Introduction
Bone homeostasis mainly relies on the balance
between bone-forming cells (osteoblasts) and bone-resorptive cells
(osteoclasts) (1). The imbalance
between bone formation and resorption leads to skeletal diseases
associated with progressive bone degeneration, such as
osteoporosis. Osteoporosis is a disease characterized by lower than
normal bone mass and greater than normal bone loss, and is
associated with increased risk of bone weakness and fracture
(2). Studies on osteoblastic
differentiation may help elucidate the pathogenesis of
osteoporosis.
Transforming growth factor (TGF)-β1 is a polypeptide
member of the TGF family that is involved in cellular functions,
including the control of cell growth, proliferation and
differentiation (3). TGF-β1 can
maintain postnatal bone mass by coupling bone resorption and bone
formation (4). TGF-β1 favors
osteoclast differentiation by binding to a receptor on osteoclasts,
and thereby activating Smad2/3, which is directly associated with
the TRAF6-TAB1-TAK1 complex (5).
The possible mechanisms of osteoblastic
differentiation and bone formation include the coordination of cell
signals and the activity of post-transcriptional factors. MicroRNAs
(miRNAs/miRs) serve an important role in osteoblastic
differentiation. miRNAs are small non-coding single-stranded RNAs
that inhibit the translation or promote the degradation of target
mRNAs through binding to their 3′-untranslated region (UTR), and
thereby attenuate protein synthesis (6). Numerous miRNAs have been characterized
in diverse biological and pathological processes, including cell
proliferation, apoptosis, differentiation and tumorigenesis
(7–10). There is also evidence that aberrant
miRNA expression is strongly associated with osteoblast
differentiation through various mechanisms (11). miR-877-3p has been demonstrated to
promote cell proliferation and differentiation in lung and bladder
cancer (12,13). In addition, miR-877-3p has been
revealed to inhibit the myofibroblast differentiation of
lung-resident mesenchymal stem cells and attenuate the lung
fibrosis induced by bleomycin (12).
However, it is unclear whether miR-877-3p is one of the key miRNAs
that regulate the proliferation and differentiation of
osteoblasts.
The aim of the present study was to define the role
of miR-877-3p in osteoblast differentiation using MC3T3-E1 cells,
an osteoblast precursor cell line derived from mouse calvaria that
has been widely used as a model in bone biology research (14). Results demonstrated that miR-877-3p
was upregulated during TGF-β1-mediated osteoblastic
differentiation. Bioinformatics analysis identified Smad7 as a
target of miR-877-3p. Osteoblastic differentiation of MC3T3-E1
cells was induced by miR-877-3p overexpression whilst Smad7
overexpression partially reversed this effect. Therefore,
miR-877-3p/Smad7 may represent a potential therapeutic target for
bone regeneration-associated diseases.
Materials and methods
Cell culture and osteoblastic
differentiation
The MC3T3-E1 cell line was obtained from the Chinese
Academy of Sciences Cell Bank (Shanghai, China) and cultured in
α-minimal essential medium (α-MEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) containing penicillin (50 U/ml), 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and
streptomycin (50 µg/ml) at 37°C in a humidified atmosphere with 5%
CO2. For TGF-β1 treatment, recombinant human TGF-β1
(cat. no. CSB-AP003861HU; CUSABIO Technology LLC, Wuhan, China)
with final concentration of 4 ng/ml was added to cell culture for 7
or 14 days. To induce osteoblastic differentiation, cells were
seeded (1×105 cells per well) in 6-well plates and
treated with osteogenic medium (10 ng/ml β-glycerophosphate; 10
mmol/l dexamethasone and 50 µg/ml vitamin C; all from
Sigma-Aldrich; Merck KGaA) for 7 and 14 days when they reached 70%
confluence (15).
Cell transfection
pcDNA3.1-Smad negative control (NC) and
pcDNA3.1-Smad7 were synthesized by GenePharma Co., Ltd. (Jiangsu,
China). miR-877-3p mimics, inhibitor and their corresponding
negative controls (NCs), miR-877-3p agomir, miR-877-3p antagomir
and their corresponding controls (mimics NC: miR01101, RiboBio Co.,
Ltd., and inhibitor NC; miR02101; RiboBio Co., Ltd.) were
synthesized by Guangzhou RiboBio Co., Ltd. (Guangzhou, China). The
sequences were as follows: miR-877-3p mimics,
5′-UGUCCUCUUCUCCCUCCUCCCA-3′; miR-877-3p inhibitor,
5′-UGGGAGGAGGGAGAAGAGGACA-3′; miR-877-3p agomir,
5′-UGUCCUCUUCUCCCUCCUCCCA-3′ and miR-877-3p antagomir,
5′-UGGGAGGAGGGAGAAGAGGACA-3′. MC3T3-E1 cells were seeded in 6-well
plates at a density of 30,000 cells/well and cultured for 24 h in
antibiotic-free fresh medium. Subsequently, the cells were
transfected with miRNA or vector (100 nM) using Lipofectamine 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. After 48 h incubation, the cells were used
for further experiments.
