Introduction
Approximately 250,000 novel central nervous system
(CNS) tumors are reported to occur each year in Europe (1). Glioma is one of the most common types
of malignant cancer and accounts for the majority of CNS
tumor-related mortality (2). Due to
the limited treatment strategies available, the overall 5-year
survival rate for patients with glioma remains poor (3). Therefore, considerable efforts are
required in order to investigate the underlying mechanisms of
glioma progression in order to facilitate the development of novel
therapeutic approaches in malignant glioma.
microRNAs (miRNAs) are endogenous RNAs that can bind
to the 3′-untranslated region (3′-UTR) of target genes (4). Emerging evidence suggests that miRNAs
may serve vital roles in regulating cellular behavior (5,6). Human
cancer progression is often associated with abnormal cellular
behavior, which includes cell proliferation, apoptosis, migration
and invasion (7). Therefore, it may
be important to investigate the roles of miRNAs in several types of
human cancer, including glioma (8,9).
miR-490-3p, a newly identified miRNA, has been previously shown to
function as a tumor suppressor in several types of cancer (10–13). Liu
et al (10) revealed that
miR-490-3p regulates colorectal cancer cell proliferation,
metastasis and apoptosis by targeting the voltage dependent anion
channel 1. Tian et al (11)
observed that miR-490-3p regulates the sensitivity of ovarian
cancer cells to cisplatin by directly targeting ABCC2. Previous
studies also demonstrated that miR-490-3p may be a potential
therapeutic target for osteosarcoma (12,13).
However, the clinical significance of miR-490-3p in glioma remains
unknown.
High-mobility group AT-hook 2 (HMGA2) belongs to the
high-motility group (HMG) protein family and is reported to
function as an oncogene in several types of human cancer (14–16). A
previous study demonstrated that HMGA2 overexpression was
correlated with poor prognosis in patients with lung cancer
(15). In addition, HMGA2
overexpression was revealed to be closely associated with
aggressive cell behaviors of glioma (16). Furthermore, HMGA2 is inversely
regulated by miRNAs in several types of human cancer including
squamous cell lung carcinoma and clear cell renal carcinoma
(17,18). However, whether miRNAs regulate HMGA2
expression in glioma remains unknown.
In the current study, miR-490-3p expression was
significantly downregulated in glioma tissues and cell lines.
Subsequently, the current study demonstrated that miR-490-3p may
regulate glioma cell proliferation and migration in vitro.
HMGA2 was identified as a target gene of miR-490-3p in glioma.
Furthermore, the inhibitory effect of miR-490-3p on glioma cell
proliferation and migration may be partially regulated by HMGA2
expression. Therefore miR-490-3p may be a potential novel target
for the treatment of glioma.
Materials and methods
Clinical samples
Glioma tissue and adjacent noncancerous tissue
samples (>3 cm from cancer tissues) were collected from 24
patients (12 male and 12 female; average age, 59.1±6.3 years; age
range, 42–79 years) with glioma at The First Affiliated Hospital of
Guangzhou University of Traditional Chinese Medicine between April
2014 and May 2016. Tumor stage was classified based on the World
Health Organization (WHO) stage and grading system (19). The Karnofsky performance scale (KPS)
ranking runs from 100 to 0, where 100 is ‘perfect health’ and 0 is
death. Its purpose is to evaluate a patient's ability to survive
chemotherapy treatment (20). All
patients involved in the current study did no receive anti-cancer
treatment prior to the surgery. All tissue samples were stored at
−80°C for further analyses. The study protocol was approved by the
Ethics Committee of The First Affiliated Hospital of Guangzhou
University of Traditional Chinese Medicine (Guangzhou, China).
Written informed consent was obtained from all patients prior to
the study.
Cell culture
Human glioma cell lines A172, T98 and U87-MG (cat.
no. TCHu138, glioblastoma of unknown origin) were purchased from
Cell Bank of the Chinese Academy of Science (Shanghai, China) and
cultured in Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Thermo Fisher
Scientific, Inc.). Normal human astrocytes (NHAs) were purchased
from Lonza Group, Ltd. and cultured in AGM™ Astrocyte Growth Medium
(Lonza Group, Ltd.) supplemented with 10% FBS (Thermo Fisher
Scientific, Inc.). Cells were maintained at 37°C in a humidified
atmosphere containing 5% of CO2.
Cell transfection
miR-490-3p mimic (5′-CAACCUGGAGGACUCCAUGCUG-3′),
inhibitor (5′-CAGCAUGGAGUCCUCCAGGUUG-3′) and negative control miRNA
(NC-miRNA; 5′-ACCGCUAAUCAUACGAAUACAC-3′) were purchased from
Guangzhou RiboBio Co., Ltd. HMGA2 overexpression vector based on
pcDNA3.1 and NC plasmid (pcDNA3.1) was purchased from GenScript
Co., Ltd. (Nanjing, China). Cells (U87-MG and T98) were seeded into
6-well plates at the density of 5×105 cells/well.
