Introduction
Diabetes mellitus (DM) is a chronic metabolic
disease that has become a major health problem worldwide. In 2014,
an estimated 387 million individuals worldwide were diagnosed with
DM and the prevalence is expected to rise to 592 million by 2035
(1). In addition, patients with DM
have a high mortality rate that is often caused by diabetic
cardiomyopathy (DCM) (2,3).
DCM is characterized by left ventricular hypertrophy
and reduced diastolic function, and is a major complication in
patients with either type 1 or type 2 DM (4). Previous studies have demonstrated that
diverse pathogenic mechanisms contribute to DCM, including
hyperglycemia, inflammation, fibrosis and apoptosis (5,6). Among
these factors, cardiomyocyte apoptosis is thought to initiate
cardiac remodeling and results in cardiac dysfunction (7). Therefore, it serves a key function in
the pathogenesis and progression of DCM. Accumulating evidence has
indicated that cardiomyocyte endoplasmic reticulum (ER) stress is
involved in the pathogenesis of cardiac dysfunction in patients
with DM and animal models (8,9). The
early stages of ER stress are considered to be an adaptive response
aiming at maintenance of ER homeostasis. This is known as the
unfolded protein response (UPR), which is monitored by
glucose-regulated protein 78 (Grp78), an ER chaperone (10). If it persists long-term, ER stress
could induce the intrinsic pathway of apoptosis (11). C/EBP homologous protein (CHOP), a
marker of ER stress, was identified to be elevated in multiple
organs of a diabetic animal model, including myocardial tissue
(9,12,13).
Furthermore, CHOP knockout mice were observed to exhibit less
pronounced hypertrophy and cardiac dysfunction in comparison with
wild-type animals (14).
Tanshinone IIA (TSIIA), a phytochemical derived from
the roots of Salvia miltiorrhiza, has the effect of
suppressing apoptosis via multiple pathways (15,16). In
previous studies, it has been demonstrated to inhibit ER-induced
apoptosis in certain tissues (17,18).
Therefore, it was hypothesized that TSIIA could improve cardiac
function by inhibiting ER stress-induced apoptosis. The present
study performed comparative studies on ER stress-associated
signaling proteins Grp78 and CHOP to explore the mechanism of TSIIA
on protecting cardiac function in a streptozotocin (STZ)-induced
diabetic rat model.
Materials and methods
Animals
The experimental procedures were reviewed and
approved by the Committee for the Care and Use of Laboratory
Animals at Zhejiang Chinese Medical University (Hangzhou, China). A
total of 40 6-week-old male Sprague-Dawley rats (160–180 g;
Shanghai SLAC Laboratory Animal Co., Ltd., Shanghai, China) were
individually housed in separately ventilated cages at a temperature
of 23±2°C and humidity of 55±2% with a 12-h light/dark cycle, and
were given free access to standard food and water.
Materials
TSIIA was purchased from Chiatai Qingchunbao
Pharmaceutical Co., Ltd., (Hangzhou, China). STZ was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). The blood glucose
test machine (FreeStyle Optium Neo) and strips were purchased from
Abbott Laboratories (Lake Bluff, IL, USA). Grp78 (cat. no. 3813)
and CHOP (cat. no. 2895) antibodies were purchased from Cell
Signaling Technology, Inc., (Danvers, MA, USA). The β-actin
antibody (cat. no. AbD12141) was purchased from Bio-Rad
Laboratories, Inc., (Hercules, CA, USA).
Induction of diabetes in rats
A total of 40 male rats were randomized into the
following groups (n=10 per group): Control group, DM group, DM rats
treated with lower-dose TSIIA (L-TSIIA group) and DM rats treated
with high-dose TSIIA (H-TSIIA group). DM was induced by a single
intraperitoneal injection of STZ (dissolved in 10 mM citrate
buffer, pH 4.5) at a dosage of 60 mg/kg body weight to induce
diabetes. Rats in groups with TSIIA injection were
intraperitoneally administered daily at a dose of 2 mg/kg (L-TSIIA
group) and 4 mg/kg (H-TSIIA group) for 6 weeks, while the rats in
the control and diabetes groups were injected with the same volume
of vehicle (PBS). At day 2 after STZ injection, tail-vein blood
glucose concentration was measured. Rats with blood glucose level
>16 mmol/l were identified as diabetic model rats. Body weight
and fasting blood glucose were determined at the end of week 6.
TSIIA was dissolved in DMSO at a concentration of 10 mg/ml and
then, diluted to 0.5 mg/ml with PBS prior to injection.
