Introduction
Platelets primarily participate in hemostasis and
antimicrobial host defense, however they also secrete cytokines,
which induce inflammation and trigger tissue repair (1,2). It is
common knowledge that platelets serve critical roles in the
pathology of cardiovascular disorders, chronic inflammatory and
atherothrombotic diseases (3). At
injury sites platelets typically aggregate on vessel walls and
prevent bleeding (4). Infections are
associated with either the sudden increase or decrease in platelets
(known as thrombocytosis and thrombocytopenia, respectively), which
are also the biomarkers for disease progression or tissue healing
(1). In a healthy individual
platelets range between 150,000 and 400,000/µl of blood and there
is a continual balance between platelet clearance and platelet
production (1,5). This balance must be carefully
maintained to avoid arterial occlusion, spontaneous bleeding and
organ damage. However, understanding of this process is limited as
has been poorly investigated.
Previous studies have reported a number of
anti-coagulation drugs, including sodium citrate, heparin, vitamin
K antagonist and nattokinase, all of which have been extensively
utilized within a clinical setting (6–8).
Physiological hemostasis has not been widely investigated in the
past, however, hemostasis is important for the clearance of
thrombus (9,10). Platelets serve a critical role in the
process of physiological hemostasis, however their specific
mechanisms and targets have not been fully clarified (9–11). The
authors of the present study speculated that the identification of
physiological hemostasis targets may be important for the
prevention of thrombogenesis. Human platelets express a series of
molecules, including immunoglobulin G immune complex receptor,
glycoprotein VI (GPVI), Fc-γ-RIIa and C-type lectin-like receptor
(12–14). The activation of these molecules
contributes to physiological hemostasis or thrombosis (15).
The leucine-rich repeat flightless-interacting
protein-1 (LRRFIP1) gene has previously been identified and
revealed to serve a key role in the regulation of gene
transcription (16). LRRFIP1 was
originally derived as a GC-rich binding-protein that repressed the
expression of platelet-derived growth factors (17). Silencing of the LRRFIP1 gene caused a
notable decrease in thrombus formation and was positively
correlated with levels of αIIbβ3 (18,19).
Therefore, it was hypothesized that the LRRFIP1 gene may serve an
important role in thrombus formation. The present study aimed to
investigate the effects of LRRFIP1 on platelet agglutination.
Materials and methods
Bacterial strain and preparation of
LRRFIP1 recombinant protein
The BL21 (DE3) Escherichia coli strain (cat.
no. GSB013; ZonHon Biopharma Institute Inc., Changzhou, China) was
cultured in lysogeny broth (LB) liquid medium (cat. no. L3152;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Full-length
recombinant LRRFIP1 protein was purified from the total crude
extract of BL21 as previously described (20). Briefly, the BL21 strain was cultured
in the LB culture medium and then re-suspended in lysis liquid
(cat. no. T9424; Sigma-Aldrich; Merck KGaA) to obtain total
protein, which was purified using glutathione-S-transferase soluble
protein (Sangon Biotech Co., Ltd., Shanghai, China). The
prokaryotic-expressed proteins were extracted using glutathione
sepharose 4B beads (GE Healthcare, Chicago, IL, USA) according to
the manufacturer's protocol. The highly purified LRRFIP1 was
identified as high-density bands on the SDS-PAGE images. Briefly,
the concentration of the obtained recombinant LRRFIP1 was
determined using a bicinchoninic acid protein assay kit according
to the manufacturer's protocol. A total of 0.2 µg protein lysates
were separated with SDS-PAGE on a 15% gel.
Animals
A total of 20 BALB/C mice with 6–8 week-old (10 male
and 10 female), weighting from 25 to 35 g were purchased from
Beijing HFK Bioscience Co., Ltd. (Beijing, China). All mice used in
the present study were housed in cages (5 mice/cage) under the same
conditions, including a controlled environment at 22°C with 50%
humidity and a 12 h light/dark cycle. The food, water and bedding
were sterilized, and the mice had free access to food and water.
All animal experiments were approved by the Ethics Committee of
Daping Hospital (Chongqing, China) and all mice were handled in
accordance with the Guidelines for Care and Use of Laboratory
Animalsby the National Institute of Health (21).
Establishment of plasmids and a mouse
model of LRRFIP1 gene knockout
To create LRRFIP1 expression plasmids the targets of
the LRRFIP1 gene were designed as detailed in Fig. 1A. Embryonic stem cells (cat. no.
CRL-11379; American Type Culture Collection, Manassas, VA, USA)
were used to clone the LRRFIP1 gene according to the previously
published studies (16,17). Several strains of the LRRFIP1 gene
are listed in Fig. 1B, andexon 2 was
the most conservative. Therefore, the LRRFIP1 gene knockout mice
were established by synthesizing the mutated exon 2 gene. Among the
listed LRRFIP1 genes, three target gene sequences were selected and
cloned into the px458 plasmid (cat. no. 3683466; BioVector NTCC
Inc., Beijing, China) to construct LRRFIP1−/− expression
plasmids by employing the BamH I and EcoR I restriction enzymes
(Fig. 1C); these plasmids were used
to develop the LRRFIP1 knockout mouse model. Target gene 3
illustrated the highest-density band (Fig. 1D), which was selected to establish
the LRRFIP1−/− expression plasmid according to previous
studies (16,17).
To establish a mouse model of LRRFIP1 gene knockout,
LIRRFIP1 genes were synthesized by Western Biotech, Inc.
(Chongqing, China). The metaphase II oocytes of fertilized embryos
were cultured in M2 medium (cat. no. M8410) for 24 h at 37°C and
treated with acidic tyrode's solution (cat. no. P4417; both
Sigma-Aldrich; Merck KGaA) for 60 sec at 37°C. The cells were
subsequently cultured with Opti-minimal essential medium (MEM; cat.
no. 31985-070; Life Technologies; Thermo Fisher Scientific, Inc.)
for 24 h at 37°C. For Cas9 plasmid (cat. no. 44758; Addgene, Inc.,
Cambridge, MA, USA) transfection, 500 ng purified Cas9 plasmid was
added to 25 µl Opti-MEM, followed by the addition of 100 ng guide
RNA (gRNA; GenScript Corporation, Piscataway, NJ, USA). A total of
2 µl Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) was diluted into 25 µl opti-MEM and mixed with
the Cas9 plasmid/gRNA sample. The mixture was incubated at 37°C for
10 min prior to the additionof the fertilized embryos. The ovigerm
stage fertilized embryos were subsequently transplanted into the
mice by injecting the embryos into the uterus in a specific
pathogenic and virus antigen free environment. The mice were
monitored for breathing problems, marasmus, roughened hair and
fever to ensure that the specific pathogenic and virus antigen free
environment was maintained. DNA was extracted from the blood of the
BALB/C mice using the commercial RNA extraction kit (cat. no.
DP432; Tiangen Biotech Co., Ltd., Beijing, China) and the first
strand cDNA synthesis kit (cat. no. K1612; Thermo Fisher
Scientific, Inc.). The PCR assay was conducted using the SYRB PCR
system (cat. no. DRR820A; Takara Biotechnology Co., Ltd., Dalian,
China) to confirm that the mice harbored the desired LRRFIP1
mutation. The primers were as follows: forward,
5-CTAAGCCGGGCACAGTAACA-3′ and reverse, 5′-TAAAGGGCAAGCTCTCAGGC-3′.
