Introduction
Stress is a complex condition with negative
emotional and biological effects. Stress can be detrimental to
human health regardless of whether it occurs during the prenatal
period, infancy, childhood, adolescence, adulthood or in old age.
Stress has an impact on the brain structures involved in cognition
and mental health (1). Early-life
stress causes critical developmental complications and impacts
brain functions in both animals and humans (2–4). Early
life stress also increases the anxiety level of fish (4) and alters social behaviour in rodents
(2). Similarly in humans, exposure
to stress in early life increases the risk of developing mental
health problems such as hyperactivity (3) and sleep problems (5). While stress is percieved to have
negative consequences, this may not always be true in the case of
acute stress (6). In contrast to
chronic stress, acute stress results from a single exposure to a
stressor on a scale of min to hours (6). The effects of chronic stress are not as
clear as it paradoxically can have both positive and negative
effects in the case of cognition (7).
We know that sleep is an important physiological and
behavioral state of human homeostasis. The negative impact of
disturbed sleep on cognitive functions, mood and overall health are
very much evident (8). Melatonin has
a significant role in sleep initiation and maintenance in humans.
Its administration promotes sleep in fish (9), birds (10) diurnal primates (11,12) and
in humans (13). The effects of
reduced melatonin levels on sleep are still under investigation.
Some studies have suggested that circulating melatonin levels are
significantly lower in aged individuals as compared to young
healthy adults, which affects their sleep (14–16).
Similarly, Zhdanova et al (17) has shown that an age-dependent decline
in nighttime brain melatonin levels reduces the sleep time in
zebrafish. Haimov et al (16), has also proven a significant
correlation between a low level of melatonin production and
insomnia. Based on other studies, stress also negatively affects
melatonin synthesis (18,19), which further modulates a variety of
physiological processes such as sleep regulation.
AANAT is the first enzyme in the conversion of
serotonin to melatonin and its activity plays an important role in
melatonin production. Roseboom et al (20), has observed that after the onset of
darkness, aanat mRNA levels increases, followed shortly by
increases in AA-NAT protein levels and hence enzyme activity.
Literature suggests that there are two aanats which are expressed
(aanat1 in the retina and aanat2 in the pineal organ)
in teleost fish. Retina aanat1 mRNA peaks in the late
afternoon and pineal annat2 mRNA peaks six h later (21).
Glucocorticoids have been identified as the main
corticosteroids that protect an organism against stress (22), but excessive exogenous
glucocorticoids can induce low birth weight in infants,
hypertension in adulthood as well as increased anxiety-related
behaviour in rats (23,24) and in zebrafish (4). In literature, it is also documented
that glucocorticoids have an inhibitory effect on melatonin
synthesis (25,26). Literature also suggests that
Nfkb is potential biomarker for oxidative stress (27) and is found in almost all animal cell
types. It is also involved in cellular responses to stimuli such as
stress, cytokines and free radicals (27). This is significant because melatonin
also prevents oxidative stress (28)
and inhibits nfkb expression (29).
Piper betle L
(PB) is one of the most popular plants in the
Asiatic region and ranks second to tea and coffee in terms of daily
consumption (30). PB leaf extract
and its purified compounds are well-known ethnomedicines and are
associated with a number of potential therapeutic efficacies. It
has been reported to have antimicrobial, antibacterial,
antioxidant, anticancer, antidiabetic and anti-inflammatory
activities (31). In recent studies,
PB has been shown to exhibit analgesic and antidepressant
activities as well (32,33).
Although most behavioural research in the past was
done with rodent models, zebrafish are an upcoming animal model due
to their prolific breeding, which results in a large number of
offspring for experimental purposes in a relatively short duration
(34). Other advantages of zebrafish
as compared to rodents in terms of an animal model are their longer
lifespan and robust phenotypes, as they display apparent and easily
quantifiable behavioural endpoints (35). The variety of behavioural and
molecular tools available for use with zebrafish makes them
outstanding models for helping to investigate neuromolecular
mechanisms. Zebrafish can provide a comparatively quick indication
of possible functional efficacy. In addition, this diurnal
vertebrate has several fundamental similarities to humans (36). All these characteristics make
zebrafish a better animal model for behavioural research. Due to
their cost effectiveness, reliability and scalability, this study
opted for zebrafish as an animal model over rodents. In the context
of this experiment, zebrafish are also emerging as a promising
model organism for experimental studies of sleep and its related
disorders as its pineal gland and retina contains circadian
oscillators that drive the rhythms for synthesizing melatonin
(37) and regulating sleep in a
similar fashion as in humans (38).
Thus, the aim of the present study was to evaluate
the effect of early life stress induced via the glucocorticoid
dexamethasone (DEX), on melatonin synthesis and adult zebrafish
behaviour. This study also aims to assess the potential role of PB
for the treatment of stress induced sleep disruption.
Materials and methods
Preparation of the plant extract
PB dried leaves were obtained from the herbal
farm, Sungai Buloh, Malaysia. Following the extraction protocol
used by Ganguly et al (39),
the dried leaves were first grounded to powder form in a hand
crusher. The powdered leaves (300 gm) were extracted twice with
absolute ethanol (900 ml each time) and filtered through Whatman
no. 1 filter paper. The alcoholic extract was subsequently dried in
a rotary evaporator to obtain a semi solid extract with an
approximate yield of 5%. Then extract was stored in −20°C until
needed. The PB extract was dissolved in 1% of Dimethyl sulfoxide
(DMSO) before being used. Liquid chromatography-tandem mass
spectrometry (LC-MS/MS) was employed to identify the chemical
constituents in the alcoholic extract of PB. PB was found to have a
qualitative composition of eugenol, allylpyrocatechol,
caryophyllene and camphor.