Blood and tissue sample
preparation
At the end of week 6 following STZ injection, rats
were anesthetized in an induction chamber with 2–3% isoflurane
(Abbott Laboratories), then intraperitoneally injected with 1%
pentobarbital sodium salt (30 mg/kg; Sigma-Aldrich; Merck KGaA).
Blood was collected from carotid arteries and left at room
temperature for 1 h. The blood samples were subsequently
centrifuged at 1,500 × g for 15 min, followed by the separation of
serum from the blood cells. Serum was preserved at −20°C for
evaluating high-sensitivity superoxide dismutase (SOD), which was
performed using a SOD assay kit (cat. no. A001-1-1; NanJing
JianCheng Bioengineering Institute, Nanjing, China) After
collecting blood, all rats were perfused with 50 ml saline. Next, 5
rats in each group were perfused with 200 ml 4% paraformaldehyde
solution for subsequent transmission electron microscopy (TEM)
observation. The myocardial tissue from another 5 rats (in each
group) was preserved in liquid nitrogen for detecting the
expression of relevant genes and proteins by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blotting, respectively.
TEM
Following perfusion with 4% paraformaldehyde, the
myocardial tissue from the left ventricle was dissected, then
rinsed in 2.5% glutaraldehyde at 4°C for 4 h. The tissues were
rinsed in buffer and post-fixed with 1% osmium tetroxide for 1 h at
room temperature, dehydrated in graded alcohol, then transferred to
propylene oxide. The tissues were embedded gradually in blocks of
Epon 812 resin for 2 days at 60°C undergoing a graded ethanol
dehydration series, and infiltrated using a mixture of 50%
propylene oxide and 50% resin overnight. Then, 24 h later, the
tissues were embedded in pure resin for 2 days at 60°C. Then, 120
nm-thick sections were cut using a histology diamond knife on an
Ultracut E microtome (Leica Microsystems GmbH, Wetzlar, Germany)
and stained with 4% uranyl acetate for 20 min and with 0.5% lead
citrate for 5 min at room temperature. The ultrastructure of
mitochondria and myofibrils was observed under TEM (Philips Tecnai
10; Philips Medical Systems B.V., Eindhoven, The Netherlands).
Mitochondrial subpopulation densities were determined within a
defined region (100 µm2) with a minimum of 12 images in
each group, which were taken at ×5,900 magnification, as described
previously (19). Analysis was
performed using Image J software (v.1.46; National Institutes of
Health, Bethesda, MD, USA).
Western blotting
Total protein from rat myocardium tissues was
extracted using ice-cold RIPA buffer (cat. no. P0013B; Beyotime
Institute of Biotechnology, Haimen, China). Protein concentration
was determined using a BCA Protein Assay Reagent kit (cat. no.
23225; Pierce; Thermo Fisher Scientific, Inc.). Protein (30 µg) was
separated by 12% SDS-PAGE and transferred to a PVDF membrane. The
membranes were blocked with 5% non-fat milk for 2 h at room
temperature, then incubated overnight at 4°C with primary
antibodies (monoclonal anti-CHOP (cat. no. 2895; Cell Signaling
Technology, Inc.; 1:1,000) and polyclonal GRP-78 (1:1,000; cat. no.
3177; Cell Signaling Technology, Inc.). After three washes with
TBST, blots were incubated with horseradish peroxidase conjugated
anti-mouse secondary antibodies (1:10,000; cat. no. BA1050; Wuhan
Boster Biological Technology, Ltd., Wuhan, China) and horseradish
peroxidase conjugated anti-rabbit secondary antibody (1,10,000;
cat. no. BA2006; BOSTER Biological Tech Co., Ltd., Wuhan, China)
for 1 h at room temperature. The immunoreactive bands were
visualized using a chemiluminescent reagent as recommended by the
Supersignal West Dura Extended Duration Substrate kit (cat. no.
34075, Pierce; Thermo Fisher Scientific, Inc.). The signals of the
bands were quantified using the VICTOR-Z 1420 multilabel counter
(PerkinElmer, Inc., Waltham, MA, USA). Equal protein loading in
each lane was confirmed by hybridization with a 1:10,000 dilution
of β-actin antibody (cat. no. AbD12141; Bio-Rad Laboratories,
Inc.). The results were expressed as density relative to β-actin as
determined via Image J software (v.1.46).