Amplification conditions for PCR were as follows: 94°C for 4 min,
followed by 35 cycles of 95°C for 20 sec, 60°C for 30 sec and 72°C
for 30 sec, and terminated at 72°C for 10 min.
The following humane endpoint was established
according to a previous study to ensure that the welfare of the
mice was protected (22): A body
weight loss of >15% compared with the weight at the time of
experiment onset, accompanied by hunched posture, lethargy and poor
grooming. A total of 10% (2/20) of the mice were euthanized as a
result of reaching the established humane endpoint. This percentage
of mortality was expected in the present study based on
prior-experiments by our group. At 1 day following the end of the
study, all remaining mice were sacrificed via150 mg/kg body weight
intraperitoneal injection of thiopental (Altana AG, Wesel,
Germany).
Platelet preparation and platelet
agglutination assay
The platelet agglutination assay was performed and
evaluated as previously described with several modifications
(18). Briefly, 1 week after the
establishment of the model, whole blood (0.1 ml) was collected from
the caudal vein and added into glass evacuated tubes containing 1.5
ml 3.28% poncirin acid sodium salt (BD Biosciences, Franklin Lakes,
NJ, USA). The platelet-rich plasma (PRP) was obtained from whole
blood by centrifuging at 1,000 × g for 10 min at room temperature.
The PRP was used immediately or was stored without agitation at 4°C
for 48 h. The PRP was centrifuged at 3,000 × g for 10 min at room
temperature to form a pellet of platelets. The platelet poor plasma
was decanted and the platelets were suspended in 2 ml
HEPES/Tyrode's buffer (Gibco; Thermo Fisher Scientific, Inc.)
without Ca2+ or Mg2+. Platelet agglutination
rate maximum [PAG (M)] and PAG at 1 min [PAG (1)] were examined and an agglutination assay
was conducted using a platelet aggregation apparatus (Order no.
H79194; Shanghai Huanxi Medical Devices Co., Ltd., Shanghai, China)
according to the manufacturer's protocol. Additionally, a type of
agglutination index (I%) was evaluated as shown in a previous study
(5).
Trial groupings
All treatments were added directly to the platelets.
For the collagen protein-induced platelet agglutination assay the
following groups were used: i) Platelet control group, which did
not receive treatment; ii) collagen (0.07 µg/ml) and low-molecular
heparin (LMH; 1.2 IU/ml; both Sigma-Aldrich; Merck KGaA) treated
platelet group (P+C+LMH); iii) collagen (0.07 µg/ml), LMH (1.2
IU/ml) and wild-type LRRFIP1 (wLRRFIP1; 50 nM) treated platelet
group (P+C+LMH+wLRRFIP150); and iv) collagen (0.07 µg/ml), LMH (1.2
IU/ml) and wLRRFIP1 (100 nM) treated platelet group
(P+C+LMH+wLRRFIP1100). A total of 24 h after the platelet
treatment, the agglutination assay was conducted.
For the thrombin receptor-mediated platelet
agglutination assay the following groups were used: i) Thrombin
receptor activator (SFLLRN; 2 µM; cat. no. S1820; Sigma-Aldrich;
Merck KGaA) treated platelet group (P+SFLLRN); ii) LMH (1.2 IU/ml)
and SFLLRN (2 µM) treated platelet group (P+LMH+SFLLRN); iii)
wLRRFIP1 (100 nM) and SFLLRN (2 µM) treated platelet group
(P+wLRRFIP1100+SFLLRN); and iv) wLRRFIP1 (200 nM) and SFLLRN (2 µM)
treated platelet group (P+wLRRFIP1200+SFLLRN).
For the monoclonal antibody regulated platelet
agglutination assay, the following groups were used (n=4/group): i)
Platelet control group; ii) collagen (0.07 µg/ml) treated platelet
group (P+C); iii) collagen (0.07 µg/ml) and anti-wLRRFIP1 antibody
(30 µg/ml; cat. no. sc-515571; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) treated platelet group (P+C+anti-wLRRFIP1); iv)
SFLLRN (2 µM) treated platelet group (P+SFLLRN); and v) SFLLRN (2
µM) and anti-wLRRFIP1 antibody (30 µg/ml) treated platelet group
(P+SFLLRN+anti-wLRRFIP1).
For the LRRFIP1 gene knockout associated platelet
agglutination assay the following groups were used: i) Platelets in
wLRRFIP1+/+ mice group (P+LRRFIP1+/+); ii)
platelets in wLRRFIP1−/− mice group
(P+LRRFIP1−/−); iii) collagen (0.07 µg/ml) treated
platelets in wLRRFIP1+/+ mice group
(P+C+LRRFIP1+/+); iv) collagen (0.07 µg/ml) treated
platelets in wLRRFIP1−/− mice group
(P+C+LRRFIP1−/−); v) SFLLRN (2 µM) treated platelets in
wLRRFIP1+/+ mice group (P+SFLLRN+LRRFIP1+/+);
vi) SFLLRN (2 µM) treated platelets in wLRRFIP1−/− mice
group (P+SFLLRN+LRRFIP1−/−).
Flow cytometry for αIIbβ3
examination
αIIbβ3 was examined in the LRRFIP1+/+,
LRRFIP1−/−, LMH, LRRFIP1+/+ + Convulxin and
LRRFIP1−/− + Convulxin groups. Convulxin (0.2 nmol/l;
Sigma-Aldrich; Merck KGaA) was added to the platelets. Then active
αIIbβ3 integrin was examined by flow cytometry as previously
described (23). Briefly, the
activated integrin αIIbβ3 was quantified by binding to the
FITC-labeled rabbit anti-mouse αIIbβ3 monoclonal antibodies
(1:1,000; cat. no. 340507; BD Biosciences). A BD FACSVantage flow
cytometer (BD Biosciences) was used to quantify the data. CELIQUEST
software (version 5.1; BD Biosciences) was used to analysis the
flow cytometry data.