Synthetic miRNAs (100 pmol) or HMGA2 plasmid (2 µg) were mixed with
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol and used to transfect
cells for 48 h.
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was extracted from tissue samples and
cells (U87-MG, A172 and T98) using TRIzol® reagent
(Thermo Fisher Scientific, Inc.). Total RNA was reverse transcribed
into first-strand cDNA using the PrimeScript™ RT Reagent kit
(Takara Biotechnology Co., Ltd.), according to the manufacturer's
protocol. qPCR was subsequently performed using an ABI7500 CPR
machine (Applied Biosystems; Thermo Fisher Scientific, Inc.) and
SYBR Premix Ex Taq II (Takara Biotechnology Co., Ltd.). The
following primer pairs were used for the qPCR: miR-490-3p, forward,
5′-TGCGGTTCAAGTAATTCAGGA-3′ and reverse, 5′-CCAGTGCAGGGTCCGAGGT-3′;
and U6 small nuclear (sn)RNA, forward, 5′-TGCGGGTGCTCGCTTCGGCAGC-3′
and reverse, 5′-CCAGTGCAGGGTCCGAGGT-3′. The following thermocycling
conditions were used: 10 min at 95°C; 40 cycles of 1 min at 95°C; 2
min at 63°C; and 1 min at 72°C. Relative miR-490-3p levels were
quantified using the 2−ΔΔCq method and normalized to U6
snRNA, the internal control (21).
Western blotting
Total protein was extracted from tissue samples and
cells (U87-MG and T98) using radioimmunoprecipitation assay buffer
(Beyotime Institute of Biotechnology) and quantified using
bicinchoninic acid kit (Beyotime Institute of Biotechnology). Total
protein (50 µg per lane) was separated by SDS-PAGE on a 12% gel.
The separated proteins were subsequently transferred onto
polyvinylidene difluoride membranes (Thermo Fisher Scientific,
Inc.) and blocked with 5% fat-free milk at 4°C for 3 h. The
membranes were incubated with primary antibodies against HMGA2
(1:1,000; ab109329), matrix metallopeptidase-9 (MMP-9; 1:1,000;
ab73734) and GAPDH (ab181602; 1:1,000; all Abcam) at 4°C for
overnight. Membranes were washed three times with Tris-buffered
saline and Polysorbate 20 (TBST). Following primary incubation,
membranes were incubated with horseradish peroxidase-conjugated
goat anti-rabbit secondary antibody (1:5,000; ab97051; Abcam) at
room temperature for 4 h. Protein bands were visualized using an
BeyoEnhanced Chemiluminescence kit (Beyotime Institute of
Biotechnology) and protein expression was quantified using ImageJ
1.48 software (National Institutes of Health).
Cell proliferation assay
Cell proliferation rate was analyzed using the cell
counting kit-8 (CCK-8; Beyotime Institute of Biotechnology). Cells
(U87-MG and T98) were seeded into 96-well plates at the density of
5×103 cells/well and incubated for 0, 24, 48 and 72 h
prior at 37°C to the addition of 10 ml CCK-8 reagent. Cells were
incubated at 37°C for a further 2 h. Optical density was measured
at a wavelength of 450 nm using microplate reader (Bio-Rad
Laboratories, Inc.). Each sample was performed in triplicate.
Cell invasion assay
Cell invasion was examined using a Transwell
invasion assay. A total of 1×105 cells (U87-MG and T98)
were plated in the upper chamber (8 mm; Corning Inc.) that
pre-coated with Matrigel at 37°C for 30 min (BD Biosciences, San
Jose, CA, USA) in DMEM. DMEM supplemented with 10% FBS (Thermo
Fisher Scientific, Inc.) was plated in the lower chambers.
Following incubation for 24 h, migratory cells in the lower
membrane were fixed with methanol and stained with 0.5% crystal
violet solution at 37°C for 4 h. Stained cells were counted from
five independent fields using an inverted fluorescent microscope
(magnification, ×200; Olympus IX53; Olympus Corporation).
Dual-luciferase assay
Bioinformatic analysis was performed using
TargetScan (www.targetscan.org/vert_72) to predict potential
targets of miR-490-3p. The wild-type (wt) 3′-UTR of HMGA2 was
cloned using the following primers: HMGA2-wt forward,
5′-CCGTCTAGACGGGGGGCGCCAACGTTCGATTTCT-3′ and reverse,
5′-CTGTTTTGACCAAACTTTATTAATAGTTTAAGATCTATTCTTAT-3′. The cloned
3′-UTR fragment of HMGA1 was subsequently inserted into the
XbaI/XbaI sites of pGL3 vector (Promega Corporation).
A site-directed mutagenesis kit (Takara Biotechnology Co., Ltd.)
was used to construct the mutant (mut) 3′-UTR of HMGA2 using the
following primers: HMGA2-mut forward,
5′-AAAAAAGGGGGGGGCAATCTCTCGGTCCAATTTCTCTCTCTCTCTTCCTC-3′ and
reverse, 5′-GAGGAAGAGAGAGAGAGAAATTGGACCGAGAGATTGCCCCCCCCTTTTTT-3′.
