesis of Alzheimer's disease, Parkinson's disease,
prion disease and other neurological diseases (17). Exosomes may therefore have potential
as clinical treatments for central nervous system diseases. The aim
of the present study was to investigate the expression of TJ
proteins claudin 5, occludin and TJ protein 1 (ZO1) in the BBB
using Hcy-induced rat brain microvascular endothelial cells
(RBMVECs).
Materials and methods
Cell culture
RBMVECs (Saiqi Biological Engineering Co., Ltd.,
Shanghai, China) were seeded in 25-cm2 polystyrene
flasks (Corning Incorporated, Corning, NY, USA) with 4.5 g/ml
glucose Dulbecco's modified Eagle's medium (DMEM) supplemented with
10% heat-inactivated fetal bovine serum (both Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The cells were incubated at
37°C in an atmosphere containing 5% CO2. DMEM was
replaced every 48 h.
Exosome extraction
Exosomes derived from normal RBMVECs following Scu
administration (SE) and exosomes derived from control RBMVECs (CE)
were extracted from cells and incubated with 100 µmol/l Scu or DMEM
for 24 h, respectably. A total of 5 ml supernatant was collected
from each flask and centrifuged at 3,000 × g for 15 min at room
temperature to exclude cell debris. The supernatant was transferred
to a new sterile centrifuge tube and the 1 ml ExoQuick-TC (Tissue
Culture Media Exosome Precipitation Solution; cat. no. 170306-001;
System Biosciences, Palo Alto, CA, USA) was added. The solution was
vortexed overnight at 4°C. The suspension was then centrifuged at
room temperature at 1,500 × g for 30 min. Exosome-containing
pellets were resuspended in 500 µl PBS for intervention
experiments.
Cell viability
Cells were seeded at a density of 1×104
cells/well in 96-well culture plates and incubated for 24 or 48 h
at 37°C in an atmosphere containing 5% CO2. Cells were
incubated with aggregated intervention medications. A total of 10
µl MTT (5 mg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was
added to each well and cells were incubated at 37°C for 4 h. The
culture medium was discarded and 150 µl of dimethyl sulfoxide
(DMSO; Sigma-Aldrich; Merck KGaA) was added to dissolve the
formazan crystals. The number of viable cells in each well was
measured at 490 nm using a microplate reader (Multiskan Mk3; Thermo
Fisher Scientific, Inc.).
Experimental design
After the concentration optimization experiments by
MTT assay, RBMVECs were divided into the following groups: Control,
model, Scu, exosomes derived from CEs and exosomes derived from
normal RBMVECs after SE. The cultured cells in all groups were
pretreated with Scu (100 µmol/l), CE (40% suspension of control
exosomes in 1 ml of DMEM) and SE (40% suspension of Scu exosomes in
of 1 ml DMEM) for 30 min, followed by Hcy exposure (2.5 mmol/l;
cat. no. Z20J8H28854; Shanghai Yuanye Bio-Technology Co., Ltd,
China) for 48 h.
Measurement of lactate dehydrogenase
(LDH) and nitric oxide (NO)
LDH and NO release into the culture medium from dead
cells was assessed using an LDH cytotoxicity assay kit (cat. no.
20170314) and an NO detection kit (cat. no. 20170407; both Nanjing
JianCheng Bioengineering Institute, Nanjing, China). Cells were
lysed with 1% SDS and centrifuged at 4°C, 2,000 × g for 10 min. The
supernatants from each group were assessed according to the
manufacturer's protocol.
ROS detection
Intracellular ROS levels were monitored using
2′,7′-dichlorofluorescein diacetate (DCFH-DA; cat. no. 170V;
Beijing Vigorous Biotechnology Co., Ltd, China) to identify the
role of ROS in Scu and Scu-treated exosomes. RBMVECs were seeded in
24 well plates at a density of 2×105 cells/well and
incubated overnight at 37°C. Cells were subsequently incubated with
or without intervention drugs (Scu, CE and SE) for 48 h. The cells
were then treated with DCFH-DA (5 µM) at 37°C for 90 min.
Fluorescence microscopy (magnification, ×400; U-SPT; Olympus
Corporation, Tokyo, Japan) was used to measure the fluorescence
intensity of the treated cells.
RBMVECs (1×104 in 200 µl) were seeded in
quadruplicate in a 96-well plate and incubated with 2.5 mmol/l Hcy
for 48 h in the presence or absence of intervention drugs (Scu, CE
and SE). The cultured cells were washed three times with PBS (pH
7.4) and incubated in DMEM containing DCFH-DA (5 µM) at 37°C for 90
min. The cells were washed with prewarmed PBS and covered with 100
µl of DMEM. The fluorescence intensity (FI) of each well was
measured using a microplate reader at 485/530 nm.
Detection of cluster of
differentiation (CD)63, claudin-5, ZO1 and occludin by
immunofluorescence
Cells were incubated with 2.5 mmol/l Hcy for 48 h in
the presence or absence of intervention drugs (Scu, CE and SE).
Briefly, cells were grown on cover slips, and 4% paraformaldehyde
was used to fix the cells for 30 min at room temperature. Cells
were subsequently blocked in 10% goat serum (Gibco; Thermo Fisher
Scientific, Inc.) for 20 min at 37°C. Cells were treated with
antibodies against CD63 (cat. no. o67346b; OmnimAbs, Alhambra, CA,
USA), claudin-5 (cat. no. 6679g62), ZO1 (cat. no. 3268f94; both
Affinity Biosciences, USA) and occludin (cat. no. o29813c;
OmnimAbs) for 1 h at 37°C (1:50 dilution). Cells were subsequently
incubated for 1 h at 37°C with corresponding secondary antibodies
conjugated to FITC. Cells were subsequently washed three times with
PBS and stained with DAPI (1 µg/ml in PBS) at 37°C in the dark. The
coverslips were mounted and cells were observed under a light
microscope (U-SPT; Olympus Corp., Tokyo, Japan). Data were analyzed
by using Medical Image Analysis Software 16.0 (MIAS, Warrendale,
WA, USA).
