Introduction
Cervical cancer is one of the most common
gynecological malignancies and is currently the fourth most common
cause of cancer-associated mortality worldwide (1). According to data obtained from Global
cancer statistics (2), it is
estimated that there were 569,847 new cases and 311,365 mortalities
in 2018 as a result of cervical cancer.
Although human papillomavirus (HPV) infection has
been demonstrated to be closely associated with the pathogenesis of
cervical cancer, HPV alone cannot be accounted for as the sole
cause of this disease (3,4). Clinical outcomes for cervical cancer
remain unsatisfactory despite improvements in diagnostic techniques
and therapeutic approaches, including surgery, radiotherapy and
chemotherapy (5). The primary
reasons for poor prognosis in patients with cervical cancer include
recurrence, invasion and metastasis (6). Therefore, it is of urgent concern to
fully elucidate the mechanism underlying the aggressive
pathophysiology of this disease, to identify effective therapeutic
interventions and to improve patient prognosis.
In the past few decades, microRNAs (miRNAs) have
been implicated in tumorigenesis and tumor development (7–9). miRNAs
are a class of non-coding short RNA molecules, 18–24 nucleotides in
length (10) that regulate gene
expression by binding to target sites in the 3′-untranslated
regions (3′-UTRs) of target mRNAs, resulting in translation
suppression and/or mRNA degradation (11). To date, >2,500 miRNAs have been
identified in the human genome (12), with ~60% of protein-coding genes
reported to be targeted by miRNAs (13). Dysregulation of miRNA function has
been documented in nearly all types of human malignant tumors such
as cervical cancer (14–16). miRNAs may be involved in the
modulation of cervical carcinogenesis and progression by acting as
either oncogenes or tumor suppressors (17–19).
Therefore, unravelling the downstream consequences of miRNA
dysregulation in cervical cancer is of clinical importance for the
identification of effective biomarkers for the diagnosis and
prognosis of cervical cancer in addition to the development of
novel therapeutic approaches for patients with this malignancy.
microRNA-889-3p (miR-889) has been previously
demonstrated to serve crucial roles in esophageal squamous cell
carcinoma (20) and hepatocellular
carcinoma (21). However, to the
best of our knowledge, no studies concerning the relationship
between miR-889 and cervical cancer have been performed. Therefore,
the aim of this study was to investigate miR-889 expression in
cervical cancer and its clinical significance, and to examine the
potential effects of miR-889 in cervical cancer development on a
molecular level. Expression and functions of fibroblast growth
factor receptor 2 (FGFR2) have been extensively studied in multiple
human cancer types (22–25). FGFR2 is highly expressed in cervical
cancer tissues (26). In the present
study, FGFR2 was predicted as a potential target of miR-889. A
series of experiments were performed to validate whether FGFR2 was
a direct target gene of miR-889 in cervical cancer, and whether a
decrease of FGFR2 was responsible for the activity of miR-889
overexpression in cervical cancer cells.
Materials and methods
Ethical approval and collection
clinical tissues
This study was approved by the Ethics Committee of
Weifang People's Hospital (Weifang, China), and written informed
consent was provided by all individuals who participated in the
present study. In total, 49 pairs of cervical cancer tissues and
matched non-cancerous tissues were obtained from patients with
cervical cancer (age range, 46–71 years) following surgical
resection at Weifang People's Hospital between June 2015 and August
2017. The inclusion criteria were patients diagnosed with cervical
cancer and treated with surgical resection. In addition, none of
the patients received radiotherapy or chemotherapy prior to
surgery. Patients treated with preoperative radiotherapy or
chemotherapy were excluded from this study. All tissues were
promptly snap-frozen in liquid nitrogen following surgical
resection and stored at −80°C until further use.
Cell lines and culture conditions
A total of four human cervical cancer cell lines
(HeLa, C-33A, CaSki and SiHa) and a normal human cervical
epithelial cell line (Ect1/E6E7) were purchased from American Type
Culture Collection. All cell lines were maintained in Dulbecco's
modified Eagle's medium (DMEM) supplemented with 10% FBS, 100 U/ml
penicillin and 100 µg/ml streptomycin (all from Invitrogen; Thermo
Fisher Scientific, Inc.) in a humidified atmosphere at 37°C under
5% CO2.
Cell transfection
The miR-889 mimic and negative control miRNA mimic
(miR-NC) were chemically constructed by Guangzhou RiboBio Co., Ltd.
The miR-889 mimics sequence was 5′-UUAAUAUCGGACAACCAUUGU-3′ and the
miR-NC sequence was 5′-UUCUCCGAACGUGUCACGUTT-3′. Small interfering
RNA (siRNA) targeting FGFR2 and negative control siRNA (NC siRNA)
were purchased from Shanghai GenePharma Co., Ltd. The FGFR2 siRNA
sequence was 5′-GGAGGUGCUUCACUUAAGATT-3′ and the NC siRNA sequence
was 5′-UUCUCCGAACGUGUCACGUTT-3′. The pcDNA3.1-FGFR2 overexpression
vector (pc-FGFR2) lacking its 3′-UTR, and the empty pcDNA3.1 vector
were purchased from the Chinese Academy of Sciences. Cells were
seeded (6×105 cells/well) into 6-well plates 24 h prior
to transfection and subsequently transfected with miR-889 mimic
(100 pmol), miR-NC (100 pmol), FGFR2 siRNA (100 pmol), NC siRNA
(100 pmol), pc-FGFR2 (4 µg) or empty pcDNA3.1 vector (4 µg) using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Transfection was performed at room temperature
according to manufacturer's protocols. Reverse
transcription-quantitative PCR (RT-qPCR) and western blot analysis
were conducted 48 and 72 h following transfection, respectively.
Cell Counting Kit-8 (CCK-8) assay was performed on transfected
cells 24 h after transfection, whereas invasion assay was conducted
48 h following transfection.
RT-qPCR
For the measurement of miR-889 and FGFR2 mRNA
expression, total RNA was isolated from tissues (50 mg) or cells
(1×106) using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to manufacturer's
protocol. A Nanodrop 2000 spectrophotometer (Thermo Fisher
Scientific, Inc.) was utilized to assess the concentration and
quality of total RNA. For the detection of miR-889 expression,
total RNA was reverse-transcribed into cDNA using miScriptII
Reverse Transcription Kit (Qiagen China Co., Ltd.) according to the
manufacturer's protocol. The cDNA was subsequently subjected to
qPCR using miScript SYBR® Green PCR Kit (Qiagen China
Co., Ltd.) according to manufacturer's protocol. The thermocycling
conditions for qPCR were as follows: 95°C for 2 min, 95°C for 10
sec, 55°C for 30 sec and 72°C for 30 sec, for 40 cycles.
