Introduction
Osteosarcoma is the third most common cancer in
children and young adults, and the most common primary malignant
bone tumor in the pediatric and adult populations. It is
characterized by the proliferation of tumor cells producing osteoid
or immature bone matrix (1,2). Approximately 15–20% of osteosarcoma
patients present with pulmonary metastasis at the time of
diagnosis, exhibiting an extremely poor 5-year survival rate.
Despite the availability of multimodality treatment, numerous
challenges remain in the clinical treatment of osteosarcoma
patients, and the majority of chemotherapeutic drugs have a limited
effect on osteosarcoma due to side effects and development of drug
resistance. The poor prognosis of osteosarcoma patients has not
improved over the three decades following the advent of modern
chemotherapy (3–5). Therefore, there is an urgent need to
develop novel therapeutic agents against osteosarcoma.
Apoptosis, regulated by complex signaling pathways,
is essential for normal tissue development (6). The abnormal expression of pro- or
anti-apoptotic genes is believed to be associated with a number of
pathologic disorders, including cancer (7,8). As a
major signaling cascade for apoptosis, the intrinsic mitochondrial
pathway serves crucial roles in the regulation of apoptotic
processes in various cancer cells (9). Studies have demonstrated that
persistent endoplasmic reticulum (ER) stress was associated with
the activation of the intrinsic mitochondrial apoptosis pathway
(10). ER stress is activated
through three sensors: Glucose-regulated protein 78 (GRP78),
inositol-requiring enzyme 1 (IRE1) and activating transcription
factor 6 (ATF6) (11). Once the
sensors are released from GRP78 (also known as binding
immunoglobulin protein), certain downstream effectors, such as
C/EBP homologous protein, are activated to trigger pro-apoptotic
signals by targeting several apoptotic genes (12,13).
Natural products isolated from plants have recently
attracted great interest due to their potent biological and
pharmaceutical activities. A novel compound known as (3R)-
5,6,7- trihydroxy-3-isopropyl-3-methylisochroman-1-one (TIM),
isolated from the whole plants of Selaginella moellendorffii
Hieron., has been reported to inhibit the proliferation of colon
cancer cells by inducing apoptosis (14). In the present study, TIM was further
studied to investigate its effects on osteosarcoma. The study
results revealed that TIM exhibited potent antitumor activities, as
evidenced by its inhibitory effect on osteosarcoma growth in a
xenograft tumor model. These results provided insights that may
facilitate the development of TIM as a new potential therapeutic
agent against osteosarcoma.
Materials and methods
Cell lines, reagents and
chemicals
The normal osteoblast cell line MC3T3-E1, breast
adenocarcinoma cell line MCF-7, pancreatic adenocarcinoma cell line
PANC-1, colon adenocarcinoma cell line HT-29, stomach
adenocarcinoma cell line BGC-823, lung adenocarcinoma cell line
A549, hepatoblastoma cell line HepG2, glioblastoma cell line U-251
MG, epidermoid carcinoma cell line A431, and osteosarcoma cell
lines U2OS, MG63, 143B and SaOS-2 were purchased from Shanghai Cell
Bank (Shanghai, China). Cell culture media and fetal bovine serum
(FBS) were purchased from Gibco (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The fluorescent probe
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine
iodide (JC-1) was purchased from Molecular Probes (Eugene, OR,
USA). The
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium-bromide (MTT)
was purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
The Annexin V-FITC/propidium iodide (PI) apoptosis detection kit
was purchased from BIO-BOX Biotech (Nanjing, China), while the
cytochrome c immunoassay kit was from R&D Systems, Inc.
(Minneapolis, MN, USA). iScript™ Reverse Transcription
Supermix for RT-qPCR (cat. no. 170-88400) was purchased from
Bio-Rad Laboratories, Inc. (Hercules, CA, USA). SYBR™
Green Master Mix kit for PCR (cat. no. 4309155) was purchased from
Thermo Fisher Scientific, Inc. Doxorubicin and Paclitaxel were
purchased from Shijiazhuang Pharmaceutical Co., Ltd. (Shijiazhuang,
China), and prostaglandin E1 (PGE1) was from Merck KGaA
(Calbiochem; La Jolla, CA, USA). All other solvents were of
analytical grade and were obtained from Sinopharm Chemical Reagent
Co., Ltd. (Shanghai, China). TIM was isolated and identified by
Professor Yi He from the Department of Pharmaceutical Science,
Hangzhou Vocational and Technical College (Hangzhou, China) for use
in the present study (14). TIM was
dissolved in DMSO for use in the in vitro experiments. For
the in vivo experiments, doxorubicin was freshly prepared
with PBS and TIM was freshly prepared in 0.5% carboxymethyl
cellulose prior to use in the in vivo experiments.
Animals
Female nude mice (BALB/c-nu/nu; age, 6 weeks) were
obtained from Vital River Laboratory Animal Technology Co., Ltd.
(Beijing, China), and housed in a sterile environment at 25°C on a
12-h light/dark cycle. The mice had free access to food and water
during the entire experimental period. All animal procedures were
conducted in accordance with the Chinese Legislation Guide for the
Care and Use of Laboratory Animals, and the study was approved by
the Institutional Animal Care and Use Committee of Qingdao
University Medical College (Yantai, China).
Cell culture, treatment and viability
assay
MC3T3-E1, MCF-7, PANC-1, HepG2, A431, MG63 and 143B
cells were cultured in Dulbecco's modified Eagle's medium, and
HT-29, BGC-823, A549, U-251 MG, U2OS and SaOS-2 cells were cultured
in RPMI 1640 medium. Both media were supplemented with 10% FBS and
100 µg/ml penicillin/streptomycin at 37°C in an atmosphere
containing 5% CO2. For the IC50 assay, cells
were treated with Paclitaxel (1, 5, 25, 125, 625, 3,125 or 5,000
nM) or TIM (1, 5, 25, 125, 625, 3,125 or 5,000 nM) alone for 48 h.