Statistical analysis
All data in the present study are presented as the
mean ± standard deviation and were analyzed using SPSS 20.0
software (IBM Corp., Armonk, NY, USA). The data was obtained from a
minimum of three independent experiments. One-way analysis of
variance was used to compare multiple groups followed by Tukey's
Honest Significant Difference post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
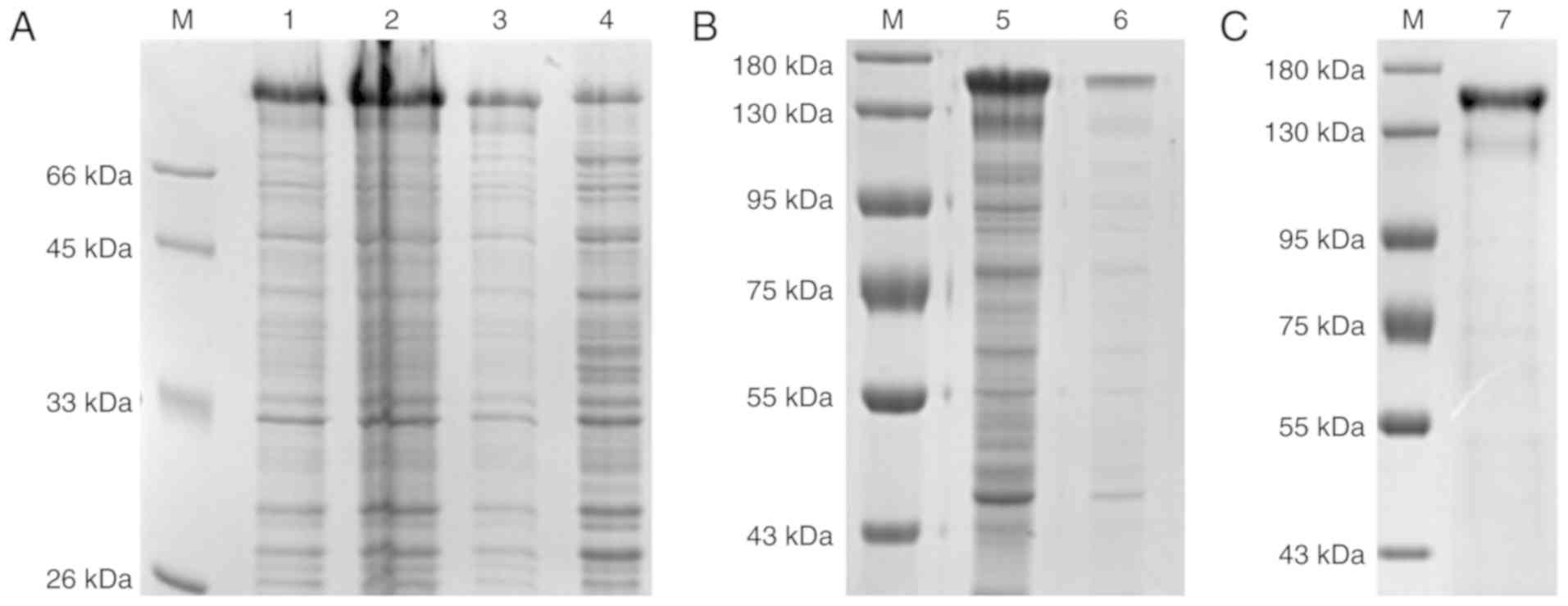
wLRRFIP1 was successfully
expressed
The wLRRFIP1 protein was successfully expressed as
determined by SDS-PAGE analysis (Fig.
2). The SDS-PAGE images illustrated that the wLRRFIP1 protein
was highly expressed in the ultrasonic precipitations.
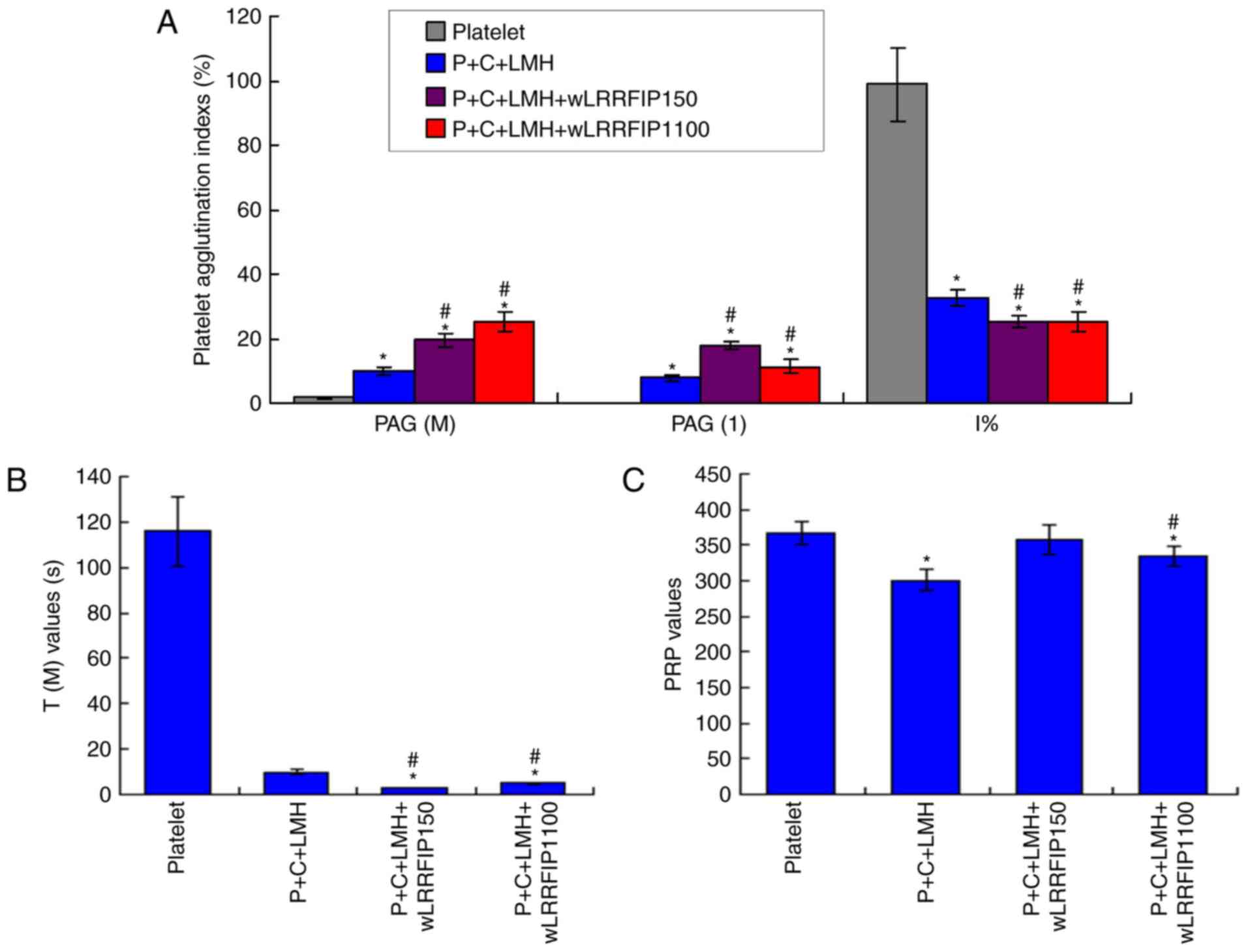
wLRRFIP1 treatment triggers platelet
agglutination of collagen treated platelets
A low dose of wLRRFIP1 (P+C+LMH+wLRRFIP150 group)
and a high-dose of wLRRFIP1 (P+C+LMH+wLRRFIP1100 group)
significantly increased the levels of adenosine diphosphate-PAG
(M), PAG (1) and significantly
decreased the I% values compared with the platelet group (all
P<0.05; Fig. 3A). Low- and
high-dose wLRRFIP1 significantly decreased the time for maximum PAG
[T(M)] values (Fig. 3B) and high
dose wLEEFIP1 significantly decreased the PRP values (Fig. 3C) compared with the platelet group
(all P<0.05), demonstrating that platelet agglutination
increased. Additionally, the P+C+LMH+wLRRFIP1100 group had a
decreased PAG (1) compared with the
P+C+LMH+wLRRFIP150 group.
