Introduction
Prostate cancer (PCa) is the most common malignancy
and the leading cause of cancer mortality among men in the USA and
Europe (1,2). The male population with a median age
>60 years is more susceptible to PCa (3,4). In
total, there are >1 million newly diagnosed cases of PCa
identified each year and ~300,000 cases succumb to the disease
(5). The incidence and mortality
rates of PCa vary due to genetic variation and environmental
changes between different age and ethnic groups (3,5–7). Exercise, obesity, smoking, vitamins,
micronutrients, metformin, statins and other medications are all
considered to be critical factors, which can affect the incidence
and mortality rates of PCa (5,6). In
addition, the incidence rate increases significantly in urban areas
of China, South Korea and other countries (1,3,6).
Testosterone serves a key role in the development,
growth and maintenance of the prostate (8,9) and
abnormal prostate cell proliferation and differentiation caused by
hormonal regulation may be involved in the initiation of
hypertrophy, hyperplasia and malignancy (8,9). Many
growth factors, including epidermal growth factor, fibroblast
growth factor (FGF), insulin-like growth factor, transforming
growth factor β1 and keratinocyte growth factor are also involved
(8,10). FGFs can activate FGF receptors
(FGFRs), which can lead to wound repair and neovascularization, as
well as tumor growth and differentiation, thereby making FGF a
potential therapeutic target in the development of novel
anti-cancer treatments (11,12).
Early diagnosis of PCa is key to reducing morbidity
and mortality (8). The main
treatment options for early-stage PCa involve monitoring the
prostate, whereas late-stage PCa requires radiation, surgery and
other treatments, such as systemic chemotherapies or targeted
therapies (8). Radical prostatectomy
followed by radiotherapy in combination with hormonal treatment is
widely used to treat patients with recurrent PCa (8,13).
Furthermore, treatment outcome varies dramatically as clinical
heterogeneity is another feature associated with PCa (8,13).
microRNAs (miRNAs or miRs) are single-stranded small
RNAs, 18–23 nucleotides in length which can regulate gene
expression through mRNA degradation or protein synthesis inhibition
(14,15). miRNAs can regulate several
physiological or pathological processes including cell
proliferation, differentiation, stress and death (14,15).
miRNAs have been identified in several types of cancer and can
function as positive or negative regulators, depending on their
target gene (16,17). miR-628, which is located at 15q21.3,
was initially identified in acute myeloid leukemia and is regulated
by interleukin-3, granulocyte-macrophage colony-stimulating factor
and granulocyte colony-stimulating factor (15). A previous study demonstrated that
miR-628 is an endotoxin-responsive gene induced by
lipopolysaccharide as well as a nuclear factor κB-dependent gene
(18). miR-628 can regulate
inflammatory responses by targeting a key adapter molecule
downstream of toll-like receptors (TLRs) to suppress TLR signaling
(18).
In addition, miR-628 mediates burn-induced skeletal
muscle atrophy through regulation of the insulin receptor substrate
1/protein kinase B serine/threonine kinase 1/forkhead box protein O
3a signaling pathway (19).
miR-628-3p is upregulated in patients with bone fractures and
miR-628-3p can exert an inhibitory effect on osteogenesis via the
suppression of runt related transcription factor 2 leading to
atrophic non-union, a complication associated with bone fractures
(20). Furthermore, miR-628-3p is
involved in rifampin-mediated cytochrome P450 family 3 subfamily A
member 4 (CYP3A4) induction as it is thought to directly target
CYP3A4 (21). miR-628-5p has
previously been demonstrated to decrease the stem-like cell
percentage of epithelial ovarian cancer (EOC) cells by inducing
their apoptosis as well as decreasing the tumorigenicity of EOC
cells by targeting FGFR2 (11).
A previous study identified miR-628 as a potential
non-invasive biomarker for PCa (14), however whether miR-628 serves a tumor
suppressing role in PCa remains unknown. In addition, whether FGFR2
is a direct target of miR-628 in PCa remains unknown. Therefore,
the aim of the present study was to investigate miR-628 expression
and its underlying mechanism in PCa cell proliferation and invasion
and the FGFR2 signaling pathway.
Materials and methods
PCa and healthy control patients
A total of 33 patients with PCa (males; age range,
55–89 years; average age, 70 years) were recruited in Wuhan Fourth
Hospital, Puai Hospital (Wuhan, China) between January 2016 and
December 2017 for inclusion in the current study. Patients were
diagnosed following prostate biopsy or postoperative pathological
examination. Patients with PCa with a Gleason score range from 4–10
were included in the study. In total, 12 patients had a Gleason
score of 4–6 (well differentiated), 9 patients had a Gleason score
of 7 (medium differentiated) and 12 patients had a Gleason score of
8–10 (low differentiated). A summary of the cancer staging and
Gleason score for patients with PCa is presented in Table I. In addition, 26 healthy control
patients (age range, 62–82 years; average age, 79 years) were
recruited following admittance to the hospital physical examination
center of Wuhan Fourth Hospital, Puai Hospital (Wuhan, China) for a
health check-up between January 2016 and December 2017. Blood
pressure, urine routine test, and liver and kidney function were
all within the normal range. Patients with other diseases affecting
heart, lungs or other vital organs, such as benign prostatic
hyperplasia and other prostate-associated diseases were excluded.
The current study was approved by the Ethics Committee of Wuhan
Fourth Hospital, Puai Hospital (Wuhan, China) and written informed
consent was obtained from all participants.
 | Table I.Serum levels of PSA, FGF1 and FGFR2
in patients with PCa. |
Table I.