Western blotting
RBMVECs were harvested and lysed using phenylmethane
sulfonyl fluoride-containing lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China). Proteins were quantified using a BCA
protein assay kit (Beyotime Institute of Biotechnology). Equal
amounts of protein (30 µg) were separated by 12% SDS-PAGE and
transferred onto a polyvinylidene fluoride membrane. The membrane
was blocked with 5% nonfat skim milk in TBST for 1 h at room
temperature followed by incubation with primary antibodies against
CD63, claudin-5, ZO1, occludin and GAPDH (cat. no. 10; Cell
Signaling Technology, Inc., Danvers, MA, USA) overnight at 4°C
(1:1,000 dilution). The membrane was washed with TBST and incubated
with horseradish peroxidase conjugated secondary antibodies for 1 h
at 37°C (1:2,000 dilution; cat/no. 7074s; Cell Signaling
Technology, Inc.). Proteins were detected using an ECL detection
kit and Image-Lab software-5.2.1 (both Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Statistical analysis
Data are expressed as the mean ± standard deviation.
Significant differences between groups were determined using
one-way analysis of variance followed by Bonferroni's post hoc test
for multiple comparisons. Correlations between LDH, NO, ROS, CD63,
claudin 5, occludin and ZO1 expression were identified using
Pearson's correlation analysis. All data analyses were performed
using SPSS 16.0 statistical software (SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
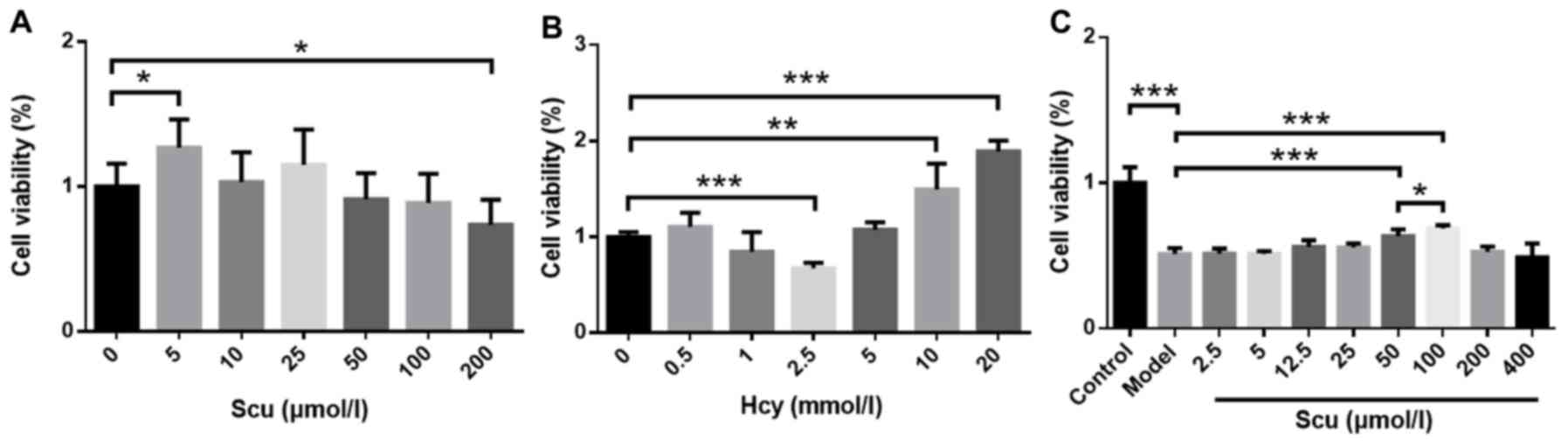
Effect of Scu on Hcy-induced RBMVEC
viability
Cell viability increased in the presence of 5 µmol/l
Scu (P<0.05) (Fig. 1A) and
decreased in the presence of 200 µmol/l Scu (P<0.05) (Fig. 1A) compared with untreated RBMVECs.
Treatment with 2.5 mmol/l Hcy significantly decreased the viability
of RBMVECs compared with the control (P<0.001; Fig. 1B), while viability was increased
following treatment 10–20 mmol/l Hcy (P<0.001) (Fig. 1B). The effects of 2.5–400 µmol/l Scu
on the activity of RBMVECs induced with 2.5 mmol/l Hcy were then
investigated. Cell viability increased significantly in the
presence of 50 or 100 µmol/l Scu compared with the model group
(P<0.001) (Fig. 1C). Furthermore,
100 µmol/l Scu-treated cell viability was significantly higher
compared with 50 µmol/l Scu-treated cells (P<0.05) (Fig. 1C).