RT-qPCR
Myocardial tissue preserved in liquid nitrogen was
used for RT-qPCR analysis. Total RNA was isolated from the
specimens using the TRIzol reagent kit (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. Total
RNA (1 µg) was reverse transcribed to cDNA using the Transcriptor
First Strand cDNA Synthesis kit (Takara Biotechnology Co., Ltd.,
Tokyo, Japan). qPCR was performed using Power SYBR Green qPCR
Master mix (Applied Biosystems; Thermo Fisher Scientific, Inc.).
cDNA was denatured by heating samples to 94°C for 3 min. The
template was amplified by 40 rounds of PCR (denaturation at 94°C
for 10 sec, annealing at 60°C for 30 sec and extension at 72°C for
30 sec) before measuring fluorescence at 72°C. PCR primers
(Invitrogen; Thermo Fisher Scientific, Inc.) for ER
stress-associated genes: Grp-78 (forward,
5′-GGTGCAGCAGGACATCAAGTT-3′; reverse,
5′-CCCACCTCCAATATCAACTTGA-3′), CHOP (forward,
5′-CTGCCTTTCACCTTGGAGAC-3′; reverse, 5′-CGTTTCCTGGGGATGAGATA-3′)
and GAPDH (forward 5′-GGTGGACCTCATGGCCTACAT-3′ and reverse
5′-GCCTCTCTCTTGCTCTCAGTATCCT-3′). The comparative Cq
method was utilized as described previously (20).
Statistical analysis
Data are expressed as the mean ± standard deviation.
A two-tailed, unpaired Student's t-test was used to compare two
groups. Multiple group comparisons were performed using one-way
analysis of variance, followed by Newman-Keuls post-hoc test.
P<0.05 was considered to indicate a statistically significant
difference. Data were analyzed using GraphPad Prism 6 software
(GarphPad Software, Inc., La Jolla, CA, USA).
Results
Effect of TSIIA on body weight and
blood glucose concentration
There were no significant differences among groups
in terms of body weight or blood glucose concentration prior to STZ
injection (Table I). However, body
weight in diabetic rats was observed to be significantly decreased
(P<0.01; Table I) at 6 weeks when
compared with the control. There were no significant differences
between the L-TSIIA and H-TSIIA groups (Table I). Furthermore, blood glucose
concentration was significantly increased at 2 days and 6 weeks
following STZ injection in the diabetes group as compared with the
control group (P<0.01; Table I).
TSIIA was beneficial for reducing blood glucose; the concentration
of blood glucose was significantly decreased in the L-TSIIA group
(20.78±2.88 mmol/l) at 6 weeks when compared with the diabetes
group (P<0.05; Table I).
 | Table I.Effect of TSIIA on body weight and
blood glucose concentration. |
Table I.
Effect of TSIIA on body weight and
blood glucose concentration.
|
| Body weight (g) | Blood glucose
concentration (mmol/l) |
|---|
|
|
|
|
|---|
| Group | Prior to STZ
injection | 2 days post-STZ
injection | 6 weeks post-STZ
injection | Prior to STZ
injection | 2 days post-STZ
injection | 6 weeks post-STZ
injection |
|---|
| Control | 133.27±3.67 | 141.59±4.92 | 404.85±31.48 | 5.71±0.55 | 6.25±0.64 | 5.67±0.43 |
| Diabetes | 134.16±4.03 | 142.02±5.63 |
299.26±47.14a | 5.64±0.66 |
21.87±3.35a |
25.07±2.59a |
| L-TSIIA | 136.40±4.23 | 144.20±5.51 |
311.99±49.92a | 6.12±0.79 |
21.78±4.73a |
20.78±2.88a,b |
| H-TSIIA | 130.08±5.39 | 138.68±6.55 |
289.50±42.18a | 5.68±0.52 |
23.89±4.18a |
23.46±3.05a |
TSIIA injection rescues injured
mitochondrial structure and reduces mitochondrial density caused by
hyperglycemia
TEM examination revealed that mitochondrial
structure was normal in the control group (Fig. 1A), while the mitochondrial cristae of
the myocardial cells of the diabetic group disappeared and the
vacuoles in the swollen mitochondrial matrix increased, which are
thought to be characteristics of apoptosis (Fig. 1B). Furthermore, disordered myofibrils
were observed in the myocardial cells of diabetic rats (Fig. 1B), but not the control group.
Administration of TSIIA was observed to attenuate the damage
induced by hyperglycemia (Fig. 1C and
D). Furthermore, it was observed that the mitochondrial density
was significantly decreased in the diabetes group (P<0.01),
which was rescued by H-TSIIA treatment (P<0.05 vs. diabetes
group; Table II).
| Table II.Quantification of mitochondrial
density under transmission electron microscopy. |
Table II.