For the measurement of FGFR2 mRNA expression, cDNA
was generated from RNA using Prime-Script Reverse Transcription
Reagent Kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocol. qPCR was performed using SYBR Premix Ex
Taq™ Kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocol. The thermocycling conditions for qPCR were
as follows: 5 min at 95°C, followed by 40 cycles of 95°C for 30 sec
and 65°C for 45 sec. U6 small nuclear RNA and GAPDH served as
loading controls and for normalization of miR-889 and FGFR2 mRNA
expression, respectively. The 2−ΔΔCq method was applied
to quantify relative gene expression (27). The following primer pairs were used
for the qPCR: miR-889, forward 5′-ACACTCCAGCTGGGTTAATATCGGACAAC-3′
and reverse 5′-TGGTGTCGTGGAGTCG-3′; U6, forward
5′-CTCGCTTCGGCAGCACA-3′ and reverse 5′-AACGCTTCACGAATTTGCGT-3′;
FGFR2, forward 5′-CGCTGGTGAGGATAACAACACG-3′ and reverse
5′-TGGAAGTTCATACTCGGAGACCC-3′; and GAPDH, forward
5′-AACGGATTTGGTCG-TATTG-3′ and reverse
5′-GGAAGATGGTGATGGGATT-3′.
CCK-8 assay
A total of 100 µl culture medium containing ~3,000
cells were seeded into each well of 96-well plates 24 h after
transfection. Five replicates were set for every group. The culture
plates were incubated at 37°C under 5% CO2 for 0, 24, 48
and 72 h before 10 µl of CCK-8 solution (Dojindo Molecular
Technologies, Inc.) was added into each well. Following a further 2
h of incubation, absorbance was measured at 450 nm using a
microplate reader (Bio-Rad Laboratories, Inc.) to determine cell
viability.
Invasion assay
The invasive ability of the cells was examined using
24-well Transwell® chambers (pore size, 8 µm; Corning
Inc.) pre-coated with Matrigel (BD Biosciences). Cells were
serum-starved for 12 h then harvested and resuspended in FBS-free
DMEM prior to being seeded into the upper chambers at a density of
5×104 cells/well 48 h following transfection. A total of
500 µl DMEM supplemented with 10% FBS was added into the lower
chambers, which served as the chemoattractant. Following 24 h of
incubation, cells remaining on the upper side of the chambers were
mechanically removed using a cotton swab. Cells that successfully
invaded were fixed with 4% paraformaldehyde at room temperature for
30 min, stained with 0.1% crystal violet at room temperature for 30
min and washed with PBS. The average number of invaded cells was
counted using a light microscope (magnification, ×200; Olympus
Corporation) from five randomly chosen microscopic fields of view
for each membrane.
Bioinformatics prediction
TargetScan (Release 7.2; http://www.targetscan.org/vert_72/) and miRDB
(www.mirdb.org) were used to search for potential
miR-889 targets. Hsa-miR-889-3p was entered into the searching bar
and a list of potential target genes were obtained.
Dual-luciferase reporter assay
The wild-type (WT) miR-889 target site in the 3′-UTR
of FGFR2 or a mutant (MUT) 3′-UTR were amplified by Shanghai
GenePharma Co., Ltd. and inserted into the pMIR-GLO™
Luciferase vector (Promega Corporation) downstream of the firefly
luciferase coding region. The generated luciferase reporter
plasmids were defined as pMIR-FGFR2-3′-UTR WT and pMIR-FGFR2-3′-UTR
MUT thereafter. Cells were seeded into 24-well plates at a density
of 1×105 cells/well one day prior to transfection.
miR-889 mimic (20 pmol) or miR-NC (20 pmol) were co-transfected
with pMIR-FGFR2-3′-UTR WT (0.2 µg) or pMIR-FGFR2-3′-UTR MUT (0.2
µg) using Lipofectamine 2000, in accordance with the manufacturer's
protocol. Co-transfection was performed at room temperature then
co-transfected cells were incubated at 37°C under 5% CO2
for 48 h. Using a Dual-Luciferase Reporter Assay System (Promega
Corporation), luciferase activity was assessed 48 h after
incubation according to the manufacturer's protocol. Firefly
luciferase activity was normalized to Renilla luciferase
activity.
Western blot analysis
Cells (1×106) and tissues (100 mg) were
washed with PBS prior to protein extraction using
radioimmunoprecipitation assay buffer (Sigma-Aldrich; Merck KGaA).
Protein concentration was measured using Bicinchoninic Acid protein
assay kit (Sigma-Aldrich; Merck KGaA). Equal amounts of protein (30
µg) were separated by 10% SDS-PAGE, transferred to polyvinylidene
difluoride membranes and blocked at room temperature for 2 h with
5% non-fat milk powder diluted in Tris-buffered saline containing
0.1% Tween-20 (TBST). Following incubation overnight at 4°C with
primary antibodies, the membranes were washed with TBST and
incubated at room temperature for 1 h with horseradish
peroxidase-conjugated goat anti-mouse secondary antibody (1:5,000
dilution; cat. no. ab205719; Abcam). To visualize the protein bands
ECL™ Western Blotting Detection Reagents (GE Healthcare)
was used. The primary antibodies used in this study were as
follows: FGFR2 (1:1,000 dilution; cat. no. ab58201; Abcam) and
GAPDH (1:1,000 dilution; cat. no. ab9482; Abcam). Quantity One
software version 4.62 (Bio-Rad Laboratories, Inc.) was employed for
performing densitometric analysis to quantify protein
expression.