RNA isolation and RT-qPCR
Total RNA was extracted from cells or tissue with
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. Total RNA was quantified
by Nanodrop 2000 spectrophotometry (Thermo Fisher Scientific,
Inc.). First-strand cDNA synthesis was then performed using a
Superscript RT kit (Biouniquer Technology, Nanjing, China)
following the manufacturer's instructions. The reaction was
performed under the following conditions: 37°C for 15 min and 98°C
for 5 min. qPCR was conducted with SYBR Green I (Biouniquer
Technology) using the LightCycler 480 II System (Roche Diagnostics,
Basel, Switzerland). Primer sequences were as follows:
miR-877-3p-F, 5′-GCGTGTATTTGACAAGCTGAGTTG-3′; collagen type I α1
chain (COL1A1) forward, 5′-TCGTGGTGAGACTGGTCCTG-3′ and reverse,
5′-TGTCACCTTGTTCGCCTGTC-3′; runt-related transcription factor 2
(RUNX2) forward, 5′-CGGACGAGGCAAGAGTTTC-3′ and reverse,
5′-TGGCTCAGATAGGAGGGGTAA-3′; osterix (OSX) forward,
5′-AGCAGCAGCAGCAACAGAAG-3′ and reverse, 5′-CTCAGGCCAGCTCACTCTTG-3′;
Smad7 forward, 5′-GCTATTCCAGAAGATGCTGTTC-3′ and reverse,
5′-GTTGCTGAGCTGTTCTGATTTG-3′; β-actin forward
5′-GTCCCTCACCCTCCCAAAAG-3′ and reverse,
5′-GCTGCCTCAACACCTCAACCC-3′. The thermocycling conditions were as
follows: 95°C for 5 min, followed by 35 cycles at 95°C for 10 sec
then 60°C for 30 sec. The 2−ΔΔCq method was used for
data analysis with normalization to β-actin (16). U6 (cat. no. B532461-0001; Sangon
Biotech Co., Ltd., Shanghai, China) was used as endogenous control
for the analysis of miR-877-3p expression.
Measurement of alkaline phosphatase
(ALP) activity
Total proteins were extracted from cells using a
radioimmunoprecipitation assay (RIPA) lysis buffer (Biouniquer
Technology) and quantified by bicinchoninic acid assay according to
the manufacturer's protocol. The cell lysate was added to 96-well
plates and incubated with ALP staining solution (Sigma-Aldrich;
Merck KGaA) for 60 min at 37°C. Following the addition of the stop
solution, ALP activity was measured spectrophotometrically at 405
nm, and normalized to the total protein concentration.
Mineralization assay
The MC3T3-E1 cells were cultured with TGF-β1 (4
ng/ml) and osteoblastic differentiation medium for 7 days in
24-well plates at a density of 2×104 cells per well, and
the mineralization of the MC3T3-E1 cells was observed via Alizarin
Red S (ARS) and ALP staining with an optical light microscope (ARS,
Solarbio, China; ALP staining kit, Beyotime Institute of
Biotechnology, Haimen, China).
In vivo transplantation
A total of 24 female NOD/SCID mice (age, 4 weeks;
weight, 17–19 g) were purchased from the Provincial Animal Center
(Guangdong, China) and housed for 1 week to adapt them to the
laboratory environment (24°C, 12-h light/dark cycles and humidity
60±10%) with free access to water and food. The mice were randomly
divided into four groups (each n=6). MC3T3-E1 cells transfected
with miR-877-3p mimics, miR-877-3p mimics inhibitor or their
corresponding controls were induced with osteogenic medium for 14
days in vitro. Then, 5×106 cells were loaded onto
20 mg hydroxyapatite-tricalcium phosphate (Sigma-Aldrich; Merck
KGaA) and subcutaneously implanted in the dorsal region of the
NOD/SCID mice. When the xenografts reached 50 mm3 in
volume, miR-877-3p agomir or antagomir or their corresponding
controls (5 nmol), respectively, was injected into the tumor every
other day. Four weeks later, the xenografts were removed, fixed
with 4% paraformaldehyde for 3 days and then decalcified in 10%
EDTA (pH 6.0) for 7 days. The xenografts were then embedded in
paraffin, sectioned (4 µm) and stained with hematoxylin and eosin
(H&E), or Masson's trichrome stain (Sigma-Aldrich; Merck KGaA)
according to the manufacturer's protocols. All animal procedures
were approved by the Animal Care Committee of Southern Medical
University (L2016113).
Western blotting
The cells were harvested and lysed with RIPA buffer
(Beyotime Institute of Biotechnology) containing phenylmethane
sulfonyl fluoride and protease inhibitors. Equal amounts of protein
(30 µg per lane) were loaded on a 10% sodium dodecyl
sulfate-polyacrylamide gel, separated by electrophoresis, and
transferred onto a 0.4-µm polyvinylidene difluoride membrane (EMD
Millipore, Billerica, MA, USA). The membranes were sealed with 5%
non-fat milk for 1 h at room temperature, then incubated with
primary antibodies overnight at 4°C. The primary antibodies were
anti-Smad7 (1:100; cat. no. sc-365846, Santa Cruz Biotechnology,
Ltd., Dallas, TX, USA), anti-OSX (1:200; cat. no. sc-393325, Santa
Cruz Biotechnology, Ltd.), anti-COL1A1 (1:1,000; cat. no. ab90395,
Abcam, Cambridge, UK), anti-RUNX2 (1:1,000; cat. no. 12556; Cell
Signaling Technology, Inc., Danvers, MA, USA), anti-phospho-Smad2
(1:1,000; cat. no. 18338; Cell Signaling Technology, Inc.),
anti-Smad2 (1:1,000; cat. no. 5339; Cell Signaling Technology,
Inc.), anti-phospho-Smad3 (1:1,000; cat. no. ab52903; Abcam,
Cambridge, UK), Smad3 (1:1,000; cat. no. 9523, Cell Signaling
Technology, Inc.) and anti-β-actin antibody (1:1,000; cat. no.
4970; Cell Signaling Technology, Inc.). The membranes were probed
with horseradish peroxidase-conjugated anti-rabbit secondary
antibody for 1 h at room temperature (1:5,000; cat. no. 7074; Cell
Signaling Technology, Inc.) and the protein signals were obtained
with ECL Plus substrate (EMD Millipore).
Luciferase reporter assay
The wild-type and mutant putative miR-877-3p binding
sites in the 3′UTR of Smad7 were synthesized and cloned into a
pmirGLO luciferase vector (Shanghai Genechem Co., Ltd., Shanghai,
China). The plasmids, miRNA (miR-877-3p and miR-NC) and Renilla
luciferase plasmid (Shanghai Genechem Co., Ltd.) were
co-transfected into cells using Lipofectamine 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.). Firefly and Renilla
luciferase activities were then measured using the Dual Luciferase
Reporter Assay system (Promega Corporation, Madison, WI, USA) at 48
h after transfection. All experiments were repeated three
times.