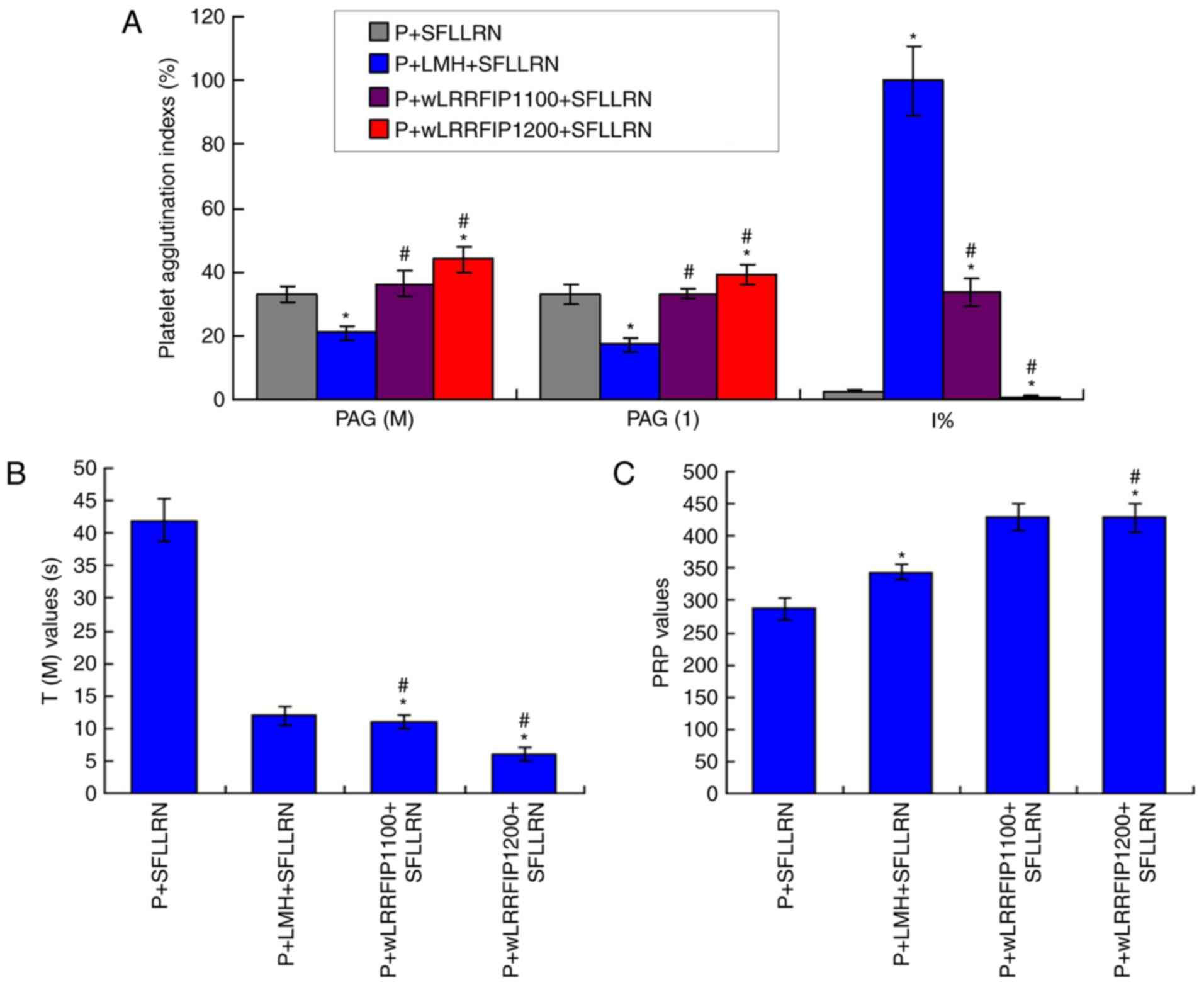
wLRRFIP1 treatment causes platelet
agglutination of thrombin receptor treated platelets
The levels of PAG (M) and PAG (1) were significantly increased, and I%
values were significantly decreased in the P+wLRRFIP1200+SFLLRN
group compared with the P+LMH+SFLLRN group (all P<0.05; Fig. 4A). PAG (M) and PAG (1) were slightly increased, and I% values
were significantly decreased in the P+wLRRFIP1100+SFLLRN group
compared with the P+SFLLRN group (P<0.05). In addition, wLRRFIP1
significantly decreased the T(M) values (Fig. 4B) and significantly increased the PRP
values (Fig. 4C) compared with the
P+SFLLRN group (all P<0.05), demonstrating that platelet
agglutination increased.
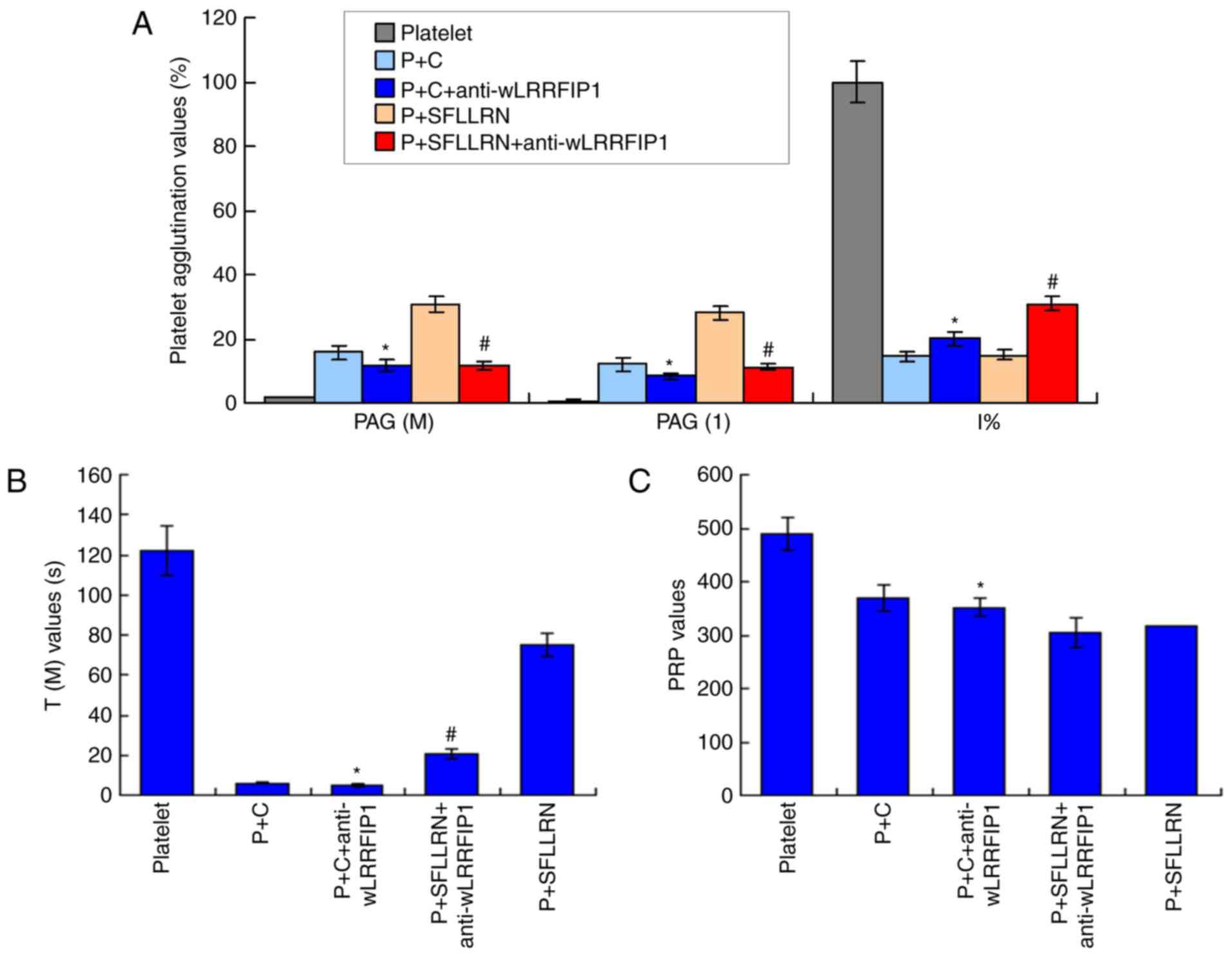
wLRRFIP1 treatment triggers platelet
agglutination of monoclonal antibody incubated platelets
The levels of PAG (M) and PAG (1) were significantly decreased and I%
values were significantly increased in P+C+anti-wLRRFIP1 group
compared with the P+C group (P<0.05; Fig. 5A). Anti-wLRRFIP1 treatment also
significantly affected the T (M) (Fig.