Luciferase reporter gene assay
A luciferase reporter gene assay was performed to
determine the affinity of PB for the melatonin receptors MT1 and
MT2. This method measures G protein signalling through changes in
the levels of the secondary messenger cAMP. In case of the
melatonin receptor assay, stimulation of Gi-coupled melatonin
receptors decreases the expression of intracellular cAMP, leading
to a decrease in the transcription level of the reporter gene
luciferase and hence the degree of bioluminescence produced. For
this experiment, the method described by Chen et al
(40) was used, with some minor
modifications for Luciferase Reporter Gene Assay. Briefly, human
embryonic kidney (HEK) (41) cells
were grown in Dulbecco's modified Eagle medium (DMEM) supplemented
with 10% fetal bovine serum (FBS) at 37°C and in an atmosphere of
5% CO2. On the second day after checking the confluency
of the cells, the transfection was done using a suspension of
plasmid (MT1 or MT2), Luciferase reporter (CRE),
OPTI-MEM (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) and Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc.). After 24 h, the media and cell morphology was checked and
then treated with either 0.25 mM of the positive controls (MT1
receptor assay positive control: 6-chloromelatonin; MT2 receptor
assay positive control: 2-iodomelatonin) or different PB extract
doses (10, 30, 50 µg/ml). Forskolin (1:1,000) was also added as a
stimulator of adenylylcyclase, to raise the cAMP level so that the
inhibition of cAMP is easier to detect. After 6 h of treatment, the
dual Luciferase assay was performed as per the manufacturer's
(Promega Corporation, Madison, WI, USA) instructions. For the data
analysis, the luciferase activity of each sample was normalized to
the renilla luciferase activity and the relative luciferase
activity was calculated with the help of the formula:
Relative
Luciferaseactivity=FLA/RLAcompoundFLA/RLAnegativecontrol
FLA=Firefly luciferase
activity,RLA=Renilla luciferase activity
Animal breeding
Adult zebrafish (Danio rerio; 3–4 months-old)
of heterogeneous wild-type stock (standard short-fin phenotype)
were obtained from a local commercial supplier (Akuarium Batu
Karang Laut Sdn Bhd, Selangor, Malaysia). All fish were housed in
the animal facility department of Monash University, Malaysia under
standard conditions. They were maintained under 14 h Light (200
lux): 10 h Dark (5 lux) regime [light onset: 8 am (ZT; Zeitgeber
Time=0); light offset: 10 pm (ZT=14)] and fed twice daily to ensure
a constant source of nourishment. They were kept under the above
conditions for two weeks to acclimatize the zebrafish before
breeding them. In the present study, 10 adult zebrafish (5 females
and 5 males) were used as breeders to procure eggs. We have used
the standard breeding method as described by Khor et al
(4).
Dexamethasone (DEX) exposure for early
life stress
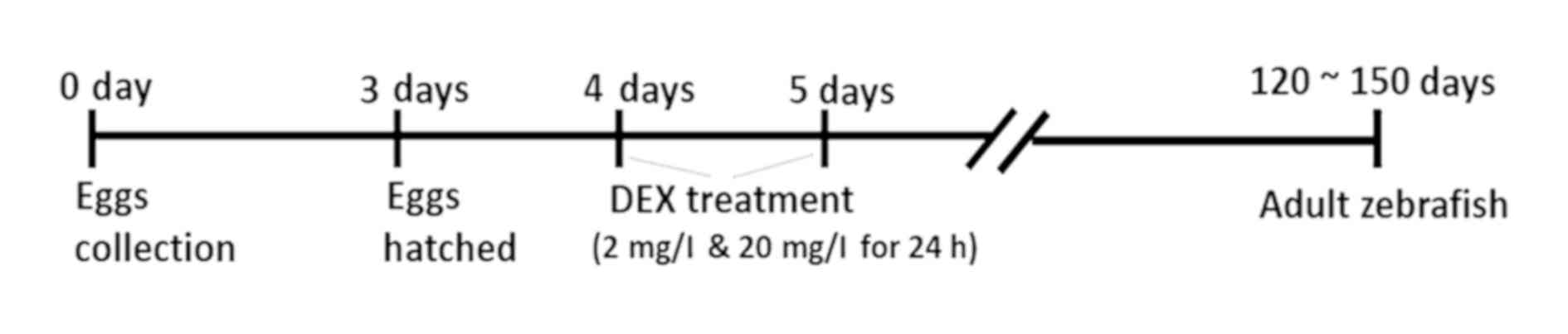
To create an early life stress model (Fig. 1), zebrafish larvae were treated with
dexamethasone 21-phosphate salt (DEX; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) (2 and 20 mg/l) and with tap water (control
group) at 4 dpf (days post fertilization) for 24 h. All the larvae
were randomly divided into groups. The doses used for DEX treatment
(2, 20 mg/l) were chosen as per the doses established by Khor et
al (4) in their model of
zebrafish anxiety like behaviour. Although the paper also tested
the effect of 200 mg/l of DEX, this dose resulted in very apprent
changes in the swimming pattern of the zebrafish. After the
treatment, the larval zebrafish were maintained until the adult
stage (120~150 dpf; weight range of 0.7–0.9 grams) under the
standard conditions previously mentioned (14:10 LD) till subjected
to experimentation. Only adult male zebrafish were used in the
study. All experimental procedures implemented in this study follow
the guidelines on the care and the use of animals, with ethical
approval by the Monash Animal Research Platform (MARP) Animal
Ethics Committee, Monash University, Australia (MARP/2014/133).
Grouping and treatment
A total of 200 zebrafish were used in the present
study. All zebrafish were randomly divided into groups using a
physical randomisation method. In the first part of the study, all
adult zebrafish which were earlier exposed to DEX treatment;
divided into three groups (n=10 per group) consisting of treatments
with DEX (2, 20 mg/l) and water (control) to explore the effect of
early life stress on behaviour (locomotor activity and % of time
spent in the upper zone of the tank), the level of melatonin and
nfkb expression in adult zebrafish. Secondly, the fish were divided
into eight groups (n=8–10 per group) consisting of early life
exposure to DEX (2 and 20 mg/l) and treatment with either PB 10
mg/Kg, PB 30 mg/Kg, melatonin 10 mg/kg or vehicle (DMSO), to
determine the effect of PB on behavior, melatonin receptor and
synthesis as well as stress related gene expression in adult
zebrafish. Intraperitoneal injections were used for the PB,
melatonin and vehicle (DMSO) treatment, using a standard method
according to Kundap et al (42). The melatonin dose was chosen based on
the work of Cuesta et al (43) whom determined that this dose produced
the highest positive effects when injected into seabream (teleost
fish).