Bioinformatics target prediction
The candidate target genes of miR-877-3p were
predicted using TargetScan online software (http://www.targetscan.org/).
Statistical analysis
Data are expressed as mean ± standard deviation and
were analyzed using SPSS 22.0 software (IBM Corp., Armonk, NY,
USA). All experiments were repeated at least three times with
comparable results unless indicated otherwise. Statistical
evaluation of the data was performed using the unpaired Student's
t-test for comparisons between two groups and one-way analysis of
variance followed by Bonferroni correction post hoc test for
comparisons amongst multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
TGF-β1 promotes the osteoblastic
differentiation of MC3T3-E1 cells
To induce osteoblastic differentiation, the MC3T3-E1
cells were treated with TGF-β1. The mRNA expression levels of three
osteogenic genes, namely RUNX2, OSX and COL1A1, were then
determined in the MC3T3-E1 cells at 0, 7 and 14 days after TGF-β1
treatment. The expression levels of RUNX2, OSX and COL1A1 mRNA in
the MC3T3-E1 cells were increased at days 7 and 14 compared with
the respective levels at day 0, with the highest levels being
observed at day 14 (Fig. 1A).
Consistently, the corresponding protein levels in the MC3T3-E1
cells appeared to increase in a time-dependent manner (Fig. 1B). Furthermore, ALP activity
gradually increased during treatment with TGF-β1 (Fig. 1C), indicating the key role of TGF-β1
in osteoblastic differentiation.
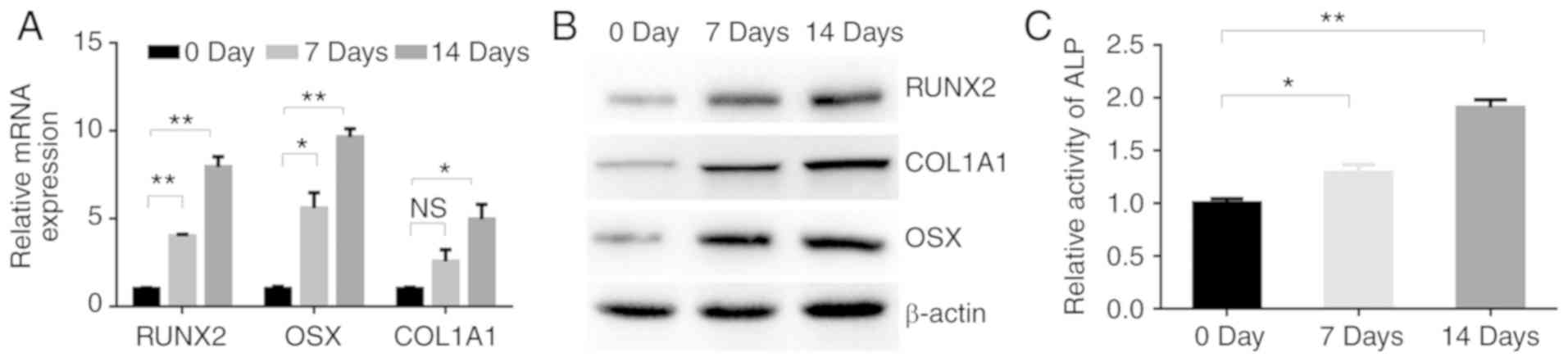 | Figure 1.TGF-β1 promotes the osteoblastic
differentiation of MC3T3-E1 cells. (A) RT-qPCR analysis of the
osteogenic-related genes RUNX2, OSX and COL1A1 in MC3T3-E1 cells
after treatment with TGF-β1 (4 ng/ml) for 0, 7 and 14 days. (B)
Western blotting of RUNX2, COL1A1-and OSX protein in MC3T3-E1 cells
cultured with TGF-β1 (4 ng/ml) for 0, 7 and 14 days. (C) ALP
activity in MC3T3-E1 cells after treatment with TGF-β1 (4 ng/ml)
for 0, 7 and 14 days. NS, no significance change; *P<0.05,
**P<0.01 as indicated. TGF, transforming growth factor; RUNX2,
runt-related transcription factor 2; OSX, osterix; COL1A1 collagen
type I α1 chain; ALP, alkaline phosphatase. |
Expression of miR-877-3p is
significantly altered during osteoblast induction and promotes the
osteoblastic differentiation of MC3T3-E1 cells
The role of miR-877-3p during osteoblast
differentiation is unclear. Therefore, the aim of the present study
was to investigate whether miR-877-3p is associated with osteogenic
differentiation. Following the treatment of MC3T3-E1 cells with
TGF-β1, the expression of miR-877-3p was detected using RT-qPCR
analysis, and a time-dependent increase in the level of miR-877-3p
was observed (Fig. 2A). To further
elucidate the functional role of miR-877-3p during osteoblastic
differentiation, miR-877-3p was overexpressed and knocked down in
MC3T3-E1 cells via transfection with miR-877-3p mimics and
miR-877-3p inhibitor, respectively, and corresponding controls were
established for comparison (Fig.
2B). The mRNA levels of the osteoblast-associated markers
RUNX2, OSX and COL1A1 were found to be significantly elevated in
miR-877-3p-overexpressing MC3T3-E1 cells, and significantly reduced
in cells with miR-877-3p knockdown (Fig.
2C). Consistently, ARS and ALP staining results demonstrated
that the staining intensity of mineralization nodes was markedly
increased in the miR-877-3p-overexpressing cells, and decreased in
the miR-877-3p-depleted cells (Fig.
2D). The effect of miR-877-3p on MC3T3-E1 osteogenesis was
further evaluated in vivo. The H&E and Masson's
trichrome staining results revealed that osteoid formation was
increased in the xenografts of mice treated with miR-877-3p agomir,
and decreased in those of the mice treated with miR-877-3p
antagomir (Fig. 2E). RT-qPCR
analysis confirmed that the expression of miR-877-3p was elevated
in the agomir-treated group and reduced in antagomir-treated group,
compared with the respective controls (Fig. 2F). These results demonstrate that
miR-877-3p promoted the osteogenic differentiation of MC3T3-E1
cells.