Following the IC50 assay, U2OS cells were treated with
TIM (4 nM) together with PGE1 (100 nM) for 48 h. Cells
treated with DMSO were used as control. Subsequently, the cell
viability was measured by an MTT assay. Briefly, treated cells were
seeded into 96-well plates at a density of 4×104/100
µl/well. MTT solution (10 µl) was added to each well and incubated
at 37°C for 4 h. Then 200 µl of DMSO was added into each well to
dissolve the formazan. The absorbance was measured at 570 nm. The
IC50 value was calculated with GraphPad Prism software
(version 8; GraphPad Software, Inc.).
For subsequent experiments except for the xenograft
tumor model establishment and terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay, U2OS cells were treated with
TIM (1, 2 or 4 nM) for 48 h. Cells treated with DMSO were used as
control.
Cytochrome c assay
U2OS cells were fractionated with Fractionation kit
(cat. no. 78840; Thermo Fisher Scientific, Inc.) subsequent to TIM
treatment. The cytochrome c release was measured using a
cytochrome c quantikine ELISA kit (cat. no. DCTC0; R&D Systems,
Inc.), according to the manufacturer's protocol.
Mitochondrial membrane potential (MMP)
measurement
MMP was measured using the fluorescent probe JC-1.
Briefly, following the indicated treatments, U2OS cells were
incubated with JC-1 for 15 min at 37°C in the dark. Subsequent to
rinsing twice with PBS, the red/green fluorescence intensity was
determined with a fluorescence microplate reader (Tecan Polarion;
Tecan Group, Ltd., Mannedorf, Switzerland) at an excitation
wavelength of 490 nm and emission wavelength of 530/590 nm,
respectively.
Caspase activity measurement
Following the indicated treatments, U2OS cells were
lysed with the fractionation kit. Next, the caspase-3/9 activities
were determined using the assay kits (cat. no. K106-100 for
caspase-3; cat. no. K119-110 for caspase-9; R&D Systems, Inc.)
according to the manufacturer's protocol, followed by examination
with a microplate reader.
DNA fragmentation measurement
Subsequent to the indicated treatments, U2OS cells
were lysed with the fractionation kit, and DNA fragmentation was
measured by the Cell Death Detection ELISAplus kit (cat.
no. 11544675001; Roche Diagnostics) according to the manufacturer's
protocol. The absorbance was measured with a microplate reader at
405 nm.
Cell cycle distribution
measurement
Subsequent to the indicated treatments, U2OS cells
were washed three times with PBS and fixed with 70% ethanol for 30
min at 4°C. Next, the cells were incubated with PI/Triton X-100 for
10 min, and the cell cycle distribution was examined by flow
cytometry (BD Biosciences, San Jose, CA, USA). The ratio of G1/S
was calculated, and changes in cell cycle distribution were
expressed as a percentage of the control, which was set to
100%.
Apoptosis analysis
For apoptosis analysis, a total of 2×106
U2OS cells were harvested, washed with pre-chilled PBS, and then
re-suspended in 500 µl binding buffer. Subsequently, 5 µl Annexin
V-FITC/PI were added to each sample, and incubated for 10 min in
the dark at room temperature. Analysis of apoptotic cells was
performed using a FACScan flow cytometer (BD Biosciences).
RT-qPCR
Total RNA was extracted with the Qiagen RNeasy
reagents (Qiagen GmbH, Hilden, Germany) following manufacturer's
protocol and the RNA concentration was measured with NanoDrop
Spectrophotometer. RNA (0.5 µg) was used to generate cDNA using an
iScript™ Reverse Transcription Supermix kit; the RT
protocol was as follows: 25°C for 5 min, 46°C for 20 min and 95°C
for 1 min. qPCR was run on a StepOne system (Thermo Fisher
Scientific, Inc.) using SYBR Green Supermix (Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows:
95°C for 10 min, and 40 cycles of 95°C for 15 sec and 60°C for 60
sec. The mRNA levels were quantified with the comparative
Cq value method (15) and
normalized to β-actin levels. The gene-specific primer
sequences were as follows: Caspase-3 forward, 5-ttg tgg aat
tga tgc gtg at-3 and reverse, 3-ggc agg cct gaa taa tga aa-5
(GenBank reference: AJ413269.1); Caspase-9 forward, 5-cag
gag aaa ggc ctc agt tg-3 and reverse, 3-gga tgt agc cgt gtg acc
tt-5 (GenBank reference: AY214168.1); Ire1 forward, 5-cgg
cct ttg cag ata gtc tc-3 and reverse, 3-act gtc cac agt cac cac
ca-5 (GenBank reference: AF059198.1); Atf6 forward, 5-tga
act tcg agg atg ggt tc-3 and reverse, 3-tca ctc cct gag ttc ctg
ct-5 (GenBank reference: AB015856.1); Grp78 forward, 5-aaa
gga cag gct ggt gct aa-3 and reverse, 3-ggg ctg gag tac agt ggt
gt-5 (GenBank reference: BC020235.1); β-actin forward, 5-aga
gct acg agc tgc ctg ac-3 and reverse, 3-agc act gtg ttg gcg tac
ag-5 (GenBank reference: AK222925.1).
Western blot analysis
In order to obtain the total protein, cells were
lysed with th fractionation kit and centrifuged at 6,000 × g at 4°C
for 15 min, following which protein concentration was determined by
the BCA method. Next, 40 µg protein samples were subjected to
sodium dodecyl sulfate-polyacrylamide gel electrophoresis with
4–12% gels and then transferred to polyvinylidene difluoride
membranes. The membranes were blocked with 1% bovine serum albumin
(cat. no. A9306; Sigma-Aldrich; Merck KGaA) for 1 h at room
temperature and then incubated at 4°C overnight with the following
primary antibodies (Abs): Anti-cleaved caspase-3 rabbit monoclonal
(m)Ab (1:1,000; cat. no. 9664; Cell Signaling Technology, Inc.);
anti-cleaved caspase-9 rabbit polyclonal (p)Ab (1:500; cat. no.