Quantification of mitochondrial
density under transmission electron microscopy.
| Groups | Mitochondrial
density/100 µm2 |
|---|
| Control |
47.56±12.90 |
| Diabetes |
28.12±8.39a |
| L-TSIIA |
28.14±8.39a |
| H-TSIIA |
40.62±14.47b |
Expression of ER stress-associated
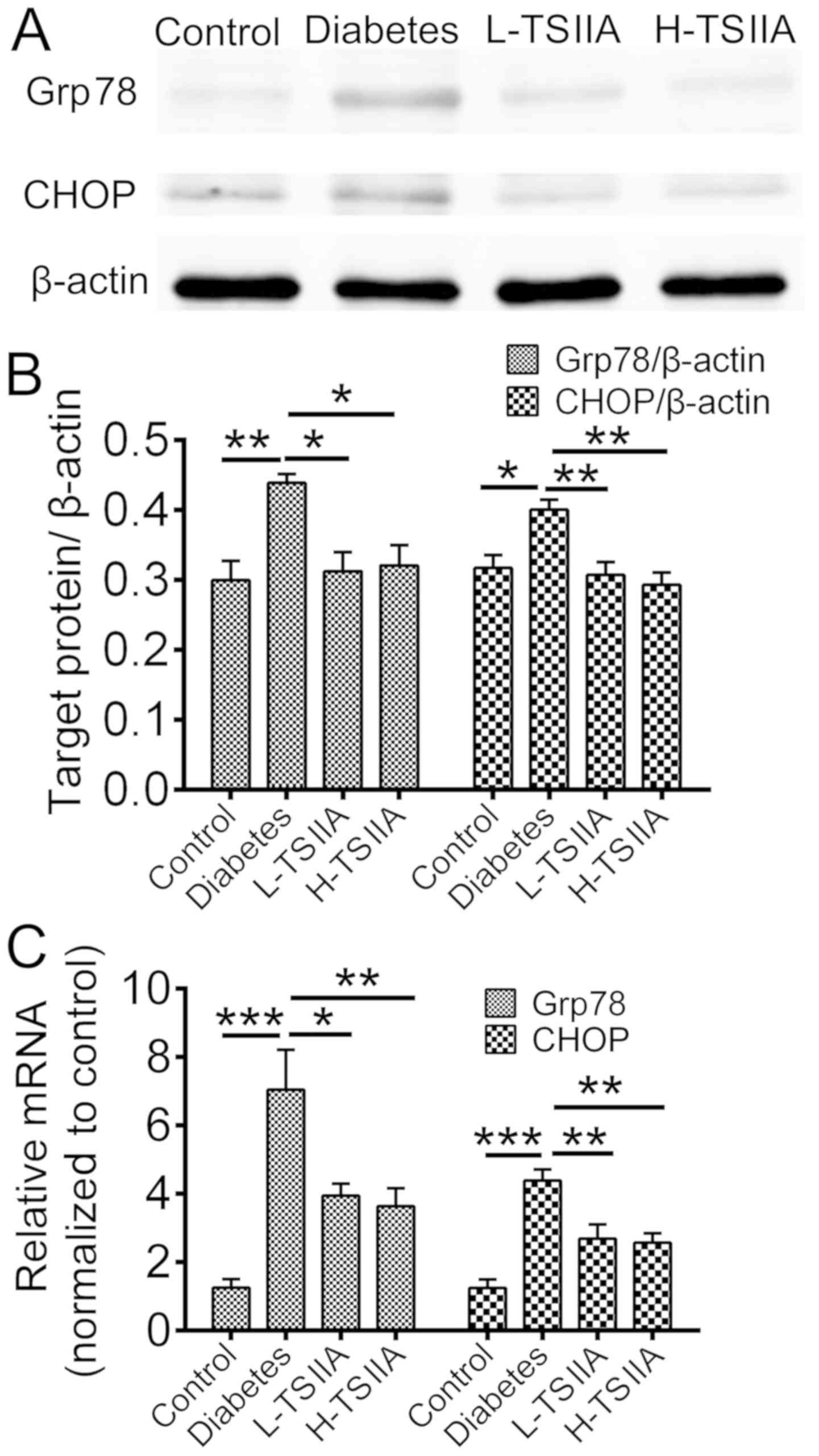
proteins
The mRNA and protein expression levels of Grp78, an
ER chaperone, and CHOP, a protein crucial to growth arrest and DNA
damage, were measured. According to RT-qPCR and western blot
analysis results, the gene and protein levels of Grp78 and CHOP
were significantly enhanced in the diabetes group as compared with
the control group (P<0.05; Fig.