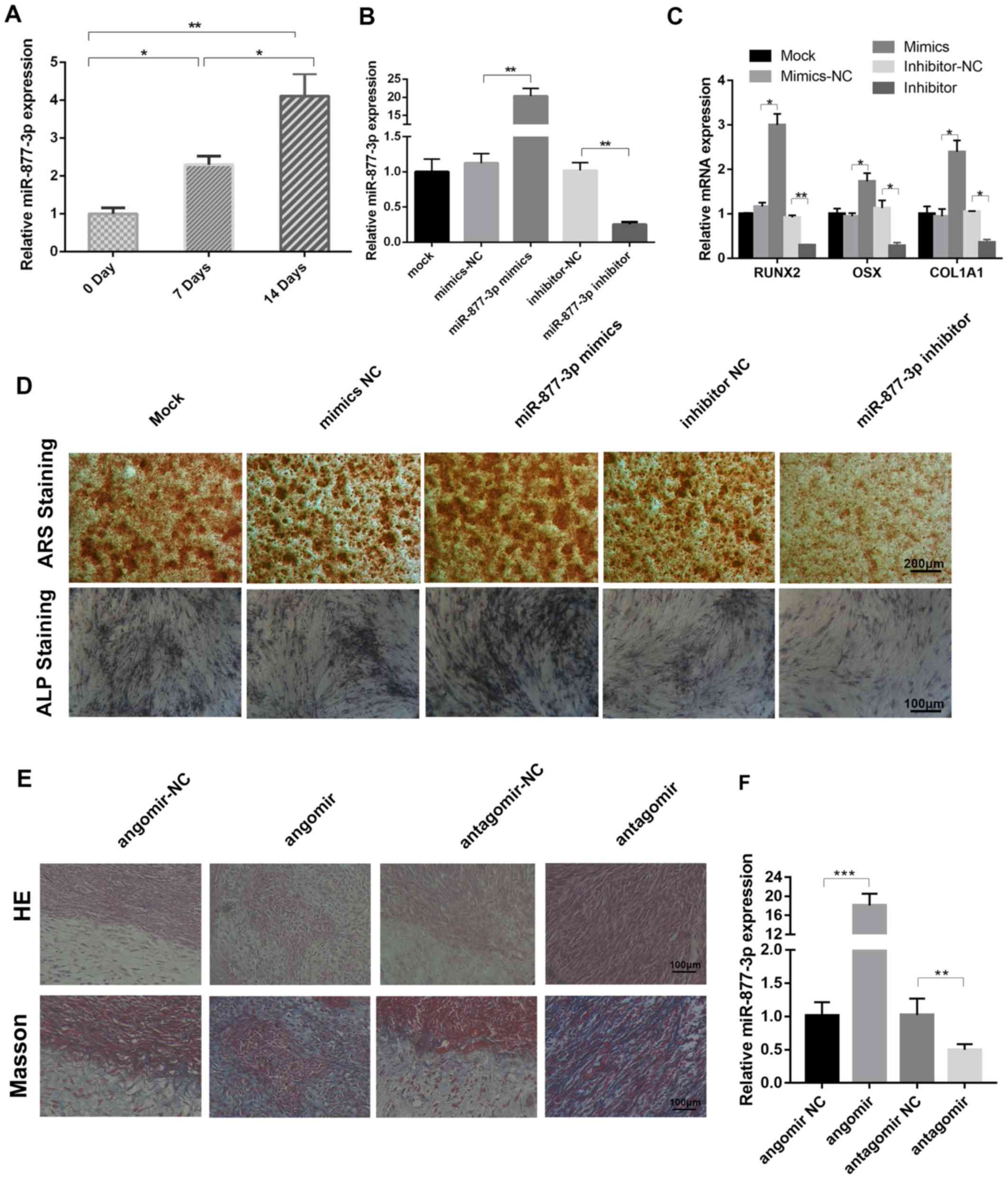 | Figure 2.miR-877-3p promotes the osteoblastic
differentiation of MC3T3-E1 cells. (A) miR-877-3p expression in
MC3T3-E1 cells after treatment with TGF-β1 (4 ng/ml) for 0, 7 and
14 days. (B) Expression of miR-877-3p was validated in MC3T3-E1
cells transfected with miR-877-3p mimics or inhibitor and their
corresponding controls by RT-qPCR. (C) RUNX2, OSX and COL1A1 mRNA
expression in MC3T3-E1 cells. (D) MC3T3-E1 cells transfected with
miR-877-3p mimics or inhibitor and their corresponding controls
were cultured in osteogenic medium. At day 14, the mineralization
of differentiated MC3T3-E1 cells was detected with ARS and ALP
staining. (E) Transfected MC3T3-E1 cells were cultured in
osteogenic medium for 14 days and then mixed with tricalcium
phosphate/hydroxyapatite and transplanted into the dorsal region of
nude mice for 4-weeks. The results were then evaluated by H&E
and Masson's trichrome staining. (F) (F) Transfected MC3T3-E1 cells
were cultured in osteogenic medium for 14 days and then mixed with
tricalcium phosphate/hydroxyapatite and transplanted into the
dorsal region of nude mice for 4-weeks. The Expression expression
of miR-877-3p in the xenografts from each group was detected by
RT-qPCR analysis. *P<0.05, **P<0.01 and ***P<0.001, as
indicated. miR, microRNA; TGF, transforming growth factor; RUNX2,
runt-related transcription factor 2; OSX, osterix; COL1A1 collagen
type I α1 chain; NC, negative control; ARS, Alizarin Red S; ALP,
alkaline phosphatase; H&E, hematoxylin and eosin. |
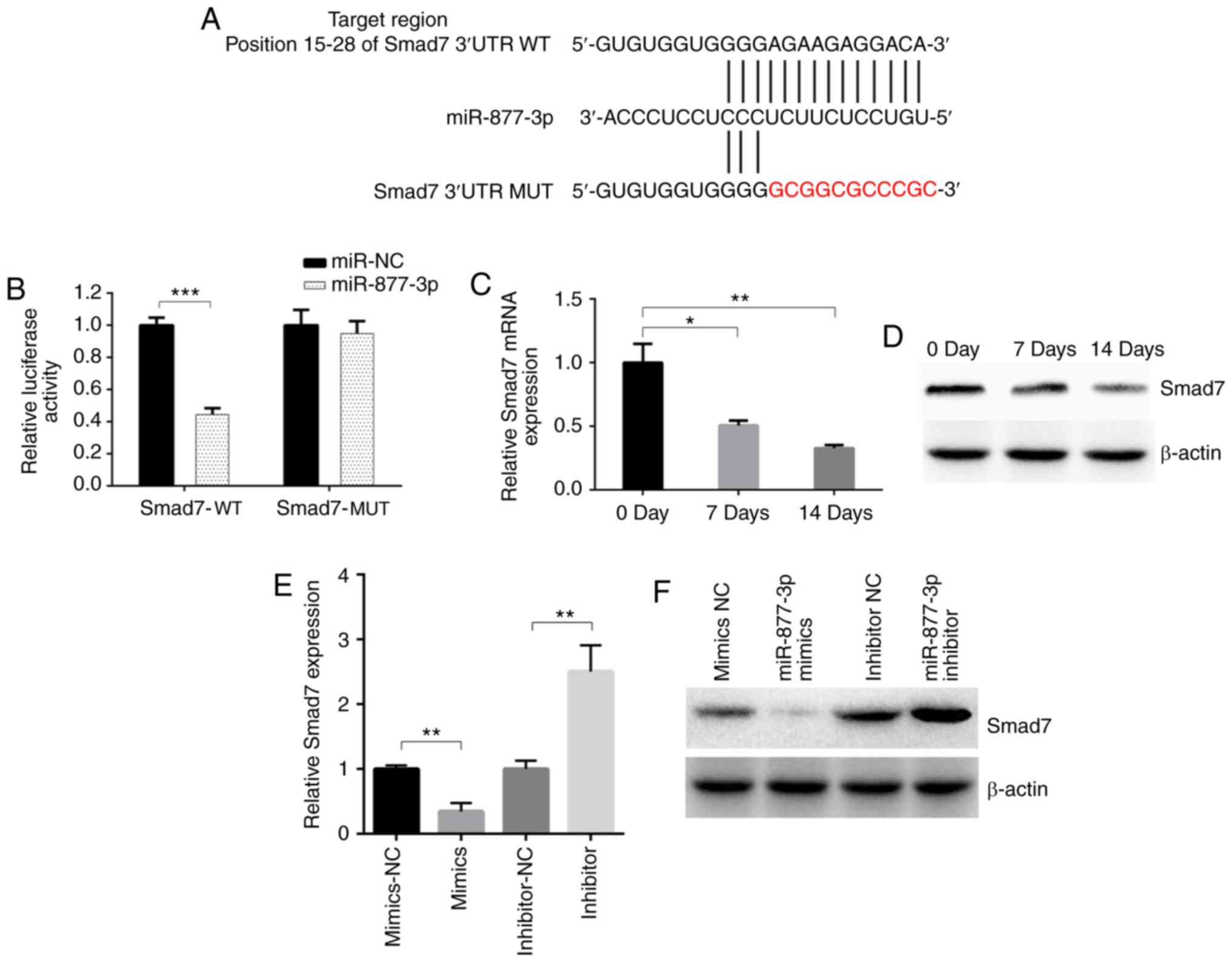
MiR-877-3p inhibits the expression of
Smad7
miR-877-3p has been demonstrated to promote the
TGF-β1-induced osteoblastic differentiation of MC3T3-E1 cells;