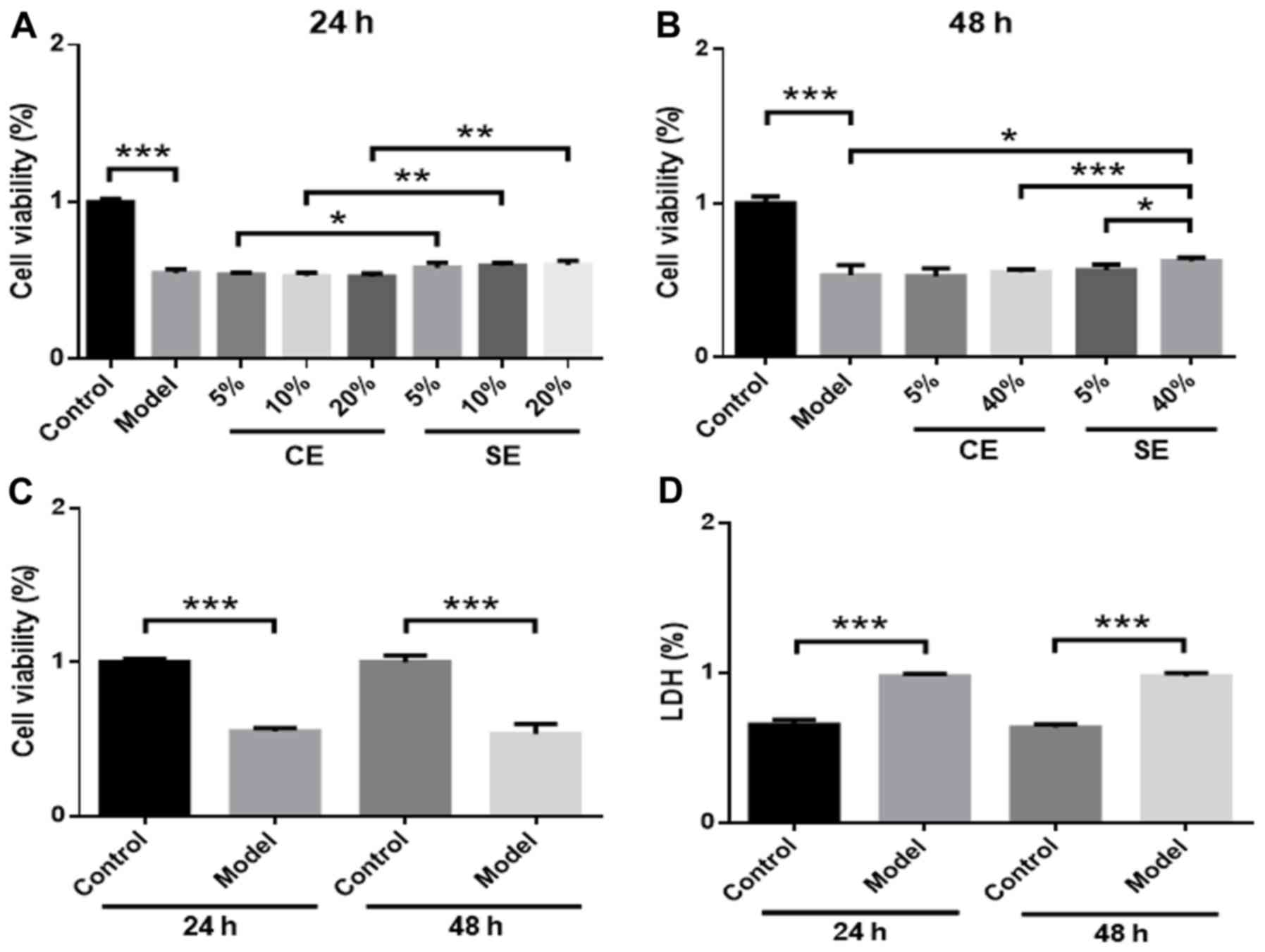
Effects of pretreated exosomes on
Hcy-induced RBMVEC viability
RBMVECs were incubated with different amounts of
exosomes (5, 10, 20 or 40% in DMEM) in the presence of 2.5 mmol/l
Hcy for 24 or 48 h. No significant changes in cell viability were
observed in the 5, 10 or 20% CE and SE groups compared with the
model group (Fig. 2A). However, cell
viability was significantly higher in the SE group compared with
the CE group at 24 h (P<0.05) (Fig.
2A). In the 40% SE group, cell viability was significantly
increased compared with the model group (P<0.05) (Fig. 2B) and the 40% CE group at 48 h
(P<0.001) (Fig. 2B). A
dose-effect relationship was observed in the SE group, as treatment
with 40% solution produced increased cell viability significantly
more than the 5% solution (P<0.05) (Fig. 2B). No significant differences in cell
viability or LDH levels were observed at 24 or 48 h, which suggests
that the effect of Hcy in the model group was not time-dependent
(Fig. 2C and D).
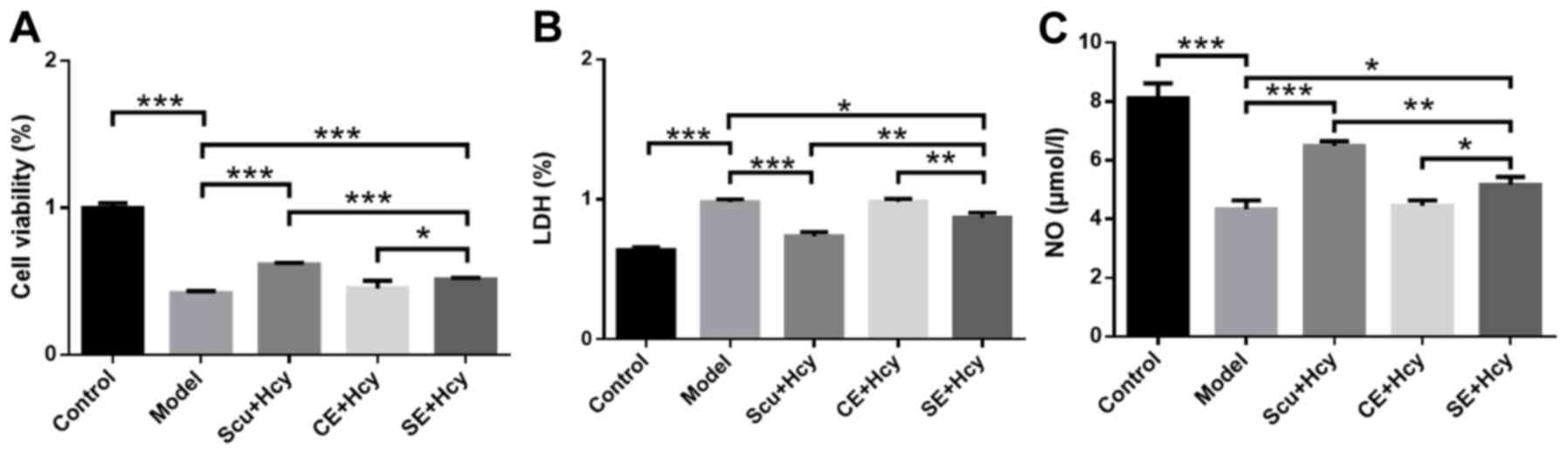
Effect of Scu and Scu-treated exosomes
on cell viability, LDH expression and NO levels
RBMVECs were incubated with 2.5 mmol/l Hcy for 48 h
along with Scu and Scu-treated exosomes. Cell viability and NO
levels in the Scu and SE group were significantly increased
(P<0.05) (Fig. 3A) compared with
the model group, while LDH levels were significantly decreased
(P<0.05) (Fig. 3B). In addition,
cell viability and NO levels were significantly higher in the Scu
group compared with the SE group (P<0.01) (Fig. 3A and C), while LDH levels were
significantly lower in the Scu group compared with the SE group
(P<0.01) (Fig. 3B). Cell
viability and NO levels were significantly increased (P<0.01)
(Fig. 3A and C) and LDH levels were
significantly decreased (P<0.05; Fig.
