Introduction
Colorectal cancer (CRC) is one of the most common
types of human malignancy worldwide (1). In recent decades, the combined
treatment for CRC has improved, which includes surgical resection,
radiotherapy and chemoradiotherapy; however, the prognosis of
patients with advanced CRC remains poor (2). It has been demonstrated that CRC is
driven by a series of successive genetic and epigenetic changes,
which cause malignant transformation and metastasis (3–5).
Non-coding RNAs are important epigenetic factors associated with
cancer development (6–10), but the functions of various
non-coding RNAs in CRC remain to be fully elucidated.
Long non-coding RNAs (lncRNAs), a class of
non-coding RNAs longer than 200 nucleotides, are 10-fold lower in
abundance than mRNAs (11). It has
been demonstrated that there are thousands of lncRNA small
nucleolar RNA host gene 12 (SNHG12) sequences in mammals and that
the majority of lncRNAs are likely to be functional; however, only
a small proportion of lncRNAs have been studied (12). Through regulation of gene expression
at transcriptional, post-transcriptional and epigenetic levels,
lncRNAs have important roles in various physiological and
pathological processes, including cell growth, proliferation,
differentiation, apoptosis and tumourigenesis (13,14).
Numerous lncRNAs have been reported to function as promoters during
cancer development and progression (15,16).
Therefore, these lncRNAs may become promising biomarkers and/or
therapeutic targets for various cancer types.
Recently, several studies have indicated that the
lncRNA SNHG12 functions as an oncogene in certain, common cancer
types, including osteosarcoma, hepatocellular carcinoma, breast
cancer, lung cancer, gastric cancer, glioma, cervical cancer,
papillary thyroid carcinoma and CRC (17–25). For
instance, Lei et al (20)
reported that SNHG12 promoted the proliferation and migration of
glioma cells by binding to ELAV like RNA binding protein 1. Ding
et al (21) indicated that
SNHG12 enhanced the proliferation and metastasis of papillary
thyroid carcinoma cells by regulating the Wnt/β-catenin signalling
pathway. Furthermore, Wang et al (17) reported that SNHG12 promotes the
proliferation and inhibits apoptosis of CRC cells. However, the
regulatory mechanisms of how SNHG12 affects CRC growth and
metastasis have remained elusive.
MicroRNAs (miRs), a class of small non-coding RNAs
of 22–25 nt in length, are key regulators of gene expression
(26). They directly bind to the
3′-untranslated region (3′-UTR) of their target mRNAs to thereby
cause translational repression or mRNA degradation (26). A large number of studies have
reported that miRs have key roles in the development and
progression of human cancer types, including CRC (27,28). In
recent years, Qian et al (29) reported that miR-16 was significantly
downregulated in CRC tissues compared with that in normal colonic
mucosa, and miR-16 expression was associated with tumour
differentiation, lymph node metastasis, TNM stage and tumour
recurrence in CRC. Furthermore, low miR-16 expression is an
independent factor predicting a poor prognosis for CRC patients
(29). In addition, miR-16 has been
indicated to inhibit CRC cell growth in vitro by regulating
the p53-survivin signalling pathway (30). However, the association between
SNHG12 and miR-16 in CRC has remained elusive.
Therefore, the present study was performed to
explore the association of SNHG12 and miR-16 in CRC and the
underlying molecular mechanisms in CRC cells in vitro.
Materials and methods
Clinical tissue samples
A total of 53 CRC tissues and their matched adjacent
non-tumour tissues were obtained from primary CRC patients at the
Second Xiangya Hospital, Central South University (Changsha, China)
between April 2011 and June 2012. These 53 CRC patients included 31
males and 22 females aged between 34 to 71 years (mean age, 53.7
years). The tissue samples were immediately snap-frozen in liquid
nitrogen after surgical resection and stored until use.
Cell culture
Normal intestinal epithelial NCM460 cells and CRC
cell lines, including HT-29, SW480, SW620 and LOVO, were obtained
from the Cell Bank of Central South University and were cultured in
Dulbecco's modified Eagle's medium (Thermo Fisher Scientific, Inc.)
with 10% foetal bovine serum (FBS; HyClone; GE Healthcare) in a
humidified atmosphere with 5% CO2 at 37°C.
Cell transfection
For cell transfection, SW620 and HT-29 cells
(1×106 per well) were seeded into 6-well plates,
cultured overnight and transfected with 100 nM non-specific short
interfering (si)RNA (NC siRNA; cat. no. 12935300; Thermo Fisher
Scientific, Inc.), 100 nM SNHG12-specific siRNA (SNHG12 siRNA; cat.
no. AM16708; Thermo Fisher Scientific, Inc.), 100 nM pcDNA3.1
vector (cat. no. V00008; Yearthbio, Inc.), 100 nM pcDNA-SNHG12
expression plasmid (cat. no. P00951; Yearthbio, Inc.), 100 nM
miR-16 inhibitor (cat. no. 4464084; Thermo Fisher Scientific, Inc.)
and/or 100 nM negative control (NC) inhibitor (cat. no. 4464076;
Thermo Fisher Scientific, Inc.) by using Lipofectamine®
2000 (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The subsequent experiments were performed
a 48 h after transfection.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from tissues and cells using
TRIzol reagent (Thermo Fisher Scientific, Inc). To detect the
expression of miR-16 or SNHG12, RNA was reverse-transcribed into
complementary (c)DNA using the PrimeScript™ II 1st Strand cDNA
Synthesis kit (Takara Bio, Inc.) according to the manufacturer's
protocol. Subsequently, qPCR was performed using the SYBR Premix
kit (Takara Bio, Inc.) or the SYBR Prime Script miRNA RT-PCR kit
(Takara Bio, Inc.). U6 and GAPDH were used as internal references.
The relative expression was determined using the 2−∆∆Cq
method (31). The following primers
were used: GAPDH forward, 5′-CTGGGCTACACTGAGCACC-3′ and reverse,
5′-AAGTGGTCGTTGAGGGCAATG-3′; and SNHG16 forward,
5′-GTGCCTCAGGAAGTCTCTTGCC-3′ and reverse,
5′-ATCCAAACAAGTTATCACACAGCAC-3′. Primers for U6 (cat. no.