however, its downstream target has not yet been determined. Using
the online bioinformatics tool TargetScan, Smad7 was predicted to
be a downstream target of miR-877-3p with a potential binding site
(Fig. 3A). Fragments of the 3′UTR of
Smad7 containing a putative miR-877-5p binding site or mutant
fragment were synthesized and inserted into plasmids. The predicted
binding site of miR-877-3p in Smad7 was confirmed using a
luciferase reporter assay (Fig. 3B).
The mRNA and protein levels of Smad7 were decreased after TGF-β1
induction (Fig. 3C and D). Whether
miR-877-3p regulated osteogenic differentiation by suppressing
Smad7 expression was then investigated. The results revealed a
notable downregulation of Smad7 expression following the
transfection of miR-877-3p mimic into MC3T3-E1 cells, whereas Smad7
was upregulated in cells transfected with miR-877-3p inhibitor, as
confirmed by RT-qPCR and western blot assays (Fig. 3E and F). These results support the
hypothesis that miR-877-3p directly binds to Smad7.
Smad7 may be a negative target of
miR-877-3p involved in osteoblast differentiation
Although Smad7 expression has been found to be
affected by miR-877-3p, the interaction between Smad7 and
miR-877-3p required further evaluation. To confirm the role of the
miR-877-3p/Smad7 axis in the regulation of osteoblastic
differentiation, MC3T3-E1 cells transfected with miR-877-3p
inhibitor or inhibitor-NC were cultured with or without TGF-β1 for
7 days. Compared with the control cells transfected with
inhibitor-NC, the expression of Smad7 was upregulated when cells
were transfected with miR-877-3p inhibitor alone, while TGF-β1
reversed the expression of Smad7 induced by the miR-877-3p
inhibitor, as confirmed by RT-qPCR (Fig.