The experimenter was unaware of the identity of the
control and treatment groups while processing the samples for
analysis.
Behavior tests
All the behaviour tests were carried out with adult
male zebrafish in the early h (9:00am-1:00pm) of the light phase,
The zebrafish had their behaviour monitored in an experimental tank
with the dimensions of 36×26×22 cm, filled with 10L of water.
Lighting conditions were fixed (~400 lux) and the water temperature
was at 28°C±1°C. A video tracking system (Sony Handycam PJ340) was
used to record their behaviour. All data was analyzed by using
SMART software (Panlab, Barcelona, Spain). For analysis, the
experimental tank was divided into two equal halves (upper and
lower zones). The sleep like behaviour parameters analysed in this
study as adapted from Zhdanova et al (17), were locomotor activity-total distance
travelled (cm) in the tank; time spent in upper zone of the tank
(%) and quiescent state-inactive period (sec) in the given period
of time (10 min). As per the criterion of Zhdanova et al
(17), inactive period was defined
as the number of sec in which the swimming speed of the fish is
below 0.1 cm/sec. of Fish were injected intraperitoneally with
their respective treatment doses of PB, melatonin or DMSO (as
mentioned above) and after 30 min, they were subjected to behaviour
monitoring.
Melatonin estimation
LCMS-MS was performed for the estimation of
melatonin in adult zebrafish brains which were earlier exposed to
DEX treatment. All zebrafish were divided into three groups
consisting of treatments with DEX (2 mg/l, n=6; 20 mg/l, n=6) and
control (water, n=6) as mentioned above. All brain samples were
harvested at ZT 16 (after the 2 h of light offset). Melatonin is
light sensitive, so as a precaution, all sampling was completed
under very dim red light. Samples were harvested and homogenized in
400 µl of ice cold methanol with 0.1% formic acid. The homogenate
was vortex mixed and centrifuged at 18,000 X g for 10 min at 4°C.
The supernatant was used for further analysis by using the standard
method of LCMS-MS (42).
Gene expression study
A new set of DEX-exposed fish groups were divided in
a similar manner to the behavior study. They were injected
intraperitoneally with their respective doses at ZT12 (2 h before
lights offset) and all brain samples were harvested at ZT16 (after
the 2 h of light offset). Because melatonin is light sensitive, all
sampling was completed under the very dim red light as a
precaution. All brain samples were collected in 200 µl of ice cold
trizole (Invitrogen; Thermo Fisher Scientific, Inc.) and
immediately stored in −80°C until needed.
RNA isolation and cDNA synthesis
Total mRNA was isolated by following the
manufacturers' protocol. In brief, the brain tissue was properly
homogenized in trizole and mixed with 40 µl of chloroform and
centrifuged at 13,500 rpm (revolutions per minute) for 15 min at
4°C. The upper aqueous supernatant was transferred into new tubes
and 100 µl of isopropanol was added, mixed and incubated for 10 min
at room temperature and centrifuged for 10 min at 13,500 rpm at
4°C. Later, the supernatant was discarded and the pellets were
subjected for rinsing with 200 µl of 75% ethanol. Then, the pellets
were left for air drying for between 5 to 8 min. Finally, 15 µl of
nuclease free water was added to each tube to dissolve the mRNA
pellet. The concentration and purity of the isolated mRNA was
measured using a NanoDrop Spectrophotometer. The mRNA samples were
converted into cDNA using the Omniscript Reverse-transcription kit
(Qiagen GmbH, Hilden, Germany) according to the manufacturer's
protocol, with a final volume of 20 µl.
Reverse transcription-quantitative-PCR
(RT-qPCR)
Gene expression levels for melatonin receptor 1A
(MT1), melatonin receptor 1B (MT2), arylalkylamine
N-acetyltransferase 1 (aanat1), arylalkylamine
N-acetyltransferase 1 (aanat2) and nuclear factor
Kappa-light-chain-enhancer of activated B cells (nfkb) were
quantified by real-time quantitative RT-qPCR (Step one; Applied
Biosystems) using QuantiTect SYRB Green dye (Qiagen, Valencia, CA,
USA). All the primer sets were provided by Qiagen
(MT1:QT02113986; MT2:PPZ00432A;
aanat1:QT02041431; aanat2:QT02183783;
nfkb:QT02149980 and eef1a1b:QT02042684). The PCR
mixture contained 1X SYBR-Green PCR master mix (Qiagen), 0.7 µM
each forward and reverse primers and 1 µl of sample cDNA. The
samples were incubated at 95°C for 2 min prior to thermal cycling
(40 cycles of 95°C for 5 sec and 60°C for 15 sec). The relative
expression values of the above genes was obtained by normalizing
threshold cycle (Ct) values of genes of interest against Ct value
of eef1a1b (housekeeping gene) [2^(Ct eef1a1b-Ct Gene
of interest)] (42).
Statistical analysis
All obtained data was analyzed by one-way analysis
of variance (one-way ANOVA) followed by Dunnett post-hoc test with
the level of significance set as *P<0.05, **P<0.01,
***P<0.001 for the DEX-2 groups and #P<0.05,
##P<0.01, ###P<0.001 for the DEX-20
groups, using Graph Pad Prism v.5.0. The data is presented as mean
± standard error of the mean.