2). However, TSIIA application during treatment could reduce
Grp78 and CHOP expression (P<0.05; Fig. 2). This suggested that ER stress was
activated in myocardium tissue of diabetic rats, which could be
suppressed by TSIIA treatment. These results indicate that TSIIA
may be able to rescue injured mitochondrial structure by
hyperglycemia via inhibition of the ER stress response.
TSIIA reduces the expression of
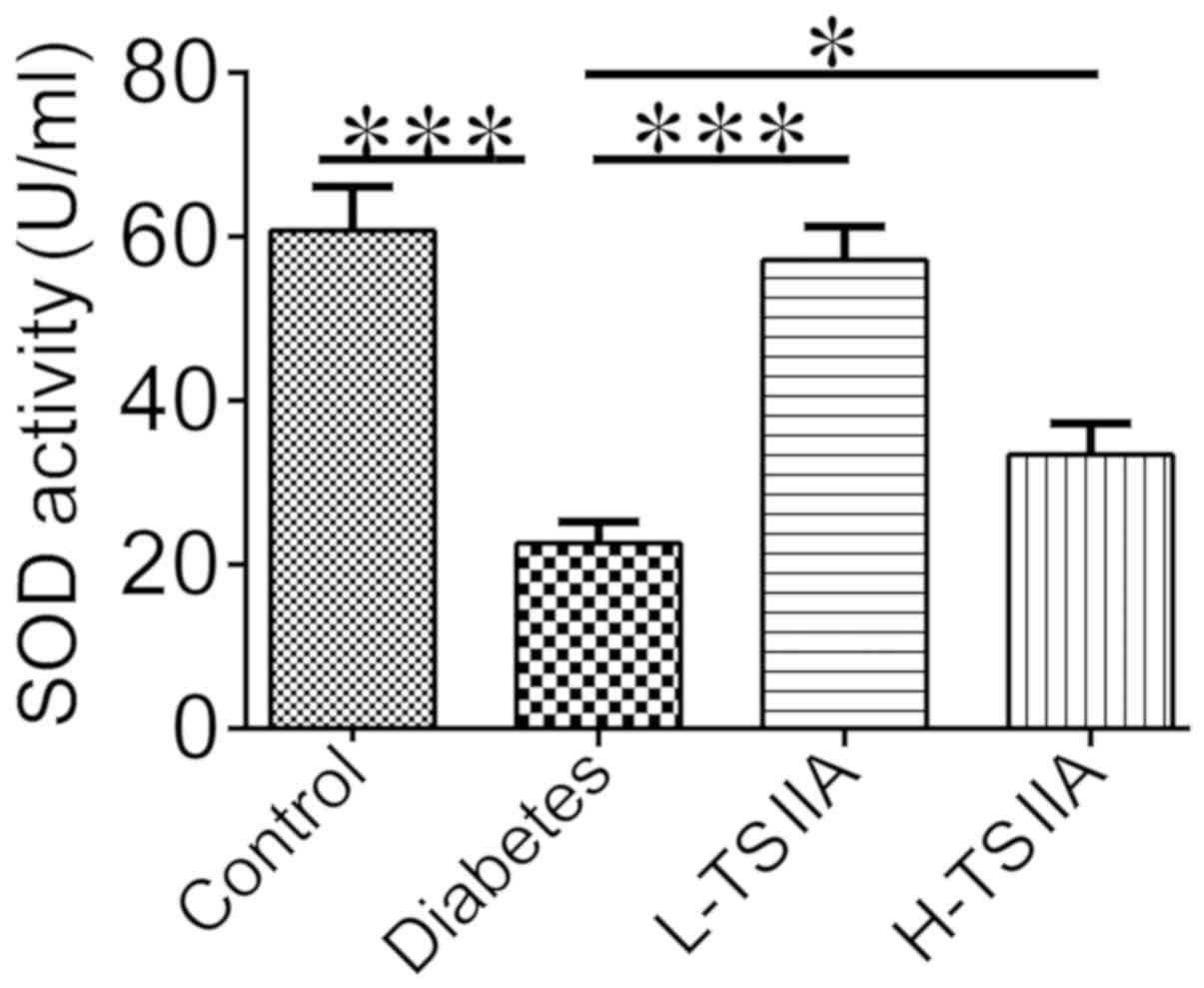
SOD
Oxidative stress could trigger ER stress activation
(21). SOD, as an important
antioxidant, was measured in the serum of rats to investigate the
mechanism of TSIIA protective effects on DCM. As indicated in
Fig. 3, SOD activity was
significantly decreased in diabetic rats (22.64±8.18 U/ml) when
compared with the control group (60.74±16.92 U/ml; P<0.001).