3B) in the SE group compared with the CE group.
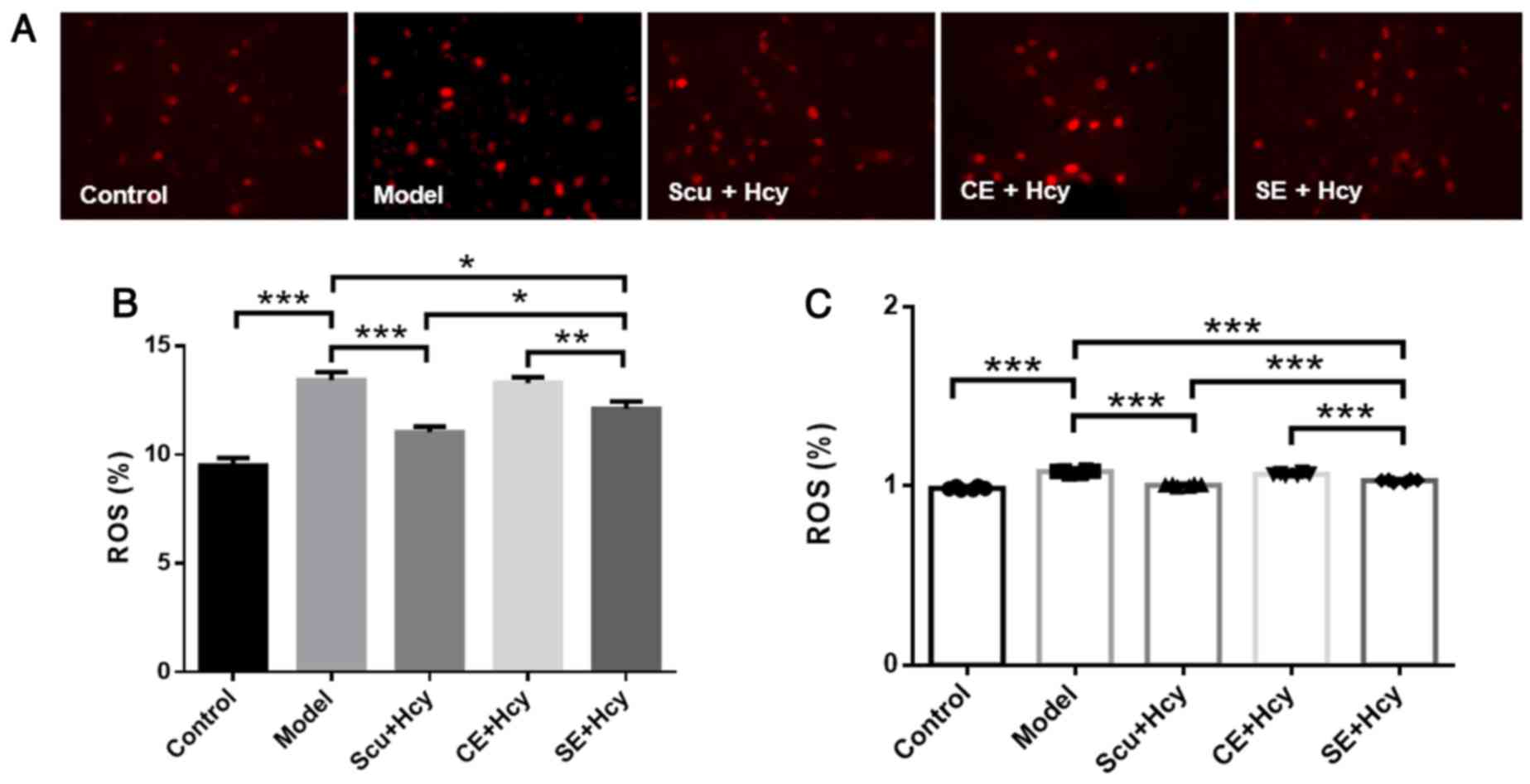
Effect of Scu and Scu-treated exosomes
on ROS
Cells were treated as described above, and ROS
levels were detected using fluorescence and spectrophotometry. ROS
levels were significantly decreased in the Scu and SE groups
compared with the model group (P<0.05) (Fig. 4). ROS levels were reduced in the Scu
and SE groups compared with the CE groups (P<0.01) (Fig. 4).
Effect of Scu and Scu-treated exosomes
on CD63, claudin-5, occludin and ZO1 expression
The detailed mechanisms underlying the effect of Scu
and Scu-treated exosomes were investigated. CD63, claudin-5,
occludin and ZO1 expression was significantly increased in the Scu
and SE groups compared with the model group (P<0.05) (Figs. 5 and 6). CD63, claudin-5, occludin and ZO1
expression was significantly increased in the Scu group compared
with the SE group (P<0.05) (Figs.
5 and 6). CD63, claudin-5,
occludin and ZO1 expression was significantly higher in the SE
group compared with the CE group (P<0.05) (Figs. 5 and 6).