Results
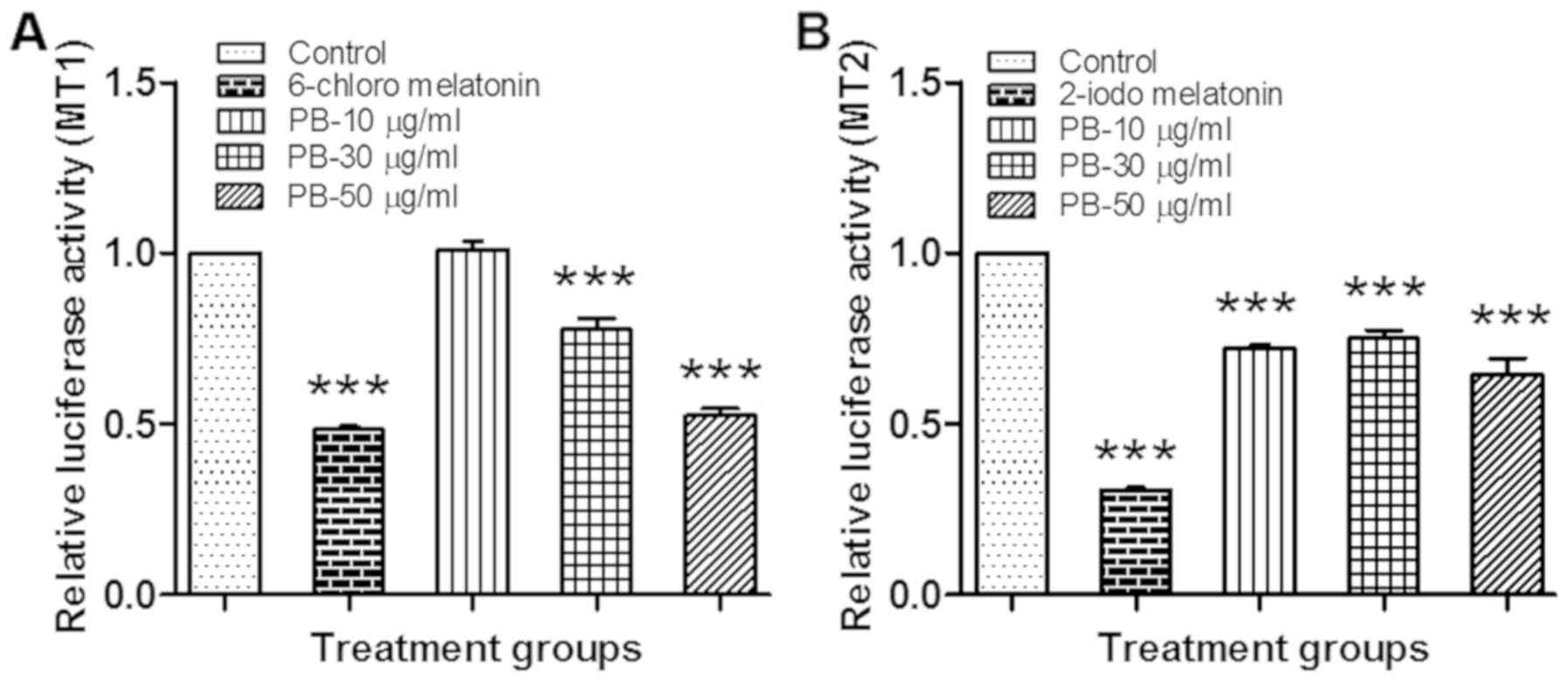
Luciferase reporter gene assay
PB significantly decreased (P<0.001) the relative
luminescence activity as compared to the vehicle control, which
indicates that Piper betel extract exhibited a strong agonist
activity towards Melatonin receptor 1A (Fig. 2A) and 1B (Fig. 2B).
Effect of early life DEX exposure in
adult zebrafish on the behaviour and the level of melatonin and
nfkb gene expression
The groups of fish which were exposed to DEX (2 and
20 mg/l) in early life, show significantly increased (P<0.01 and
P<0.05) total distance travelled in a given period of time (10
min) in comparison to control group of fish (Figs. 3 and 4A). Furthermore, the percentage of time
spent in the upper zone was significantly decreased in DEX 2 mg/l
(P<0.001) and 20 mg/l (P<0.001) treated fish in comparison to
control group (Figs. 3 and 4B).
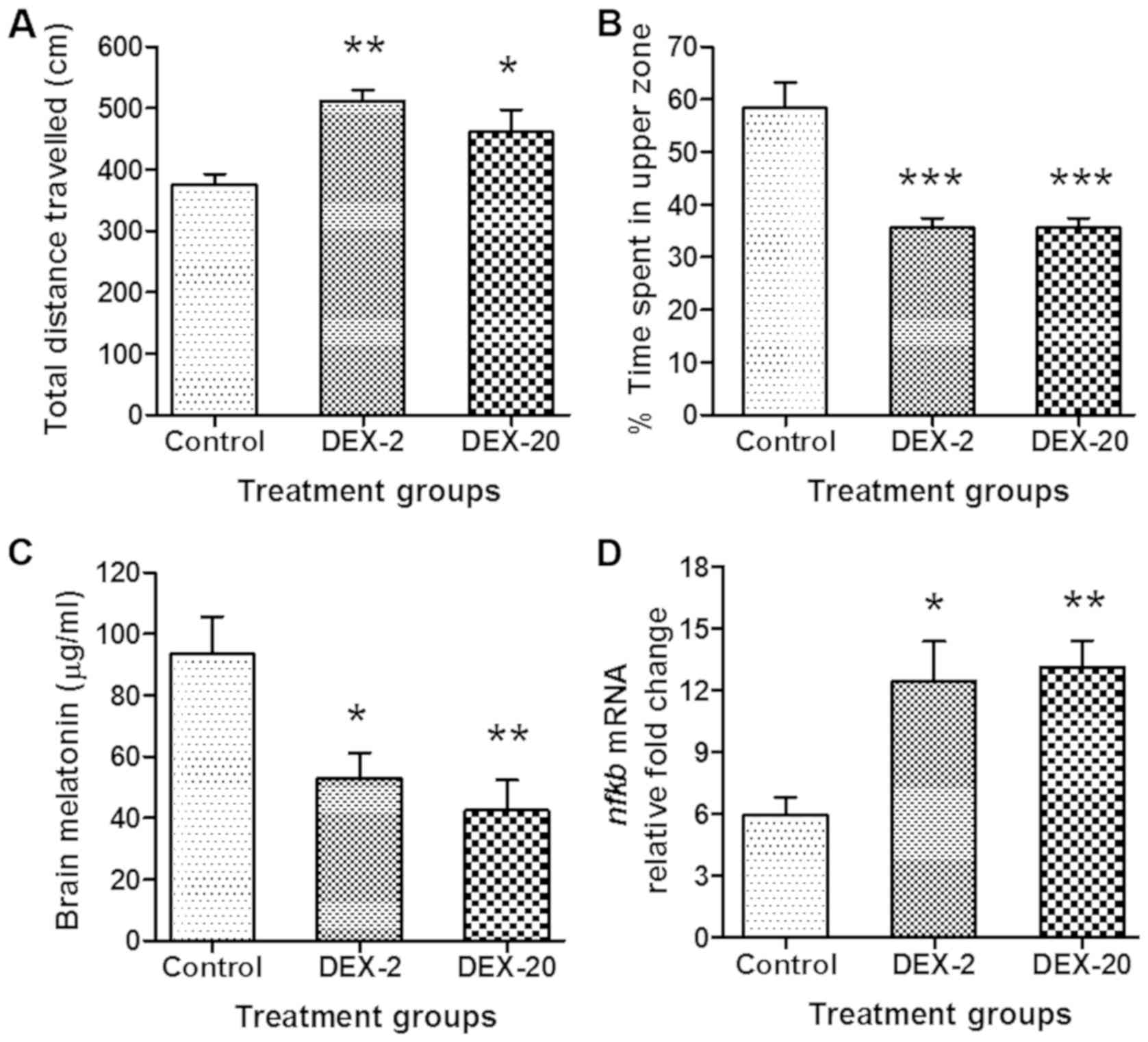 | Figure 4.Effect of early life DEX treatment on
the locomotion parameters, brain melatonin levels and nfkb
mRNA expression level of adult zebrafish. (A) Effect of early life
DEX treatment (2 and 20 mg/l) in adult zebrafish; total distance
travelled, (B) % time spent in the upper zone of tank, (C) level of
brain melatonin and (D) nfkb mRNA expression level. The data were
analyzed by one way ANOVA followed by dunnett post-hoc test, and
was represented as the mean ± SEM, n=8. *P<0.05; **P<0.01;