Statistical analysis
All data were presented as the mean ± standard
deviation from at least three separate experiments. SPSS software
version 16.0 (SPSS, Inc.) was used to perform all statistical
analyses. χ2 test was applied to determine the
association between miR-889 expression and clinicopathological
factors in patients with cervical cancer. Student's t-test was used
to analyze differences between two groups. Differences between
multiple groups were compared using one-way analysis of variance
followed by Tukey's post-hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-889 expression is reduced in
cervical cancer tissues and cell lines
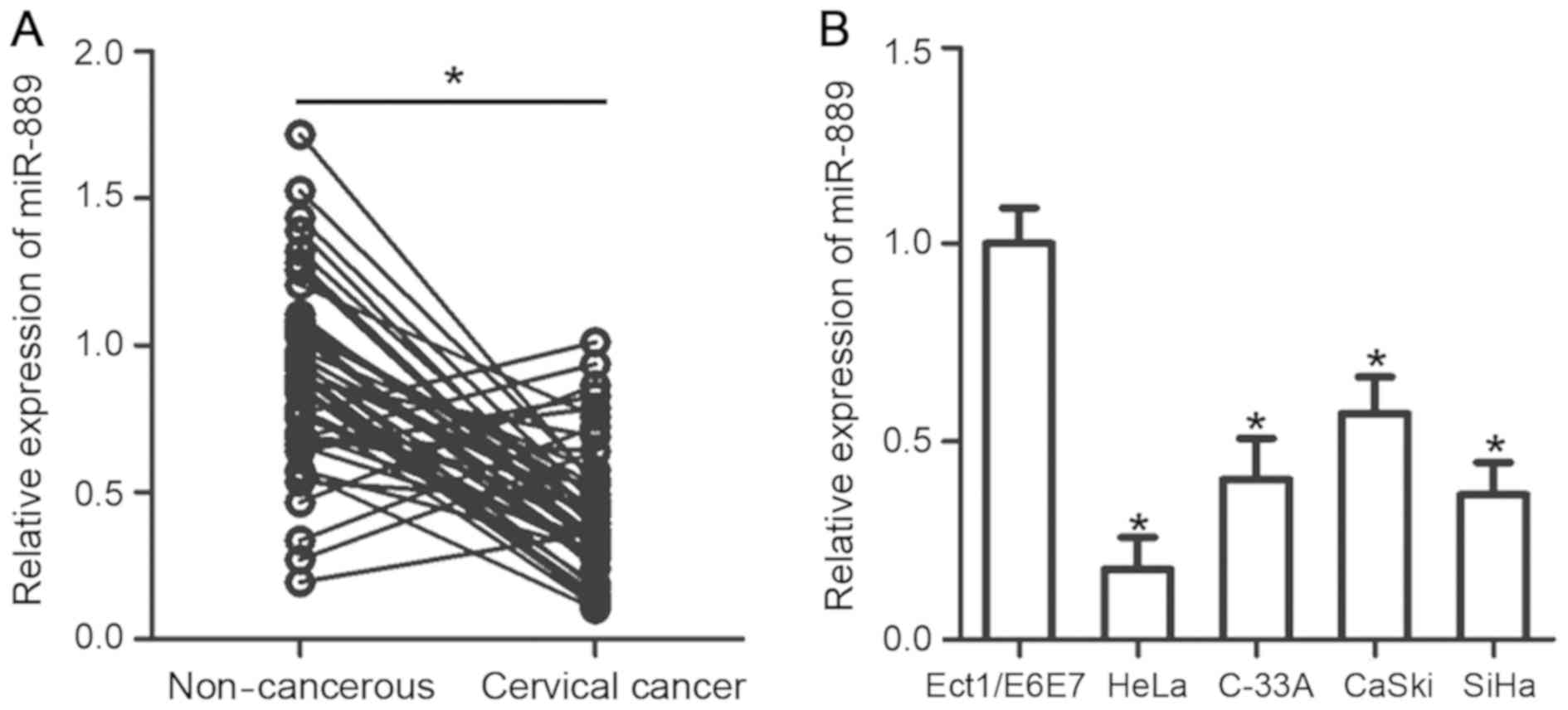
RT-qPCR analysis was performed to measure miR-889
expression levels in 49 pairs of cervical cancer tissues and
matched non-cancerous tissues. A significantly lower level of
miR-889 expression was observed in cervical cancer tissues compared
with their corresponding matched noncancerous tissues (P<0.05;
Fig. 1A). Subsequently, the
association between miR-889 and a number of clinicopathologic
factors were investigated in patients with cervical cancer. All
patients were divided into either low or high miR-889 expression
groups using the median value of miR-889 in cervical cancer tissues
as the cutoff value. Low miR-889 expression was demonstratively
associated with International Federation of Gynecology and
Obstetrics (FIGO) stages (P=0.030) and increased lymph node
metastasis (P=0.012; Table I).
 | Table I.Association between miR-889 expression
and clinicopathological features of patients with cervical
cancer. |
Table I.
Association between miR-889 expression
and clinicopathological features of patients with cervical
cancer.
|
| miR-889
expression |
|
|---|
|
|
|
|
|---|
| Clinicopathological
feature | Low | High | P-value |
|---|
| Age (years) |
|
| 0.444 |
|
<55 | 11 | 8 |
|
|
≥55 | 14 | 16 |
|
| Tumor size
(cm) |
|
| 0.588 |
|
<4 | 8 | 6 |
|
| ≥4 | 17 | 18 |
|
| Family history of
cancer |
|
| 0.413 |
|
Yes | 4 | 2 |
|
| No | 21 | 22 |
|
| FIGO stage |
|
| 0.030 |
|
I–II | 10 | 17 |
|
|
III–IV | 15 | 7 |
|
| Lymph node
metastasis |
|
| 0.012 |
| No | 11 | 19 |
|
|
Yes | 14 | 5 |
|
To support this observation, the level of miR-889
mRNA expression was measured and assessed in four human cervical
cancer cell lines (HeLa, C-33A, CaSki and SiHa) and a normal human
cervical epithelial cell line (Ect1/E6E7) using RT-qPCR. miR-889
was demonstrated to be significantly lower in all four of the
cervical cancer cell lines compared with expression in Ect1/E6E7
cells (P<0.05; Fig. 1B). These
results suggested that miR-889 expression is downregulated in
cervical cancer, and may be linked with the development of this
malignancy.