For luciferase activity assays, cells (U87-MG and T98) were
co-transfected with wt or mut 3′-UTR HMGA2 and miR-490-3p mimic or
NC-miRNA using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). Luciferase activity was measured 48 h
post-transfection using a Dual-Luciferase® (DLR™)
Reporter Assay System (Promega Corporation) with Renilla
luciferase activity as the internal control.
Statistical analysis
Data were analyzed with SPSS 19.0 (IBM Corp.) and
presented as the mean ± standard deviation. Differences between
groups were analyzed using Wilcoxon signed-rank test or one-way
analysis of variance followed by Tukey's post hoc test. Association
between miR-490-3p expression and clinicopathological parameters
was analyzed by χ2 test. Correlation between miR-490-3p
and HMGA2 was conducted using Pearson's correlation co-efficient.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-490-3p expression is downregulated
in glioma tissues and cell lines
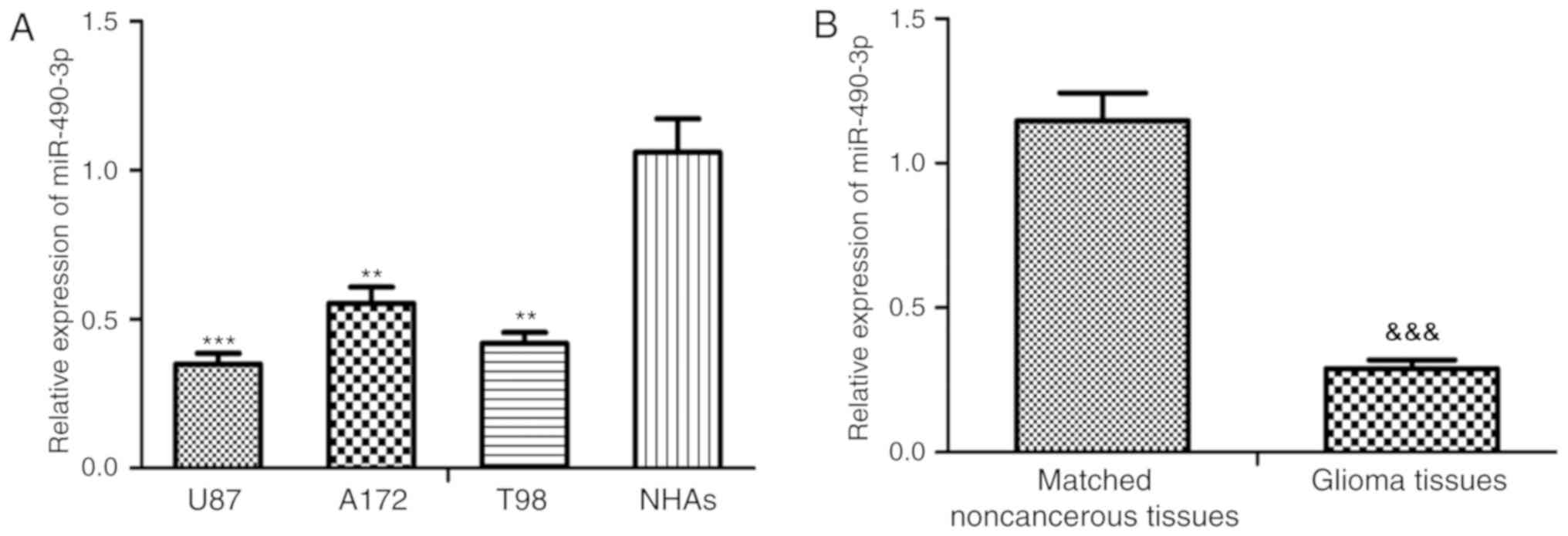
The relative miR-490-3p expression level was
significantly downregulated in all glioma cell lines, U87-MG, A172
and T98, compared with the NHAs (P<0.01 and P<0.001; Fig. 1A). In addition, the U87-MG and T98
cells lines demonstrated the greatest decrease and therefore the
U87-MG and T98 cells lines were selected for subsequent
experimentation. Furthermore, the relative miR-490-3p expression
level was significantly downregulated in glioma tissue compared
with adjacent noncancerous tissue samples (P<0.001; Fig. 1B). Patients with glioma were
classified into a high (n=9) or low (n=15) miR-490-3p expression
group based on the relative miR-490-3p levels (cut-off value:
0.27). The current study demonstrated that low miR-490-3p
expression was closely associated with WHO grade (P=0.031) and KPS
(P=0.014) in patients with glioma, however, there was no
significant association observed between miR-490-3p expression with
age or sex (Table I).
 | Table I.Association between miR-490-3p
expression and clinicopathological features of patients with
glioma. |
Table I.