Serum levels of PSA, FGF1 and FGFR2
in patients with PCa.
| Group | n | PSA (ng/l) | FGF1 (pg/ml) | FGFR2 (pg/ml) |
|---|
| Control | 26 | 1.03±0.62 | 203.72±25.27 | 64.14±11.11 |
| PCa | 33 | 21.65±12.18 | 304.02±45.79 | 88.26±10.09 |
| Gleason score |
|
4–6 | 12 | 13.42±5.69 | 304.93±57.13 | 88.60±12.92 |
| 7 | 9 | 20.87±12.86 | 318.36±29.26 | 88.40±8.19 |
|
8–10 | 12 | 30.46±10.97 | 292.36±43.54 | 87.03±9.24 |
| Clinical stage |
| T1 | 4 | 8.09±1.94 | 314.42±60.76 | 89.125±12.17 |
| T2 | 6 | 15.15±5.51 | 304.03±54.08 | 88.65±14.13 |
| T3 | 15 | 20.34±10.35 | 308.78±43.85 | 88.19±8.56 |
| T4 | 8 | 35.76±8.27 | 289.89±41.58 | 86.49±10.86 |
Specimen collection
Venous blood samples (4 ml) were collected from all
participants (PCa and control) in the fasting state prior to any
treatment, which included medication, hormonal treatment, surgery,
as well as rectal examination, prostate massage and puncture within
1 week before blood extraction. Blood was centrifuged at 1,200 × g
at room temperature for 12 min to collect serum. Serum
prostate-specific antigen was measured immediately using an ADVID
Centaur CP analyzer (Siemens Healthineers, Erlangen, Germany). The
remaining serum was stored at −20°C and used to determine serum
expression levels of miR-628, FGF-1 and FGFR2 using an
IMMULITE®1000 immunoassay system (Siemens
Healthineers).
Cell culture
Human prostatic adenocarcinoma cell line LNCaP
(ATCC® CRL-1740™) was purchased from the American Type
Culture Collection (Manassas, VA, USA). Cells were cultured in
RPMI-1640 medium (ATCC) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100
U/ml penicillin and 100 mg/ml streptomycin, and maintained at 37°C
in an atmosphere containing 5% CO2.
Cell transfection
MiR-628-5p mimic (5-AUG CUG ACA UAU UUA CUA GAGG-3;
cat. no. MCH03275) and negative control miRNA (cat. no. MCH00000),
as well as miR-628-5p inhibitor (cat. no. MIH03275) or negative
control inhibitor (cat. no. MIH00000) was purchased from Applied
Biological Materials Inc., (Richmond, BC, Canada). Lipofectamine
2000 (Thermo Fisher Scientific, Inc.) was used to transfect 35 nM
miR-628-5p mimic and negative control miRNA (negative control), as
well as 35 nM miR-628-5p inhibitor or negative control inhibitor
(negative control) into 105 cells. Untransfected cells
were considered control cells. Subsequent experiments were
performed at 24 h post-transfection.
MTT cytotoxicity assays
LNCaP cells were seeded into 96-well plates
(3×104 cells in 0.1 ml culture medium per well) and
transfected with miR-628-5p mimic (cat. no. MCH03275), miR-628-5p
inhibitor (cat. no. MIH03275; both Applied Biological Materials
Inc., Richmond, BC, Canada) or control. Following 12-h incubation,
10 µl MTT solution (5 mg/ml) was added and cells were incubated for
a further 2 h at 37°C. Following incubation, cell culture medium
was removed and dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was added to dissolve the purple formazan
crystals. Optical density was measured at 570 nm using a
luminometer microplate reader (Berthold Technologies GmbH, Bad
Wildbad, Germany).
Cell invasion assay
Transwell inserts (8 µm) were used. Upper chambers
were pre-coated with 40 ml Matrigel® (BD Biosciences,
Franklin Lakes, NJ, CA, USA) dissolved in 20% culture medium and
incubated overnight at 37°C. In total, 5×104 cells
collected at 24 h after transfection were suspended in 200 ml
serum-free RPMI-1640 medium and were plated in the upper chamber.
RPMI-1640 medium supplemented with 20% FBS (Gibco; Thermo Fisher
Scientific, Inc.) was plated in the lower chamber. Following
incubation for 36 h at 37°C, cells were stained with 0.1% crystal
violet at room temperature for 20 min. Stained cells were observed
under an optical microscope (magnification, ×40).
ELISA
Prostate-specific antigen (PSA), fibroblast growth
factor (FGF)1 and FGFR2 in plasma were detected by performing ELISA
using human Prostate Specific Antigen ELISA Kit (cat. no. ab188388;
Abcam), Human FGF1 ELISA Kit (cat. no. ab219636; Abcam) and FGFR2
ELISA kit (cat. no. MBS921985; MyBioSource; Thermo Fisher
Scientific, Inc.) respectively.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from LNCaP cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA (≤500 ng) was reverse transcribed into cDNA using
the High Capacity RNA-to-cDNA kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.). To examine the mRNA expression level of
FGFR2, qPCR was performed using the SYBR™ Green Master Mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.). To examine the
expression level of miR-628 expression, qPCR was performed using
the TaqMan probe kit (Roche Applied Science, Penzberg, Germany).
qPCR was performed using an ABI 7900 Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The following primer
pairs were used for qPCR: FGFR2 forward,
5′-CGCTGGTGAGGATAACAACACG-3′ and reverse,
5′-TGGAAGTTCATACTCGGAGACCC-3′. GAPDH forward,
5′-GTCTCCTCTGACTTCAA-3′ and reverse, 5′-ACCACCCTGTTGCTGTA-3′.
miR-628 forward primer: 5′-GTCGTATCCAGTGCAGTTATATCC-3′. miR-628
reverse primer and U6 primers were included in the kit. The
thermocycling conditions for qPCR were: Initial denaturation at
95°C for 10 sec; 40 cycles of 95°C for 10 sec and 60°C for 60 sec.
FGFR2 and miR-628 expression was quantified using the
2−ΔΔCq method (15) and
normalized to the internal reference gene GAPDH or U6 small
non-coding RNA, respectively.