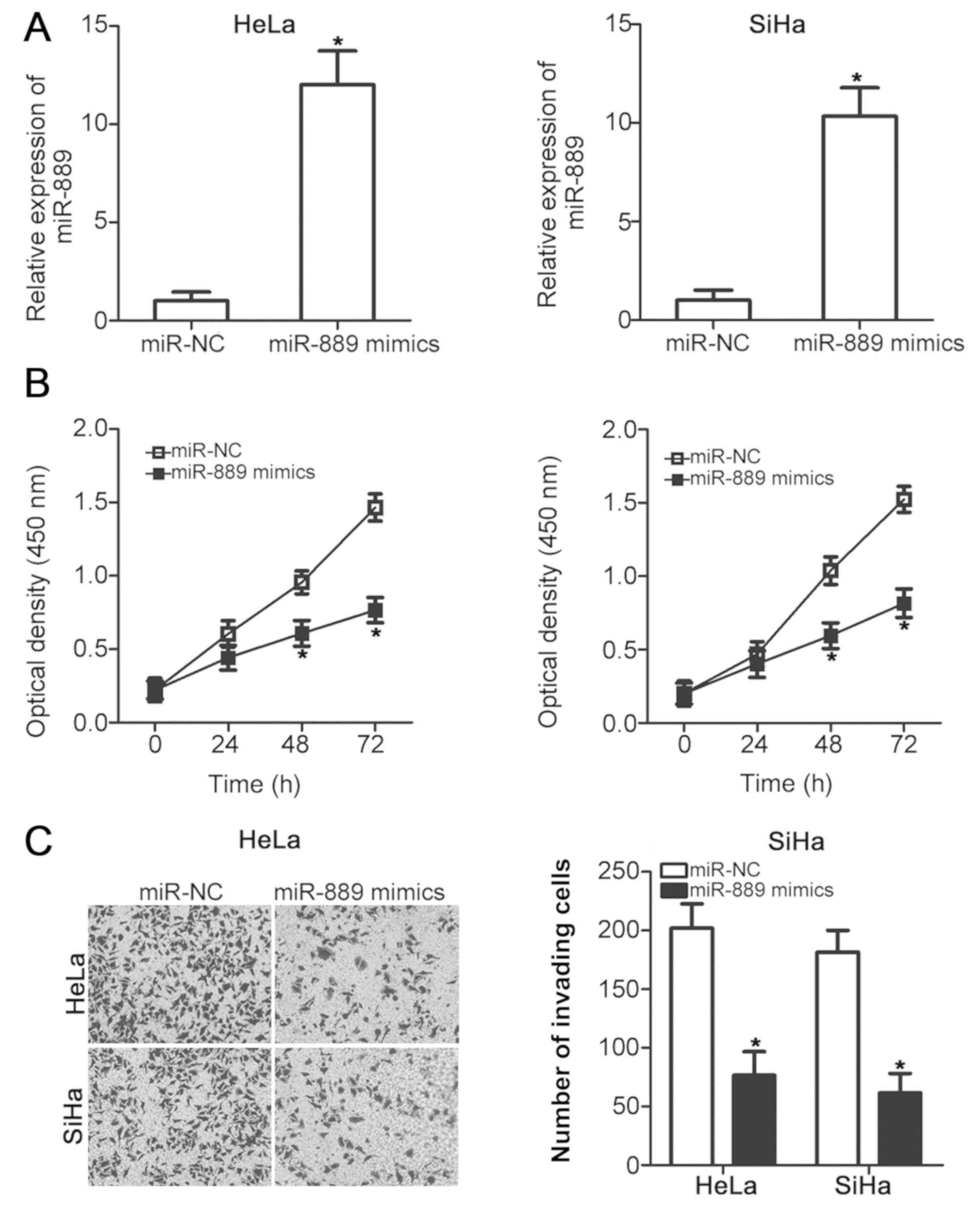
miR-889 inhibits cervical cancer cell
viability and invasion
HeLa and SiHa cells exhibited the lowest expression
levels of miR-889 among the four cervical cancer cell lines
(Fig. 1B); therefore, these two
cervical cancer cell lines were chosen for subsequent experiments.
To assess the role of miR-889 in cervical cancer malignant
phenotype, HeLa and SiHa cells were transfected with either the
miR-889 mimic or miR-NC. Successful upregulation of miR-889
expression after miR-889 mimic transfection was verified in the two
cell lines using RT-qPCR (P<0.05; Fig. 2A). The effect of miR-889 upregulation
on HeLa and SiHa cell viability was investigated using CCK-8 assay;
miR-889 mimic transfection significantly inhibited HeLa and SiHa
cell viability compared with cells transfected with miR-NC at 48
and 72 h post-transfection (P<0.05; Fig. 2B). The role of miR-889 transfection
on cervical cancer cell invasion was also examined. HeLa and SiHa
cells transfected with the miR-889 mimic exhibited a significantly
reduced number of invading cells compared with miR-NC-transfected
cells (P<0.05; Fig. 2C). These
results suggested that miR-889 may serve a tumor suppressive role
in cervical cancer.