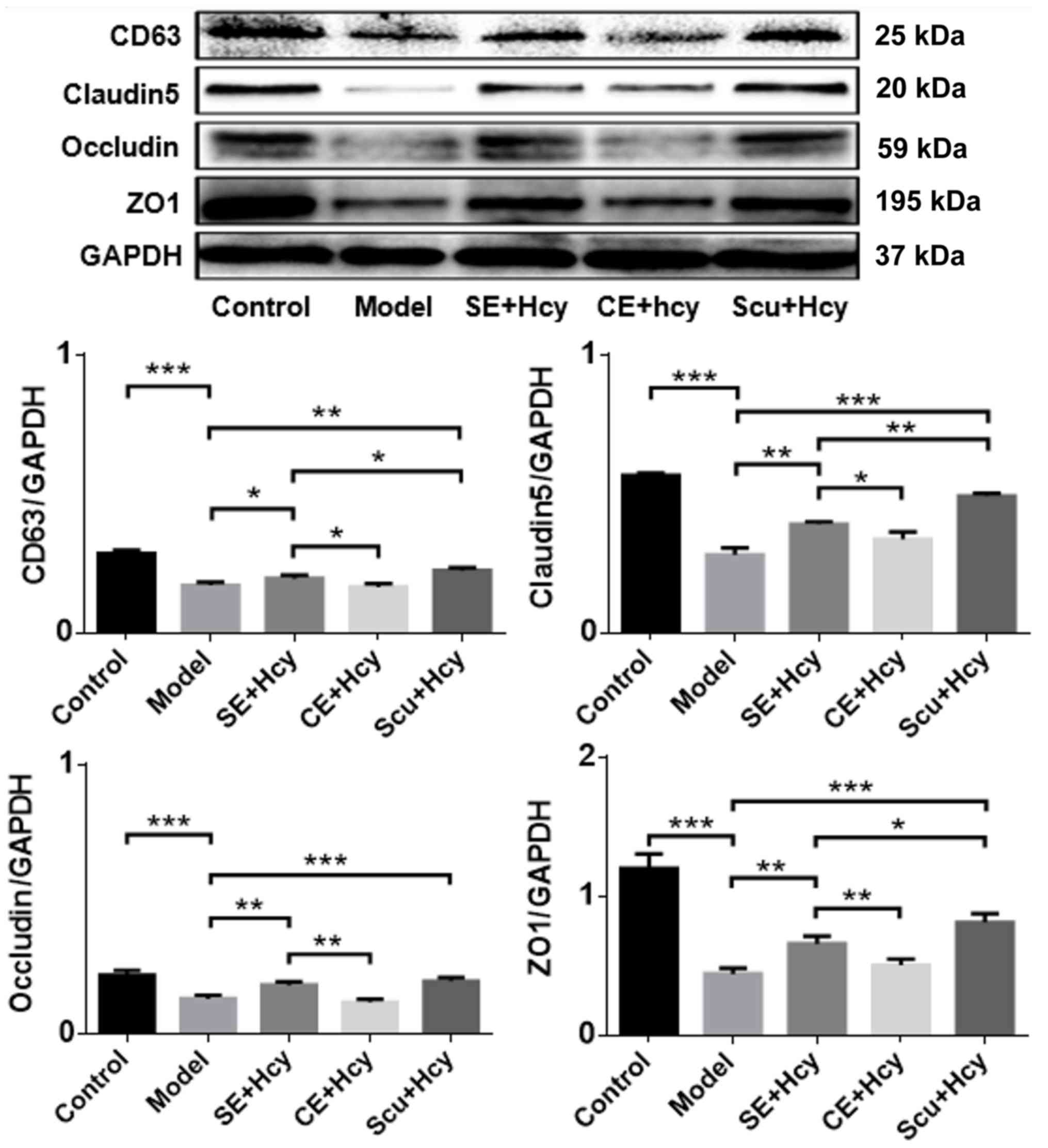 | Figure 5.Representative western blot and
quantified results for CD63, claudin-5, occludin and ZO1
expression. *P<0.05, **P<0.01 and ***P<0.001 with a bar
linking the groups with an asterisk. n=3. CD, cluster of
differentiation; ZO1, tight junction protein 1; Model, 2.5 mmol/l
Hcy-induced rat brain microvascular endothelial cells; CE,
control-treated exosomes; Scu, Scutellarin; SE, Scu-treated
exosomes; Hcy, homocysteine. |
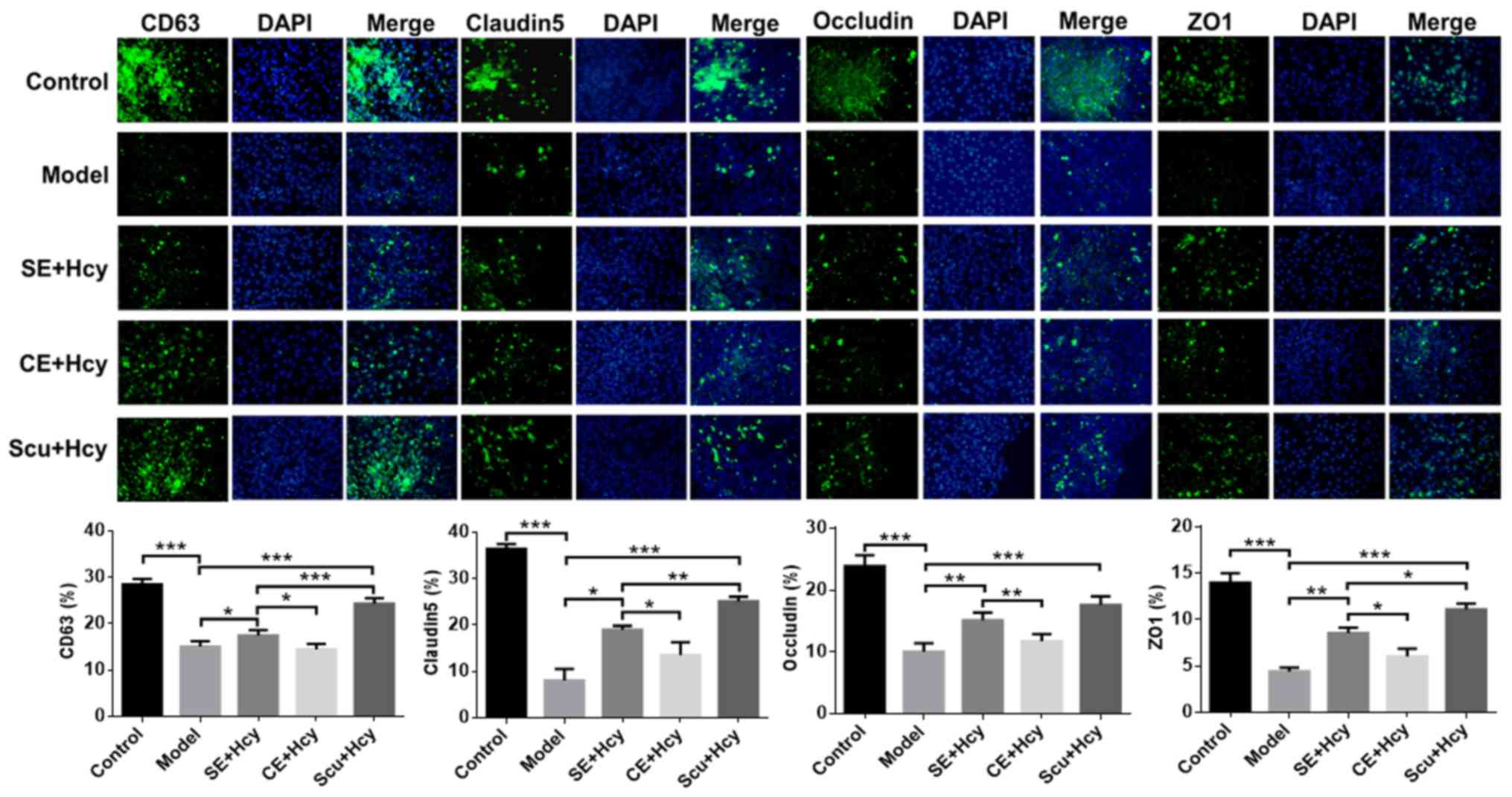 | Figure 6.Immunofluorescence of CD63,
claudin-5, occludin and ZO1 expression. Magnification, ×400; n=3.