Western blot analysis
Total protein was extracted from LNCaP cells using
the M-PER™ Mammalian Protein Extraction reagent (Pierce; Thermo
Fisher Scientific, Inc.). Protein concentration was determined by
BCA assay. An equal volume of protein was mixed with an equal
volume of 2X SDS loading buffer and boiled for 5 min. Proteins (30
µg per lane) were separated via SDS-PAGE on a 10% gel and
transferred onto polyvinylidene difluoride membranes (Zhongshan
Jinqiao Biology & Technology Co., Ltd., Beijing, China).
Membranes were blocked with 5% skimmed milk at room temperature for
2 h. The membranes were incubated with primary antibodies against
FGFR2 (1:800; cat. no. ab58201; Abcam, Cambridge, MA, USA) and
GAPDH (1:800; cat. no. sc-47724; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) overnight at 2–8°C. Following primary incubation,
membranes were incubated with horseradish peroxidase-labeled goat
anti-mouse immunoglobulin G secondary antibody (1:1,200; cat. no.
ZB2305; Zhongshan Jinqiao Biology & Technology Co., Ltd.) at
room temperature for 3 h. Protein bands were visualized using a
Amersham ECL Prime Western Blotting Detection Reagent (GE
Healthcare Life Sciences, Shanghai, China).
Dual-luciferase reporter assay
TargetScan (http://www.targetscan.org/vert_72/) and miRDB
(http://mirdb.org/) bioinformatics software was used to
identify the 3′ untranslated region (UTR) of FGFR2 as a putative
target of miR-628. The wild-type (wt) and mutant (mut) 3′UTR of
FGFR2, which contains a sequence targeting miR-628, were cloned
into the psi-CHECK vector (Promega Corporation, Madison, WI, USA).
Cells were co-transfected with either miR-628-5p mimic (or
miR-628-5p inhibitor) and psiFGFR2-wt (or psiFGFR2-mut) using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.).
Control miRNA or control inhibitor were used as controls. Following
48-h transfection, cells were harvested and luciferase activities