5B) and PRP values (Fig. 5C) in
the P+C+anti-wLRRFIP1 group when compared with that in the P+C
group. In addition, the levels of PAG (M) and PAG (1) were significantly decreased and the I%
values significantly increased in the P+SFLLRN+anti-wLRRFIP1 group
compared with the P+SFLLRN group (P<0.05; Fig. 5A). Anti-wLRRFIP1 treatment also
significantly affected the T (M) (Fig.
5B) and PRP values (Fig. 5C) in
the P+SFLLRN+anti-LRRFIP1 group compared with that in the P+SFLLRN
group. Therefore, wLRRFIP1 triggered platelet agglutination.
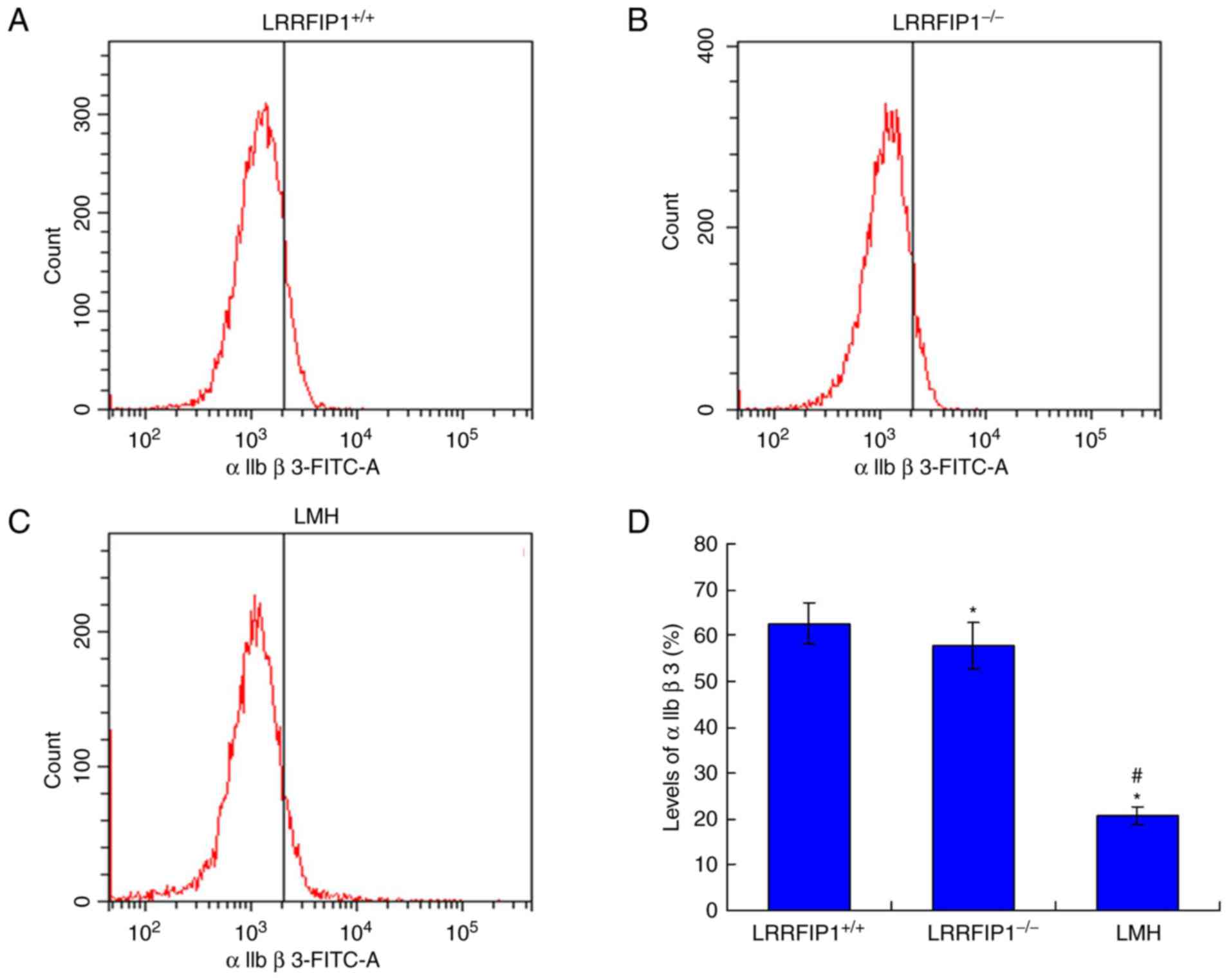
wLRRFIP1 knockout decreases αIIbβ3
levels
The αIIbβ3 integrin levels were examined by flow
cytometry. The results indicated that the levels of αIIbβ3 in the
LRRFIP1−/− group were significantly decreased compared
with the LRRFIP1+/+ group (P<0.05; Fig. 6). The levels of αIIbβ3 in the LMH
group were also significantly decreased compared with the
LRRFIP1+/+ group (P<0.05).
Platelet agglutination is
significantly inhibited in a mouse model of
LRRFIP1−/−
The results demonstrated that PAG (M), PAG (1) and I% values in the
P+LRRFIP1−/−, P+C+LRRFIP1−/− and
P+SFLLRN+LRRFIP1−/− groups were significantly decreased
compared with that in the P+LRRFIP1+/+,
P+C+LRRFIP1+/+ and P+SFLLRN+LRRFIP1+/+
groups, respectively (P<0.05; Fig.