Association between miR-490-3p
expression and clinicopathological features of patients with
glioma.
|
|
| miR-490-3p
expression |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
features | No. of cases | Low (n=15) | High (n=9) | P-value |
|---|
| Age, years |
|
|
|
|
|
>60 | 12 | 9 | 3 | 0.083 |
|
<60 | 12 | 6 | 6 |
|
| Sex |
|
|
|
|
| Male | 12 | 8 | 4 | 0.558 |
|
Female | 12 | 7 | 5 |
|
| KPS |
|
|
|
|
|
>90 | 7 | 5 | 2 | 0.014 |
|
<90 | 17 | 10 | 7 |
|
| WHO grade |
|
|
|
|
|
I–II | 8 | 4 | 4 | 0.031 |
|
III | 16 | 11 | 5 |
|
miR-490-3p directly regulates of HMGA2
expression
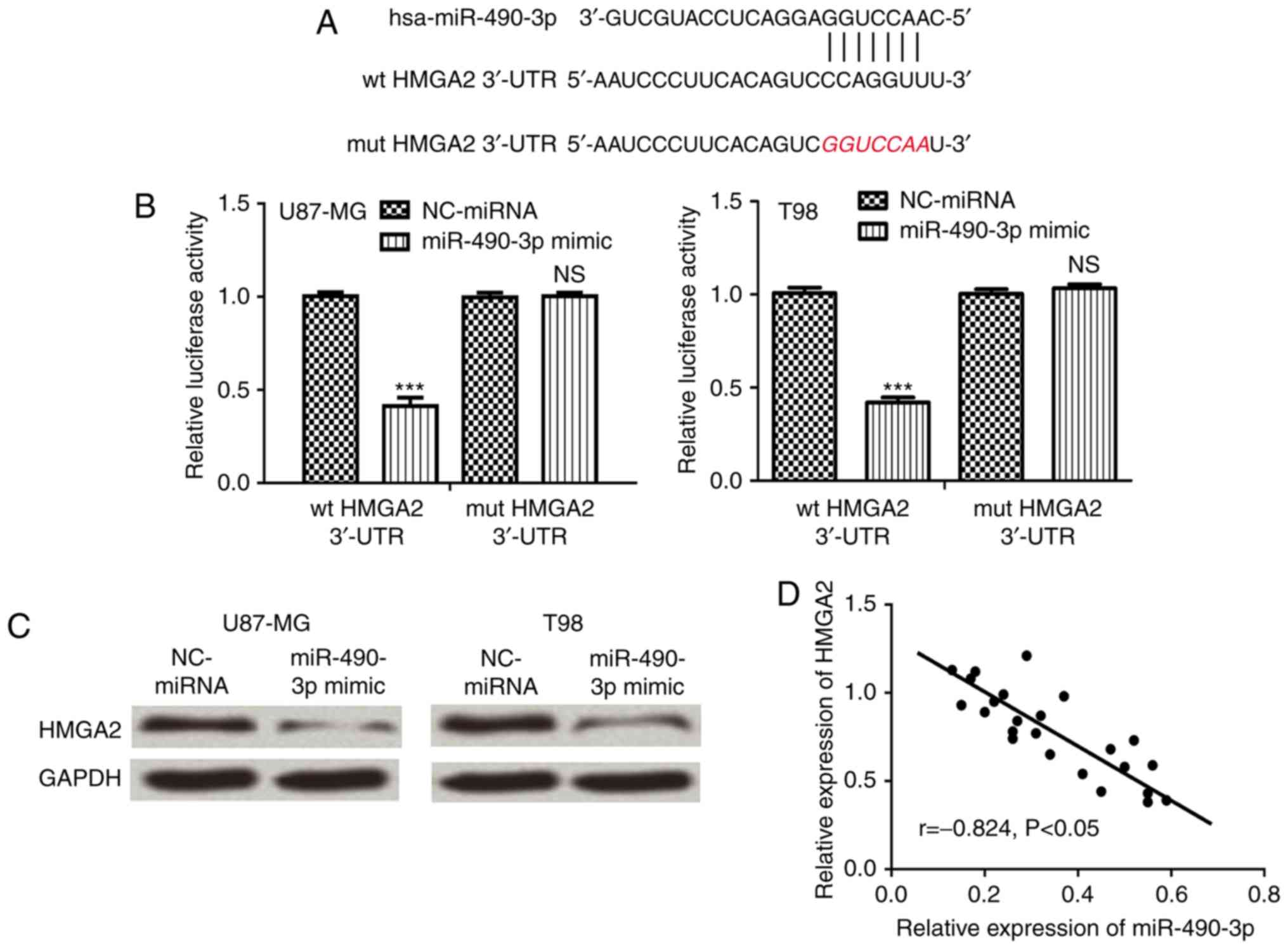
Bioinformatics analysis was performed to predict
target genes of miR-490-3p. HMGA2 was identified as a putative
target gene of miR-490-3p and bioinformatics analysis was used to
predict the miR-490-3p target sequence in the 3′-UTR of HMGA2
(Fig. 2A). Luciferase reporter
assays demonstrated that miR-490-3p overexpression significantly
reduced wt HMGA2 luciferase activity compared with mut HMGA2 3′-UTR
(P<0.001), which did not affect luciferase activity (Fig. 2B). Furthermore, HMGA2 protein
expression was analyzed in cells following transfection with
miR-490-3p mimic or NC-miRNA. The relative HMGA2 protein expression
level was markedly reduced in cells following transfection with
miR-490-3p mimic compared with NC-miRNA (Fig. 2C). Furthermore, correlation analysis
demonstrated that there was a negative correlation between
miR-490-3p and HMGA2 expression in glioma tissues (Fig. 2D).