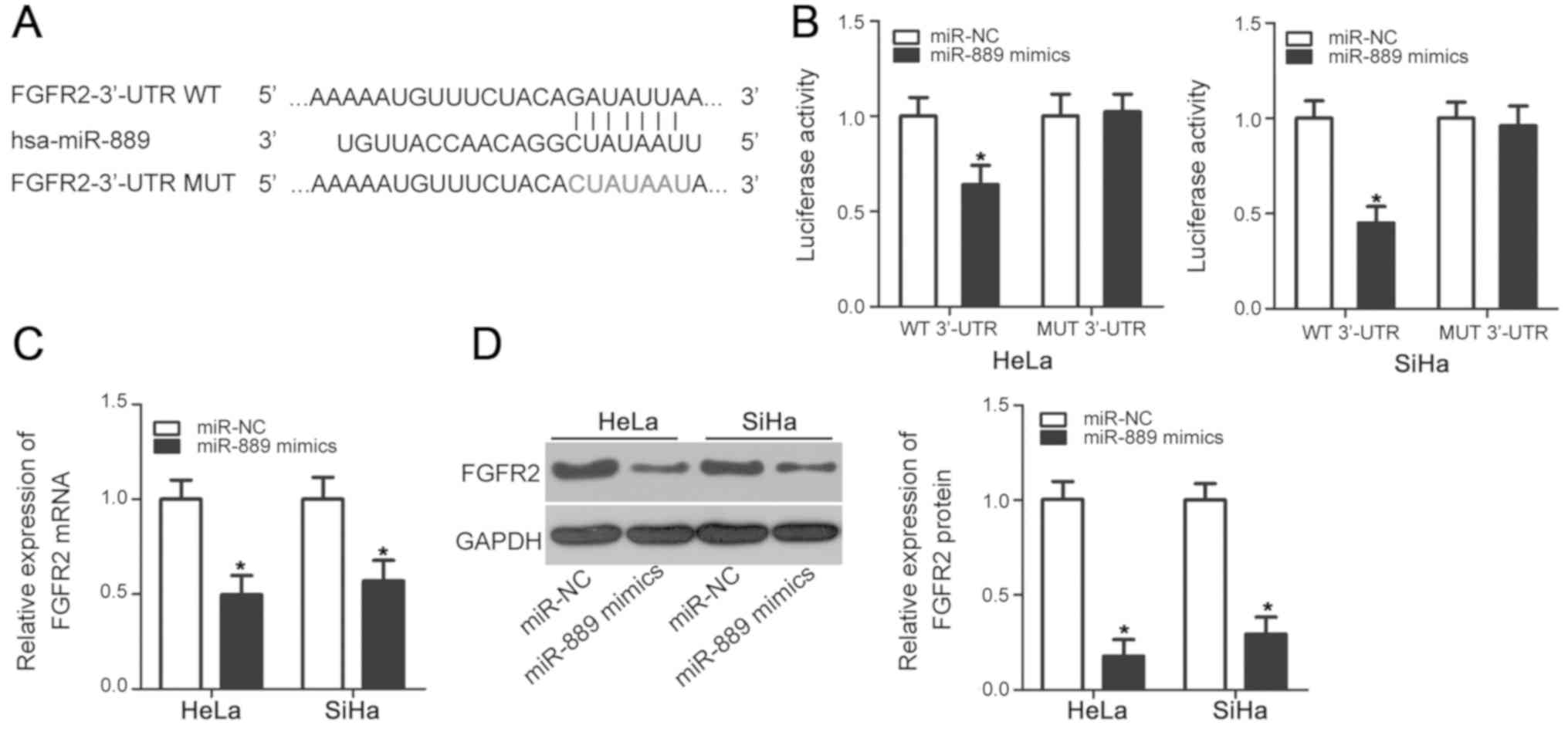
FGFR2 is a direct target of miR-889 in
cervical cancer cells
To decipher the mechanism of involvement in cervical
cancer tumorigenesis, bioinformatic screens were used to predict
the putative targets of miR-889. In particular, FGFR2 was predicted
to be a potential target of miR-889 according to TargetScan and
miRDB (Fig. 3A). Since FGFR2 has
been previously reported to be closely associated with cervical
cancer tumorigenesis in addition to being directly targeted by a
number of miRNAs in other malignancies (23,26,28–30), it
was chosen for further study. Luciferase reporter assay was used to
determine if miR-889 can directly bind to the 3′-UTR of FGFR2 in
cervical cancer cell lines. Compared with cells transfected with
the miR-NC, miR-889 mimic overexpression significantly reduced
luciferase activity in HeLa and SiHa cells transfected with the
pMIR-FGFR2-3′-UTR WT plasmid (P<0.05), but exerted no
significant effects on luciferase activity in cells transfected
with the p-MIR-FGFR2-3′-UTR MUT plasmid (Fig. 3B). RT-qPCR and western blot analysis
were subsequently performed to evaluate the effects of miR-889
mimic transfection on endogenous FGFR2 expression in cervical
cancer cells. The levels of FGFR2 mRNA (P<0.05; Fig. 3C) and protein (P<0.05; Fig. 3D) expression were significantly
reduced in HeLa and SiHa cells following miR-889 mimic transfection
compared with cells transfected with miR-NC. These results
suggested that FGFR2 is a direct target of miR-889 in cervical
cancer cells.
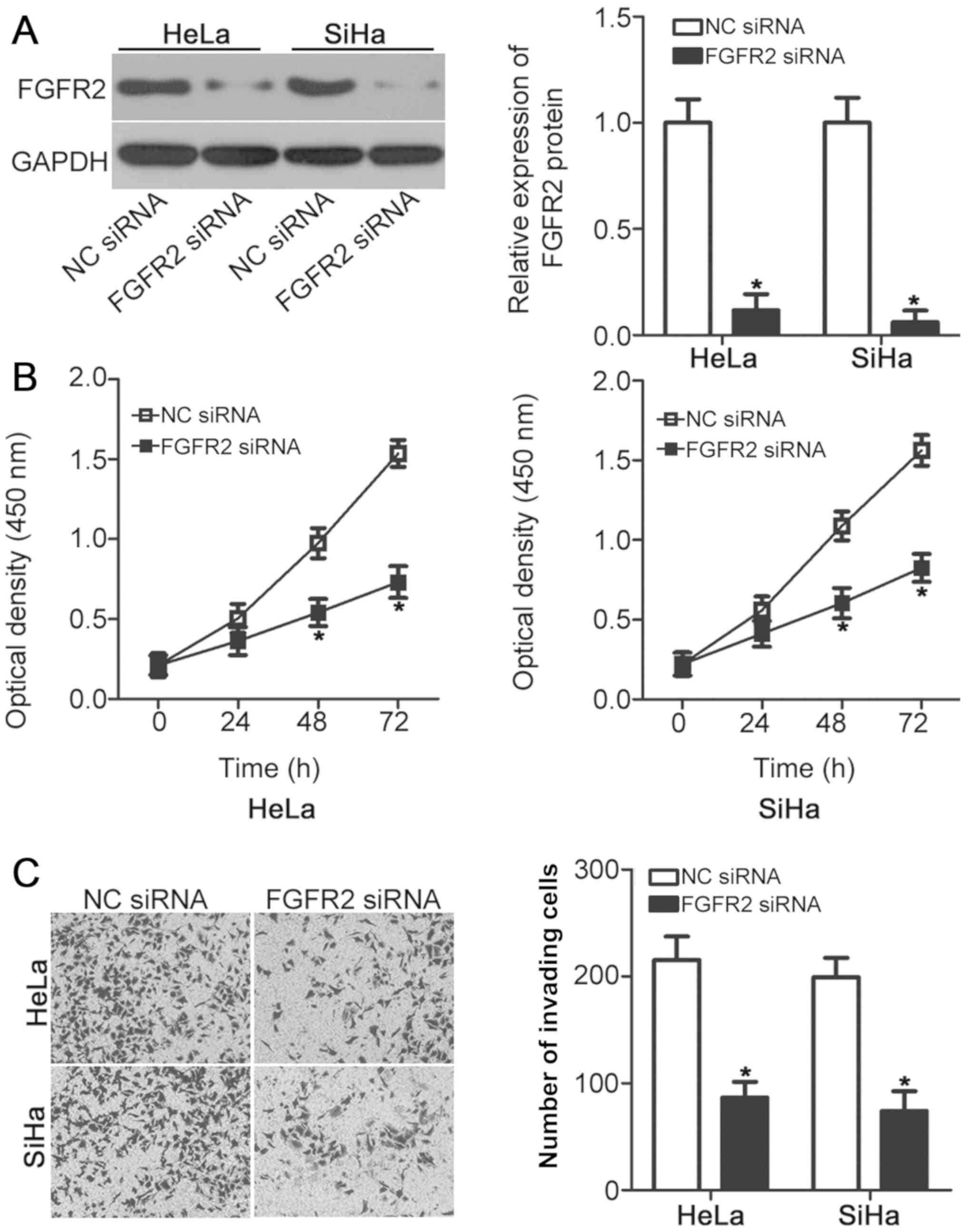
FGFR2 knockdown mimics the suppressive
effects of miR-889 overexpression on cervical cancer cell
lines
Since FGFR2 was identified as a direct target of
miR-889, the physiological function of FGFR2 was investigated in
cervical cancer cell lines. FGFR2 siRNAs were chemically
synthesized and transfected into HeLa and SiHa cells to suppress
endogenous FGFR2 expression. Western blot analysis verified the
efficiency of FGFR2 knockdown in HeLa and SiHa cells following
FGFR2 siRNA transfection (P<0.05; Fig. 4A). FGFR2 knockdown resulted in a
significant reduction in HeLa and SiHa cell viability (P<0.05;
Fig. 4B) and invasive ability
(P<0.05; Fig. 4C), an observation
that was similar to that caused by miR-889 overexpression (Fig. 2). These results further suggested
that FGFR2 may be a functional target of miR-889 in cervical cancer
cells.