were measured using the Dual-Luciferase Reporter Assay System
(Promega Corporation) using a luminometer (Berthold Technologies
GmbH). Firefly luciferase activity was normalized to Renilla
luciferase activity.
Statistical analysis
Data were expressed as mean ± standard deviation.
All statistical analyses were performed using Sigmaplot v.11
(Systat Software Inc., Chicago, IL, USA) and SPSS 16.0 software
(SPSS, Inc., Chicago, IL, USA). Welch's t-test was used to analyze
differences between two groups, while one-way analysis of variance
and Tukey's post hoc test were used to analyze comparisons among
multiple groups. Correlation analysis between the serum expression
level of miR-628 and FGF-1 or FGFR2 was analyzed by Pearson's
correlation analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-628 expression is decreased in the
serum of patients with PCa
Serum samples from patients with PCa and healthy
controls were collected and used for gene expression analyses. The
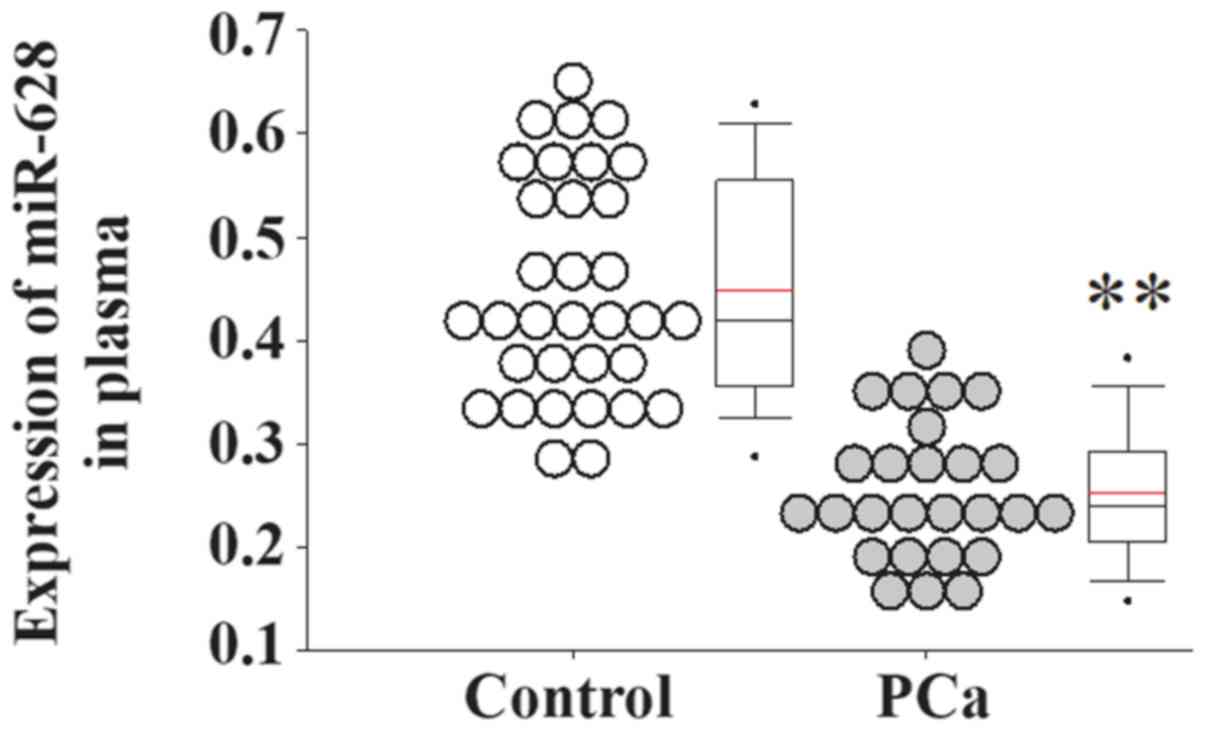
serum expression level of miR-628 was significantly decreased in
patients with PCa compared with healthy controls (Fig. 1). In addition, the serum expression
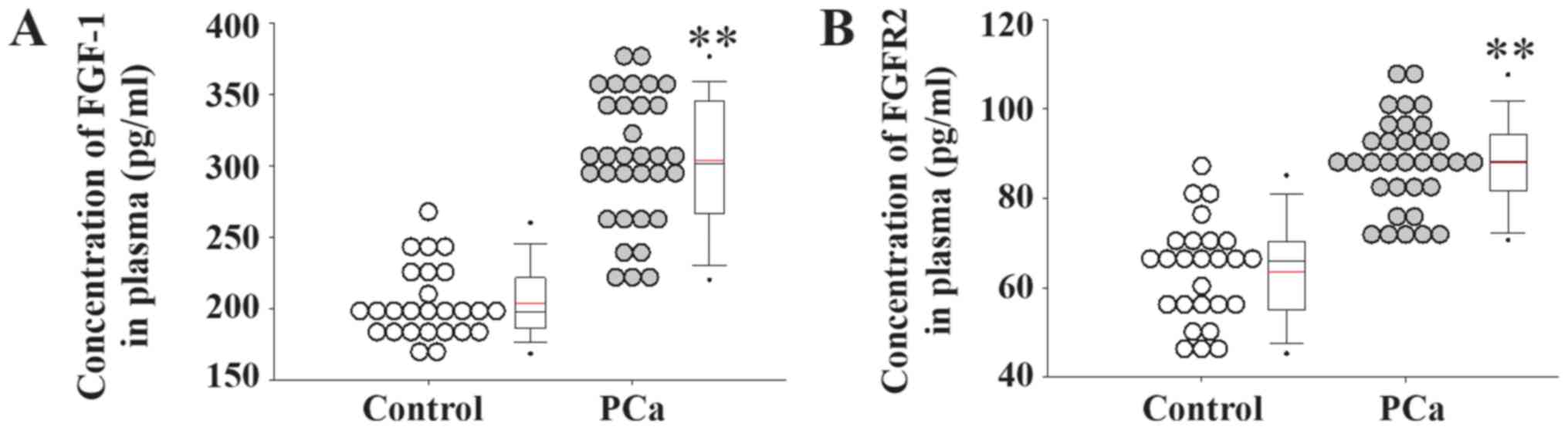
levels of FGF1 and its receptor FGFR2 were significantly increased
in patients with PCa compared with healthy controls (Fig. 2).
miR-628 expression negatively
correlates with FGFR2
Serum levels of PSA were increased in patients with
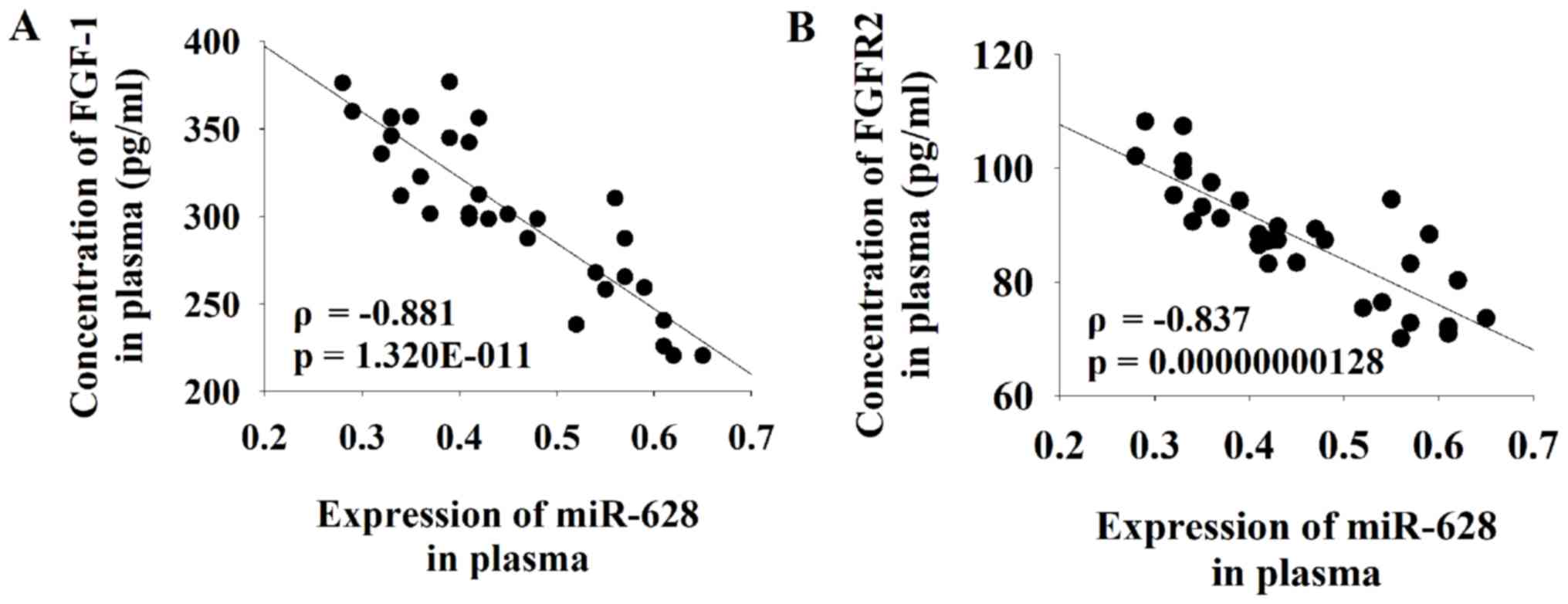
PCa compared with healthy controls (Table I). Pearson correlation coefficient
analysis was performed to determine the association between the
serum expression level of miR-628 and FGF1 or FGFR2. The results of
the current study demonstrated that there was a significant
negative correlation between the serum level of miR-628 and both
FGF1 and FGFR2 in patients with PCa (Fig. 3).