HmiRQP9001) and miR-16 (cat. no. HmiRQP0226) were obtained from
Guangzhou FulenGen, Co., Ltd.
Bioinformatics analysis and luciferase
reporter gene assay
TargetScan (www.targetscan.org) software predicted that SNHG12 was
a potential target gene of miR-16. SW620 and HT-29 cells were
transfected with a wild-type (WT) or a mutant (MT) SNHG12
pMIR-REPORT™ miRNA Expression Reporter Vector (Ambion; Thermo
Fisher Scientific, Inc.) containing a 4-bp mutation at the
predicted miR-16 binding site within SNHG12, along with miR-16
mimics (cat. no. 4464066; Thermo Fisher Scientific, Inc.) or miR-NC
mimics (cat. no. 4464058; Thermo Fisher Scientific, Inc.) using
Lipofectamine 2000. After transfection for 48 h, a luciferase
reporter gene assay was performed using the
Dual-Luciferase® Reporter Assay System (Promega
Corporation). The activity of firefly luciferase was normalized to
the activity of Renilla luciferase.
Cell Counting Kit (CCK)-8 assay
The transfected cells (2,000 cells/well) were seeded
in 96-well plates and cultured at 37°C with 5% CO2 for
0, 24, 48, 72 and 96 h, respectively. The cell proliferation was
then assessed using a CCK-8 (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The optical density value
was read at a wavelength of 450 nm using a microplate reader
(Bio-Rad Laboratories, Inc.).
Cell invasion assay
Cell invasion was assessed using Transwell chambers
pre-coated with Matrigel® (BD Biosciences). The
transfected cells (1×105 per chamber) were re-suspended
in 300 µl serum-free DMEM and added to the upper chamber, and the
lower chamber was filled with medium containing 10% FBS. After
incubation at 37°C for 48 h, cells that had transgressed through
the membrane were fixed with 37% methanol at room temperature for
30 min, and stained with 0.5% crystal violet at room temperature
for 10 min. The invaded cells were counted under a microscope.
Statistical analysis
Values are expressed as the mean ± standard
deviation. SPSS 19.0 (IBM Corp.) was used for statistical analysis.
An unpaired or paired Student's t-test was used for comparison
between two groups. One-way analysis of variance followed by
Tukey's post-hoc test was used for comparisons of more than two
groups. A chi-square test was used to analyse the association
between SNHG12 expression and clinicopathological characteristics
in patients with CRC. Pearson correlation analysis was used to
determine the association between miR-16 and SNHG12 expression in
CRC tissues. The Kaplan-Meier method and a log-rank test were used
to analyse the overall survival of patients with CRC. P<0.05 was
considered to indicate statistical significance.
Results
Upregulation of SNHG12 in CRC tissues
and cell lines
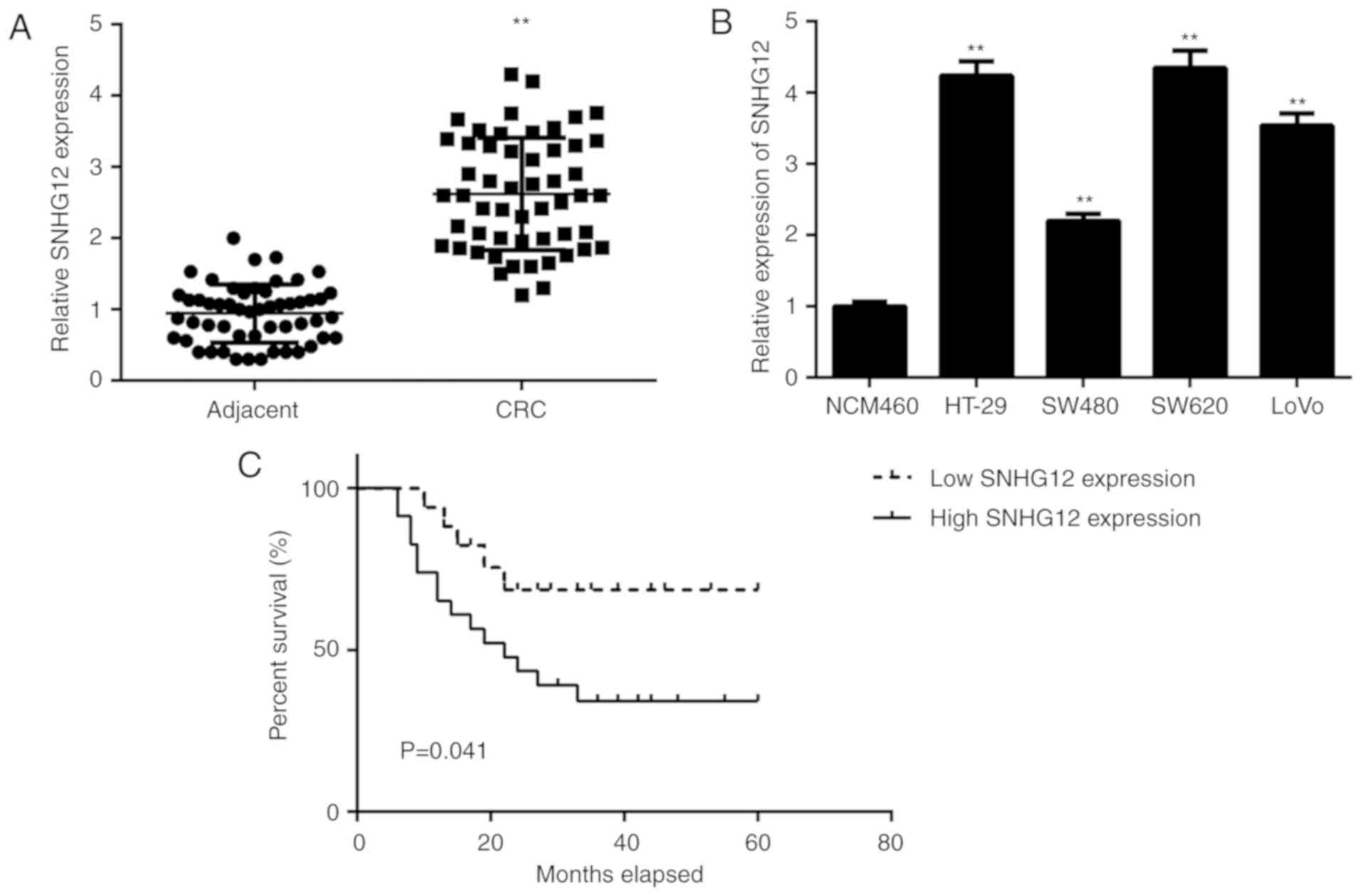
First, the expression of lncRNA SNHG12 in CRC and
adjacent non-tumour tissue samples was examined using RT-qPCR. As
indicated in Fig. 1, the levels of
SNHG12 were significantly higher in CRC tissues compared with those
in adjacent non-tumour tissue samples. Consistently, the expression
levels of SNHG12 were also markedly increased in CRC cell lines,
including HT-29, SW480, SW620 and LOVO, when compared with those in
NCM460 cells (Fig. 1B).