7A). Platelet agglutination was significantly inhibited in the
P+SFLLRN+LRRFIP1−/− group compared with the
P+SFLLRN+LRRFIP1+/+ group as indicated by changes in the
levels of PAG (M), PAG (1) and I%.
The T (M) values in the P+LRRFIP1−/− and
P+C+LRRFIP1−/− groups were significantly decreased
compared with that in the P+LRRFIP1+/+ and
P+C+LRRFIP1+/+ groups, respectively (P<0.05; Fig. 7B). The T (M) values in the
P+SFLLRN+LRRFIP1−/− group were significantly increased
compared with that in the P+SFLLRN+LRRFIP1+/+ group
(P<0.05). The PRP values in the P+LRRFIP1−/−,
P+C+LRRFIP1−/− and P+SFLLRN+LRRFIP1−/− groups
were significantly increased compared with that in thr
P+LRRFIP1+/+, P+C+LRRFIP1+/+ and
P+SFLLRN+LRRFIP1+/+ groups, respectively (P<0.05;
Fig. 7C). Therefore, the changes
observed in the T (M) and PRP values also illustrated the
inhibition of platelet agglutination in the LRRFIP1−/−
mouse model compared with the LRRFIP1+/+ mouse
model.
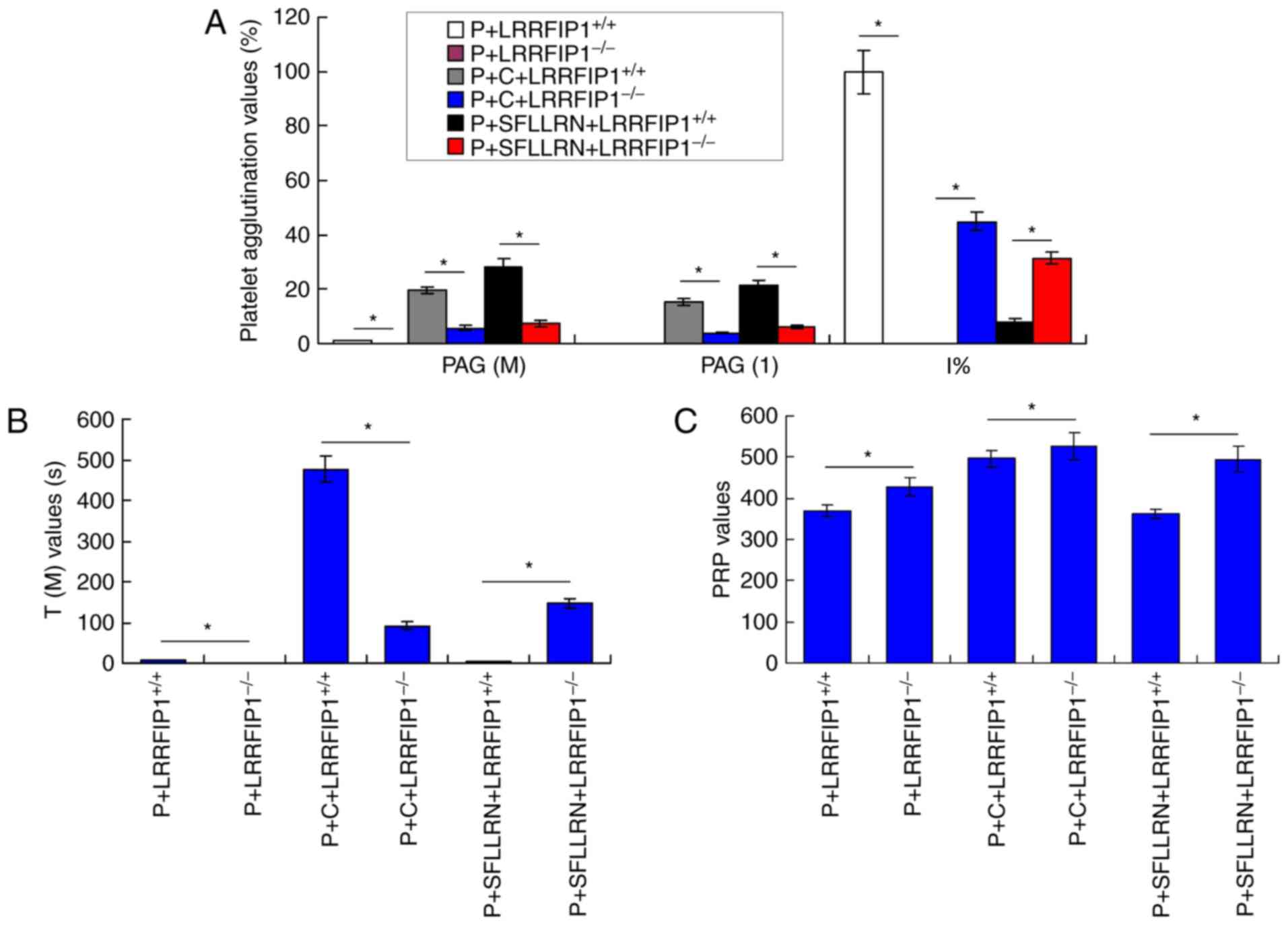 | Figure 7.Observation of the platelet
agglutination in a mouse model of LRRFIP1−/−. (A)
Platelet agglutination indexes of PAG (M), PAG (1) and I% of platelet agglutination. (B) T
(M) and (C) PRP values of platelet agglutination. *P<0.05. PAG,
adenosine diphosphate-platelet agglutination rate; PRP,
platelet-rich plasma; PAG (M), adenosine diphosphate-platelet
agglutination rate maximum; PAG (1),
PAG at 1 min; T (M), the time for maximum PAG; P, platelets; C,
collagen; SFLLRN, thrombin receptor activator;
LRRFIP1−/−, leucine-rich repeat flightless-interacting
protein-1 knockout mice; LRRFIP1+/+; leucine-rich repeat
flightless-interacting protein-1 positive mice. |
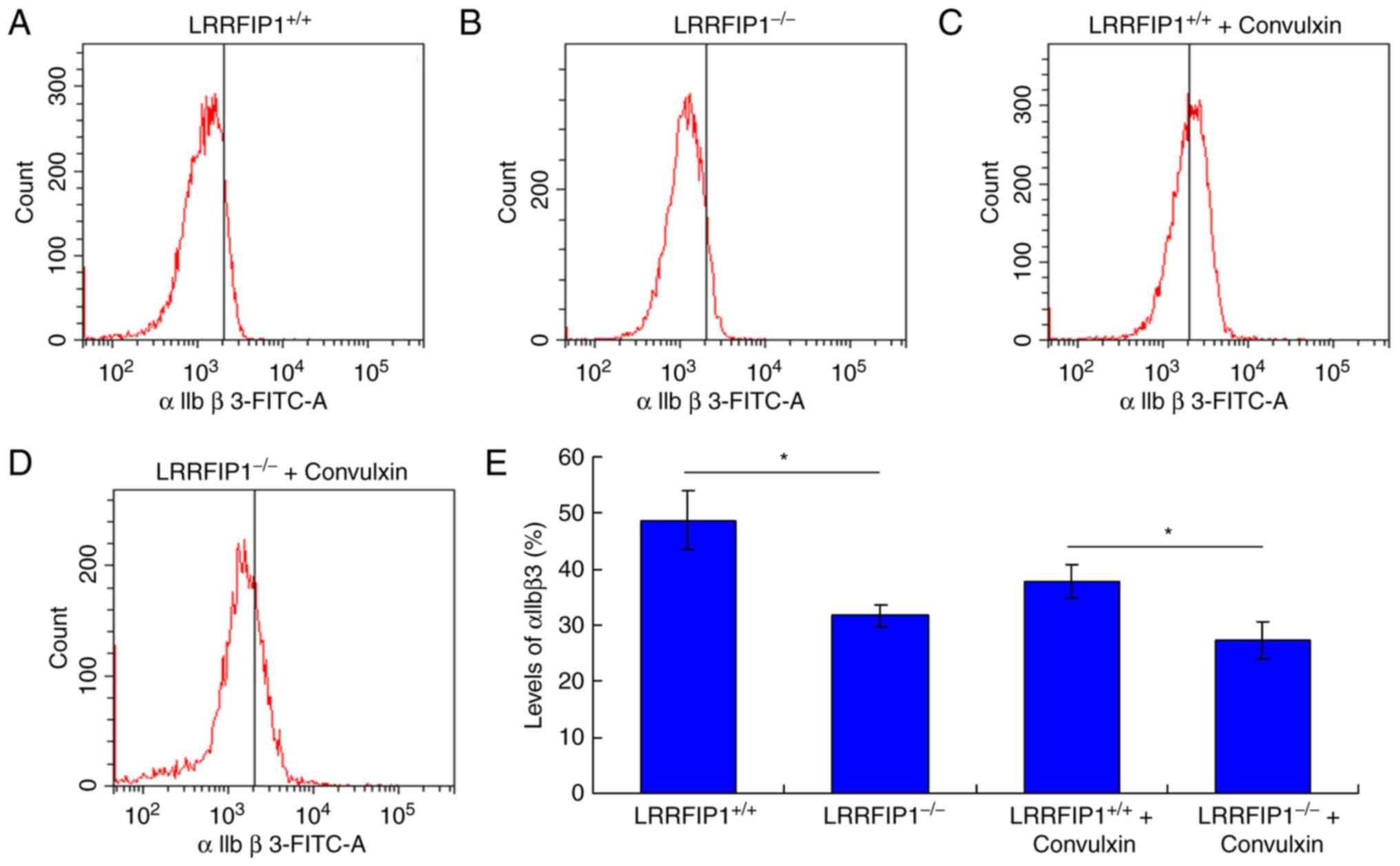
wLRRFIP1 knockout decreases αIIbβ3
levels in platelets treated with Convulxin
The αIIbβ3 integrin levels were examined by flow
cytometry. The results revealed that levels of αIIbβ3 in the
LRRFIP1−/− group were significantly decreased compared
with the LRRFIP1+/+ group (P<0.05; Fig. 8). The levels of αIIbβ3 in the
LRRFIP1−/− + Convulxin group were also significantly
decreased compared with the LRRFIP1+/+ + Convulxin group
(P<0.05; Fig. 8). This result
suggests that knocking out wLRRFIP1 reduced αIIbβ3 levels in the
platelets treated with Convulxin.
Discussion
Platelets are critical for atherothrombosis,
hemostasis and wound healing (24,25). The
platelet count and volume and the response of platelets are
independent risk factors for myocardial infarction; anti-platelet
therapy cannot reverse the effects of hyperactive platelets
(26). Although a number of
regulatory factors/proteins of platelet function have been
identified, the transcription profile of platelets is incomplete.
Goodall et al (18)
identified the nonsynonymous single nucleotide polymorphism (SNP;
SNP site: rs3739038) in the LRRFIP1 as a putative and novel signal
correlated with myocardial infarction. Therefore, the present study
investigated the effects of the LRRFIP1 on platelet
agglutination.
The LRRFIP1 gene is correlated with the regulation
of gene transcription, however its function in platelet regulation
has not been fully elucidated (20).
LRRFIP1 has also been identified as a repressor for the
transcription of genes, including platelet-derived growth factor
subunit A, epidermal growth factor receptor and tumor necrosis
factor α, by closely binding with their promoter regions (16). Previous studies have suggested that