C7729; Sigma-Aldrich; Merck KGaA); anti-Bcl-2 rabbit mAb (1:1,000;
cat. no. ab32124; Abcam); anti-Bax rabbit mAb (1:1,000; cat. no.
ab182733; Abcam); anti-myeloid cell leukemia 1 (MCL-1) rabbit mAb
(1:2,000; cat. no. ab32087; Abcam); anti-NOXA rabbit pAb (1:500;
cat. no. ab36833; Abcam); anti-IRE1 (phosphor S724) rabbit pAb
(1:1,000; cat. no. ab48187; Abcam); anti-ATF6 rabbit pAb (1:500;
cat. no. ab37149; Abcam); anti-GRP78 rabbit mAb (1:5,000; cat. no.
ab108615; Abcam); anti-β-actin rabbit mAb (1:5,000; cat. no.
ab179467; Abcam). Membranes were then incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibodies
(1:8,000; cat. no. ab6721; Abcam) for 1 h at room temperature. The
bands were then detected with an ECL system (EMD Millipore,
Billerica, MA, USA). The bands were quantified with ImageJ software
(version 1.4; National Institutes of Health, Bethesda, MD,
USA).
Xenograft tumor model
A total of 5×106 U2OS cells were injected
subcutaneously into the right flank of nude mice. When the tumor
volume reached 50–100 mm3, the tumor-bearing mice were
divided into four groups (5 mice per group), and administered
different treatments. One of the groups was intraperitoneally
injected with doxorubicin (2 mg/kg) twice weekly, while two other
mouse groups received intragastric treatment with TIM (1 and 2
mg/kg), 5 days per week. Mice treated with PBS served as the
negative controls. Once the tumor volume reached 1,500
mm3 (after ~3 weeks), all tumor-bearing mice were
sacrificed to collect the tumor tissue to record the tumor weight
and detect apoptosis.
TUNEL assay
Tumor samples were fixed with 4% paraformaldehyde
for 4 h, and then dehydrated by graded sucrose solution.
Subsequently, the tumor samples were embedded in Tissue-Tek Optimal
Cutting Temperature compound (Sakura, Alphen aan den Rijn, The
Netherlands), and 10-µm frozen sections were cut sagittally with a
freezing microtome. Apoptosis was detected by the In Situ
Cell Death Detection kit (Roche Applied Science, IN, USA). Briefly,