*P<0.05, **P<0.01 and ***P<0.001 with a bar linking the
groups with an asteris. CD, cluster of differentiation; ZO1, tight
junction protein 1; Scu, Scutellarin; SE, Scu-treated exosomes;
Hcy, homocysteine; CE, control-treated exosomes; Model, 2.5 mmol/l
Hcy-induced rat brain microvascular endothelial cells. |
Correlations between experimental
indicators
The association of LDH levels with NO, ROS, CD63,
claudin-5, occludin and ZO1 expression was assessed using Pearson's
correlation analysis. The results indicated that LDH expression was
significantly negatively correlated with NO, CD63, claudin-5,
occludin and ZO1 levels (P<0.05) (Table I). Furthermore, NO levels were
significantly negatively correlated with ROS expression (P<0.05)
(Table I), while ROS expression was
significantly negatively correlated with CD63, claudin-5, occludin
and ZO1 levels (P<0.05) (Table
I). Significant positive correlations were observed between
CD63 and claudin-5, CD63 and occludin, and CD63 and ZO1 expression
(all P<0.01) (Table I). Claudin-5
expression was significantly positively correlated with occludin
and ZO1 levels (P<0.01) (Table
I).
 | Table I.Correlation analysis. |
Table I.
Correlation analysis.
|
| LDH | NO | ROS | CD63 | Claudin-5 | Occludin | ZO1 |
|---|
| LDH | 1.000 |
|
|
|
|
|
|
| NO | −0.891a | 1.000 |
|
|
|
|
|
| ROS | 0.277 | −0.889a | 1.000 |
|
|
|
|
| CD63 | −0.721a | −0.327 | −0.866a | 1.000 |
|
|
|
| Claudin-5 | −0.721a | −0.327 | −0.866a | 0.999b | 1.000 |
|
|
| Occludin | −0.721a | −0.327 | −0.866a | 0.999b | 1.000b | 1.000 |
|
| ZO1 | −0.721a | −0.327 | −0.866a | 0.999b | 1.000b | 1.000b | 1.000 |
Discussion
High Hcy levels induce neuronal apoptosis and BBB
destruction via peroxidation, which eventually leads to stroke
(18). A recent study demonstrated
that TJs close the gap between cerebral vascular endothelial cells,
are involved in the blood-brain barrier, transmit intracellular
signals and maintain cell growth and differentiation, thereby
protecting against neurological diseases (19).
Exosomes are an important intercellular mediator in
post-stroke nerve repair and injury, with beneficial effects for
patient recovery (20). Scu
increases blood flow, dilates blood vessels and lowers blood
prophylaxis (12). However, whether
high Hcy levels damage the structure and function of the BBB via TJ
destruction has not previously been reported. The aim of the
present study was to clarify the protective mechanisms of Scu and
Scu-treated exosomes on Hcy-induced RBMVECs.
The effects of Scu and Scu-treated exosomes on
Hcy-induced RBMVECs were investigated via assessing the biological
activity of cells following treatment with Hcy at different
concentrations and for different intervals. An LDH detection kit
was used to measure changes in LDH release rate. Oxidative
stress-related injury and apoptosis increase cell membrane
permeability, resulting in the release of LDH (21). As such, LDH activity in the cell
culture medium is proportional to cell death (22). For normal RBMVECs, 5–200 µmol/l Scu
was not observed to cause significant toxicity at 24 h. However,
2.5 mmol/l Hcy was observed to be toxic to normal RBMVECs, and so
this concentration was selected for subsequent experiments. It was
also demonstrated that 50–100 µmol/l Scu protected RBMVECs against
Hcy-induced toxicity. Normal exosomes and Scu-treated exosomes
(0–20%) produced no significant effects on 2.5 mmol/l Hcy-induced
RBMVECs after 24 h. However, 40% Scu-treated exosomes had a
significant protective effect on cells in the model group after 48
h. Scu-treated exosomes were collected from the cell culture medium
following treatment, and the exosomes concentration likely did not
reach a maximum. Therefore, a higher concentration (40%) of
Scu-treated exosomes and a longer exposure duration (48 h) were
required to produce significant pharmacological effects. The
effects of Hcy on cell viability at different time points (24 and
48 h) were investigated using MTT and LDH assays. The results
indicated that Hcy-induced damage was not time-dependent in
RBMVECs. As such, a treatment duration of 48 h was selected to
ensure consistency across groups. Scu and Scu-treated exosomes
reduced Hcy-induced cell damage in the model group, with a greater
efficacy observed in the Scu group compared with the SE group in
four cell injury measurements.