miR-628 regulates FGFR2 signaling
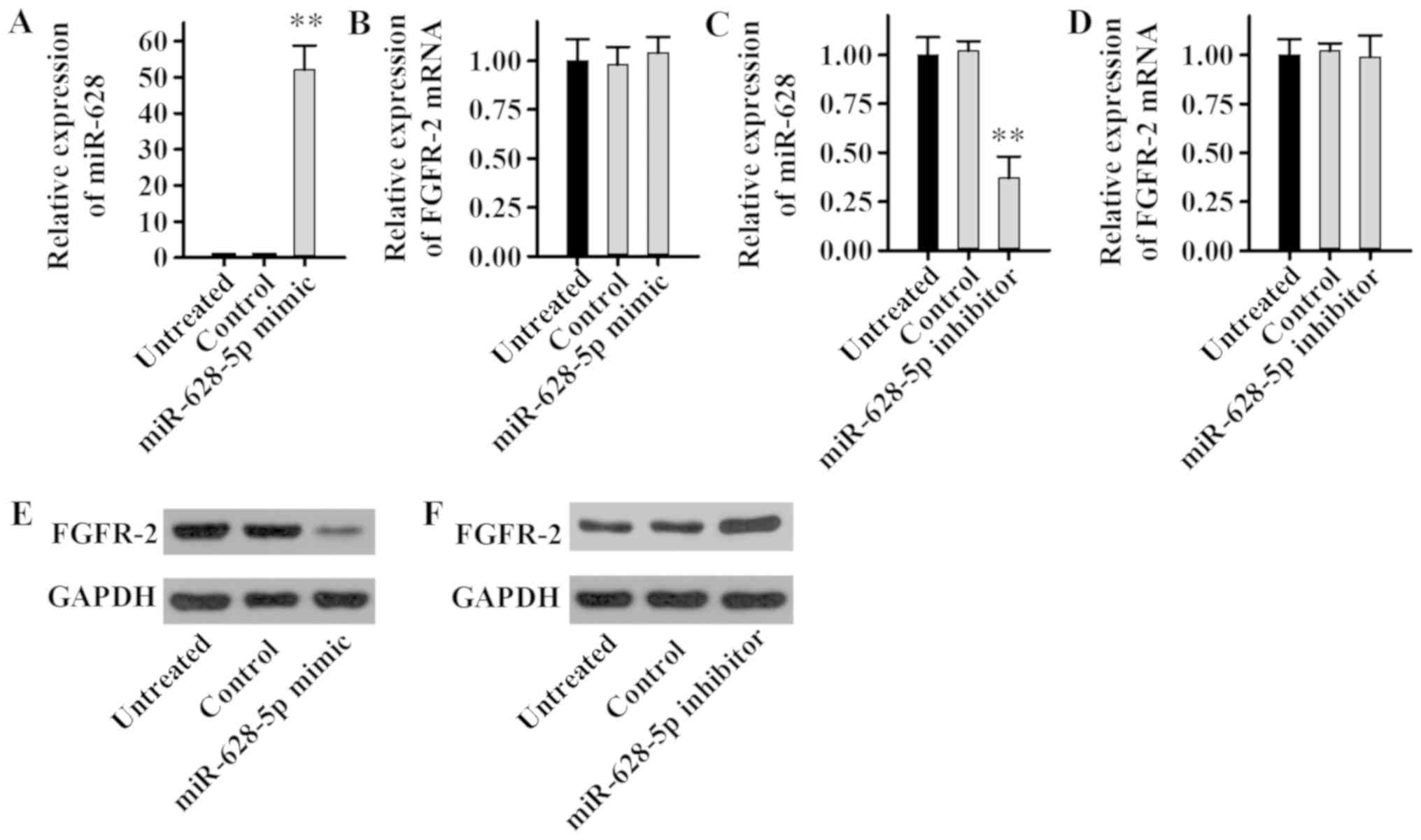
To determine the effect of miR-628 on the expression
level of FGFR2, LNCaP cells were transfected with miR-628-5p mimic,
inhibitor or negative controls, and miR-628 and FGFR2 expression
levels were examined. The current study demonstrated that the
miR-628-5p mimic significantly increased miR-628 expression
(Fig. 4A), whereas the miR-628-5p
mimic had no significant effect on the mRNA expression level of
FGFR2 in LNCaP cells (Fig. 4B). In
addition, the miR-628-5p inhibitor significantly decreased miR-628
expression (Fig. 4C), while the
miR-628-5p inhibitor had no significant effect on the mRNA
expression level of FGFR2 in LNCaP cells (Fig. 4D). Although there was no effect on
the FGFR2 mRNA expression level, the FGFR2 protein expression level
was revealed to be potentially regulated by miR-628. The protein
expression level of FGFR2 decreased markedly following transfection
with miR-628-5p mimic (Fig. 4E),
while the protein expression level of FGFR2 increased following
transfection with miR-628-5p inhibitor (Fig. 4F). Taken together, these results
suggest that miR-628 can inhibit FGFR2 protein expression in PCa
cell lines without affecting transcriptional levels and therefore
miR-628 may regulate FGFR2 expression.