TSIIA treatment significantly increased SOD activity in L-TSIIA
(P<0.001) and H-TSIIA rats (P<0.05) when compared with
non-treated diabetic rats.
Discussion
DCM, which is now recognized to have a high
prevalence among patients with diabetes, leads to structural and
functional changes of myocardial tissue (22). The aim of the current study was to
investigate the effect and mechanism of TSIIA on cardiac
dysfunction in a diabetic rat model. The current results indicated
that improved cardiac pathological changes were accompanied by
rescued myocardial damage following TSIIA treatment. This suggested
that TSIIA exerts a beneficial effect on DCM. The underlying
mechanism was also investigated in the present study; to the best
of our knowledge, this has not been reported previously.
As a key component of S. miltiorrhiza, TSIIA
has been used to treat multiple diseases, including Alzheimer's
disease, chronic kidney disease, cerebral ischemic injury and DCM
(23–26). Furthermore, previous studies have
demonstrated that TSIIA could attenuate myocardial ischemia
reperfusion injury due to its antioxidant capacity (27). In the present study, it was observed
that SOD activity was significantly reduced in the serum of
diabetic rats compared with the control group, while TSIIA at
different dosages could elevate the activity of SOD. SOD is known
to be a major scavenger of superoxide radicals, which may attenuate
oxidative stress (28). Therefore,
it is proposed that TSIIA may alleviate the injury induced by
hyperglycemia via increased SOD activity. In the current study, the
ultrastructure of mitochondria and myofibrils was also observed
under TEM. Previous studies have reported that STZ-induced
hyperglycemia elevates free radicals and represses antioxidant
defense mechanisms, which may lead to cell disruption, degenerated
mitochondria and disordered myofibrils (29,30) and
may induce DCM after 6 weeks (31).
Consistent with these findings, in the current study, a lack of
mitochondrial cristae and disordered myofibrils were observed in
diabetic rats, which was accompanied by a reduction in
mitochondrial density. However, it was identified that
mitochondrial cristae were partially restored, and mitochondrial
density was rescued, by application of TSIIA, which may be
associated with increased activity of SOD. A previous study also
reported that mitochondrial structure plays an important role in
cardiac function (32). Degradation
in mitochondrial structure results in a progressive increase in
mitochondrial Ca2+ sequestration and mitochondrial
depolarization, finally leading to cardiac dysfunction (33).
Previous research has identified direct associations
between hyperglycemia, oxidative stress and protein folding
(21). Alterations in redox state
could result in unfolded/misfolded proteins accumulating in the ER
and UPR being initiated, generally referred as ER stress (34). Accumulation of unfolded/misfolded
proteins could also increase reactive oxygen species production and
therefore exacerbate ER stress. One major goal of UPR/ER stress is
to induce ER chaperones that promote protein folding, which is
controlled by the ER chaperone, Grp78 (35). Activation of Grp78 protein expression
is a widely used indicator of ER stress. In the current study, the
expression of Grp78 was upregulated in the myocardial tissue of
diabetic rats, which was accompanied by the increased expression of
CHOP, a key mediator of ER stress-induced apoptosis. These results
suggested that ER stress was activated in myocardial tissue of the
STZ-induced diabetic animal model. However, the expression of Grp78
was significantly decreased following TSIIA treatment when compared
with that in the diabetic group. Furthermore, CHOP is involved in
one of the ER stress-induced cell apoptosis signaling pathways that
could trigger cell apoptosis (36).
In the present study, TSIIA inhibited CHOP apoptosis signaling
pathways. Therefore, it was suggested that TSIIA may alleviate
cardiomyocyte ER stress-induced apoptosis.
The present results demonstrated that TSIIA may
exert a therapeutic effect on DCM. TSIIA was observed to improve
cardiac pathological changes, and this effect may be via inhibiting
ER stress-mediated myocardial cell apoptosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Zhejiang Province, China (grant no. LY
16H270003).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
YZ designed the study. Diabetes animal model
induction, blood and tissue sample preparation was performed by ST
and LC. Sample preparation for TEM was done by JS, NZ and XS
performed Western blot and real time PCR. RS and GG analyzed data.