To further assess the clinical significance of
SNHG12 expression in CRC, the CRC patients were divided into high
and low SNHG12 expression groups using the mean expression level of
SNHG12 as the cut-off value. As presented in Table I, high expression of SNHG12 was
significantly associated with lymph node metastasis and advanced
clinical stage. Furthermore, the CRC patients with high SNHG12
expression had a poorer overall survival when compared with that in
the group with low SNHG12 expression (Fig. 1C), suggesting that high SNHG12
expression may predict poor prognosis for CRC patients.
 | Table I.Association between SNHG12 expression
and clinicopathological characteristics in patients with colorectal
cancer (n=53). |
Table I.
Association between SNHG12 expression
and clinicopathological characteristics in patients with colorectal
cancer (n=53).
|
|
| SNHG12
expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Variable | N | Low (n=27) | High (n=26) | Chi-square value | P-value |
|---|
| Age (years) |
|
|
| 0.930 | 0.335 |
| ≤55 | 27 | 12 | 15 |
|
|
|
>55 | 26 | 15 | 11 |
|
|
| Sex |
|
|
| 0.195 | 0.442 |
|
Male | 31 | 15 | 16 |
|
|
|
Female | 22 | 12 | 10 |
|
|
|
Differentiation |
|
|
| 2.163 | 0.141 |
| Well
and moderate | 43 | 24 | 19 |
|
|
|
Poor | 10 | 3 | 7 |
|
|
| Lymph node
metastasis |
|
|
| 2.419 | 0.016 |
|
Present | 18 | 5 | 13 |
|
|
|
Absent | 35 | 22 | 13 |
|
|
| Tumor, nodes and
metastasis stage |
|
|
| 2.419 | 0.016 |
|
I–II | 35 | 22 | 13 |
|
|
|
III–IV | 18 | 5 | 13 |
|
|
Function of SNHG12 in CRC cell
proliferation and invasion
Next, the function of SNHG12 in the proliferation
and invasion of CRC cells was examined. As the above results had
indicated that SNHG12 was upregulated in CRC, SW620 and HT-29 cells
were transfected with SNHG12 siRNA to reduce its expression. After
transfection, the expression of SNHG12 was markedly downregulated
in the siSNHG12 group when compared with that in the siNC group
(Fig. 2A). A CKK-8 assay indicated
that the proliferation of SW620 and HT-29 cells was significantly
decreased in the siSNHG12 group compared with that in the siNC
group, indicating that knockdown of SNHG12 expression inhibits CRC
cell proliferation (Fig. 2B and C).
Similarly, the Transwell assay suggested that the invasive capacity
of SW620 and HT-29 cells was markedly decreased after silencing of
SNHG12 expression (Fig. 2D).