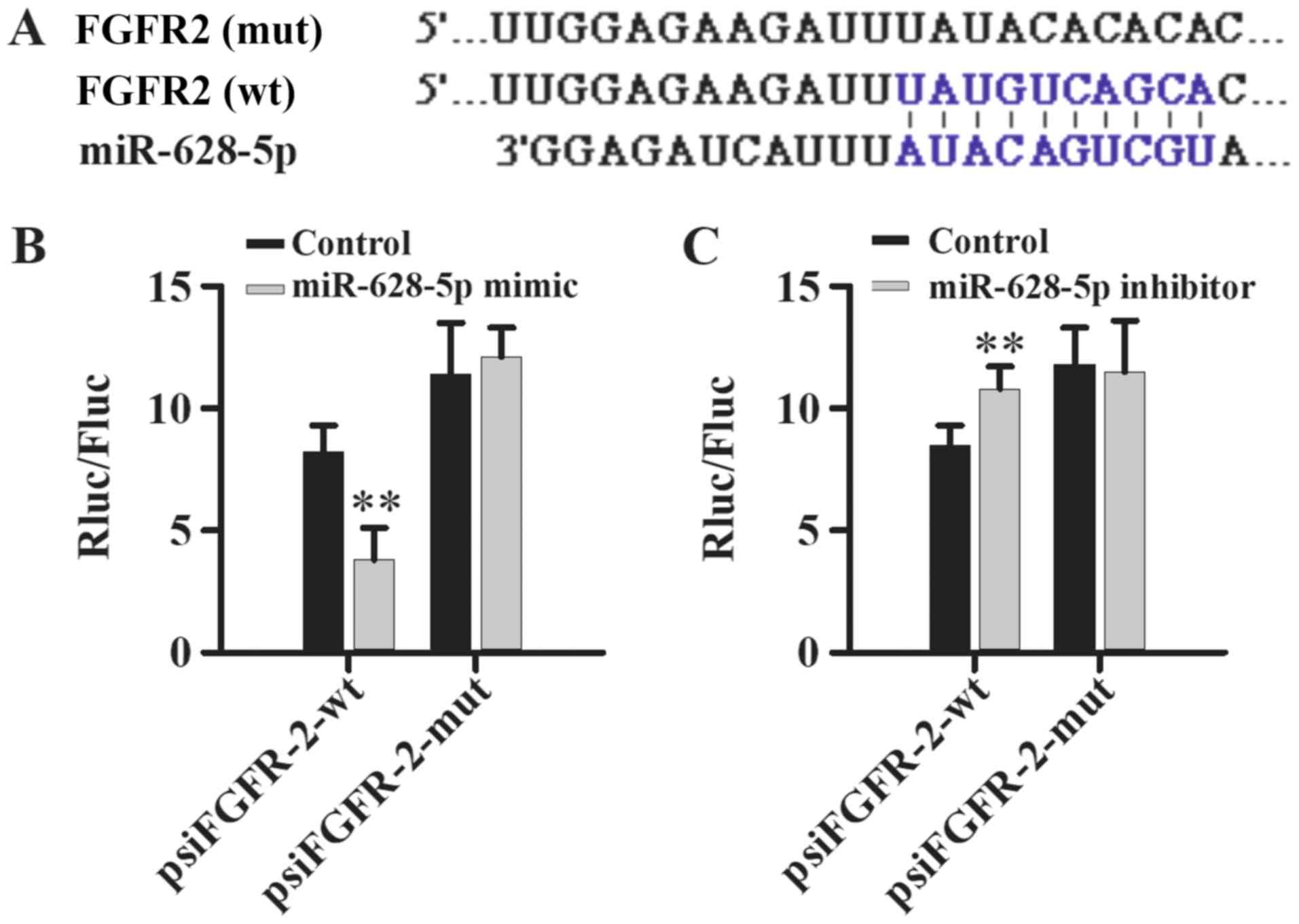
miR-628 directly targets FGFR2
TargetScan and miRDB bioinformatics software was
used to identify the 3′UTR of FGFR2 as a putative target of miR-628
(Fig. 5A) and dual-luciferase
reporter assays were performed to validate the direct interaction
between miR-628 and FGFR2. Luciferase activity was significantly
decreased following co-transfection with miR-628-5p mimic and
psiFGFR2-wt compared with psiFGFR2-mut, which had no significant
effect on luciferase activity (Fig.
5B). In addition, luciferase activity was significantly
increased following co-transfection with miR-628-5p inhibitor and
psiFGFR2-wt compared with psiFGFR2-mut, which had no effect on
luciferase activity (Fig. 5C). Taken
together, these results suggest that miR-628 directly interacts
with FGFR2.
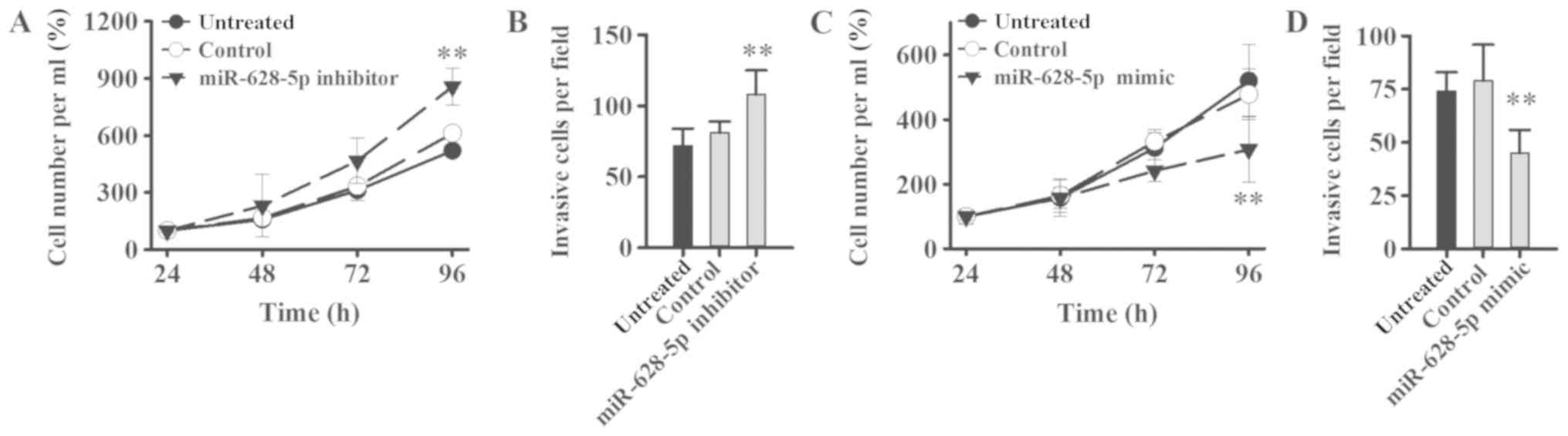
miR-628 regulates tumorigenesis via
FGFR2 signaling pathway
The effect of miR-628 on PCa cell proliferation and
invasion was examined in LNCaP cells following transfection with
miR-628-5p mimic, inhibitor or negative controls. Cell
proliferation and invasion was significantly increased in LNCaP
cells following transfection with miR-628-5p inhibitor compared
with the control group (Fig. 6A and
B). By contrast, cell proliferation and invasion was
significantly decreased in LNCaP cells following transfection with
miR-628-5p mimic compared with the control group (Fig. 6C and D). FGF is known to promote the
proliferation of tumor cells and the formation of tumor blood
vessels (11) and therefore,
downregulating miR-628 expression may increase the expression level
of FGF in PCa and thereby promote tumor initiation, expansion and
metastasis.