4A). By contrast, the osteogenesis-associated genes RUNX2, OSX
and COL1A1 exhibited the opposite tendency (Fig. 4B). The protein levels of RUNX2 and
Smad7 were also verified by western blotting, and were consistent
with the mRNA results. Western blotting also revealed that the
levels of phospho-Smad2 and phospho-Smad3 were decreased by the
miR-877-3p inhibitor, and this effect was reversed by TGF-β1
(Fig. 4C). Furthermore,
Smad7-overexpressing and control MC3T3-E1 cells were established by
transfection with a pcDNA3.1-Smad7 plasmid (pcDNA-Smad7) and
control plasmid (pcDNA-NC), and then co-transfected with miR-877-3p
mimics or mimics-NC, and the success of transfection was verified
using RT-qPCR (Fig. 4D and E). The
promoting effects of miR-877-3p mimics on osteogenesis-related
genes, phospho-Smad2 and phospho-Smad3 were markedly inhibited by
pcDNA-Smad7 (Fig. 4F and G). ALP
activity, ARS and ALP staining exhibited the same tendency
(Fig. 4H and I). Collectively, these
results confirmed that Smad7 inhibits the osteoblastic
differentiation of MC3T3-E1 cells and is able to reverse the
osteoblastic differentiation mediated by miR-877-3p.
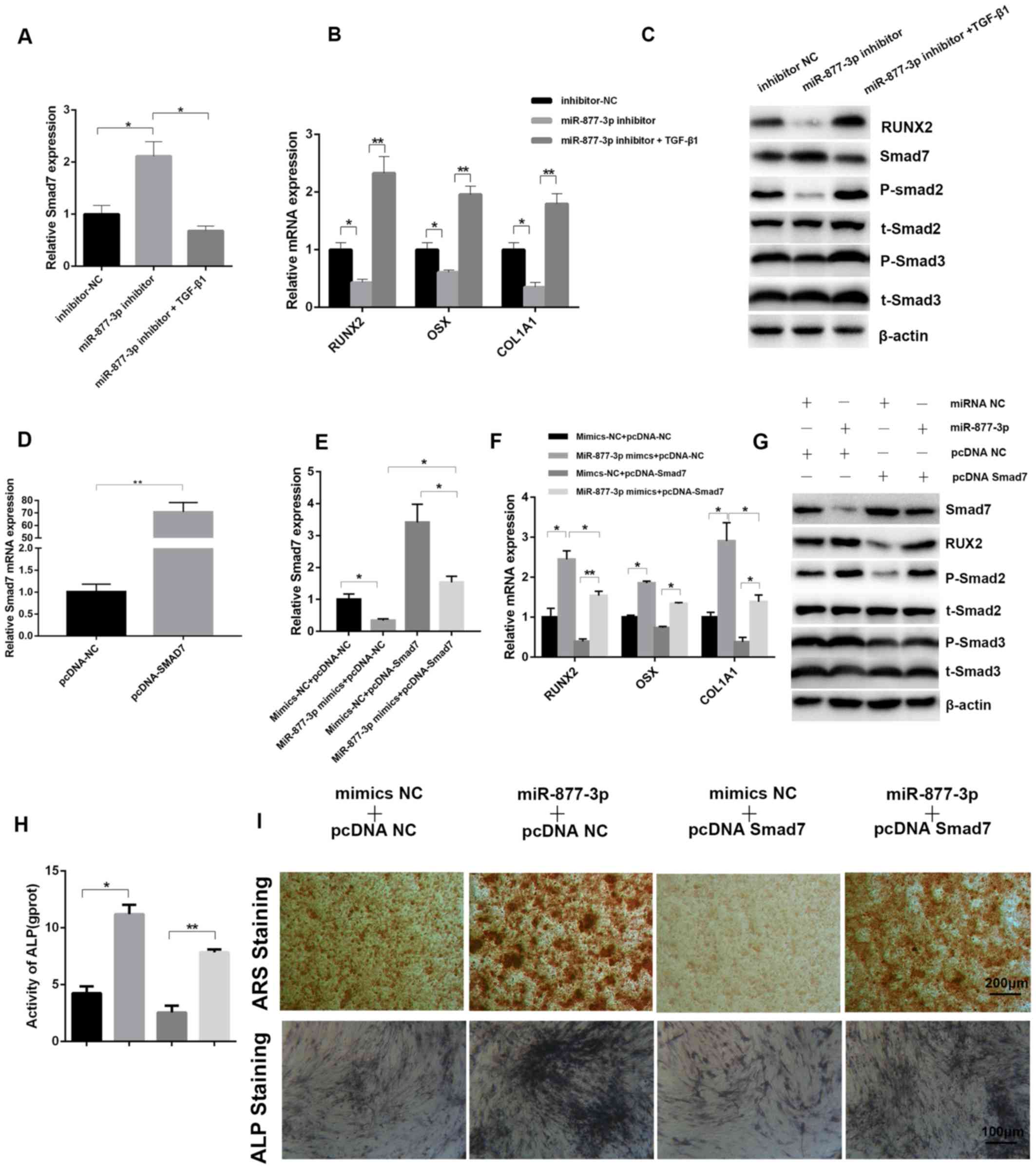 | Figure 4.Smad7 is identified as a downstream
target of miR-877-3p during the osteoblastic differentiation of
MC3T3-E1 cells. mRNA expression of (A) Smad7 and (B) osteoblast
differentiation-associated genes in MC3T3-E1 cells transfected with
inhibitor-NC, miR-877-3p inhibitor or miR-877-3p inhibitor plus
TGF-β1. (C) Expression of RUNX2, Smad7, p-Smad2, t-Smad2, p-Smad3
and t-Smad3 protein in MC3T3-E1 cells transfected with
inhibitor-NC, miR-877-3p inhibitor or miR-877-3p inhibitor plus
TGF-β1. (D) The mRNA expression of Smad7 was validated in MC3T3-E1
cells transfected with pcDNA-NC or pcDNA-Smad7. The mRNA expression
of (E) Smad7 and (F) osteoblast differentiation-associated genes in
MC3T3-E1 cells transfected with miR-877-3p mimics and/or
pcDNA-Smad7. (G) Protein levels of Smad7, RUNX2, p-Smad2, t-Smad2,
p-Smad3 and t-Smad3 in MC3T3-E1 cells transfected with miR-877-3p
mimics and/or pcDNA-Smad7. (H) ALP activity of MC3T3-E1 cells
transfected with miR-877-3p mimics and/or pcDNA-Smad7 after culture
with osteogenic medium for 7 days. (I) MC3T3-E1 cells transfected
with miR-877-3p mimics and/or pcDNA-Smad7 were induced with
osteogenic medium. At day 14, the mineralization of differentiated
MC3T3-E1 cells was detected with ARS and ALP staining. *P<0.05,
**P<0.01 as indicated. miR, microRNA; NC, negative control; TGF,
transforming growth factor; RUNX2, runt-related transcription
factor 2; p, phospho; t, total; ALP, alkaline phosphatase; ARS,
Alizarin Red S. |
Discussion
Bone formation and mineralization are triggered by
the differentiation of mesenchymal precursor cells into
osteoblasts. Osteoblast differentiation is significantly affected
by a key transcriptional factor, RUNX2, that regulates the
expression of several osteogenic genes, including ALP, OSX and
COL1A1 (17–19). These osteogenic genes are associated
with the expression of bone matrix genes such as type 1 collagen,
osteopontin, osteocalcin and bone sialoprotein (20–23). In
addition, several signaling pathways, including Wnt/β-catenin,
BMP/TGF-β, JAK/STAT and MAPK, have been reported to promote
osteoblast differentiation in vivo and in vitro
(23–25). TGF-β1 is a polypeptide member of the
TGF-β superfamily of cytokines that regulate several cellular
functions, including cell proliferation, survival and
differentiation (26–28). The combined actions of these
biological processes rely on the effect of TGF-β1 on bone formation
(29).