miR-490-3p suppresses glioma cell
proliferation and invasion
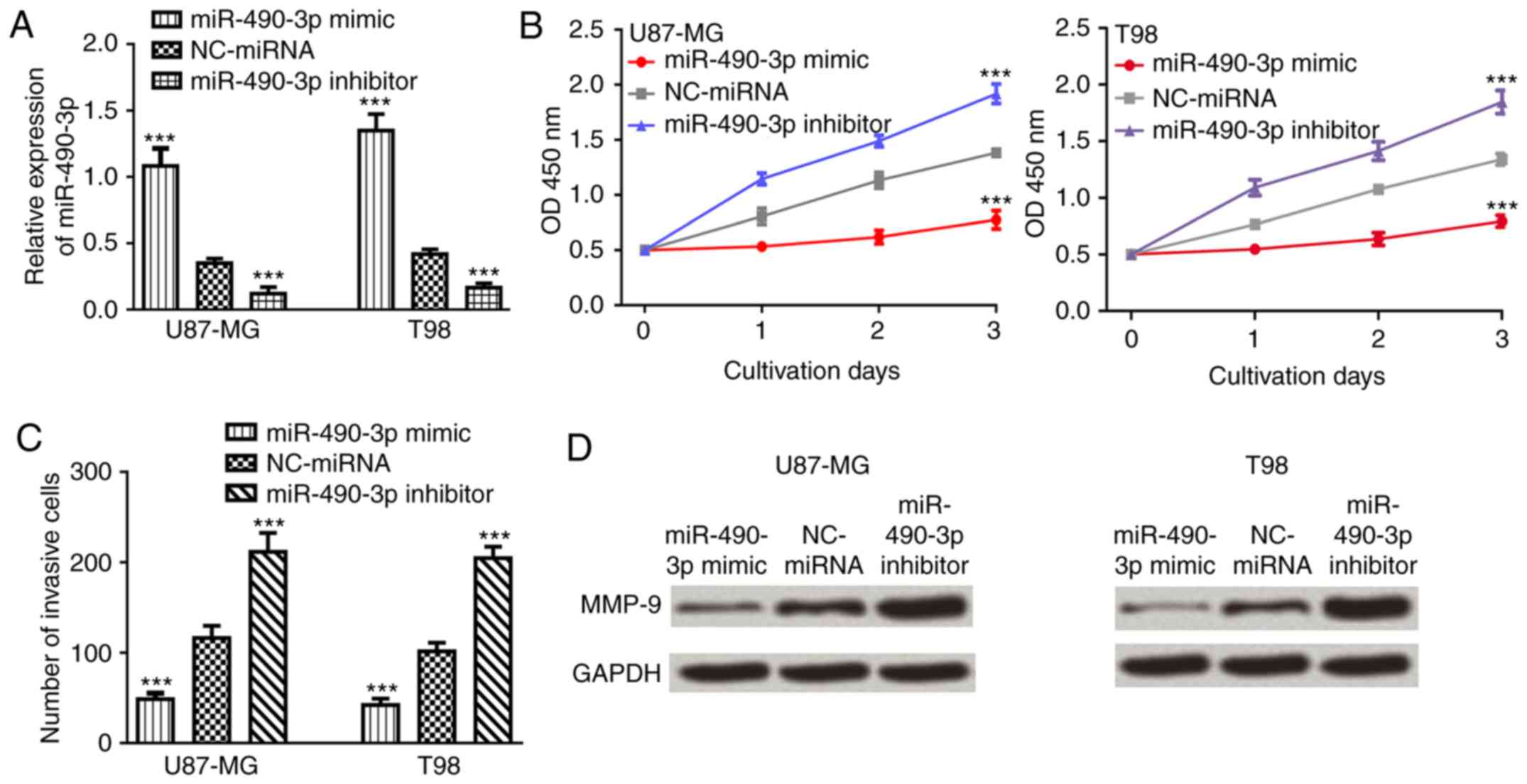
To investigate the role of miR-490-3p in glioma,
CCK-8 and Transwell invasion assays were used to examine the effect
of miR-490-3p on glioma cell proliferation and invasion following
transfection with miR-490-3p mimic, NC-miRNA or miR-490-3p
inhibitor. The relative miR-490-3p expression level was
significantly increased in glioma cell lines following transfection
with miR-490-3p mimic compared with NC-miRNA (P<0.001; Fig. 3A). By contrast, the relative
miR-490-3p expression level was significantly decreased in glioma
cell lines following transfection with miR-490-3p inhibitor
(P<0.001; Fig. 3A). Cell
proliferation and invasion were significantly decreased in glioma
cells following transfection with miR-490-3p mimic compared with
NC-miRNA (P<0.001; Fig. 3B and
C). Furthermore, the expression level of MMP-9, an invasion
marker (22), was examined in glioma
cell lines following transfection with miR-490-3p mimic, NC-miRNA
or miR-490-3p inhibitor. The relative protein expression level of
MMP-9 was markedly decreased following transfection with miR-490-3p
mimic, while relative protein expression level of MMP-9 was
markedly increased following transfection with miR-490-3p inhibitor
compared with NC-miRNA (Fig.