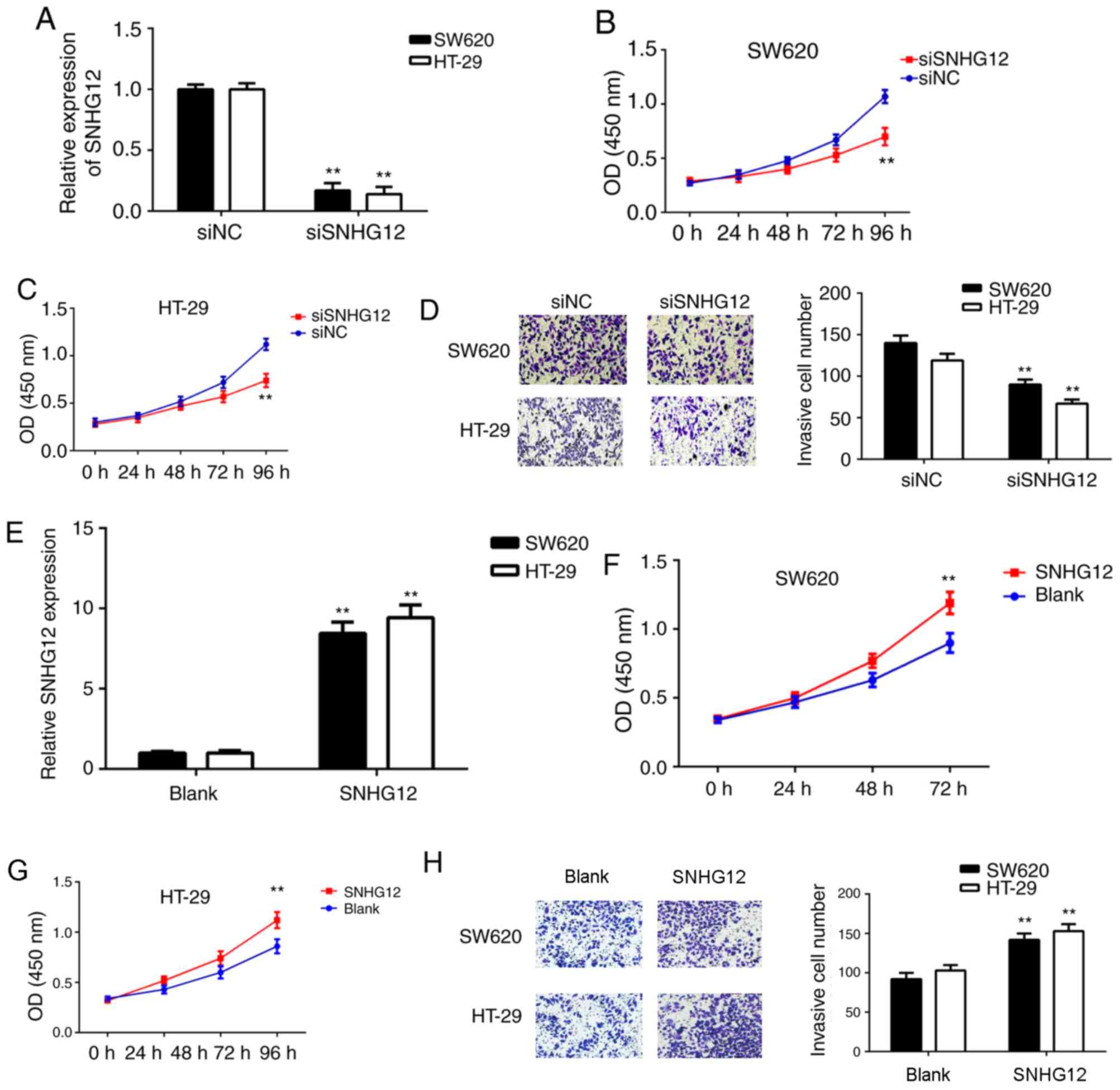 | Figure 2.Inhibition of SNHG12 inhibits
colorectal cancer cell proliferation and invasion. (A-D) SW620 and
HT-29 cells were transfected with SNHG12 siRNA or NC siRNA,
respectively. (A) RT-qPCR was used to examine the levels of SNHG12.
CCK-8 assay using (B) SW620 and (C) HT-29 cells, and (D) a
Transwell assay (magnification, ×200) were performed to assess cell
proliferation and invasion, respectively. For A-D, **P<0.01 vs.
NC siRNA. (E-H) Next, SW620 and HT-29 cells were transfected with
SNHG12 expression plasmid or a blank vector, respectively. (E)
RT-qPCR was used to determine the levels of SNHG12. CCK-8 assay
using (F) SW620 and (G) HT-29 cells, and (H) a Transwell assay
(magnification, ×200) were performed to study cell proliferation
and invasion. For E-H, **P<0.01 vs. Blank. Groups: siNC, cells
transfected with NC siRNA; siSNHG12, cells transfected with siRNA
targeting SNHG12; SNHG12, cells transfected with SNHG12
overexpression vector; Blank, cells transfected with empty vector.
SNHG12, small nucleolar RNA host gene 12; CKK-8, Cell Counting Kit
8; siRNA, small interfering RNA; NC, negative control; OD, optical
density; RT-qPCR, reverse transcription-quantitative polymerase
chain reaction. |
To further confirm the above results, the effects of
SNHG12 overexpression on the proliferation and invasion of CRC
cells were then assessed. SW620 and HT-29 cells were transfected
with SNHG12 plasmid or blank vector. After transfection, SNHG12 was
significantly upregulated in the SNHG12 group when compared with
that in the blank vector group (Fig.
2E). Furthermore, as indicated in Fig. 2F-H, the proliferation and invasion of
SW620 and HT-29 cells was significantly increased in the SNHG12
group compared with that in the blank vector group. These results
suggest that SNHG12 may have a role in promoting CRC growth and
metastasis.
Targeting association between SNHG12
and miR-16 in CRC cells
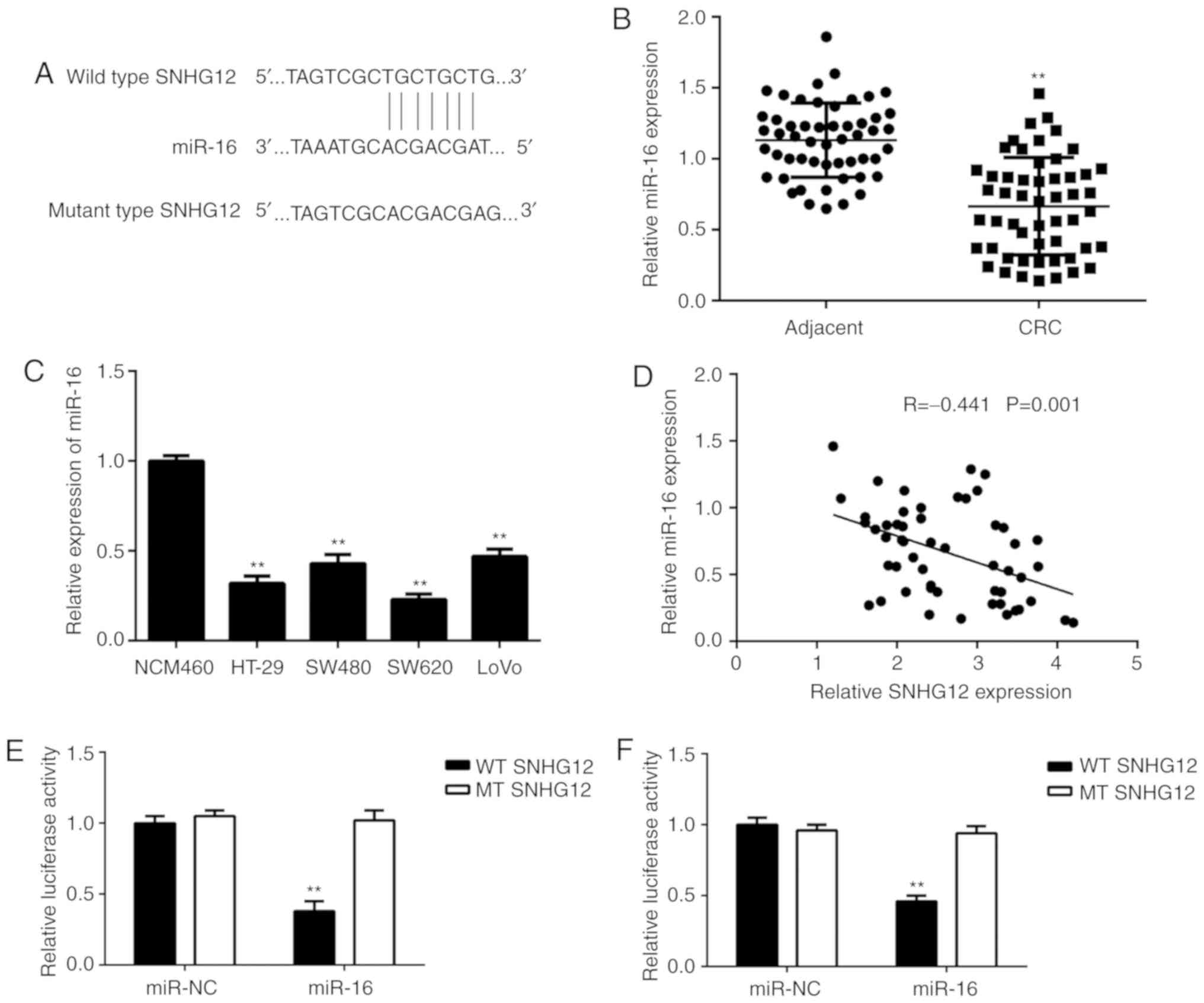
Next, a Bioinformatics analysis was performed to
identify miRs targeted by SNHG12, and it was indicated that miR-16
and SNHG12 had complementary binding sites (Fig. 3A). Therefore, the expression levels
of miR-16 in CRC tissue and cell lines were examined. As indicated
in Fig. 3B and C, the expression
levels of miR-16 were markedly reduced in CRC tissues and cell
lines when compared with those in adjacent non-tumour tissues and