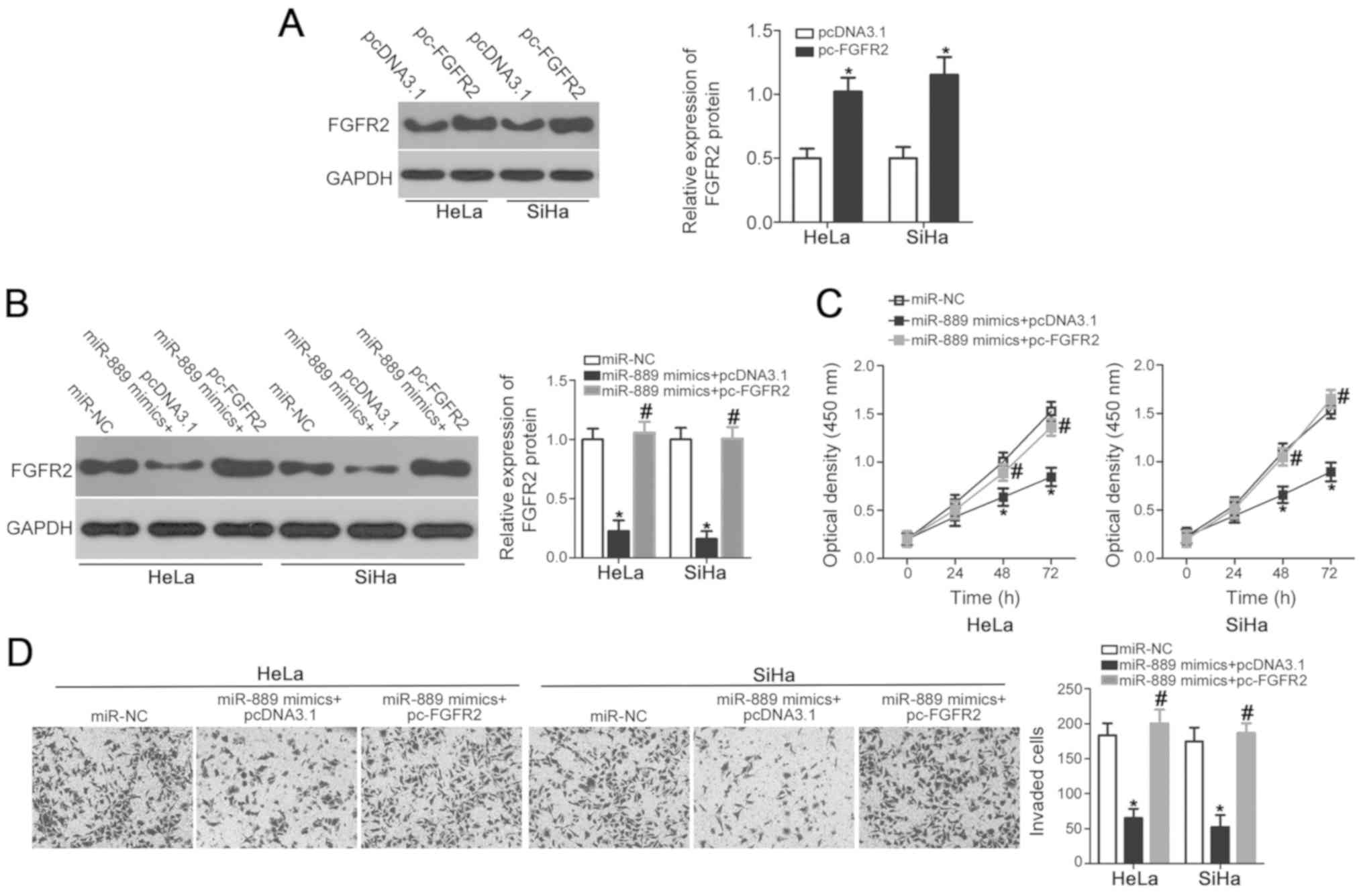
FGFR2 overexpression rescues
miR-889-mediated suppression of cervical cancer cell viability and
invasion
To explore whether FGFR2 is a direct downstream
mediator of miR-889, rescue experiments were performed in HeLa and
SiHa cells by co-transfecting cells with pc-FGFR2 overexpression
plasmids. First, western blot analysis confirmed that FGFR2 protein
expression was significantly upregulated in HeLa and SiHa cells
transfected with pc-FGFR2 compared with pcDNA3.1 empty vector
(P<0.05; Fig. 5A). Subsequently,
HeLa and SiHa cells were co-transfected with miR-889 mimic and
either pc-FGFR2 or empty pcDNA3.1 vector. Western blot analysis
demonstrated that miR-889 mimic-induced suppression of FGFR2
expression was rescued in HeLa and SiHa cells co-transfected with
pc-FGFR2, but not in those transfected with pcDNA3.1 (P<0.05;
Fig. 5B). Similarly, FGFR2
overexpression reversed the suppressive effects of miR-889 mimic on
HeLa and SiHa cell viability (P<0.05; Fig. 5C) and invasive ability (P<0.05;
Fig. 5D). Taken together, these
results further suggested that FGFR2 may be at least partially
involved in the miR-889-mediated suppression of cervical cancer
cell viability and invasion.