3D).
Overexpression of HMGA2 partially
reverses the inhibitory effect of miR-490-3p on glioma cell
proliferation and invasion
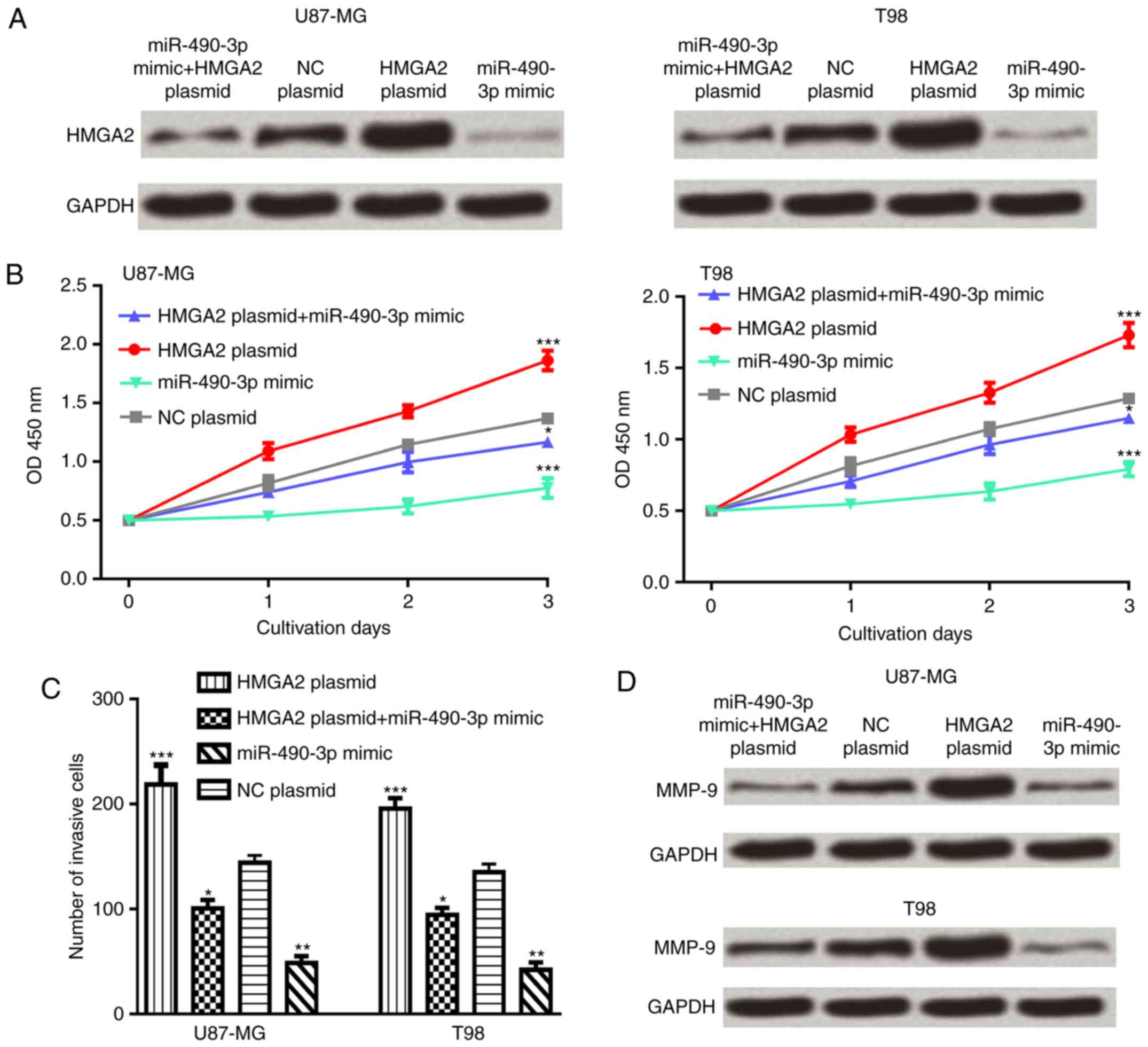
To investigate whether HMGA2 was an effector for
miR-490-3p, glioma cell lines were co-transfected with the HMGA2
expression plasmid and miR-490-3p mimic. The results demonstrated
that overexpression of HMGA2 partially reversed the inhibitory
effect of miR-490-3p on the HMGA2 protein expression level in
glioma cells (Fig. 4A). In addition,
overexpression of HMGA2 partially reversed the inhibitory effect of
miR-490-3p on glioma cell proliferation and invasion (Fig. 4B and C). Furthermore, the protein
expression level of MMP-9 was increased following co-transfection
with the HMGA2 expression plasmid and miR-490-3p mimic compared
with the miR-490-3p mimic alone (Fig.