NCM460 cells, respectively. Furthermore, a significant inverse
correlation was observed between SNHG12 and miR-16 expression in
CRC tissues (Fig. 3D). To further
confirm the association between SNHG12 and miR-16 in CRC,
luciferase reporter plasmids containing WT or MT miR-16 binding
sites of SNHG12 were constructed (Fig.
3E). The luciferase reporter assay indicated that transfection
with miR-16 mimics markedly decreased the luciferase activity of
the vector containing WT SNHG12 but had no effect on that in the
vector carrying MT SNHG12 in SW620 and HT-29 cells (Fig. 3F-G), suggesting that SNHG12 acts as a
sponge for miR-16 in CRC cells.
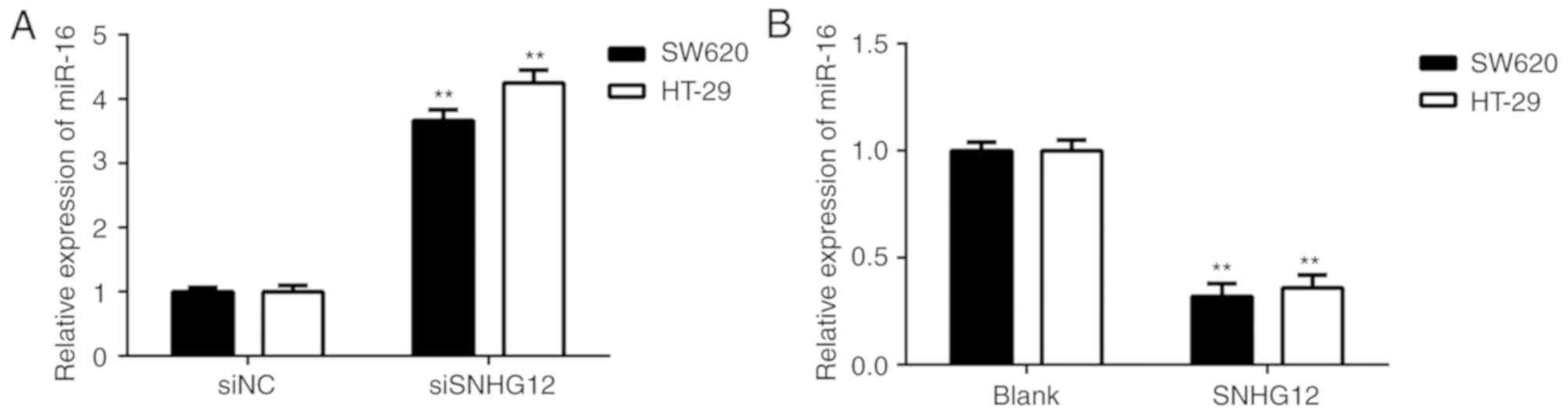
SNHG12 inhibits miR-16 expression in
CRC cells
The effects of SNHG12 on miR-16 expression in CRC
cells were then further studied. As presented in Fig. 4A, silencing of SNHG12 increased the
expression of miR-16 in SW620 and HT-29 cells. Furthermore,
overexpression of SNHG12 markedly reduced the expression of miR-16
in CRC cells (Fig. 4B). It was
therefore indicated that SNHG12 negatively affects the miR-16
levels in CRC cells.
miR-16 is involved in SNHG12-induced
CRC cell proliferation and invasion
Based on the above results, it was speculated that
SNHG12 promotes CRC cell proliferation and invasion by acting as a
sponge of miR-16. To validate this speculation, SW620 and HT-29
cells were co-transfected with NC siRNA and NC inhibitor (siNC+NC
in group), NC siRNA and miR-16 inhibitor (siNC+miR-16 in group),
SNHG12 siRNA and NC inhibitor (siSNHG12+NC in group), or with
SNHG12 siRNA and miR-16 inhibitor (siSNHG12+miR-16 in group). As
presented in Fig. 5A, the expression
levels of miR-16 were markedly lower in the siNC+miR-16 in group,
while they were higher in the siSNHG12+NC group, when compared with
those in the siNC+NC in group. Furthermore, the expression levels
of miR-16 were lower in the siSNHG12+miR-16 in group when compared
with those in the siSNHG12+NC in group. CKK-8 assay and Transwell
assays further indicated that the proliferation and invasion of CRC
cells were significantly reduced in the siSNHG12+NC in group when
compared with those in the siNC+NC in group, while these effects
were rescued in the siSNHG12+miR-16 in group (Fig. 5B-E). These results suggested that
knockdown of SNHG12 inhibited CRC cell proliferation and invasion
by upregulating miR-16.