frozen sections were incubated with permeabilization solution for 2
min on ice. After washing, the sections were incubated with the
TUNEL reaction mixture for 60 min at 37°C in the dark and then
rinsed with PBS. Subsequently, the sections were sealed with
VECTASHIELD mounting medium with DAPI (Vector Laboratories, Inc.,
Burlingame, CA, USA) and visualized under the Olympus BX60
microscope (Olympus Corporation, Shinjuku, Japan).
Statistical analysis
Data are expressed as the mean ± standard deviation,
and the software SPSS (version 12.0; SPSS, Inc., Chicago, IL, USA)
was used for statistical analysis. Data were analyzed by one-way
analysis of variance, followed by the least significant difference
test. A P-value of <0.05 was considered as an indicator of a
statistically significant difference.
Results
TIM inhibits cell proliferation
To explore the anti-tumor spectrum of TIM, MTT assay
was used to determine the anti-proliferative activities of TIM on
different cancer cell lines. Lower IC50 values indicated
stronger anti-proliferative activities. As shown in Table I, TIM selectively inhibited the
proliferation of colon carcinoma and osteosarcoma cells.
Furthermore, TIM exhibited low toxicity on the normal osteoblast
cells MC3T3-E1 (Table I).
 | Table I.Anti-proliferative effects of TIM on
various cancer cells. |
Table I.
Anti-proliferative effects of TIM on
various cancer cells.
| Cell line | Origin | TIM (IC50;
nM) | Paclitaxel
(IC50; nM) |
|---|
| MC3T3-E1 | Normal osteoblast
cell line | >5,000 | >5,000 |
| MCF-7 | Breast
adenocarcinoma | 2,561.5±189.3 |
6.1±1.2 |
| PANC-1 | Pancreatic
adenocarcinoma |
155.7±14.3 | 15.3±1.7 |
| U2OS | Osteosarcoma |
8.1±1.5 | 16.7±2.9 |
| MG63 | Osteosarcoma |
9.3±1.8 | 12.5±2.2 |
| 143B | Osteosarcoma |
11.5±1.9 | 19.2±3.5 |
| SaOS-2 | Osteosarcoma |
15.4±2.6 | 11.3±2.1 |
| HT-29 | Colon
adenocarcinoma |
9.5±1.7 | 11.2±3.0 |
| BGC-823 | Stomach
adenocarcinoma | 1,332.4±127.6 | 16.6±2.3 |
| A549 | Lung
adenocarcinoma |
44.5±5.1 |
9.9±1.8 |
| HepG2 | Hepatoblastoma | 1,851.5±163.2 | 35.5±3.3 |