***P<0.001 vs. the Control Group. DEX, dexamthasone;
nfkb, nuclear factor kappa-light-chain-enhancer of activated
B cells; mRNA, messenger RNA; ANOVA, analaysis of variance; SEM,
standard error of the mean. |
In the case of melatonin estimation, fish which were
exposed to DEX (2 and 20 mg/l) in early life show significantly
reduced (P<0.05 and P<0.01) levels of melatonin in the adult
zebrafish brain in comparison to control group (Fig. 4C).
The results also showed a significant elevation
(P<0.05) and (P<0.01) in the expression level of nfkb
in the DEX treated groups (2 and 20 mg/l) as compared to control
group (Fig. 4D).
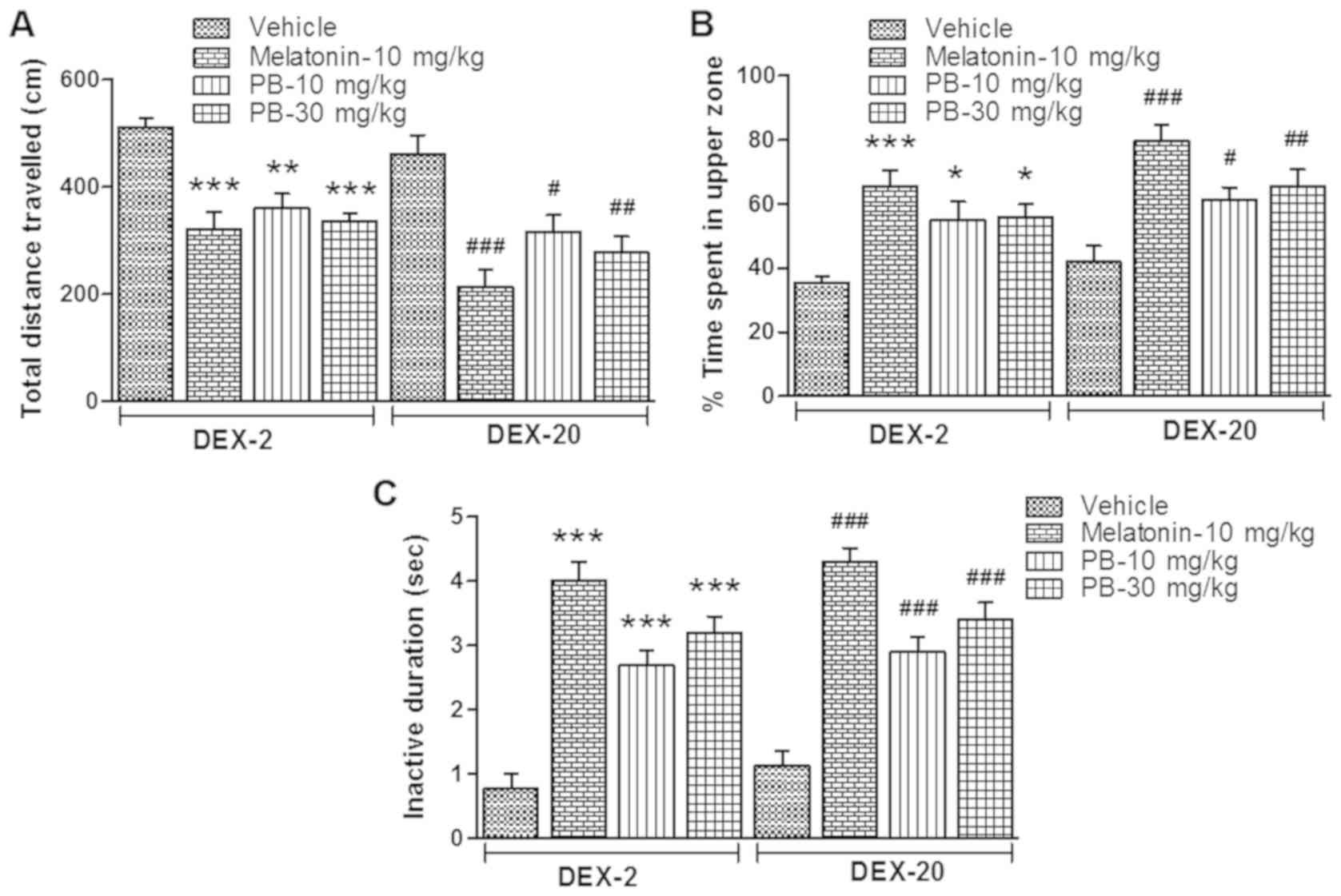
Effect of PB on adult zebrafish
behaviour which were earlier exposed to DEX treatment
After the treatment of melatonin (positive control;
10 mg/kg) and PB (10 and 30 mg/kg) on the groups of fish which were
earlier exposed to DEX (2 and 20 mg/l), they showed significantly
reduced [(DEX-2; P<0.001, P<0.01 and P<0.001) (DEX-20;
P<0.001, P<0.05 and P<0.01)] locomotor activity in given
period of time (10 min) (Figs. 5 and
6A) and their time spent in the
upper zone of the tank significantly increased [(DEX-2; P<0.001,
P<0.05 and P<0.05) (DEX-20; P<0.001, P<0.05 and
P<0.01)] (Figs. 5 and 6B). Furthermore their quiescent state
(inactive duration) also significantly increased [(DEX-2;
P<0.001, P<0.001 and P<0.001) (DEX-20; P<0.001,
P<0.001 and P<0.001)] in comparison to their respective
vehicle treated group (Fig. 6C).