Discussion
Several miRNAs, which include miR-21, miR-221 and
miR-141, have been associated with PCa (20). Most of these miRNAs are involved in
cell cycle, migration, metastasis, apoptosis, proliferation,
angiogenesis and epithelial-to-mesenchymal transition (22). A previous study demonstrated that the
serum expression level of miR-628 was significantly downregulated
in African American and Caucasian American patients with PCa
(14). Similarly, the current study
confirmed that the serum expression level of miR-628 was
significantly downregulated in Chinese patients with PCa. The FGFR2
signaling pathway is known to be involved in several types of
cancer. A recent study demonstrated that miR-628 targeted and
downregulated the expression of FGFR2 in ovarian cancer (11). In the current study, serum expression
levels of FGF1, FGFR2 and PSA were examined in patients with PCa
and controls. Serum FGF1 and FGFR2 were negatively correlated with
the serum expression level of miR-628. In addition, dual-luciferase
reporter assays revealed that FGFR2 is a direct target of miR-628.
Furthermore, the current study demonstrated that miR-628 inhibited
PCa cell proliferation and invasion via the FGFR2 signaling
pathway.
The FGFR2 gene at human chromosome 10q26 encodes two
isoforms, FGFR2b and FGFR2c, which function as FGFRs and are
involved in tumorigenesis (23–25).
Missense mutations, copy number variation, splicing and single
nucleotide polymorphisms within intron 2 of the FGFR2 gene are
associated with breast, bladder and gastric cancer whereby aberrant
FGFR2 signaling activation induces proliferation and survival of
tumor cells (23–25). FGFR2 alternative splicing serves a
role in the progression of PCa from an androgen-sensitive to
androgen-insensitive tumor (26–28).
Although FGFR2 inhibitors are approved and used for cancer
treatment (11), the current study
identified miR-628, which may be used as a potential molecular
target for the development of novel therapeutic treatment
strategies for patients with PCa. In addition, targeting miR-628
may also be used to treat ovarian cancer as well as inflammation
regulation (15,19).
Several technologies including human antibody,
peptide mimetic, RNA aptamer, small interfering RNA and synthetic
miRNA are emerging technologies, which can be applied and used as
potential therapeutic treatment strategies in FGFR2-mediated cancer
(24). In particular, these
technologies may be used for the treatment of patients with PCa
with low expression of miR-628 and upregulated FGFR2 signaling.
Curcumin, for example, has a variety of pharmacological effects
including anti-cancer effects and curcumin exerts its therapeutic
effects through regulation of miRNA expression (17). miRNAs are thought to be potential
targets for cancer therapy and as such there have been a number of
studies regarding miRNA regulation via miRNA activators and
inhibitors (16,29–32).
However, developing novel delivery systems for miRNA treatment is
key to its success as a potential therapy (31).
Further experiments are required to validate the
findings of the current study using in vivo mouse models,
such as examining tumorigenesis in a xenograft mouse model. In
vivo mouse models for PCa could be used to study miR-628
expression and regulation of the FGFR2 signaling pathway, as well
as testing newly designed miR-628 activators. However, a critical
step will be to develop a delivery system for the efficient
delivery of miRNAs into in vivo mouse models for PCa. The
current study identified the FGFR2 signaling pathway as a potential
therapeutic target, however, patients may become resistant to
FGFR2-mediated therapy and alternative treatment may be required.
Sequencing technology may be used to develop treatments combining
FGFR2 inhibition and miR-628 upregulation, which may be a
potentially effective approach for the treatment of patients with
PCa.
In conclusion, the present study demonstrated that
miR-628 directly targets FGFR2 and miR-628 can regulate
tumorigenesis via the FGFR2 signaling pathway in PCa cells.
Therefore, FGFR2 and miR-628 may be potential molecular targets for
the development of novel and effective therapeutic treatment
strategies for patients with PCa.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JC and YZ designed experiments. YZ and PH performed
experiments. TZ analyzed data. YZ drafted this manuscript and all
authors approved this manuscript.
Ethics approval and consent to
participate
The current study was approved by the Ethics
Committee of Wuhan Fourth Hospital, Puai Hospital (Wuhan, China).
All patients and healthy volunteers provided written informed
consent prior to their inclusion in the study.
Patient consent for publication
Patients provided consent for publication of data in
this paper.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Han HH, Park JW, Na JC, Chung BH, Kim CS
and Ko WJ: Epidemiology of prostate cancer in South Korea. Prostate
Int. 3:99–102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bashir MN: Epidemiology of prostate
cancer. Asian Pac J Cancer Prev. 16:5137–5141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ye D and Zhu Y: Epidemiology of prostate
cancer in China: An overview and clinical implication. Zhonghua Wai
Ke Za Zhi. 53:249–252. 2015.(In Chinese). PubMed/NCBI
|
|
4
|
Tao ZQ, Shi AM, Wang KX and Zhang WD:
Epidemiology of prostate cancer: Current status. Eur Rev Med
Pharmacol Sci. 19:805–812. 2015.PubMed/NCBI
|
|
5
|
Cooperberg MR and Chan JM: Epidemiology of
prostate cancer. World J Urol. 35:8492017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kimura T and Egawa S: Epidemiology of
prostate cancer in Asian countries. Int J Urol. 25:524–531. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tourinho-Barbosa RR, Pompeo AC and Glina
S: Prostate cancer in Brazil and Latin America: Epidemiology and
screening. Int Braz J Urol. 42:1081–1090. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Beltran H, Antonarakis ES, Morris MJ and
Attard G: Emerging molecular biomarkers in advanced prostate
cancer: Translation to the clinic. Am Soc Clin Oncol Educ Book.