| U-251 MG | Glioblastoma | 1,305.1±112.3 | 11.3±2.2 |
| A431 | Epidermoid
carcinoma |
19.3±2.8 | 17.5±1.9 |
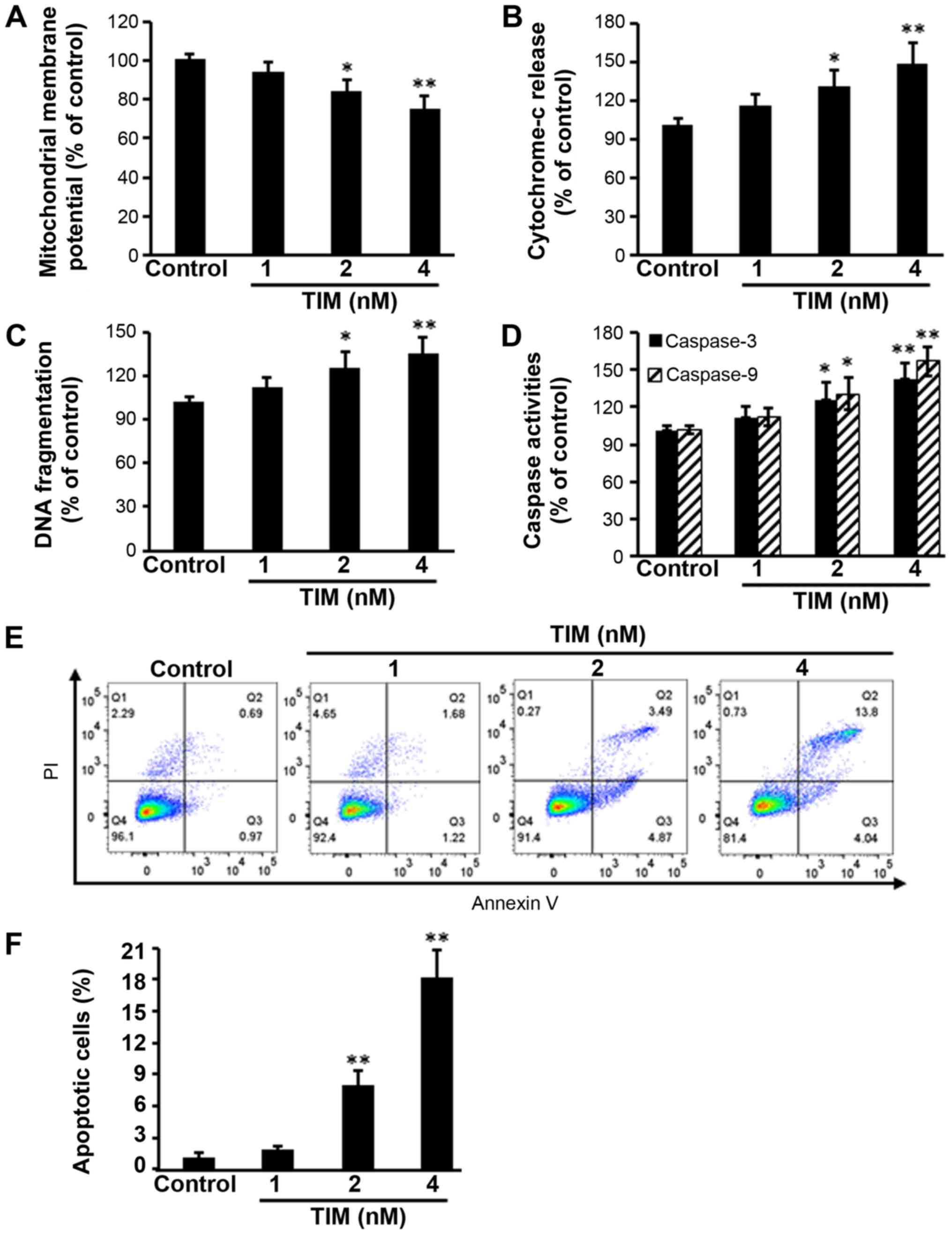
TIM induces the apoptosis of U2OS
cells
U2OS osteosarcoma cells were subjected to TIM
treatment at concentrations of 1, 2 and 4 nM, and then a number of
apoptosis-associated markers were detected. Comparing with the
control group, TIM treatment at concentrations of 2 and 4 nM
significantly decreased the MMP (Fig.
1A), while it markedly increased the cytochrome c
release (Fig. 1B), DNA fragmentation
(Fig. 1C) and Caspase-3/9 activities
(Fig. 1D). In addition, TIM
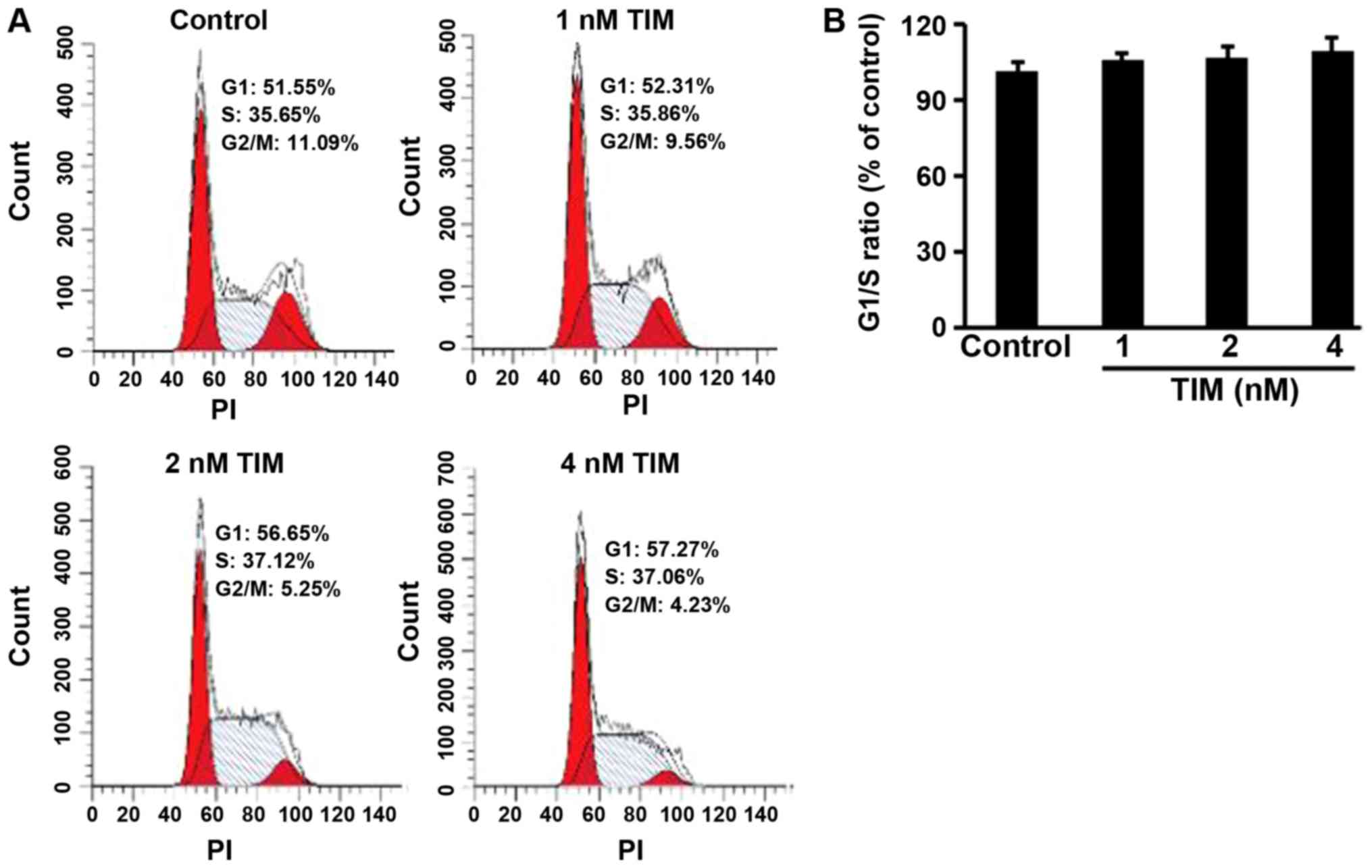
treatment (2 and 4 nM) induced apoptosis in U2OS cells (Fig. 1E and F). However, as shown in
Fig. 2, TIM treatment did not
significantly alter the cell cycle distribution. Comparing with the
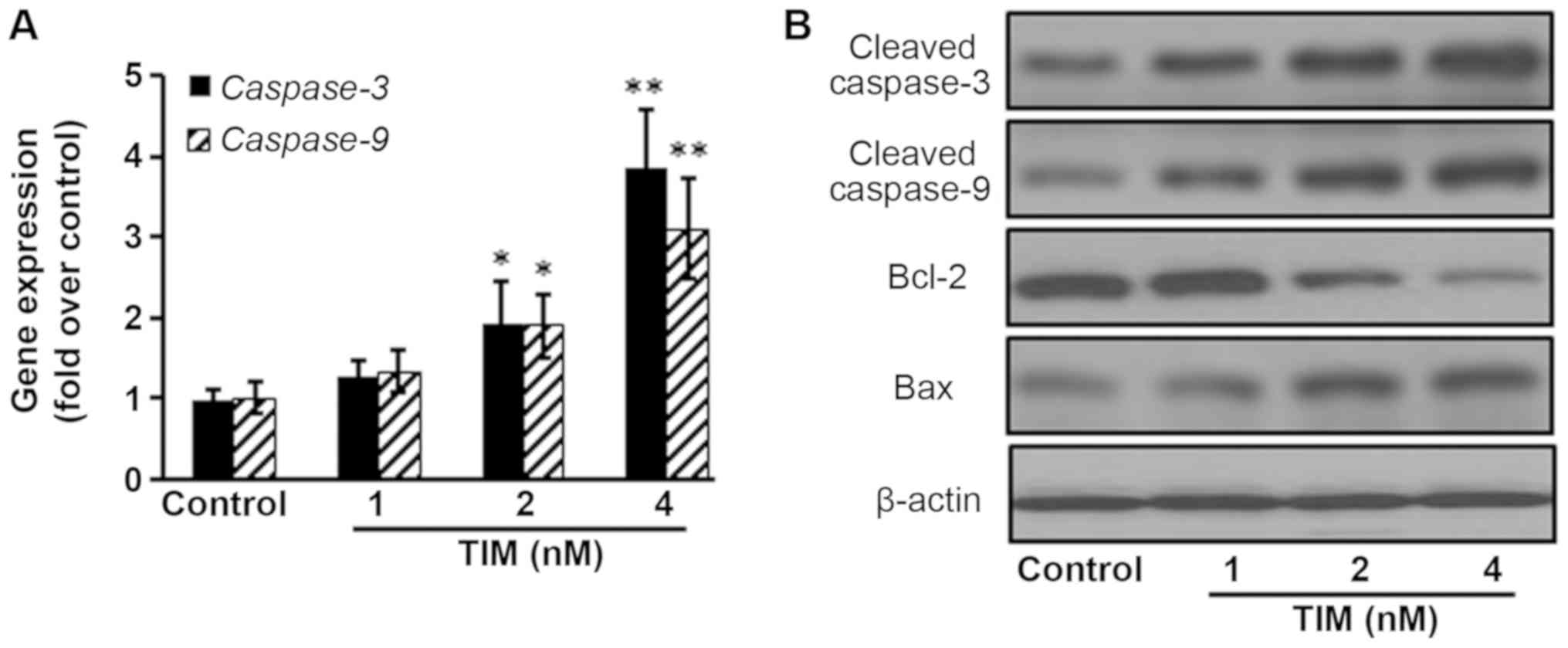
control group, pro-apoptotic gene (Caspase-3 and
Caspase-9) expression levels were significantly increased
following 4 nM TIM treatment. Pro-apoptotic protein (Cleaved
caspase-3, Cleaved caspase-9 and Bax) expression levels were
markedly increased following TIM treatment, whereas anti-apoptotic
protein (Bcl-2) expression was evidently decreased (Fig. 3).
TIM induces apoptosis through the
NOXA/MCL-1 axis and by triggering ER stress
The NOXA/MCL-1 axis is involved in
chemotherapeutic-induced apoptosis in several tumor types (16). The present study results demonstrated
that the protein expression of MCL-1 was decreased following TIM
treatment, whereas the protein expression of NOXA was upregulated.
Addition of the MCL-1 stabilization agent PGE1
significantly antagonized the anti-proliferative effects of TIM on
U2OS cells, indicating that the NOXA/MCL-1 axis was involved in
TIM-induced apoptosis in U2OS cells (Fig. 4A and B). Furthermore, TIM treatment
triggered ER stress, as evidenced by the upregulation of the gene
and protein expression levels of ER stress markers IRE1, ATF6 and
GRP78 (Fig. 4C and D).