Discussion
miRNAs have regularly been found to be dysregulated
in cervical cancer (31). Indeed,
the dysregulation of miRNAs has been reported to contribute to a
number of processes associated with cancer pathophysiology,
including cell proliferation, cell cycle, apoptosis,
epithelial-mesenchymal transition, metastasis, angiogenesis and
resistance to chemo- and radiotherapy (32–34).
Therefore, a thorough understanding of the mechanism underlying
cervical cancer carcinogenesis and progression is crucial for early
diagnosis and improving patient outcome (14). In particular, further exploration on
the specific roles of dysregulated miRNAs in cervical cancer and
their associated mechanism may provide an avenue for the
development of effective therapeutic approaches.
miR-889 is upregulated in esophageal squamous cell
carcinoma tissues and cell lines (20). Upregulation of miR-889 expression
prohibits esophageal squamous cell carcinoma cell growth in
vitro and in vivo by directly targeting disabled
homolog-2 interactive protein (20).
miR-889 is also downregulated by histone deacetylase inhibitors in
hepatocellular carcinoma cells (21). miR-889 overexpression attenuates
hepatocellular carcinoma cell susceptibility to natural killer
cell-mediated lysis by downregulating major histocompatibility
complex class I chain-related gene B (21). However, the role of miR-889 in
cervical cancer remains unclear. The present study revealed that
miR-889 expression was lower in cervical cancer tissues and was
found to be negatively associated with FIGO stages and lymph node
metastasis. miR-889 overexpression inhibited viability and invasive
ability of cervical cancer cell lines. These results suggested that
miR-889 may be an attractive therapeutic target for patients with
this malignancy.
miRNAs exert their functions by directly targeting
different genes to regulate cellular processes (11). Specifically, identifying the direct
targets of miR-889 in cervical cancer is important for elucidating
their action on the malignant signatures, which may be useful for
the development of novel therapeutic interventions. Therefore, to
explore the mechanism in which miR-889 attenuated the malignant
phenotype of cervical cancer cells, a bioinformatics screen was
performed to search for genes directly targeted by miR-889. This
screen identified FGFR2 as a potential miR-889 target, which was
subsequently validated using luciferase reporter assay. Expression
analysis of mRNA and protein levels found that miR-889 upregulation
markedly suppressed FGFR2 expression in cervical cancer cell lines.
Supporting this, the effects of miR-889 overexpression on the same
cell lines could be successfully mimicked by FGFR2 knockdown.
Finally, FGFR2 overexpression abrogated the suppressive effects of
miR-889 on cervical cancer cell viability and invasion. These
findings strongly suggested that FGFR2 is a direct and functional
downstream target of miR-889 in cervical cancer cells.
The FGFR2 gene, located on human chromosome 10q26,
is a member of the FGF2/FGFR2 signaling pathway (35). Previous studies have reported that
FGFR2 is highly expressed in a number of human cancers, including
gastric (22), thyroid (23), bladder (24) and breast cancer (25). FGFR2 serves a role in tumorigenesis
by regulating a number of physiological processes, including cell
growth, apoptosis, survival, metastasis, motility and angiogenesis
(36). FGFR2 is expressed strongly
in cervical cancer tissues and cell lines in addition to
associating with lymph node metastasis in previous studies
(26,28). Results from the present study
indicated that miR-889 directly targeted FGFR2 to suppress the
malignant phenotype of cervical cancer. Based on these data, the
miR-889/FGFR2 axis may serve as a potential therapeutic target for
patients with cervical cancer in the future.
In conclusion, to the best of our knowledge, this
was the first study to demonstrate the reduction of miR-889
expression in cervical cancer tissues and cell lines, which was
associated with FIGO stages and lymph node metastasis. miR-889
overexpression suppressed cervical cancer cell viability and
invasion by directly targeting FGFR2. Therefore, miR-889 may serve
important roles in cervical cancer physiology, suggesting this
miRNA to be a potentially novel therapeutic target. However, the
correlation between miR-889 and the prognosis of patients with
cervical cancer was not analyzed in this study; which is a
limitation of this study. In future investigations, prognosis data
of patients with cervical cancer will be collected to explore the
association between miR-889 expression and prognosis.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
KH designed this research and performed reverse
transcription-quantitative PCR and statistical analysis. YS and YC
conducted Cell Counting Kit-8 and invasion assays. Dual-luciferase
reporter assays and western blot analyses were carried out by YZ.
All authors read and approved the final draft of the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Weifang People's Hospital (Weifang, China), and was
performed in accordance with the Declaration of Helsinki and
guidelines of the Ethics Committee of Weifang People's Hospital.
Written informed consent was provided by all individuals that
participated in this study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Castellsague X: Natural history and
epidemiology of HPV infection and cervical cancer. Gynecol Oncol
110 (3 Suppl 2). S4–S7. 2008. View Article : Google Scholar
|
|
4
|
Du CX and Wang Y: Expression of P-Akt,
NFkappaB and their correlation with human papillomavirus infection
in cervical carcinoma. Eur J Gynaecol Oncol. 33:274–277.
2012.PubMed/NCBI
|
|
5
|
Ghebre RG, Grover S, Xu MJ, Chuang LT and
Simonds H: Cervical cancer control in HIV-infected women: Past,
present and future. Gynecol Oncol Rep. 21:101–108. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hildesheim A and Wang SS: Host and viral
genetics and risk of cervical cancer: A review. Virus Res.
89:229–240. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Link A and Kupcinskas J: MicroRNAs as
non-invasive diagnostic biomarkers for gastric cancer: Current
insights and future perspectives. World J Gastroenterol.