Effect of PB on gene expression of
adult zebrafish which were earlier exposed to DEX treatment
After the treatment of melatonin (positive control;
10 mg/kg) and PB (10 and 30 mg/Kg) on the groups of fish which were
earlier exposed to DEX (2 and 20 mg/l), they show significantly
reduced mRNA expression level of aanat2 [(DEX-2; P<0.01
and P<0.05) (DEX-20; P<0.001, P<0.01 and P<0.001)]
(Fig. 7B) and nfkb [(DEX-2;
P<0.001, P<0.001 and P<0.01) (DEX-20; P<0.001,
P<0.001 and P<0.001)] (Fig.
7E). In the case of MT2, the mRNA expression level
significantly increased [(DEX-2; P<0.001 and P<0.05) (DEX-20;
P<0.01 and P<0.01)] in comparison to their respective vehicle
treated group (Fig. 7D), but there
was no significant change in aanat1 and MT1 gene
expression.
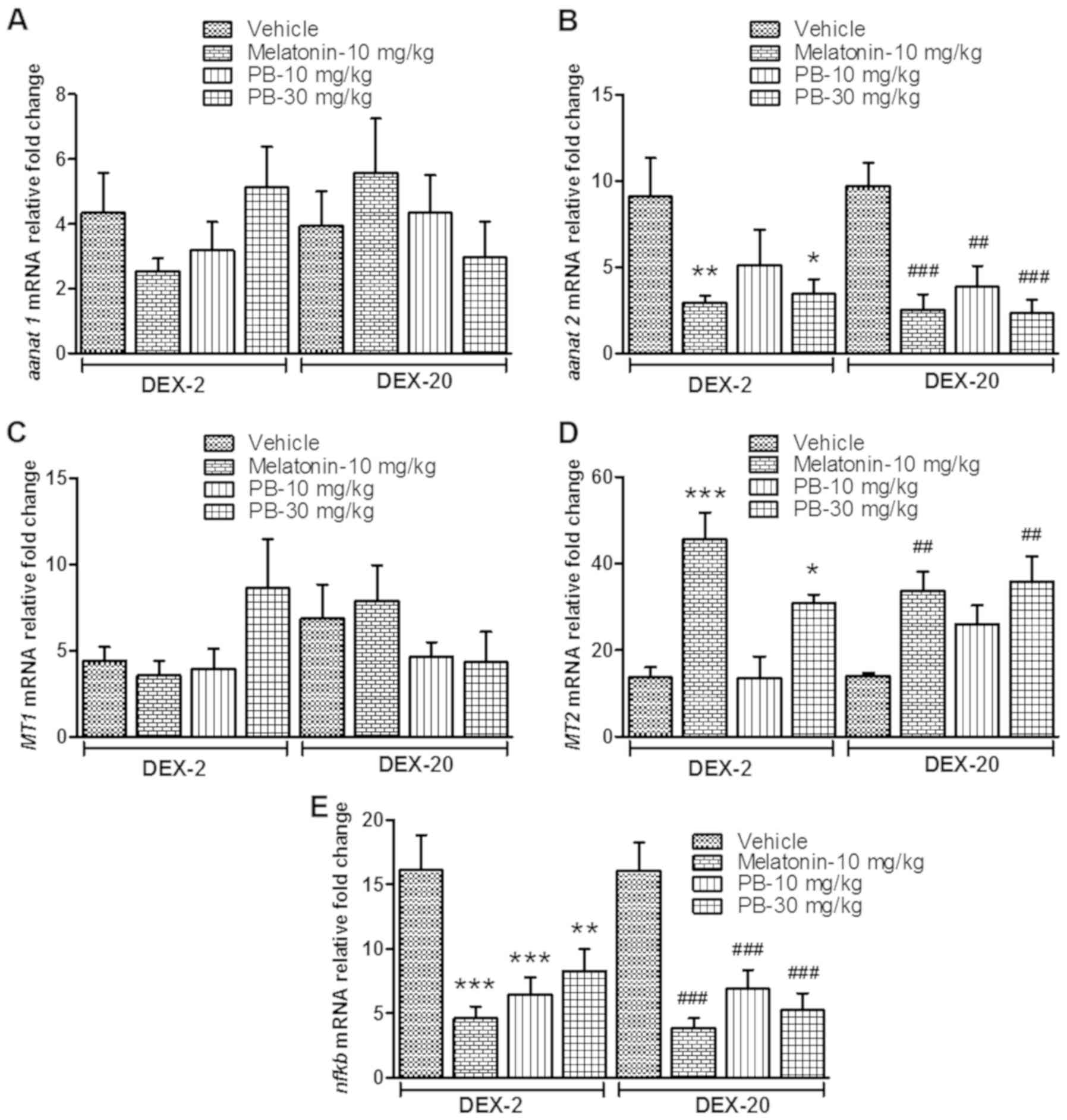 | Figure 7.Effect of PB on sleep-related genes
in early life DEX treated adult zebrafish. Effect of PB (10 and 30
mg/kg) on early life DEX treated (2 and 20 mg/ml) adult zebrafish
on sleep related gene; (A) aanat1 mRNA expression level, (B)
aanat2 mRNA expression level, (C) MT1 mRNA expression
level, (D) MT2 mRNA expression level and (E) stress related
gene nfkb mRNA expression level. The data were analyzed by
one way ANOVA followed by dunnett post-hoc test, and was
represented as the mean ± SEM, n=8. **P<0.01; ***P<0.001 vs.