4D).
Discussion
Approximately 60% of human genes are thought to be
regulated by miRNAs, which suggests that miRNAs may be involved in
multiple cellular processes (23).
Numerous miRNAs have been identified to be critical players in the
progression of glioma (9,24,25). For
example, miR-30b-5p overexpression significantly inhibited glioma
cell proliferation in vitro by downregulating the expression
of metadherin (24). Ding et
al (25) demonstrated that
miR-122 expression was downregulated in glioma tissues. In
addition, miR-122 may function as tumor suppressor through
targeting runt-related transcription factors (25).
The current study demonstrated that miR-490-3p
expression was significantly downregulated in glioma tissue
compared with adjacent noncancerous tissue samples. Furthermore,
the current study demonstrated that low miR-490-3p expression was
closely associated with advanced tumor grade and high KPS scores in
patients with glioma. Taken together, these results suggest that
miR-490-3p may act as a tumor suppressor in glioma and decreased
miR-490-3p expression may be associated with tumor progression.
In the current study, the biological role of
miR-490-3p in glioma was further investigated. The results
demonstrated that ectopic expression of miR-490-3p suppresses
glioma cell proliferation and invasion in vitro, while the
opposite effects were observed following downregulation of
miR-490-3p. Furthermore, the protein expression level of MMP-9 in
glioma cell lines was increased following transfection with
miR-490-3p inhibitor, while the protein expression level of MMP-9
in glioma cell lines was decreased following transfection with
miR-490-3p mimic. Previous studies demonstrated that miR-490-3p has
different targets in several types of cancer (10,11).
To examine the underlying mechanism of miR-490-3p in
glioma cell proliferation and invasion, bioinformatics analysis was
used to identify target genes of miR-490-3p. In the current study,
luciferase reporter assays and western blot analysis confirmed
HMGA2 as a direct target of miR-490-3p in glioma. HMGA2 was
previously reported to be involved in the regulation of cellular
behavior in several types of human cancer (15–18). In
addition, HMGA2 was recently identified to be dispensable for
pancreatic cancer progression, metastasis and therapy resistance
(26). Hawsawi et al
(27) revealed that HMGA2 may
promote epithelial-mesenchymal transition of prostate cancer via
the mitogen-activated protein kinase pathway. In the current study,
functional assays demonstrated that HMGA2 overexpression partially
reversed the inhibitory effect of miR-490-3p on the malignant
phenotype of glioma cells, which suggests that miR-490-3p may
function as a tumor suppressor in glioma by targeting HMGA2.
Several studies demonstrated that HMGA2 overexpression promotes
stemness, invasion and tumorigenicity in glioma (16,28). A
previous study revealed that the miRNA let-7a may target HMGA2
expression to regulate glioma cell proliferation, invasion and
metastasis via the transforming growth factor-β/Smad3 signaling
pathway (29). Furthermore, the
expression levels of let-7a and HMGA2 were correlated with glioma
grades (29). These results suggest
that targeting HMGA2 may have therapeutic benefits in glioma
therapy. A previous study revealed an inverse correlation between
miR-490-3p and HMGA2 expression and miR-490-3p may regulate cell
proliferation and apoptosis in osteosarcoma (13). In the current study, an inverse
correlation between miR-490-3p and HMGA2 expression was observed in
glioma, which confirmed the results from previous studies.
Furthermore, the current study revealed the inhibitory effect of
miR-490-3p on cell proliferation and invasion. Taken together,
these results demonstrate the importance of the miR-490-3p/HMGA2
axis, which may be valuable in understanding the roles of
miR-490-3p and HMGA2 in different types of human cancer.
In conclusion, the current study demonstrated that
miR-490-3p serves an important role in regulating glioma cell
proliferation and invasion. In addition, a correlation between
miR-490-3p and HMGA2 expression in glioma was identified, which may
be valuable in understanding the underlying mechanisms of glioma
progression. Taken together, these results suggest that the
miR-490-3p/HMGA2 axis may be a potential therapeutic target for
glioma.
Acknowledgements
Not applicable.
Funding
This work was supported by the Foundation of the
Science and Technology Program of Guangzhou, People's Republic of
China (grant no. 201607010365).