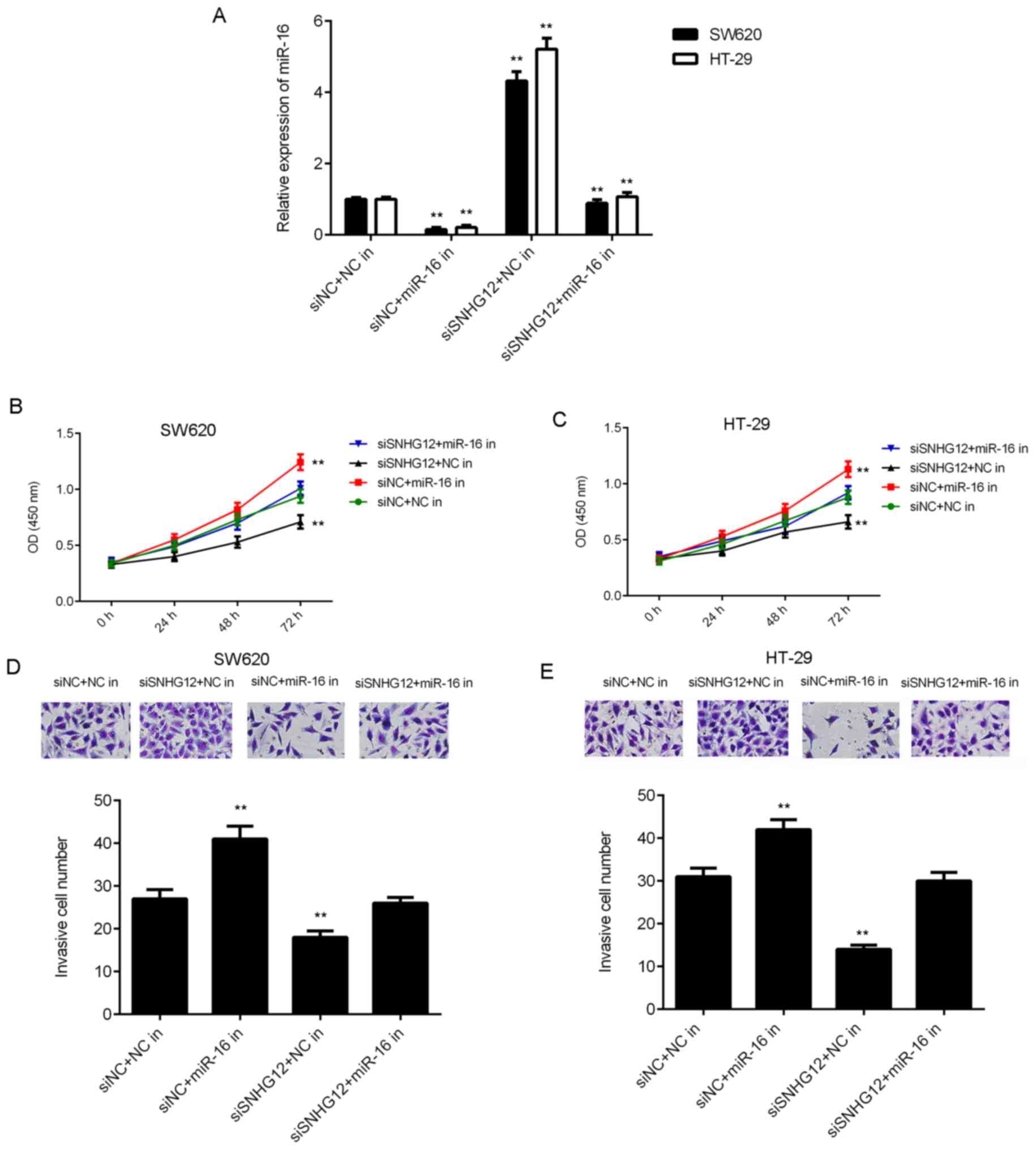 | Figure 5.miR-16 is involved in SNHG12-mediated
colorectal cancer cell proliferation and invasion. SW620 and HT-29
cells were co-transfected with NC siRNA and NC inhibitor (siNC+NC
in), NC siRNA and miR-16 inhibitor (siNC+miR-16 in), SNHG12 siRNA
and NC inhibitor (siSNHG12+NC in), or with SNHG12 siRNA and miR-16
inhibitor (siSNHG12+miR-16 in). After transfection, (A) reverse
transcription-quantitative polymerase chain reaction was used to
examine the levels of miR-16. CCK-8 assay using (B) SW620 and (C)
HT-29 cells, and (D and E) a Transwell assay (magnification, ×200)
were performed to examine cell proliferation and invasion,
respectively. **P<0.01 vs. siSNHG12+NC. siSNHG12, siRNA
targeting SNHG12; in, inhibitor; SNHG12, small nucleolar RNA host
gene 12; siRNA, small interfering RNA; NC, negative control; OD,
optical density; miR, microRNA. |
Discussion
The regulatory mechanism of SNHG12 in CRC cells has
remained largely elusive. In the present study, SNHG12 was
identified to be markedly upregulated in CRC tissue when compared
with adjacent non-tumour tissues, and its upregulation was markedly
associated with CRC progression, as well as poor prognosis of
patients. In addition, SNHG12 expression was higher in CRC cell
lines when compared with that in normal intestinal epithelial
cells. Knockdown of SNHG12 significantly repressed CRC cell
proliferation and invasion, while SNHG12 overexpression
significantly increased the proliferation and invasion of CRC
cells. A Bioinformatics analysis indicated that SNHG12 and miR-16
had complementary binding sites, which was confirmed by a
luciferase reporter gene assay. The expression levels of miR-16
were markedly downregulated in CRC tissues and cell lines, and were
inversely correlated with the expression of SNHG12 in CRC tissues.