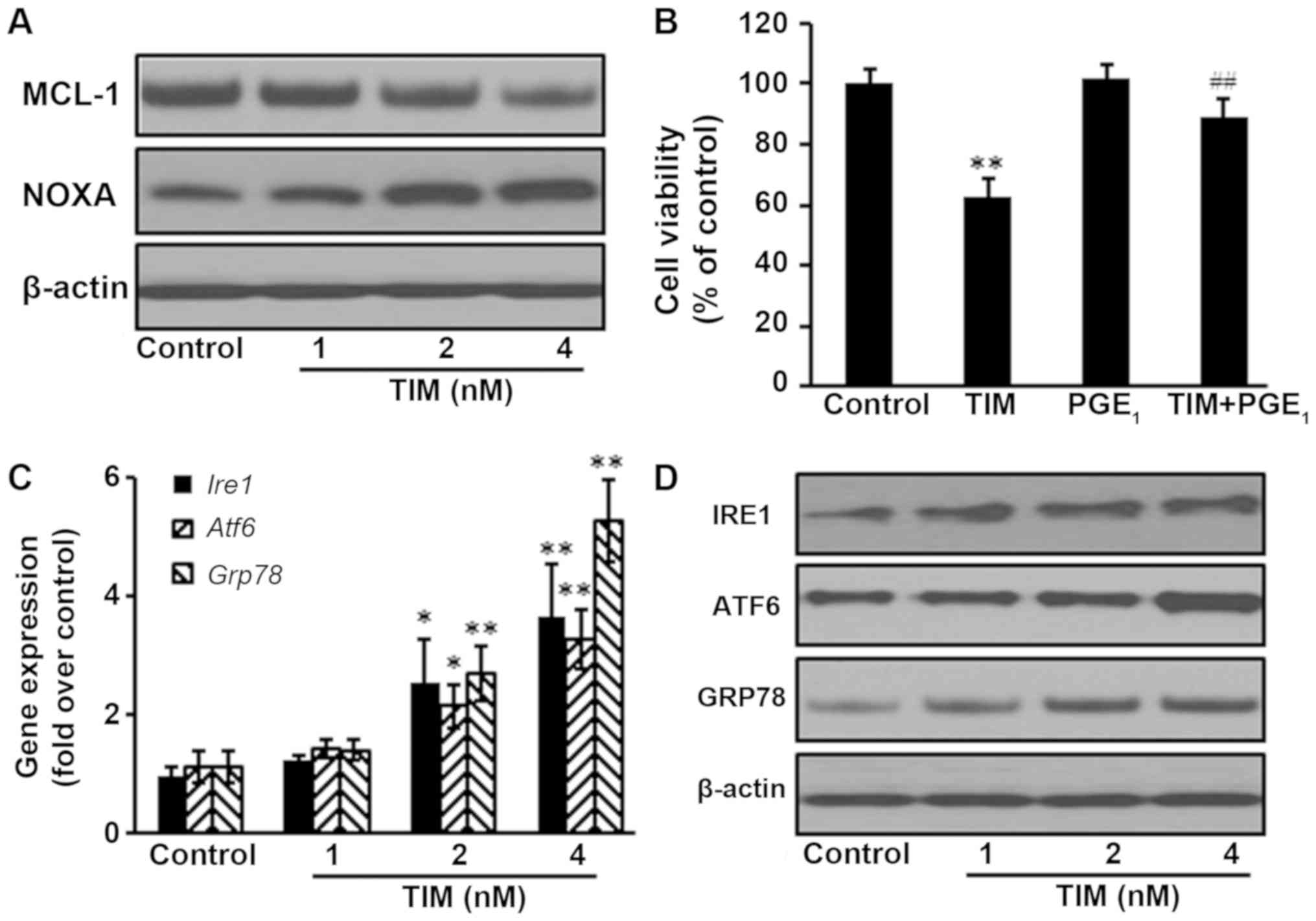 | Figure 4.TIM induced apoptosis through the
NOXA/MCL-1 axis and triggering ER stress. U2OS cells were treated
with 4 nM TIM for 48 h. (A) TIM treatment decreased the protein
expression of MCL-1, but increased the protein expression of NOXA.
(B) PGE1 (100 nM) inhibited the anti-proliferative effects of TIM.
(C) Gene and (D) protein expression levels of the ER stress-marker
proteins IRE1, ATF6 and GRP78 were increased by TIM. Data are
expressed as the mean ± standard deviation (n=3). *P<0.05 and
**P<0.01, vs. control group; ##P<0.01, vs. TIM
alone. TIM,
(3R)-5,6,7-trihydroxy-3-isopropyl-3-methylisochroman-1-one;
ER, endoplasmic reticulum; GRP78, glucose-regulated protein 78;
IRE1, inositol-requiring enzyme 1; ATF6, activating transcription
factor 6; PGE1, prostaglandin E1. |
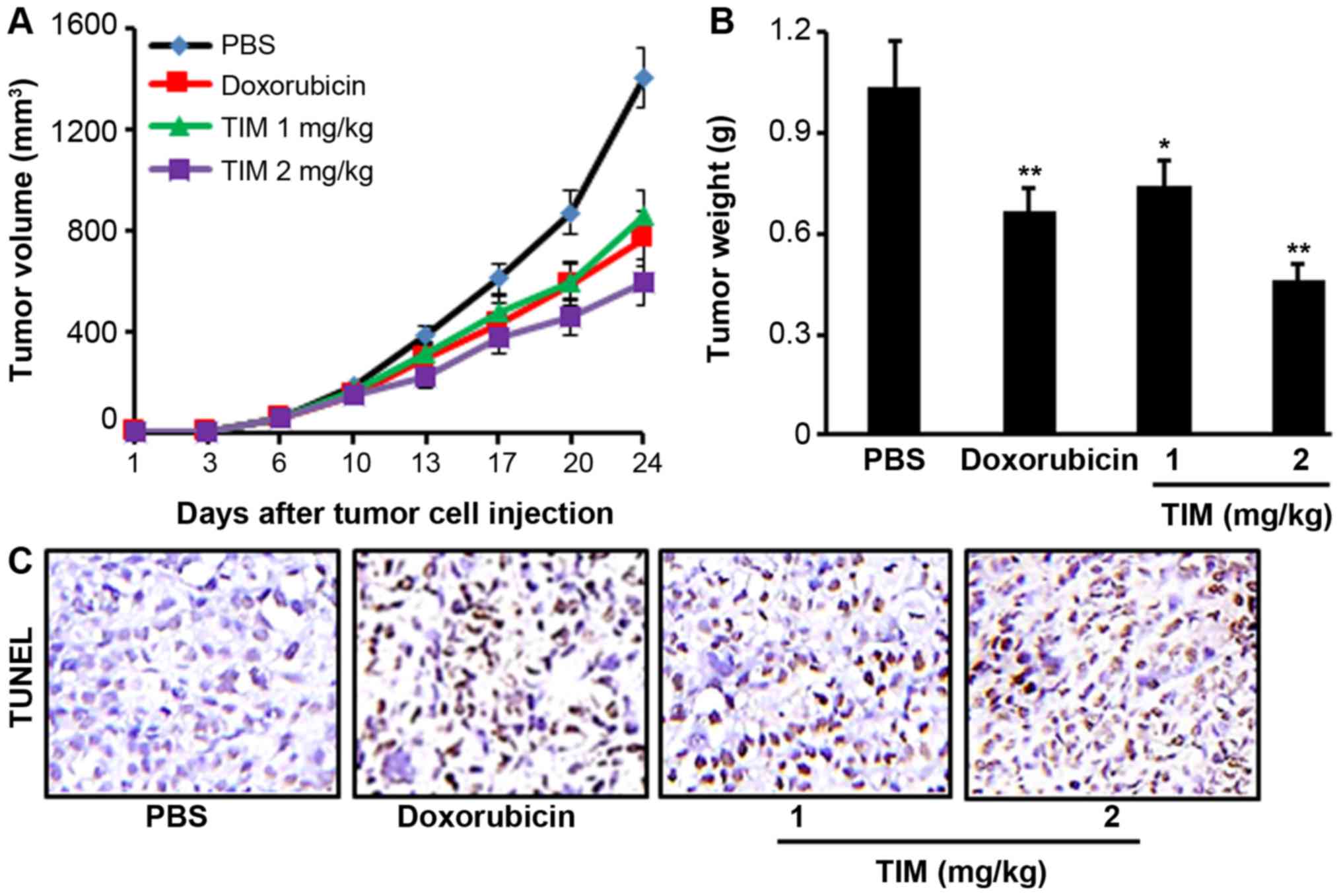
TIM inhibits tumor growth in a
xenograft mouse model
Tumor-bearing mice were treated with TIM at a
concentration of 1 or 2 mg/kg for 24 days. It was observed that TIM
treatment at 2 mg/kg significantly inhibited the U2OS tumor growth,
as indicated by the reduction in tumor volume and weight, which
showed stronger anti-tumor activities than the positive control
drug, doxorubicin (Fig. 5A and B).
Furthermore, TIM treatment was found to significantly increase the
DNA fragmentation, as indicated by the TUNEL assay results
(Fig. 5C).
Discussion
Current treatments for osteosarcoma consist of
multi-agent chemotherapy, followed by complete surgical resection,
radiation therapy and surveillance for lung metastasis. However,
patients with metastasis at the time of diagnosis exhibit a poor
response to conventional therapies due to high tumor malignancy and
inadequate treatment efficacy (17,18).
Therefore, it is of utmost importance to search for effective novel
agents and treatments. Herbal compounds are becoming an important
source of medicinal products. In the present study, it was reported
that TIM, a natural compound extracted from whole plants of
Selaginella moellendorffii Hieron., inhibited osteosarcoma
growth by inducing apoptosis.
The intrinsic mitochondrial pathway is a major
signaling cascade for apoptosis. The role of mitochondria during
the process of cell death regulation is crucial, and the decrease
of MMP would induce the release of cytochrome c from
mitochondria to the nucleus, activating caspase-associated
apoptotic proteins (19,20). Caspase-3 and caspase-9 activate
endonucleases to cleave nuclear DNA and ultimately lead to DNA
fragmentation (21). The current
study results revealed that TIM decreased the MMP, while it
increased the cytochrome c release, DNA fragmentation, and