Hcy produces ROS via self-oxidation, which causes
peroxidation of the cell membrane. O2− formed through
the oxidation process combines with NO and reduces the
bioavailability of NO (23). ROS
also oxidizes NOS and reduces its NO production capacity (24). Changes in endothelial NO biological
activity may be an important marker of vascular endothelial damage
(25). NO levels were significantly
decreased in the model group, which suggests that Hcy decreased NO
activity via superoxide activation and induced cerebral vascular
endothelial injury. NO levels in the Scu and SE groups were
significantly higher compared with the group, which indicated that
Scu and Scu-treated exosomes exerted protective effects on cells in
the model group. However, the efficacy of Scu was greater than that
of Scu-treated exosomes. ROS levels were increased in the model
group, confirming that Hcy causes cell damage. ROS levels in the
Scu and SE groups were significantly lower compared with the model
group, suggesting that RBMVECs in the Scu and SE groups suffered
less Hcy-induced damage.
Exosome-mediated communication between cells is
achieved via the transfer of biomolecules from a source cell to a
target cell (26). CD63 is a
lysosomal membrane glycoprotein that is a surface marker protein
for exosomes (27). In the present
study, CD63 expression was increased in the SE group compared with
the Scu group, which in turn was higher compared with the CE group
These results suggest that exosomes were enriched in the SE group
and provided a certain resistance to the toxic effects of Hcy.
These results suggest that Scu is able to stimulate cells in the
model group to secrete exosomes. However, this experiment primarily
measured the pharmacological effects of Scu.
It has previously been demonstrated that Scu and
Scu-treated exosomes exhibit protective effects on Hcy-induced
RBMVECs, suggesting that their protective mechanism reduces
Hcy-mediated damage to TJ proteins in the BBB. ZO1 is located in
the subcapsular part of the TJ of epithelial cells, which are
interconnected via a variety of proteins (28). Occludin and claudin form the primary
chain of the linear TJ structure (29). Occludin is directly involved in TJ
formation in brain microvascular endothelial cells and its
expression is associated with the degree of epithelial cell closure
(30). Claudin serves a role in the
formation of TJs, but is not specifically expressed in TJs
(31). In the present study,
claudin-5, occludin and ZO1 expression was significantly reduced in
the model group, suggesting that Hcy damages TJ proteins in the
BBB. Claudin-5, occludin and ZO1 expression increased significantly
following the administration of Scu and its exosomes, with greater
efficacy in the Scu group compared with the SE group. These results
suggest that Scu and its exosomes protect cells by increasing the
expression of TJ proteins in the BBB.
Correlation analyses were performed to clarify the
associations between experimental indicators. LDH levels were
significantly and negatively correlated with NO levels, which
suggests that cell damage was associated with oxidative stress. LDH
expression was also significantly and negatively correlated with
claudin-5, occludin and ZO1 expression, indicating that cell injury
and TJ proteins are closely related. ROS levels were negatively
correlated with CD63 expression, which suggests that oxidative
stress was associated with decreased lysosomal glycoprotein levels.
Finally, CD63 expression was significantly and positively
correlated with claudin-5, occludin and ZO1 expression. Together,
these results suggest that an increase in lysosomal glycoprotein
levels or in exosomes is associated with the integrity of TJ
proteins.
In conclusion, Scu and Scu-treated exosomes
increased NO levels as well as CD63, claudin-5, occludin and ZO1
expression in Hcy-induced RBMVECs, while LDH and ROS levels were
reduced. Together, these effects reduce cell damage and protect the
structure and function of the BBB. However, the present study was
limited by the lack of accurate identification of exosomes and the
lack of in-depth exosome study.
Acknowledgements
Not applicable.
Funding
The present study was supported by the China
Postdoctoral Science Foundation (grant no. 2016M592513), the
Project of Administration of Traditional Chinese Medicine of
Guangdong Province of China (grant no. 20181114), Guangzhou Science
and Technology Project (grant no. 201508020050) and the Guangzhou
Science and Technology Project (grant no. 201604020003).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XZ and MD conceived the study, designed the
experiments, analyzed the data and prepared the manuscript. MD and
MZ were major contributors in writing the manuscript. XZ, CL, MD
and MZ selected the subject and obtained samples for the present
study. XZ, MD and MZ performed the experiments. All authors have
read and approved the manuscript.
Ethics approval and consent to
participate
The Ethics Committee of the Guangdong Provincial
Hospital of Chinese Medicine (Guangzhou, China) approved the study
protocol and all participants provided written informed
consent.
Patient consent for publication
All participants provided written informed consent
for publication.
Competing interests
The authors declare no conflicts of interest.
References
|
1
|
Yang G, Wang Y, Zeng Y, Gao GF, Liang X,
Zhou M, Wan X, Yu S, Jiang Y, Naghavi M, et al: Rapid health
transition in China, 1990–2010: Findings from the global burden of
disease study 2010. Lancet. 381:1987–2015. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lozano R, Naghavi M, Foreman K, Lim S,
Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, et
al: Global and regional mortality from 235 causes of death for 20
age groups in 1990 and 2010: A systematic analysis for the global
burden of disease study 2010. Lancet. 380:2095–2128. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He Y, Li Y, Chen Y, Feng L and Nie Z:
Homocysteine level and risk of different stroke types: A
meta-analysis of prospective observational studies. Nutr Metab
Cardiovasc Dis. 24:1158–1165. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao M, Wang X, He M, Qin X, Tang G, Huo
Y, Li J, Fu J, Huang X, Cheng X, et al: Homocysteine and stroke
risk: Modifying effect of methylenetetrahydrofolate reductase C677T
polymorphism and folic acid intervention. Stroke. 48:1183–1190.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boldyrev AA: Molecular mechanisms of
homocysteine toxicity. Biochemistry (Mosc). 74:589–598. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wallin A, Sjögren M, Edman A, Blennow K
and Regland B: Symptoms, vascular risk factors and blood-brain
barrier function in relation to CT white-matter changes in
dementia. Eur Ncuro1. 44:229–235. 2000.
|
|
7
|
Lominadze D, Roberts AM, Tyagi N, Moshal
KS and Tyagi SC: Homocy-steine causes cerebrovascular leakage in
mice. Am J Physiol Heart Circ Physiol. 290:H1206–H1213. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kamath AF, Chauhan AK, Kisucka J, Dole VS,
Loscalzo J, Handy DE and Wagner DD: Elevated levels of homocysteine
compromise blood-brain barrier integrity in mice. Blood.