In addition, silencing of miR-16 eliminated the suppressive effects
of SNHG12 downregulation on CRC cell proliferation and
invasion.
It has been reported that SNHG12 has a role in
promoting certain common human cancer types. For instance, SNHG12
has been indicated to promote cell proliferation and migration by
increasing the expression of angiomotin in osteosarcoma cells
(19). It was also reported to
promote tumourigenesis and metastasis by targeting miR-199a/b-5p in
hepatocellular carcinoma (18).
Furthermore, SNHG12 contributes to multidrug resistance by
activating the MAPK/Slug pathway by sponging miR-181a in
non-small-cell lung cancer (32). In
addition, C-MYC-induced upregulation of lncRNA SNHG12 promoted cell
proliferation and migration, while inhibiting cell apoptosis in
triple-negative breast cancer (25).
Thus, SNHG12 has been suggested as a promising therapeutic target
for specific cancer types (18,32,33).
Furthermore, knockdown of SNHG12 was demonstrated to inhibit
non-small cell lung cancer cell growth and induce apoptosis by
upregulating miR-138 (33).
Recently, Wang et al (17)
reported that SNHG12 promotes the proliferation and inhibits
apoptosis of CRC cells. However, the underlying mechanisms of the
regulatory function of SNHG12 during CRC growth and metastasis has
remained elusive. The present study confirmed that the expression
of SNHG12 was markedly upregulated in CRC tissues and cell lines
when compared with that in adjacent normal tissues and normal
intestinal epithelial cells, respectively. Consistent with the
present results, Wang et al (17) also observed an increased expression
of SNHG12 in CRC tissues and cell lines. Furthermore, the present
study revealed that high expression of SNHG12 was associated with
lymph node metastasis, advanced clinical stage and poor overall
survival of CRC patients.
Next, the exact roles of SNHG12 to regulate CRC cell
proliferation and invasion were investigated. It was demonstrated
that inhibition of SNHG12 expression led to a significant reduction
in CRC cell proliferation and invasion. Consistent with these
results, Wang et al (17)
also reported that SNHG12 promoted CRC cell proliferation and that
overexpression of SNHG12 enhanced the cell cycle progression of
SW480 CRC cells, while knockdown of SNHG12 repressed the cell cycle
progression of HT29 CRC cells. Furthermore, they demonstrated that
SNHG12 also inhibited CRC cell apoptosis by increasing the
expression of cell cycle-associated proteins and suppressing the
expression of caspase 3 (17). These
previous and the present results indicated that SNHG12 may have a
role in promoting CRC proliferation and metastasis.
Next, the underlying mechanisms of the regulatory
effect of SNHG12 on CRC cell proliferation and invasion were
assessed in vitro. TargetScan software predicted that miR-16
and SNHG12 had complementary binding sites. To confirm this
predicted binding interaction, the expression of miR-16 was
examined in CRC, revealing that miR-16 was markedly downregulated
in CRC tissues and cell lines. Furthermore, an inverse correlation
was observed between miR-16 and SNHG12 in CRC tissues. To further
clarify the association between miR-16 and SNHG12 in CRC, a
luciferase reporter gene assay was performed, and the results
indicated that transfection with miR-16 mimics markedly reduced the
luciferase activity of WT SNHG12 reporter vector but did not affect
the luciferase activity of MT SNHG12 in SW620 and HT-29 cells.
These results suggest that SNHG12 may sponge miR-16 in CRC cells.
As it was further observed that miR-16 expression was negatively
regulated by SNHG12 in CRC cells, it was speculated that miR-16 is
involved in SNHG12-mediated CRC cell proliferation and invasion,
which was further confirmed by the result that inhibition of miR-16
expression abrogated the suppressive effects of SNHG12
downregulation on CRC cell proliferation and invasion.
In summary, the present study demonstrated that
SNHG12 is upregulated in CRC and has a role in promoting CRC cell
proliferation and invasion by acting as a sponge of miR-16. Thus,
SNHG12 may be a promising therapeutic target for CRC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analysed during the present
study are included in this published article.
Authors' contributions
FR designed the study and wrote the manuscript. YL
and J-YZ collected clinical tissues. SW, YS, J-PZ and FR performed
the in-vitro experiments and statistical analysis.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of the Second Xiangya Hospital, Central South
University (Changsha, China) and written informed consent had been
obtained from all subjects.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li JW, Huang CZ, Li JH, Yuan JH, Chen QH,
Zhang WF, Xu ZS, Liu YP, Li Y, Zhan MX and Lu LG: Knockdown of
metadherin inhibits cell proliferation and migration in colorectal
cancer. Oncol Rep. 40:2215–2223. 2018.PubMed/NCBI
|
|
4
|
Cen C, Li J, Liu J, Yang M, Zhang T, Zuo
Y, Lin C and Li X: Long noncoding RNA LINC01510 promotes the growth
of colorectal cancer cells by modulating MET expression. Cancer
Cell Int. 18:452018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Price TJ, Tang M, Gibbs P, Haller DG,
Peeters M, Arnold D, Segelov E, Roy A, Tebbutt N, Pavlakis N, et
al: Targeted therapy for metastatic colorectal cancer. Expert Rev
Anticancer Ther. 18:991–1006. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Islam F, Gopalan V, Vider J, Wahab R,
Ebrahimi F, Lu CT, Kasem K and Lam AKY: MicroRNA-186-5p
overexpression modulates colon cancer growth by repressing the
expression of the FAM134B tumour inhibitor. Exp Cell Res.
357:260–270. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xiao Z, Qu Z, Chen Z, Fang Z, Zhou K,
Huang Z, Guo X and Zhang Y: LncRNA HOTAIR is a prognostic biomarker
for the proliferation and chemoresistance of colorectal cancer via
MiR-203a-3p-mediated Wnt/β-catenin signaling pathway. Cell Physiol
Biochem. 46:1275–1285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xie B, Deng Z, Pan Y, Fu C, Fan S, Tao Y,
Zhou J and Xiao D: Post-transcriptional regulation DPC4 gene by
miR-190 in colorectal cancer cells. J Cancer Res Ther. 14:838–843.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu K, Yao H, Wen Y, Zhao H, Zhou N, Lei S
and Xiong L: Functional role of a long non-coding RNA
LIFR-AS1/miR-29a/TNFAIP3 axis in colorectal cancer resistance to
pohotodynamic therapy. Biochim Biophys Acta. 1864:2871–2880. 2018.