The manuscript was written by ST. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedures were reviewed and
approved by the Committee for the Care and Use of Laboratory
Animals at Zhejiang Chinese Medical University (Hangzhou,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Deng F, Wang S and Zhang L: Endothelial
microparticles act as novel diagnostic and therapeutic biomarkers
of diabetes and its complications: A literature review. Biomed Res
Int. 2016:98020262016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Boudina S and Abel ED: Diabetic
cardiomyopathy, causes and effects. Rev Endocr Metab Disord.
11:31–39. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li WF, Wang P, Li H, Li TY, Feng M and
Chen SF: Oleanolic acid protects against diabetic cardiomyopathy
via modulation of the nuclear factor erythroid 2 and insulin
signaling pathways. Exp Ther Med. 14:848–854. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Asghar O, Al-Sunni A, Khavandi K, Khavandi
A, Withers S, Greenstein A, Heagerty AM and Malik RA: Diabetic
cardiomyopathy. Clin Sci (Lond). 116:741–760. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Westermann D, Walther T, Savvatis K,
Escher F, Sobirey M, Riad A, Bader M, Schultheiss HP and Tschöpe C:
Gene deletion of the kinin receptor B1 attenuates cardiac
inflammation and fibrosis during the development of experimental
diabetic cardiomyopathy. Diabetes. 58:1373–1381. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Falcao-Pires I and Leite-Moreira AF:
Diabetic cardiomyopathy: Understanding the molecular and cellular
basis to progress in diagnosis and treatment. Heart Fail Rev.
17:325–344. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xia Y, Gong L, Liu H, Luo B, Li B, Li R,
Li B, Lv M, Pan J and An F: Inhibition of prolyl hydroxylase 3
ameliorates cardiac dysfunction in diabetic cardiomyopathy. Mol
Cell Endocrinol. 403:21–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang L, Zhao D, Ren J and Yang J:
Endoplasmic reticulum stress and protein quality control in
diabetic cardiomyopathy. Biochim Biophys Acta. 1852:209–218. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang X, Liu S, Zhang G, Zhong M, Liu T,
Wei M, Wu D, Huang X, Cheng Y, Wu Q and Hu S: Bariatric surgery
ameliorates diabetic cardiac dysfunction by inhibiting ER stress in
a diabetic rat model. Obes Surg. 27:1324–1334. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu LH, Xie H, Shi ZH, Du LD, Wing YK, Li
AM, Ke Y and Yung WH: Critical role of endoplasmic reticulum stress
in chronic intermittent hypoxia-induced deficits in synaptic
plasticity and long-term memory. Antioxid Redox Signal. 23:695–710.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iurlaro R and Muñoz-Pinedo C: Cell death
induced by endoplasmic reticulum stress. FEBS J. 283:2640–2652.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang X, Xu L, He D and Ling S:
Endoplasmic reticulum stress-mediated hippocampal neuron apoptosis
involved in diabetic cognitive impairment. Biomed Res Int.
2013:9243272013.PubMed/NCBI
|
|
13
|
Liu X, Xu Q, Wang X, Zhao Z, Zhang L,
Zhong L, Li L, Kang W, Zhang Y and Ge Z: Irbesartan ameliorates
diabetic cardiomyopathy by regulating protein kinase D and ER
stress activation in a type 2 diabetes rat model. Pharmacol Res.
93:43–51. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Minamino T, Komuro I and Kitakaze M:
Endoplasmic reticulum stress as a therapeutic target in
cardiovascular disease. Circ Res. 107:1071–1082. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cui ZT, Liu JP and Wei WL: The effects of
tanshinone IIA on hypoxia/reoxygenation-induced myocardial
microvascular endothelial cell apoptosis in rats via the JAK2/STAT3
signaling pathway. Biomed Pharmacother. 83:1116–1126. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Z, Li Y, Sheng C, Yang C, Chen L and
Sun J: Tanshinone IIA inhibits apoptosis in the myocardium by
inducing microRNA-152-3p expression and thereby downregulating
PTEN. Am J Transl Res. 8:3124–3132. 2016.PubMed/NCBI
|
|
17
|
Zhang X, He D, Xu L and Ling S: Protective
effect of tanshinone IIA on rat kidneys during hypothermic
preservation. Mol Med Rep. 5:405–409. 2012.PubMed/NCBI
|
|
18
|
Yang GL, Jia LQ, Wu J, Ma YX, Cao HM, Song
N and Zhang N: Effect of tanshinone IIA on oxidative stress and
apoptosis in a rat model of fatty liver. Exp Ther Med.