LRRFIP1 serves as an important role in platelet activation, while
also regulating platelet agglutination (20,27).
The results of the present study indicated that
while cells were undergoing treatment with collagen, LMH and
SFLLRN, LRRFIP1 significantly increased the PAG (M), PAG (1) and PRP values and significantly
decreased the I% and T (M) values of platelets compared with the
controls. In addition, when the platelets were treated with
anti-wLRRFIP1 antibodies the PAG (M), PAG (1) and PRP values were significantly
decreased and the I% and T (M) values were significantly increased
compared with the untreated groups. A mouse model of LRRFIP1 gene
knockout was established and the results demonstrated that knockout
of the LRRFIP1 gene clearly increased the PAG (M), PAG (1) and PRP values and significantly
decreased the I% and T (M) values compared with the
wLRRFIP1+/+ mice. These results suggest that LRRFIP1 may
cause platelet agglutination and LRRFIP1 inhibition may alleviate
the platelet agglutination process. These findings are consistent
with a previously published study, which reported that LRRFIP1 acts
as a component of the platelet cytoskeleton and regulates platelet
responses (18).
A previous study reported that LRRFIP1 is correlated
with the interaction between fibrinogen and αIIbβ3 (28). Platelet aggregation and activation
are complex processes and involve a number of protein interactions
(29). Aggregation processes are
initiated by the activation of αIIbβ3, which binds to fibrinogen
and adjacent platelets (30,31). Chen et al (32) previously reported that anti-αIIbβ3
complex antibodies inhibited the aggregation of platelets. The
present study examined the levels of αIIbβ3 to identify the effects
of the LRRFIP1 on platelet agglutination. The results demonstrated
that the level of αIIbβ3 in the LRRFIP1−/− group was
significantly decreased compared with the LRRFIP1+/+
group. This suggests that LRRFIP1 gene knockout inhibits the
expression of αIIbβ3.
In conclusion, LRRFIP1 treatment induces platelet
agglutination and LRRFIP1 gene knockout inhibits platelet
agglutination. In addition, LRRFIP1 gene knockout significantly
decreases the levels of αIIbβ3. Therefore, LRRFIP1 triggers
platelet agglutination by enhancing the expression of αIIbβ3.
Acknowledgements
Not applicable.
Funding
This study was granted by the National Science
Foundation for Young Scientists of China (grant no. 81501883).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XY, PL, YYL, MYL and WLF performed the agglutination
assay. BYL conducted the SDS-PAGE. XY, PL and YYL performed the
preparation of LRRFIP1 recombinant protein and established the
LRRFIP1 knockout models. XY and JHZ wrote the manuscript and were
the primary designers of the study. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by Ethics Committee
of Daping Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Grozovsky R, Giannini S, Falet H and
Hoffmeister KM: Regulating billions of blood platelets: Glycans and
beyond. Blood. 126:1877–1884. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hattori H and Ishihara M: Feasibility of
improving platelet-rich plasma therapy by using chitosan with high
platelet activation ability. Exp Ther Med. 13:1176–1180. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ed Rainger G, Chimen M, Harrison MJ, Yates
CM, Harrison P, Watson SP, Lordkipanidzé M and Nash GB: The role of
platelets in the recruitment of leukocytes during vascular disease.