activities of caspase-3 and caspase-9. Furthermore, apoptosis
significantly increased following TIM treatment. It was observed
that the expression of the anti-apoptotic protein B-cell lymphoma 2
(Bcl-2) decreased, whereas the pro-apoptotic protein
Bcl-2-associated X protein (Bax) and caspase expression levels
increased upon TIM treatment.
Members of the Bcl-2 family serve key roles in the
regulation of apoptotic processes in numerous cancer cells
(9). NOXA, a crucial pro-apoptotic
protein in the Bcl-2 family, interacts with the anti-apoptotic
Bcl-2 protein MCL-1, interfering with the polymerization of Bax and
Bak to trigger apoptosis (22,23). The
NOXA/MCL-1 axis has been reported to be involved in apoptosis
induced by chemotherapeutics in a number of tumor models (9,23). In
the present study, the expression of MCL-1 decreased upon TIM
treatment, while the pro-apoptotic protein NOXA was upregulated.
Furthermore, the addition of PGE1, an MCL-1
stabilization agent (24),
antagonized the anti-proliferative effects of TIM on U2OS cells.
These results indicated that TIM induced intrinsic mitochondrial
apoptosis through the NOXA/MCL-1 axis in osteosarcoma cells.
Various chemotherapeutics activate ER response while
inducing apoptosis in cancer cells (25). As a central regulator of ER stress to
control the activation of transmembrane ER stress sensors (IRE1 and
ATF6), GRP78 has been established by a large number of studies as a
marker of ER stress (26). As
signaling proteins in ER stress, IRE1 activates the
apoptosis-signaling kinase-1, which further activates downstream of
stress kinases to promote apoptosis, and ATF6 translocates as a
transcriptional factor to the Golgi compartment where it is cleaved
upon ER stress (27). The current
study results demonstrated that ER stress marker proteins were
upregulated upon TIM treatment, suggesting that TIM triggered ER
stress in osteosarcoma cells.
However, in the current study, only one osteosarcoma
cell line was analyzed in detail, which poses a limitation. Thus,
further studies are required to validate the effects of TIM on
osteosarcoma.
In conclusion, the present study demonstrated that
TIM, isolated from whole plants of Selaginella
moellendorffii Hieron., inhibited osteosarcoma growth, by
inducing apoptosis via the NOXA/MCL-1 axis and ER stress. The
results provide novel insight that may assist in the development of
TIM as a therapeutic agent against osteosarcoma.
Acknowledgements
The authors thank Professor Yi He from Department of
Pharmaceutical Science, Hangzhou Vocational and Technical College
for providing TIM in the current study.
Funding
No funding received.