24:3313–3329. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
To KK, Tong CW, Wu M and Cho WC: MicroRNAs
in the prognosis and therapy of colorectal cancer: From bench to
bedside. World J Gastroenterol. 24:2949–2973. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang F, Li B and Xie X: The roles and
clinical significance of microRNAs in cervical cancer. Histol
Histopathol. 31:131–139. 2016.PubMed/NCBI
|
|
10
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He L and Hannon GJ: MicroRNAs: small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kozomara A and Griffiths-Jones S: miRBase:
Annotating high confidence microRNAs using deep sequencing data.
Nucleic Acids Res. 42:D68–D73. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Laengsri V, Kerdpin U, Plabplueng C,
Treeratanapiboon L and Nuchnoi P: Cervical Cancer Markers:
Epigenetics and microRNAs. Lab Med. 49:97–111. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu B, Ju S, Chu H, Shen X, Zhang Y, Luo X
and Cong H: The potential function of microRNAs as biomarkers and
therapeutic targets in multiple myeloma. Oncol Lett. 15:6094–6106.
2018.PubMed/NCBI
|
|
16
|
Lu J, Zhan Y, Feng J, Luo J and Fan S:
MicroRNAs associated with therapy of non-small cell lung cancer.
Int J Biol Sci. 14:390–397. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cai N, Hu L, Xie Y, Gao JH, Zhai W, Wang
L, Jin QJ, Qin CY and Qiang R: MiR-17-5p promotes cervical cancer
cell proliferation and metastasis by targeting transforming growth
factor-beta receptor 2. Eur Rev Med Pharmacol Sci. 22:1899–1906.
2018.PubMed/NCBI
|
|
18
|
Li GC, Cao XY, Li YN, Qiu YY, Li YN, Liu
XJ and Sun XX: MicroRNA-374b inhibits cervical cancer cell
proliferation and induces apoptosis through the p38/ERK signaling
pathway by binding to JAM-2. J Cell Physiol. 233:7379–7390. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sanches JGP, Xu Y, Yabasin IB, Li M, Lu Y,
Xiu X, Wang L, Mao L, Shen J, Wang B, et al: miR-501 is upregulated
in cervical cancer and promotes cell proliferation, migration and
invasion by targeting CYLD. Chem Biol Interact. 285:85–95. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu Y, He J, Wang Y, Zhu X, Pan Q, Xie Q
and Sun F: miR-889 promotes proliferation of esophageal squamous
cell carcinomas through DAB2IP. FEBS Lett. 589:1127–1135. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie H, Zhang Q, Zhou H, Zhou J, Zhang J,
Jiang Y, Wang J, Meng X, Zeng L and Jiang X: microRNA-889 is
downregulated by histone deacetylase inhibitors and confers
resistance to natural killer cytotoxicity in hepatocellular
carcinoma cells. Cytotechnology. 70:513–521. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cha Y, Kim HP, Lim Y, Han SW, Song SH and
Kim TY: FGFR2 amplification is predictive of sensitivity to
regorafenib in gastric and colorectal cancers in vitro. Mol
Oncol. 12:993–1003. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu YT, Zheng HB, Zhang DQ, Zhou L and Sun
H: MicroRNA-1266 suppresses papillary thyroid carcinoma cell
metastasis and growth via targeting FGFR2. Eur Rev Med Pharmacol
Sci. 22:3430–3438. 2018.PubMed/NCBI
|
|
24
|
Marzioni D, Lorenzi T, Mazzucchelli R,
Capparuccia L, Morroni M, Fiorini R, Bracalenti C, Catalano A,
David G, Castellucci M, et al: Expression of basic fibroblast
growth factor, its receptors and syndecans in bladder cancer. Int J
Immunopathol Pharmacol. 22:627–638. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lei H and Deng CX: Fibroblast growth
factor receptor 2 signaling in breast cancer. Int J Biol Sci.
13:1163–1171. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi CH, Chung JY, Kim JH, Kim BG and
Hewitt SM: Expression of fibroblast growth factor receptor family
members is associated with prognosis in early stage cervical cancer
patients. J Transl Med. 14:1242016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kawase R, Ishiwata T, Matsuda Y, Onda M,
Kudo M, Takeshita T and Naito Z: Expression of fibroblast growth
factor receptor 2 IIIc in human uterine cervical intraepithelial
neoplasia and cervical cancer. Int J Oncol. 36:331–340.
2010.PubMed/NCBI
|
|
29
|
Li M, Qian Z, Ma X, Lin X, You Y, Li Y,
Chen T and Jiang H: MiR-628-5p decreases the tumorigenicity of
epithelial ovarian cancer cells by targeting at FGFR2. Biochem
Biophys Res Commun. 495:2085–2091. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang X, Ruan H, Hu X, Cao A and Song L:
miR-381-3p suppresses the proliferation of oral squamous cell
carcinoma cells by directly targeting FGFR2. Am J Cancer Res.
7:913–922. 2017.PubMed/NCBI
|
|
31
|
Wang JY and Chen LJ: The role of miRNAs in
the invasion and metastasis of cervical cancer. Biosci Rep.
39:2019.
|
|
32
|
Pedroza-Torres A, Lopez-Urrutia E,
Garcia-Castillo V, Jacobo-Herrera N, Herrera LA, Peralta-Zaragoza
O, López-Camarillo C, De Leon DC, Fernández-Retana J, Cerna-Cortés
JF and Pérez-Plasencia C: MicroRNAs in cervical cancer: Evidences
for a miRNA profile deregulated by HPV and its impact on
radio-resistance. Molecules. 19:6263–6281. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Banno K, Iida M, Yanokura M, Kisu I, Iwata
T, Tominaga E, Tanaka K and Aoki D: MicroRNA in cervical cancer:
OncomiRs and tumor suppressor miRs in diagnosis and treatment.
TheScientificWorldJournal. 2014:1780752014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shishodia G, Verma G, Das BC and Bharti
AC: miRNA as viral transcription tuners in HPV-mediated cervical
carcinogenesis. Front Biosci (Schol Ed). 10:21–47. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Katoh Y and Katoh M: FGFR2-related
pathogenesis and FGFR2-targeted therapeutics (Review). Int J Mol
Med. 23:626–639. 2018.
|
|
36
|
Ishiwata T: Role of fibroblast growth
factor receptor-2 splicing in normal and cancer cells. Front Biosci
(Landmark Ed). 23:626–639. 2018. View
Article : Google Scholar : PubMed/NCBI
|