Platelets. 26:507–520. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gibbins JM: Platelet adhesion signalling
and the regulation of thrombus formation. J Cell Sci.
117:3415–3425. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Grozovsky R, Giannini S, Falet H and
Hoffmeister KM: Novel mechanisms of platelet clearance and
thrombopoietin regulation. Curr Opin Hematol. 22:445–451. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kurosawa Y, Nirengi S, Homma T, Esaki K,
Ohta M, Clark JF and Hamaoka T: A single-dose of oral nattokinase
potentiates thrombolysis and anti-coagulation profiles. Sci Rep.
5:116012015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xie J, Ma J, Huang Q, Yue C and Pei F:
Comparison of enoxaparin and rivaroxaban in balance of
anti-fibrinolysis and anticoagulation following primary total knee
replacement: A pilot study. Med Sci Monit. 23:704–711. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Z, Chen B, Yao XQ, Gui BS, Ou Y and
Ouyang JM: Anticoagulation of diethyl citrate and its comparison
with sodium citrate in an animal model. Blood Purif. 33:30–36.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yeung J, Tourdot BE, Fernandez-Perez P,
Vesci J, Ren J, Smyrniotis CJ, Luci DK, Jadhav A, Simeonov A,
Maloney DJ, et al: Platelet 12-LOX is essential for
FcgammaRIIa-mediated platelet activation. Blood. 124:2271–2279.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Violi F and Pignatelli P: Platelet NOX, a
novel target for anti-thrombotic treatment. Thromb Haemost.
111:817–823. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dütting S, Bender M and Nieswandt B:
Platelet GPVI. A target for antithrombotic therapy?! Trends
Pharmacol Sci. 33:583–590. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tsuji M, Ezumi Y, Arai M and Takayama H: A
novel association of Fc receptor gamma-chain with glycoprotein VI
and their co-expression as a collagen receptor in human platelets.
J Biol Chem. 272:23528–23531. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Suzuki-Inoue K, Fuller GL, García A, Eble
JA, Pöhlmann S, Inoue O, Gartner TK, Hughan SC, Pearce AC, Laing
GD, et al: A novel Syk-dependent mechanism of platelet activation
by the C-type lectin receptor CLEC-2. Blood. 107:542–549. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rosenfeld SI, Looney RJ, Leddy JP, Phipps
DC, Abraham GN and Anderson CL: Human platelet Fc receptor for
immunoglobulin G. Identification as a 40,000-molecular-weight
membrane protein shared by monocytes. J Clin Invest. 76:2317–2322.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhi H, Rauova L, Hayes V, Gao C, Boylan B,
Newman DK, McKenzie SE, Cooley BC, Poncz M and Newman PJ:
Cooperative integrin/ITAM signaling in platelets enhances thrombus
formation in vitro and in vivo. Blood. 121:1858–1867. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suriano AR, Sanford AN, Kim N, Oh M,
Kennedy S, Henderson MJ, Dietzmann K and Sullivan KE: GCF2/LRRFIP1
represses tumor necrosis factor alpha expression. Mol Cell Biol.
25:9073–9081. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Santiago FS and Khachigian LM: Ets-1
stimulates platelet-derived growth factor A-chain gene
transcription and vascular smooth muscle cell growth via
cooperative interactions with Sp1. Circ Res. 95:479–487. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Goodall AH, Burns P, Salles I, Macaulay
IC, Jones CI, Ardissino D, de Bono B, Bray SL, Deckmyn H, Dudbridge
F, et al: Transcription profiling in human platelets reveals
LRRFIP1 as a novel protein regulating platelet function. Blood.
116:4646–4656. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Burns P, Gusnanto A, Macaulay IC, Rankin
A, Tom B, Langford CF, Dudbridge F, Ouwehand WH and Watkins NA;
Bloodomics Consortium, : Identification of variation in the
platelet transcriptome associated with glycoprotein 6 haplotype.
Platelets. 19:258–267. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fong KS and de Couet HG: Novel proteins
interacting with the leucine-rich repeat domain of human
flightless-I identified by the yeast two-hybrid system. Genomics.
58:146–157. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jones-Bolin S: Guidelines for the care and
use of laboratory animals in biomedical research. Curr Protoc
Pharmacol Appendix 4. Appendix 4B. 2012. View Article : Google Scholar
|
|
22
|
Dennis MB Jr: Humane endpoints for
genetically engineered animal models. ILAR J. 41:94–98. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Langer HF, Choi EY, Zhou H, Schleicher R,
Chung KJ, Tang Z, Gobel K, Bdeir K, Chatzigeorgiou A, Wong C, et
al: Platelets contribute to the pathogenesis of experimental
autoimmune encephalomyelitis. Circ Res. 110:1202–1210. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sorrentino S, Studt JD, Medalia O and
Tanuj Sapra K: Roll, adhere, spread and contract: Structural
mechanics of platelet function. Eur J Cell Biol. 94:129–138. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ding C, Zhang J, Li R, Wang J, Hu Y, Chen
Y, Li X and Xu Y: Investigation of standardized administration of
anti-platelet drugs and its effect on the prognosis of patients
with coronary heart disease. Exp Ther Med. 14:3207–3212. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Breet NJ, van Werkum JW, Bouman HJ, Kelder
JC, Ruven HJ, Bal ET, Deneer VH, Harmsze AM, van der Heyden JA,
Rensing BJ, et al: Comparison of platelet function tests in
predicting clinical outcome in patients undergoing coronary stent
implantation. JAMA. 303:754–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu YT and Yin HL: Identification of the
binding partners for flightless I, A novel protein bridging the
leucine-rich repeat and the gelsolin superfamilies. J Biol Chem.
273:7920–7927. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yin X, Wu S, Wang Z, Wang Y, Du Q and Wang
A: Anti-thrombosis effect of LRRFIP1 shRNA lentivirus in a mouse
model of deep vein thrombosis. Thromb Res. 132:127–131. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bhoria P, Varma N, Malhotra P, Varma S and
Luthra-Guptasarma M: Immunodiagnosis of platelet activation in
immune thrombocytopenia through scFv antibodies cognate to
activated IIb3 integrins. MAbs. 7:1212–1220. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fullard JF: The role of the platelet
glycoprotein IIb/IIIa in thrombosis and haemostasis. Curr Pharm
Des. 10:1567–1576. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bennett JS: Structure and function of the
platelet integrin alphaIIbbeta3. J Clin Invest. 115:3363–3369.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen P, Sun CX and Liu JN: A novel
anti-platelet monoclonal antibody (3C7) specific for the complex of
integrin alpha IIb beta3 inhibits platelet aggregation and
adhesion. J Biol Chem. 280:25403–25408. 2005. View Article : Google Scholar : PubMed/NCBI
|