Availability of data and materials
The data are available from the corresponding author
upon reasonable request.
Authors' contributions
LJL design and conceived in the current study. MZS
and FLZ performed the studies and analyzed the data. LJL, MZS and
FLZ prepared the manuscript.
Ethics approval and consent to
participate
All animal experiments were performed in compliance
with the Chinese legislation on the use and care of laboratory
animals, and the study was approved by the Ethical Committee on
Animal Care and Use of Qingdao University Medical College (Yantai,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lussier DM, Johnson JL, Hingorani P and
Blattman JN: Combination immunotherapy with α-CTLA-4 and α-PD-L1
antibody blockade prevents immune escape and leads to complete
control of metastatic osteosarcoma. J Immunother Cancer. 3:212015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li Y, Du W, Han J and Ge J: LAMP3 promotes
the invasion of osteosarcoma cells via SPP1 signaling. Mol Med Rep.
16:5947–5953. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carrle D and Bielack S: Osteosarcoma lung
metastases detection and principles of multimodal therapy. Cancer
Treat Res. 152:165–184. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Harting MT and Blakely ML: Management of
osteosarcoma pulmonary metastases. Semin Pediatr Surg. 15:25–29.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Strauss SJ, Ng T, Mendoza-Naranjo A,
Whelan J and Sorensen PH: Understanding micrometastatic disease and
Anoikis resistance in ewing family of tumors and osteosarcoma.
Oncologist. 15:627–635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Feng Y, Yang Y, Fan C, Di S, Hu W, Jiang
S, Li T, Ma Z, Chao D, Feng X, et al: Pterostilbene inhibits the
growth of human esophageal cancer cells by regulating endoplasmic
reticulum stress. Cell Physiol Biochem. 38:1226–1244. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Uemura N and Kondo T: Current advances in
esophageal cancer proteomics. Biochim Biophys Acta. 1854:687–695.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang Q, Mas A, Diamond MP and Al-Hendy A:
The mechanism and function of epigenetics in uterine leiomyoma
development. Reprod Sci. 23:163–175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao X, Kong F, Wang L and Zhang H: c-FLIP
and the NOXA/Mcl-1 axis participate in the synergistic effect of
pemetrexed plus cisplatin in human choroidal melanoma cells. PLoS
One. 12:e01841352017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang H, Yang Y, Chen H, Dan J, Cheng J,
Guo S, Sun X, Wang W, Ai Y, Li S, et al: The predominant pathway of
apoptosis in THP-1 macrophage-derived foam cells induced by
5-aminolevulinic acid-mediated sonodynamic therapy is the
mitochondria-caspase pathway despite the participation of
endoplasmic reticulum stress. Cell Physiol Biochem. 33:1789–1801.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Beck D, Niessner H, Smalley KS, Flaherty
K, Paraiso KH, Busch C, Sinnberg T, Vasseur S, Iovanna JL, Drieben
S, et al: Vemurafenib potently induces endoplasmic reticulum
stress-mediated apoptosis in BRAFV600E melanoma cells. Sci Signal.
6:ra72013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee JH, Kwon EJ and Kim DH: Calumenin has
a role in the alleviation of ER stress in neonatal rat
cardiomyocytes. Biochem Biophys Res Commun. 439:327–332. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chow SE, Kao CH, Liu YT, Cheng ML, Yang
YW, Huang YK, Hsu CC and Wang JS: Resveratrol induced ER expansion
and ER caspase-mediated apoptosis in human nasopharyngeal carcinoma
cells. Apoptosis. 19:527–541. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou XF, Tong GT, Wang XW and He Y:
Anti-proliferative constituents from Selaginella
moellendorffii. Nat Prod Commun. 11:623–626. 2016.PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hauck P, Chao BH, Litz J and Krystal GW:
Alterations in the Noxa/Mcl-1 axis determine sensitivity of small
cell lung cancer to the BH3 mimetic ABT-737. Mol Cancer Ther.
8:883–892. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saraf AJ, Fenger JM and Roberts RD:
Osteosarcoma: Accelerating progress makes for a hopeful future.
Front Oncol. 8:42018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Z, Lu W, Li Y and Tang B: Alpinetin
promotes Bax translocation, induces apoptosis through the
mitochondrial pathway and arrests human gastric cancer cells at the
G2/M phase. Mol Med Rep. 7:915–920. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xue S, Chen YX, Qin SK, Yang AZ, Wang L,
Xu HJ and Geng HY: Raltitrexed induces mitochondrial-mediated
apoptosis in SGC7901 human gastric cancer cells. Mol Med Rep.
10:1927–1934. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xin BR, Liu JF, Kang J and Chan WP: (2R,
3S)-pinobanksin- 3-cinnamate, a new flavonone from seeds of
Alpinia galanga willd., presents in vitro neuroprotective
effects. Mol Cell Toxicol. 10:165–172. 2014. View Article : Google Scholar
|
|
22
|
Zhao X, Liu X and Su L: Parthenolide
induces apoptosis via TNFRSF10B and PMAIP1 pathways in human lung
cancer cells. J Exp Clin Cancer Res. 33:32014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu X, Lv Z, Zou J, Liu X, Ma J, Wang J,
Sa N, Jing P and Xu W: Afatinib down-regulates MCL-1 expression
through the PERK-eIF2α-ATF4 axis and leads to apoptosis in head and
neck squamous cell carcinoma. Am J Cancer Res. 6:1708–1719.
2016.PubMed/NCBI
|
|
24
|
Kato T, Kutsuna H, Oshitani N and Kitagawa
S: Cyclic AMP delays neutrophil apoptosis via stabilization of
Mcl-1. FEBS Lett. 580:4582–4586. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qin L, Wang Z, Tao L and Wang Y: ER stress
negatively regulates AKT/TSC/mTOR pathway to enhance autophagy.
Autophagy. 6:239–247. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee AS: The ER chaperone and signaling
regulator GRP78/BiP as a monitor of endoplasmic reticulum stress.
Methods. 35:373–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sano R and Reed JC: ER stress-induced cell
death mechanisms. Biochim Biophys Acta. 1833:3460–3470. 2013.
View Article : Google Scholar : PubMed/NCBI
|