MC3T3-E1 cells, which are mouse pre-osteoblast cells
considered to be good models for the study of in vitro
osteoblast differentiation (14),
were used in the present study. These cells were treated with
TGF-β1 to induce osteoblastic differentiation, which was evaluated
by the expression of the aforementioned osteoblast gene markers. It
must be acknowledged that a limitation of the present study is that
only one cell line was investigated; different cell lines, such as
primary osteoblasts, should be included in further studies.
However, the experiments in the present study were, to the best of
our knowledge, the first to provide evidence that miR-877-3p is
upregulated during the osteoblastic differentiation of MC3T3-E1
cells in a time-dependent manner. Consistent with this observation,
miR-877-3p has also been found to be highly upregulated during
myofibroblast differentiation (12).
Subsequently, the effect of miR-877-3p on TGF-β1-induced osteoblast
differentiation and mineralization was investigated, by
overexpression and knockdown experiments performed in the
osteoblast cell line. The results clearly demonstrated that
miR-877-3p promotes osteoblast differentiation.
miRNAs have been identified as post-transcriptional
inhibitors that exert their biological effects via the
downregulation of target gene expression. Bioinformatics analysis
in the present study identified Smad7 as a putative target of
miR-877-3p. By transient transfection with miR-877-3p mimics or
inhibitor, it was demonstrated that miR-877-3p suppressed Smad7
mRNA and protein expression in MC3T3-E1 cells. Smad7 is a
well-defined negative regulator of the TGF-β1 signaling pathway and
bone metabolism (30). As regards to
bone formation initiated by osteoblasts, decreasing expression of
Smad7 during osteoblast differentiation of MC3T3-E1 cells was
observed in the present study, which is consistent with previously
reported findings (31). The present
study also indicated that the expression of Smad7 increased in
MC3T3-E1 cells with miR-877-3p downregulation, while this effect
was eliminated in the presence of TGF-β1. Therefore, it was
hypothesized that Smad7 is a negative target of miR-877-3p and
TGF-β1, and that one of the important mechanisms by which
miR-877-3p promotes the osteoblastic differentiation induced by
TGF-β1 may be through suppressing the expression of the Smad7 gene.
This hypothesis was confirmed in the co-transfection experiment.
The results suggested that overexpression of Smad7 partially
inhibited the miR-877-3p-mediated osteoblastic differentiation of
MC3T3-E1 cells. However, evidence is required to demonstrate that
microRNAs-877-3p regulate osteoblast differentiation by interacting
with Smad7. In addition, the role of miRNAs in osteoblast
differentiation is complex; the bypass or crosstalk of
microRNAs-877-3p in this process was not excluded, and requires
further study.
In summary, the results of the present study
revealed the mechanism underlying the regulatory effect of
miR-877-3p on the osteoblastic differentiation of MC3T3-E1 cells.
Through this mechanism, miR-877-3p promoted osteoblastic
differentiation of MC3T3-E1 cells induced by TGF-β1 via silencing
Smad7. These findings provided novel insights into the effects of
miR-877-3p on osteoblast differentiation and bone formation, which
may provide a new therapeutic approach for degenerative bone
diseases.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
DH conceived and designed the experiments, conducted
the data analysis, contributed reagents/materials/tools and wrote
the manuscript. GH and JC performed the experiments. All authors
read and approved the final manuscript. All authors contributed to
the data analysis, drafting and critical revision of the paper, and
agree to be accountable for all aspects of the work.
Ethics approval and consent to
participate
All animal procedures were approved by the Animal
Care Committee of Southern Medical University (L2016113).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Takayanagi H: Osteoimmunology: Shared
mechanisms and crosstalk between the immune and bone systems. Nat
Rev Immunol. 7:292–304. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Manolagas SC and Jilka RL: Bone marrow,
cytokines, and bone remodeling. Emerging insights into the
pathophysiology of osteoporosis. N Engl J Med. 332:305–311. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bernabeu C, Lopez-Novoa JM and Quintanilla
M: The emerging role of TGF-beta superfamily coreceptors in cancer.
Biochim Biophys Acta. 1792:954–973. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Crane JL, Xian L and Cao X: Role of TGF-β
signaling in coupling bone remodeling. Methods Mol Biol.
1344:287–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yasui T, Kadono Y, Nakamura M, Oshima Y,
Matsumoto T, Masuda H, Hirose J, Omata Y, Yasuda H, Imamura T, et
al: Regulation of RANKL-induced osteoclastogenesis by TGF-β through
molecular interaction between Smad3 and Traf6. J Bone Miner Res.