View Article : Google Scholar
|
|
10
|
Liao D, Li T, Ye C, Zeng L, Li H, Pu X,
Ding C, He Z and Huang GL: miR-221 inhibits autophagy and targets
TP53INP1 in colorectal cancer cells. Exp Ther Med. 15:1712–1717.
2018.PubMed/NCBI
|
|
11
|
Yunusov D, Anderson L, DaSilva LF, Wysocka
J, Ezashi T, Roberts RM and Verjovski-Almeida S: HIPSTR and
thousands of lncRNAs are heterogeneously expressed in human
embryos, primordial germ cells and stable cell lines. Sci Rep.
6:327532016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou Y, Chen Y, Ding W, Hua Z, Wang L, Zhu
Y, Qian H and Dai T: LncRNA UCA1 impacts cell proliferation,
invasion, and migration of pancreatic cancer through regulating
miR-96/FOXO3. IUBMB Life. 70:276–290. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peng Z, Liu C and Wu M: New insights into
long noncoding RNAs and their roles in glioma. Mol Cancer.
17:612018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smolle MA and Pichler M: The role of long
non-coding RNAs in osteosarcoma. Noncoding RNA. 4:E72018.PubMed/NCBI
|
|
16
|
Xu S, Kong D, Chen Q, Ping Y and Pang D:
Oncogenic long noncoding RNA landscape in breast cancer. Mol
Cancer. 16:1292017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang JZ, Xu CL, Wu H and Shen SJ: LncRNA
SNHG12 promotes cell growth and inhibits cell apoptosis in
colorectal cancer cells. Braz J Med Biol Res. 50:e60792017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lan T, Ma W, Hong Z, Wu L, Chen X and Yuan
Y: Long non-coding RNA small nucleolar RNA host gene 12 (SNHG12)
promotes tumorigenesis and metastasis by targeting miR-199a/b-5p in
hepatocellular carcinoma. J Exp Clin Cancer Res. 36:112017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ruan W, Wang P, Feng S, Xue Y and Li Y:
Long non-coding RNA small nucleolar RNA host gene 12 (SNHG12)
promotes cell proliferation and migration by upregulating
angiomotin gene expression in human osteosarcoma cells. Tumour
Biol. 37:4065–4073. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lei W, Wang ZL, Feng HJ, Lin XD, Li CZ and
Fan D: Long non-coding RNA SNHG12promotes the proliferation and
migration of glioma cells by binding to HuR. Int J Oncol.
53:1374–1384. 2018.PubMed/NCBI
|
|
21
|
Ding S, Qu W, Jiao Y, Zhang J, Zhang C and
Dang S: LncRNA SNHG12 promotes the proliferation and metastasis of
papillary thyroid carcinoma cells through regulating
wnt/beta-catenin signaling pathway. Cancer Biomark. 22:217–226.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang BF, Cai W and Chen B: LncRNA SNHG12
regulated the proliferation of gastric carcinoma cell BGC-823 by
targeting microRNA-199a/b-5p. Eur Rev Med Pharmacol Sci.
22:1297–1306. 2018.PubMed/NCBI
|
|
23
|
Dong J, Wang Q, Li L and Xiao-Jin Z:
Upregulation of long non-coding RNA small nucleolar RNA host gene
12 contributes to cell growth and invasion in cervical cancer by
acting as a sponge for MiR-424-5p. Cell Physiol Biochem.
45:2086–2094. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang H and Lu W: LncRNA SNHG12 regulates
gastric cancer progression by acting as a molecular sponge of
miR320. Mol Med Rep. 17:2743–2749. 2018.PubMed/NCBI
|
|
25
|
Wang O, Yang F, Liu Y, Lv L, Ma R, Chen C,
Wang J, Tan Q, Cheng Y, Xia E, et al: C-MYC-induced upregulation of
lncRNA SNHG12 regulates cell proliferation, apoptosis and migration
in triple-negative breast cancer. Am J Transl Res. 9:533–545.
2017.PubMed/NCBI
|
|
26
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li Y, Sun Z, Liu B, Shan Y, Zhao L and Jia
L: Tumor-suppressive miR-26a and miR-26b inhibit cell
aggressiveness by regulating FUT4 in colorectal cancer. Cell Death
Dis. 8:e28922017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lv H, Zhang Z, Wang Y, Li C, Gong W and
Wang X: MicroRNA-92a promotes colorectal cancer cell growth and
migration by inhibiting KLF4. Oncol Res. 23:283–290. 2016.
View Article : Google Scholar
|
|
29
|
Qian J, Jiang B, Li M, Chen J and Fang M:
Prognostic significance of microRNA-16 expression in human
colorectal cancer. World J Surg. 37:2944–2949. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma Q, Wang X, Li Z, Li B, Ma F, Peng L,
Zhang Y, Xu A and Jiang B: microRNA-16 represses colorectal cancer
cell growth in vitro by regulating the p53/survivin
signaling pathway. Oncol Rep. 29:1652–1658. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang P, Chen D, Ma H and Li Y: LncRNA
SNHG12 contributes to multidrug resistance through activating the
MAPK/Slug pathway by sponging miR-181a in non-small cell lung
cancer. Oncotarget. 8:84086–84101. 2017.PubMed/NCBI
|
|
33
|
Wang X, Qi G, Zhang J, Wu J, Zhou N, Li L
and Ma J: Knockdown of long noncoding RNA small nucleolar RNA host
gene 12 inhibits cell growth and induces apoptosis by upregulating
miR-138 in nonsmall cell lung cancer. DNA Cell Biol. 36:892–900.
2017. View Article : Google Scholar : PubMed/NCBI
|