the DEX-2 Vehicle Group. ##P<0.01;
###P<0.001 vs. the DEX-20 Vehicle Group. PB, Piper
betle L. leaf extract; DEX, dexamthasone; aanat1,
arylalkylamine N-acetyltransferase 1; aanat2, arylalkylamine
N-acetyltransferase 2; MT1, melatonin receptor type 1A; mRNA,
messenger RNA; MT2, melatonin receptor type 1B; ANOVA, snalaysis of
variance; SEM, standard error of the mean. |
Discussion
Melatonin receptors (MT1 and MT2) are valuable
molecular targets for the drug discovery of sleep related problems.
In preliminary in-vitro studies, PB significantly decreased
the relative luminescence activity as compared the vehicle control,
with 50 µg/ml of PB showing a level of activity that is similar to
that of the MT1 positive control (6-chloro melatonin). This
indicates that PB can act as a full agonist against MT1 at a
concentration of 50 µg/ml as it shows similar efficacy to the
melatonin analogue 6-chloro melatonin. In contrast, PB is only a
partial agonist of MT2 as it produced a sub-maximal response as
compared to the MT2 positive control (2-iodomelatonin), at all
tested concentrations. This could be due to the slight difference
between the MT1 and MT2 receptors, in which the MT2 receptor
possesses an additional hydrophobic pocket at the region which
corresponds to the melatonin N1-C2 binding region (44). This suggests that the PB constituents
which bind to the MT1 and MT2 receptors, do not possess a
lipophilic side chain at the melatonin N1 or C2 topological
equivalent. While we did not perform a quantitive LCMS-MS analysis
of our extract, any active constituents are likely to have a
similar structure to that of melatonin and thus future research
could focus on the isolation of the active constituents for further
testing and development. Future studies could also utilise MT1
(Fabomotizole) and MT2 (Luzindole) antagonists as another
confirmatory test.
Melatonin is the principal hormone involved in
multiple physiological processes, including the sleep-wake cycle.
It is functionally associated with sleep only in diurnally active
species (45). For this reason, we
have chosen melatonin as a positive control in our experiments with
the diurnal zebrafish. Zebrafish larvae exposed to DEX (2 and 20
mg/l) show a decrease in the level of melatonin production at night
time in the adult zebrafish brain. This clearly indicates the
interference of the glucocorticoid DEX in the formation of
melatonin. A number of studies have shown that melatonin synthesis
is influenced by glucocorticoids and is negatively affected by
stress in numerous animals (19,25,46,47).
These findings suggest that neonatal exposure of DEX could affect
the neuronal and hormonal signaling pathways related to melatonin
synthesis.
In addition to this, our data shows increased
zebrafish locomotor activity and decreased time spent in the upper
zone of the tank, which is characteristic of anxiety like behaviour
as seen in previous zebrafish studies (4). Some other studies have also
demonstrated that the DEX can disrupt the glucocorticoid receptor
signaling pathway and alter the neuroendocrine system, which
impacts behaviour such as hyper activity in adulthood (3,23), that
can interfere with sleep behaviour. Furthermore, nfkb mRNA
expression is significantly higher in DEX treated fish in
comparison to normal fish, which suggests that early life stress
has an impact on the expression of stress-activated proteins, which
can further influence normal sleep regulation as sleep is a resting
state with anti-oxidative properties (48).
Nowadays, sleep disorders are becoming a general
societal problem in all age groups of people. Studies suggest that
sleep problems have a negative impact on health and cognitive
performance (49). Stress is the
most powerful contributor to poor sleep and exposure to stress
during childhood has been suggested to be a crucial factor for the
development of sleep disorders in adulthood (5). Currently, many substances are available
for the treatment of sleep disorders, but they have some sort of
side effects such as daytime sleepiness (50). The interest of researchers in
medicinal plants as a natural source of many active components has
noticeably increased in the past 20 years as drugs against various
pharmacological targets are developed. Nowadays, natural products
are in great demand due to their prevalent biological properties
and for the discovery of many types of effective bioactive
compounds in natural products (33).
In the present study, PB has enhanced sleep like behaviour
(decrease in total distance travelled, increased time spent in the
upper zone and in a quiescent state), which is similar to melatonin
treated group which we have used as a positive control in the
study. Additionally, PB was found to significantly suppress
nfkb mRNA expression in early life stressed fish, similar to
that seen in the melatonin treated group. This suggets that PB has
similar anti-oxidative properties and potentially it can help to
reduce oxidative stress induced by early life stress.
In the present study, we did not find any changes in
aanat1 mRNA expression. This is interesting as in zebrafish,
aanat1 is thought to be expressed exclusively in the retina
and is naturally lower at night when we collected our brain
samples, as it peaks in the late afternoon (51). In the case of aanat2, mRNA
expression was suppressed after the treatment of PB and melatonin
in DEX treated fish. We speculate that increased levels of
melatonin initiated a negative feedback mechanism to suppress
aanat2 expression and downregulate the further production of
melatonin as it has reached peak levels. This is because rhythmic
transcription of aanat ensures melatonin production is turned on
only after the onset of darkness (52). For a clear mechanism of action of
aanat2 in zebrafish brain, further research should be
carried out.