Availability of data and materials
The datasets generated/analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FZ, AW, YW and JL designed the study, performed the
experiments, analyzed the data and wrote the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The current study was approved by the Ethics
Committee of The First Affiliated Hospital of Guangzhou University
of Traditional Chinese Medicine (Guangzhou, China). All patients
provided written informed consent prior to the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Crocetti E, Trama A, Stiller C, Caldarella
A, Soffietti R, Jaal J, Weber DC, Ricardi U, Slowinski J and
Brandes A; RARECARE working group, : Epidemiology of glial and
non-glial brain tumours in Europe. Eur J Cancer. 48:1532–1542.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chhabda S, Carney O, D'Arco F, Jacques TS
and Mankad K: The 2016 world health organization classification of
tumours of the central nervous system: What the paediatric
neuroradiologist needs to know. Quant Imaging Med Surg. 6:486–489.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mittal S, Pradhan S and Srivastava T:
Recent advances in targeted therapy for glioblastoma. Expert Rev
Neurother. 15:935–946. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Banelli B, Forlani A, Allemanni G,
Morabito A, Pistillo MP and Romani M: MicroRNA in glioblastoma: An
overview. Int J Genomics. 2017:76390842017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu X, He B, Xu T, Pan Y, Hu X, Chen X and
Wang S: MiR-490-3p functions as a tumor suppressor by inhibiting
oncogene VDAC1 expression in colorectal cancer. J Cancer.
9:1218–1230. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tian J, Xu YY, Li L and Hao Q: MiR-490-3p
sensitizes ovarian cancer cells to cisplatin by directly targeting
ABCC2. Am J Transl Res. 9:1127–1138. 2017.PubMed/NCBI
|
|
12
|
Tang B, Liu C, Zhang QM and Ni M:
Decreased expression of miR-490-3p in osteosarcoma and its clinical
significance. Eur Rev Med Pharmacol Sci. 21:246–251.
2017.PubMed/NCBI
|
|
13
|
Liu W, Xu G, Liu H and Li T:
MicroRNA-490-3p regulates cell proliferation and apoptosis by
targeting HMGA2 in osteosarcoma. FEBS Lett. 589:3148–3153. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hammond SM and Sharpless NE: HMGA2,
microRNAs and stem cell aging. Cell. 135:1013–1016. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sarhadi VK, Wikman H, Salmenkivi K, Kuosma
E, Sioris T, Salo J, Karjalainen A, Knuutila S and Anttila S:
Increased expression of high mobility group A proteins in lung
cancer. J Pathol. 209:206–212. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu B, Pang B, Hou X, Fan H, Liang N,
Zheng S, Feng B, Liu W, Guo H, Xu S and Pang Q: Expression of high
mobility group AT-hook protein 2 and its prognostic significance in
malignant gliomas. Hum Pathol. 45:1752–1758. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu L, Du B, Lu QJ, Fan XW, Tang K, Yang L
and Liao WL: miR-541 suppresses proliferation and invasion of
squamous cell lung carcinoma cell lines via directly targeting
high-mobility group AT-hook 2. Cancer Med. 7:2581–2591. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao H, Zhao H, Xia X and Liu X:
MicroRNA-599 targets high-mobility group AT-hook 2 to inhibit cell
proliferation and invasion in clear cell renal carcinoma. Mol Med
Rep. 17:7451–7459. 2018.PubMed/NCBI
|
|
19
|
Komori T: The 2016 WHO classification of
tumours of the central nervous system: The major points of
revision. Neurol Med Chir (Tokyo). 57:301–311. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karnofsky DA and Burchenal JH: Evaluation
of chemotherapeutic agents. In: The Clinical Evaluation of
Chemotherapeutic Agents in Cancer. MacLeod CM (ed). Columbia
University Press; New York, NY: pp. 191–205. 1949
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang D, Liu Z, Zheng N, Wu H, Zhang Z and
Xu J: MiR-30b-5p modulates glioma cell proliferation by direct
targeting MTDH. Saudi J Biol Sci. 25:947–952. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ding CQ, Deng WS, Yin XF and Ding XD:
MiR-122 inhibits cell proliferation and induces apoptosis by
targeting runt-related transcription factors 2 in human glioma. Eur
Rev Med Pharmacol Sci. 22:4925–4933. 2018.PubMed/NCBI
|
|
26
|
Chiou SH, Dorsch M, Kusch E, Naranjo S,
Kozak MM, Koong AC, Winslow MM and Grüner BM: Hmga2 is dispensable
for pancreatic cancer development, metastasis, and therapy
resistance. Sci Rep. 8:140082018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hawsawi O, Henderson V, Burton LJ, Dougan
J, Nagappan P and Odero-Marah V: High mobility group A2 (HMGA2)
promotes EMT via MAPK pathway in prostate cancer. Biochem Biophys
Res Commun. 504:196–202. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kaur H, Ali SZ, Huey L, Hütt-Cabezas M,
Taylor I, Mao XG, Weingart M, Chu Q, Rodriguez FJ, Eberhart CG and
Raabe EH: The transcriptional modulator HMGA2 promotes stemness and
tumorigenicity in glioblastoma. Cancer Lett. 377:55–64. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Y, Zhang X, Chen D and Ma C: Let-7a
suppresses glioma cell proliferation and invasion through
TGF-β/Smad3 signaling pathway by targeting HMGA2. Tumour Biol.
37:8107–8119. 2016. View Article : Google Scholar : PubMed/NCBI
|