26:1447–1456. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang J, Zhao L, Xing L and Chen D:
MicroRNA-204 regulates Runx2 protein expression and mesenchymal
progenitor cell differentiation. Stem Cells. 28:357–364.
2010.PubMed/NCBI
|
|
8
|
Xiang LY, Ou HH, Liu XC, Chen ZJ, Li XH,
Huang Y and Yang DH: Loss of tumor suppressor miR-126 contributes
to the development of hepatitis B virus-related hepatocellular
carcinoma metastasis through the upregulation of ADAM9. Tumour
Biol. 39:10104283177091282017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cao M, Hou D, Liang H, Gong F, Wang Y, Yan
X, Jiang X, Wang C, Zhang J, Zen K, et al: miR-150 promotes the
proliferation and migration of lung cancer cells by targeting SRC
kinase signalling inhibitor 1. Eur J Cancer. 50:1013–1024. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen JF, Mandel EM, Thomson JM, Wu Q,
Callis TE, Hammond SM, Conlon FL and Wang DZ: The role of
microRNA-1 and microRNA-133 in skeletal muscle proliferation and
differentiation. Nat Genet. 38:228–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Taipaleenmäki H, Bjerre Hokland L, Chen L,
Kauppinen S and Kassem M: Mechanisms in endocrinology: micro-RNAs:
Targets for enhancing osteoblast differentiation and bone
formation. Eur J Endocrinol. 166:359–371. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang C, Gu S, Cao H, Li Z, Xiang Z, Hu K
and Han X: miR-877-3p targets Smad7 and is associated with
myofibroblast differentiation and bleomycin-induced lung fibrosis.
Sci Rep. 6:301222016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li S, Zhu Y, Liang Z, Wang X, Meng S, Xu
X, Xu X, Wu J, Ji A, Hu Z, et al: Up-regulation of p16 by
miR-877-3p inhibits proliferation of bladder cancer. Oncotarget.
7:51773–51783. 2016.PubMed/NCBI
|
|
14
|
Niu YB, Kong XH, Li YH, Fan L, Pan YL, Li
CR, Wu XL, Lu TL and Mei QB: Radix Dipsaci total saponins stimulate
MC3T3-E1 cell differentiation via the bone morphogenetic
protein-2/MAPK/Smad-dependent Runx2 pathway. Mol Med Rep.
11:4468–4472. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang L, Wu F, Song Y, Li X, Wu Q, Duan Y
and Jin Z: Long noncoding RNA related to periodontitis interacts
with miR-182 to upregulate osteogenic differentiation in
periodontal mesenchymal stem cells of periodontitis patients. Cell
Death Dis. 7:e23272016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dacic S, Kalajzic I, Visnjic D, Lichtler
AC and Rowe DW: Col1a1-driven transgenic markers of osteoblast
lineage progression. J Bone Miner Res. 16:1228–1236. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nakashima K, Zhou X, Kunkel G, Zhang Z,
Deng JM, Behringer RR and de Crombrugghe B: The novel zinc
finger-containing transcription factor osterix is required for
osteoblast differentiation and bone formation. Cell. 108:17–29.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pratap J, Galindo M, Zaidi SK, Vradii D,
Bhat BM, Robinson JA, Choi JY, Komori T, Stein JL, Lian JB, et al:
Cell growth regulatory role of Runx2 during proliferative expansion
of preosteoblasts. Cancer Res. 63:5357–5362. 2003.PubMed/NCBI
|
|
20
|
Baek WY and Kim JE: Transcriptional
regulation of bone formation. Front Biosci (Schol Ed). 3:126–135.
2011.PubMed/NCBI
|
|
21
|
Bruderer M, Richards RG, Alini M and
Stoddart MJ: Role and regulation of RUNX2 in osteogenesis. Eur Cell
Mater. 28:269–286. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ohata Y and Ozono K: Bone and stem cells.
The mechanism of osteogenic differentiation from mesenchymal stem
cell. Clin Calcium. 24:501–508. 2014.(In Japanese). PubMed/NCBI
|
|
23
|
Bodine PV and Komm BS: Wnt signaling and
osteoblastogenesis. Rev Endocr Metab Disord. 7:33–39. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Levy JB, Schindler C, Raz R, Levy DE,
Baron R and Horowitz MC: Activation of the JAK-STAT signal
transduction pathway by oncostatin-M cultured human and mouse
osteoblastic cells. Endocrinology. 137:1159–1165. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Beederman M, Lamplot JD, Nan G, Wang J,
Liu X, Yin L, Li R, Shui W, Zhang H, Kim SH, et al: BMP signaling
in mesenchymal stem cell differentiation and bone formation. J
Biomed Sci Eng. 6:32–52. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Massagué J, Blain SW and Lo RS: TGFbeta
signaling in growth control, cancer, and heritable disorders. Cell.
103:295–309. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park JS, Chu JS, Tsou AD, Diop R, Tang Z,
Wang A and Li S: The effect of matrix stiffness on the
differentiation of mesenchymal stem cells in response to TGF-β.
Biomaterials. 32:3921–3930. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Siegel PM and Massagué J: Cytostatic and
apoptotic actions of TGF-beta in homeostasis and cancer. Nat Rev
Cancer. 3:807–821. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Janssens K, ten Dijke P, Janssens S and
Van Hul W: Transforming growth factor-beta1 to the bone. Endocr
Rev. 26:743–774. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang F, Ren CC, Liu L, Chen YN, Yang L
and Zhang XA: HOXC6 gene silencing inhibits epithelial-mesenchymal
transition and cell viability through the TGF-β/smad signaling
pathway in cervical carcinoma cell. Cancer Cell Int. 18:2042018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yano M, Inoue Y, Tobimatsu T, Hendy G,
Canaff L, Sugimoto T, Seino S and Kaji H: Smad7 inhibits
differentiation and mineralization of mouse osteoblastic cells.
Endocr J. 59:653–662. 2012. View Article : Google Scholar : PubMed/NCBI
|