107:591–593. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee H, Kim JM, Kim HJ, Lee I and Chang N:
Folic acid supplementation can reduce the endothelial damage in rat
brain microvasculature due to hyperhomocysteinemia. J Nutr.
135:544–548. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tyagi SC, Lominadze D and Roberts AM:
Homocysteine in microvascular endothelial cell barrier
permeability. Cell Biochem Biophys. 43:37–44. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mo J, Yang R, Li F, Zhang X, He B, Zhang
Y, Chen P and Shen Z: Scutellarin protects against vascular
endothelial dysfunction and prevents atherosclerosis via
antioxidation. Phytomedicine. 42:66–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan Y, Fang M, Wu CY and Ling EA:
Scutellarin as a potential therapeutic agent for microglia-mediated
neuroinflammation in cerebral ischemia. Neuromolecular Med.
18:264–273. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Du X, Chen C, Zhang M, Cai D, Sun J, Yang
J, Hu N, Ma C, Zhang L, Zhang J and Yang W: Scutellarin reduces
endothelium dysfunction through the PKG-I pathway. Evid Based
Complement Alternat Med. 2015:4302712015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo LL, Guan ZZ, Huang Y, Wang YL and Shi
JS: The neurotoxicity of β-amyloid peptide toward rat brain is
associated with enhanced oxidative stress, inflammation and
apoptosis, all of which can be attenuated by scutellarin. Exp
Toxicol Pathol. 65:579–584. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Conlan RS, Pisano S, Oliveira MI, Ferrari
M and Mendes Pinto I: Exosomes as reconfigurable therapeutic
systems. Trends Mol Med. 23:636–650. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xin H, Li Y, Cui Y, Yang JJ, Zhang ZG and
Chopp M: Systemic administration of exosomes released from
mesenchymal stromal cells promote functional recovery and
neurovascular plasticity after stroke in rats. J Cereb Blood Flow
Metab. 33:1711–1715. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Janas AM, Sapoń K and Janas T, Stowell MH
and Janas T: Exosomes and other extracellular vesicles in neural
cells and neurodegenerative diseases. Biochim Biophys Acta.
1858:1139–1151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lehotsky J, Petras M, Kovalska M, Tothova
B, Drgova A and Kaplan P: Mechanisms involved in the ischemic
tolerance in brain: Effect of the homocysteine. Cell Mol Neurobiol.
35:7–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu CY, Fang M, Karthikeyan A, Yuan Y and
Ling EA: Scutellarin attenuates microglia-mediated
neuroinflammation and promotes astrogliosis in cerebral ischemia-a
therapeutic consideration. Curr Med Chem. 24:718–727. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Corrigan F, Mander KA, Leonard AV and Vink
R: Neurogenic inflammation after traumatic brain injury and its
potentiation of classical inflammation. J Neuroinflammation.
13:2642016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun X, Chen RC, Yang ZH, Sun GB, Wang M,
Ma XJ, Yang LJ and Sun XB: Taxifolin prevents diabetic
cardiomyopathy in vivo and in vitro by inhibition of oxidative
stress and cell apoptosis. Food Chem Toxicol. 63:221–232. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang J, Ju B, Yan Y, Xu H, Wu S, Zhu D,
Cao D and Hu J: Neuroprotective effects of phenylethanoid
glycosides in an in vitro model of Alzheimer's disease. Exp
Ther Med. 13:2423–2428. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang JS, Bojovic D, Chen Y and Lindgren
CA: Homocysteine sensitizes the mouse neuromuscular junction to
oxidative stress by nitric oxide. Neuroreport. 29:1030–1035. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang X, Cui L, Joseph J, Jiang B, Pimental
D, Handy DE, Liao R and Loscalzoa J: Homocysteine induces
cardiomyocyte dysfunction and apoptosis through p38 MAPK-mediated
increase in oxidant stress. J Mol Cell Cardiol. 52:753–760. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sydow K, Hornig B, Arakawa N, Bode-Böger
SM, Tsikas D, Münzel T and Böger RH: Endothclial dysfunction in
patients with peripheral artcrial disease and chronic
hyperhomocys-teinemia:potential role of ADMA. Vasc Med. 9:93–101.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Stremersch S, Vandenbroucke RE, Van
Wonterghem E, Hendrix A, De Smedt SC and Raemdonck K: Comparing
exosome-like vesicles with liposomes for the functional cellular
delivery of small RNAs. J Control Release. 232:51–61. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang HG and Grizzle WE: Exosomes: A novel
pathway of local and distant intercellular communication that
facilitates the growth and metastasis of neoplastic lesions. Am J
Pathol. 184:28–41. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiao H, Wang Z, Liu Y, Wang P and Xue Y:
Specific role of tight junction proteins claudin-5, occludin, and
ZO-1 of the blood-brain barrier in a focal cerebral ischemic
insult. J Mol Neurosci. 44:130–139. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Y, Li Q, Pan CS, Yan L, Hu BH, Liu YY,
Yang L, Huang P, Zhao SY, Wang CS, et al: Bushen huoxue attenuates
diabetes-induced cognitive impairment by improvement of cerebral
microcirculation: Involvement of RhoA/ROCK/moesin and src signaling
pathways. Front Physiol. 9:5272018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Furuse M, Hirase T, Itoh M, Nagafuchi A,
Yonemura S and Tsukita S: Occludin: A novel integral membrane
protein localizing at tight junctions. J Cell Biol. 123:1777–1788.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ohtsuki S, Sato S, Yamaguchi H, Kamoi M,
Asashima T and Terasaki T: Exogenous expression of claudin-5
induces barrier properties in cultured rat brain capillary
endothelial cells. J Cell Physiol. 210:81–86. 2007. View Article : Google Scholar : PubMed/NCBI
|