14:4639–4646. 2017.PubMed/NCBI
|
|
19
|
Ji W, Chen X, Lv J, Wang M, Ren S, Yuan B,
Wang B and Chen L: Liraglutide exerts antidiabetic effect via PTP1B
and PI3K/Akt2 signaling pathway in skeletal muscle of KKAy Mice.
Int J Endocrinol. 2014:3124522014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nakka VP, Prakash-Babu P and Vemuganti R:
Crosstalk between endoplasmic reticulum stress, oxidative stress,
and autophagy: Potential Therapeutic targets for acute CNS
injuries. Mol Neurobiol. 53:532–544. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goyal BR and Mehta AA: Diabetic
cardiomyopathy: Pathophysiological mechanisms and cardiac
dysfuntion. Hum Exp Toxicol. 32:571–590. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang ZY, Liu JG, Li H and Yang HM:
Pharmacological effects of active components of chinese herbal
medicine in the treatment of Alzheimer's disease: A review. Am J
Chin Med. 44:1525–1541. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jiang C, Zhu W, Yan X, Shao Q, Xu B, Zhang
M and Gong R: Rescue therapy with Tanshinone IIA hinders transition
of acute kidney injury to chronic kidney disease via targeting
GSK3β. Sci Rep. 6:366982016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wen PY, Li J, Lu BL, Liu J, Yang FZ, Zhou
L, Luo H, Li WW and Zhou J: Tanshinone IIA increases levels of
NeuN, protein disulfide isomerase, and Na+/K+-ATPase and decreases
evidence of microglial activation after cerebral ischemic injury.
Neuroreport. 27:435–444. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun D, Shen M, Li J, Li W, Zhang Y, Zhao
L, Zhang Z, Yuan Y, Wang H and Cao F: Cardioprotective effects of
tanshinone IIA pretreatment via kinin B2 receptor-Akt-GSK-3β
dependent pathway in experimental diabetic cardiomyopathy.
Cardiovasc Diabeto. 10:42011. View Article : Google Scholar
|
|
27
|
Li Q, Shen L, Wang Z, Jiang HP and Liu LX:
Tanshinone IIA protects against myocardial ischemia reperfusion
injury by activating the PI3K/Akt/mTOR signaling pathway. Biomed
Pharmacother. 84:106–114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Slemmer JE, Shacka JJ, Sweeney MI and
Weber JT: Antioxidants and free radical scavengers for the
treatment of stroke, traumatic brain injury and aging. Curr Med
Chem. 15:404–414. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kara A, Unal D, Simsek N, Yucel A, Yucel N
and Selli J: Ultra-structural changes and apoptotic activity in
cerebellum of post-menopausal-diabetic rats: A histochemical and
ultra-structural study. Gynecol Endocrinol. 30:226–231. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang M, Xu Z, Zhang R, Zhang P, Weng Y,
Shen Y and Zhang X: Protection of myocardium in
streptozotocin-induced diabetic rats by water extracts of
Hsian-tsao (Mesona procumbens Hemsl.). Asia Pac J Clin Nutr.
17:23–29. 2008.PubMed/NCBI
|
|
31
|
Liu XY, Liu FC, Deng CY, Zhang MZ, Yang M,
Xiao DZ, Lin QX, Cai ST, Kuang SJ, Chen J, et al: Left ventricular
deformation associated with cardiomyocyte Ca(2+) transients delay
in early stage of low-dose of STZ and high-fat diet induced type 2
diabetic rats. BMC Cardiovasc Disord. 16:412016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kyrychenko V, Poláková E, Janíček R and
Shirokova N: Mitochondrial dysfunctions during progression of
dystrophic cardiomyopathy. Cell Calcium. 58:186–195. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang HP, Zhang WH, Wang XF, Zhu J, Zheng
YQ, Xia Q and Zhi JM: Exposure to AT1 receptor autoantibodies
during pregnancy increases susceptibility of the maternal heart to
postpartum ischemia-reperfusion injury in rats. Int J Mol Sci.
15:11495–11509. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pluquet O, Pourtier A and Abbadie C: The
unfolded protein response and cellular senescence. A review in the
theme: Cellular mechanisms of endoplasmic reticulum stress
signaling in health and disease. Am J Physiol Cell Physiol.
308:C415–C425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ron D and Walter P: Signal integration in
the endoplasmic reticulum unfolded protein response. Nat Rev Mol
Cell Biol. 8:519–529. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sano R and Reed JC: ER stress-induced cell
death mechanisms. Biochim Biophys Acta. 1833:3460–3470. 2013.
View Article : Google Scholar : PubMed/NCBI
|