35:131–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen W, Wang GM, Guo JM, Sun LA and Wang
H: NGF/γ-IFN inhibits androgen-independent prostate cancer and
reverses androgen receptor function through downregulation of FGFR2
and decrease in cancer stem cells. Stem Cells Dev. 21:3372–3380.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Adeola HA, Smith M, Kaestner L, Blackburn
JM and Zerbini LF: Novel potential serological prostate cancer
biomarkers using CT100+ cancer antigen microarray platform in a
multi-cultural South African cohort. Oncotarget. 7:13945–13964.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li M, Qian Z, Ma X, Lin X, You Y, Li Y,
Chen T and Jiang H: MiR-628-5p decreases the tumorigenicity of
epithelial ovarian cancer cells by targeting at FGFR2. Biochem
Biophys Res Commun. 495:2085–2091. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schwertfeger KL: Fibroblast growth factors
in development and cancer: Insights from the mammary and prostate
glands. Curr Drug Targets. 10:632–644. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pernar CH, Ebot EM, Wilson KM and Mucci L:
The epidemiology of prostate cancer. Cold Spring Harb Perspect Med.
8(pii): a0303612018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Srivastava A, Goldberger H, Dimtchev A,
Marian C, Soldin O, Li X, Collins SP, Suy S and Kumar D:
Circulatory miR-628-5p is downregulated in prostate cancer
patients. Tumour Biol. 35:4867–4873. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Favreau AJ and Sathyanarayana P:
miR-590-5p, miR-219-5p: miR-15b and miR-628-5p are commonly
regulated by IL-3, GM-CSF and G-CSF in acute myeloid leukemia. Leuk
Res. 36:334–341. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bryzgunova OE, Konoshenko MY and Laktionov
PP: MicroRNA-guided gene expression in prostate cancer: Literature
and database overview. J Gene Med. 20:e30162018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mirzaei H, Masoudifar A, Sahebkar A, Zare
N, Sadri Nahand J, Rashidi B, Mehrabian E, Mohammadi M, Mirzaei HR
and Jaafari MR: MicroRNA: A novel target of curcumin in cancer
therapy. J Cell Physiol. 233:3004–3015. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jun H, Ying H, Daiwen C, Bing Y, Xiangbing
M, Ping Z, Jie Y, Zhiqing H and Junqiu L: miR-628, a microRNA that
is induced by Toll-like receptor stimulation, regulates porcine
innate immune responses. Sci Rep. 5:122262015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu Y, Li X, Liu L, Chai J, Haijun Z, Chu
W, Yin H, Ma L, Duan H and Xiao M: miR-628Promotes burn-induced
skeletal muscle atrophy via targeting IRS1. Int J Biol Sci.
12:1213–1224. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen H, Ji X, She F, Gao Y and Tang P:
miR-628-3p regulates osteoblast differentiation by targeting RUNX2:
Possible role in atrophic non-union. Int J Mol Med. 39:279–286.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rosini P, Bonaccorsi L, Baldi E,
Chiasserini C, Forti G, De Chiara G, Lucibello M, Mongiat M, Iozzo
RV, Garaci E, et al: Androgen receptor expression induces FGF2,
FGF-binding protein production and FGF2 release in prostate
carcinoma cells: Role of FGF2 in growth, survival and androgen
receptor down-modulation. Prostate. 53:310–321. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Giri D, Ropiquet F and Ittmann M:
Alterations in expression of basic fibroblast growth factor (FGF) 2
and its receptor FGFR-1 in human prostate cancer. Clin Cancer Res.
5:1063–1071. 1999.PubMed/NCBI
|
|
24
|
Ropiquet F, Huguenin S, Villette JM,
Ronflé V, Le Brun G, Maitland NJ, Cussenot O, Fiet J and Berthon P:
FGF7/KGF triggers cell transformation and invasion on immortalised
human prostatic epithelial PNT1A cells. Int J Cancer. 82:237–243.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Katoh Y and Katoh M: FGFR2-related
pathogenesis and FGFR2-targeted therapeutics (Review). Int J Mol
Med. 23:307–311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Carstens RP, Eaton JV, Krigman HR, Walther
PJ and Garcia-Blanco MA: Alternative splicing of fibroblast growth
factor receptor 2 (FGF-R2) in human prostate cancer. Oncogene.
15:3059–3065. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Muh SJ, Hovhannisyan RH and Carstens RP: A
Non-sequence-specific double-stranded RNA structural element
regulates splicing of two mutually exclusive exons of fibroblast
growth factor receptor 2 (FGFR2). J Biol Chem. 277:50143–50154.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Naimi B, Latil A, Fournier G, Mangin P,
Cussenot O and Berthon P: Down-regulation of (IIIb) and (IIIc)
isoforms of fibroblast growth factor receptor 2 (FGFR2) is
associated with malignant progression in human prostate. Prostate.
52:245–252. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Catto JW, Alcaraz A, Bjartell AS, De Vere
White R, Evans CP, Fussel S, Hamdy FC, Kallioniemi O, Mengual L,
Schlomm T and Visakorpi T: MicroRNA in prostate, bladder, and
kidney cancer: A systematic review. Eur Urol. 59:671–681. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Curtin CM, Castaño IM and O'Brien FJ:
Scaffold-based microRNA therapies in regenerative medicine and
cancer. Adv Healthc Mater. 7:2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hosseinahli N, Aghapour M, Duijf PHG and
Baradaran B: Treating cancer with microRNA replacement therapy: A
literature review. J Cell Physiol. 233:5574–5588. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Weidle UH, Birzele F, Kollmorgen G and
Nopora A: Potential microRNA-related targets for therapeutic
intervention with ovarian cancer metastasis. Cancer Genomics
Proteomics. 15:1–15. 2018.PubMed/NCBI
|