Moreover, it is well established that the effect of
melatonin on sleep is mediated by melatonin receptors (9). In the present study, PB significantly
increased the MT2 mRNA expression in the DEX treated fish,
which is similar to melatonin treated group which we have used as a
positive control. Increased expression of MT2 mRNA suggests
that it facilitates the expression of melatonin to maintain an
optimum level throughout the dark h to expedite physiological
activities. Pandi-Perumal et al (53) has suggested that melatonin exerts its
physiological actions by interacting with melatonin receptors. On
the other hand, there was no significant change in MT1 mRNA
expression. Further investigations are necessary to examine whether
MT1 exhibits different expression patterns in the zebrafish
brain which might explain the discrepancies observed between both
types of receptors. Alternatively, the results could be explained
by the high level of exogenous melatonin in the positive control
group. This would trigger the same negative feedback mechanisms
that result in a reduced level of annat2 and hence melatonin
production as previously discussed. In addition, melatonin
undergoes biexponential decay in blood plasma, with a first
distribution half-life of 2 min and a second metabolic half-life of
20 min due to melatonin catabolism by the liver (54). It is possible that these correction
mechanisms results in an overshoot, causing melatonin levels to
fall below their normal concentration at ZT16 when the zebrafish
were removed for gene expression analysis, four h after treatment.
As the level of melatonin falls below normal, MT2 expression
is upregulated to compensate for this, but not MT1 as
melatonin has a higher affinity for MT1 as compared to MT2
(55). The PB extract at a dose of
30 mg/kg also shows a similar effect on the melatonin related
genes, lending credence to the idea that it can be used to improve
sleep disruption as a result of a deficiency in melatonin.
Taken together, the results of the present study
demonstrated that early life exposure of glucocorticoid (DEX) has a
negative impact on the development of the brain and produces
long-term effects that persist till adulthood. This is
chacracterised by anxious behaviour, a decrease in melatonin
production and increased nfkb mRNA expression. Our results
also suggest that PB could be effective against such conditions and
it is a promising candidate for the improvement of sleep disruption
related to early life stress.
Acknowledgements
Not applicable.
Funding
The present study was supported by Fundamental
Research Grant Scheme (FRGS), Ministry of Education Malaysia (MOE)
(grant no. FRGS/1/2014/SG03/MUSM/03/2).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YK performed all the experiments and was responsible
for the writing of the manuscript in its entirety. BC, MFS and IO
were involved in conceptualizing and proofreading. All authors gave
their final approval for the submission of the manuscript.
Ethics approval and consent to
participate
All experimental procedures implemented in this
study follow the guidelines on the care and the use of animals,
with ethical approval by the Monash Animal Research Platform (MARP)
Animal Ethics Committee, Monash University, Australia
(MARP/2014/133).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DEX
|
dexamethasone
|
|
PB
|
Piper betle L. leaves
extract
|
|
ZT
|
Zeitgeber time
|
|
MT1
|
melatonin receptor type 1A
|
|
MT2
|
melatonin receptor type 1B
|
|
aanat1
|
arylalkylamine N-acetyltransferase
1
|
|
aanat2
|
arylalkylamine N-acetyltransferase
2
|
|
nfkb
|
nuclear factor of kappa light
polypeptide gene enhancer in B-cells
|
|
eef1a1b
|
eukaryotic translation elongation
factor 1 alpha 1b
|
References
|
1
|
Lupien SJ, McEwen BS, Gunnar MR and Heim
C: Effects of stress throughout the lifespan on the brain,
behaviour and cognition. Nat Rev Neurosci. 10:434–445. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Welberg LA and Seckl JR: Prenatal stress,
glucocorticoids and the programming of the brain. J
Neuroendocrinol. 13:113–128. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nagano M, Ozawa H and Suzuki H: Prenatal
dexamethasone exposure affects anxiety-like behaviour and
neuroendocrine systems in an age-dependent manner. Neurosci Res.
60:364–371. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Khor YM, Soga T and Parhar IS: Caffeine
neuroprotects against dexamethasone-induced anxiety-like behaviour
in the Zebrafish (Danio rerio). Gen Comp Endocrinol. 181:310–315.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Greenfield EA, Lee C, Friedman EL and
Springer KW: Childhood abuse as a risk factor for sleep problems in
adulthood: Evidence from a US national study. Ann Behav Med.
42:245–256. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kirby ED, Muroy SE, Sun WG, Covarrubias D,
Leong MJ, Barchas LA and Kaufer D: Acute stress enhances adult rat
hippocampal neurogenesis and activation of newborn neurons via
secreted astrocytic FGF2. Elife. 2:e003622013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kohn N, Hermans EJ and Fernández G:
Cognitive benefit and cost of acute stress is differentially
modulated by individual brain state. Soc Cogn Affect Neurosci.
12:1179–1187. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Walker MP: Cognitive consequences of sleep
and sleep loss. Sleep Med. 9 (Suppl 1):S29–S34. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhdanova IV, Wang SY, Leclair OU and
Danilova NP: Melatonin promotes sleep-like state in zebrafish.
Brain Res. 903:263–268. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mintz EM, Phillips NH and Berger RJ:
Daytime melatonin infusions induce sleep in pigeons without
altering subsequent amounts of nocturnal sleep. Neurosci Lett.
258:61–64. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhdanova IV, Cantor ML, Leclair OU,
Kartashov AI and Wurtman RJ: Behavioral effects of melatonin
treatment in non-human primates. Sleep Res Online. 1:114–118.
1998.PubMed/NCBI
|
|
12
|
Zhdanova IV, Geiger DA, Schwagerl AL,
Leclair OU, Killiany R, Taylor JA, Rosene DL, Moss MB and Madras
BK: Melatonin promotes sleep in three species of diurnal nonhuman
primates. Physiol Behav. 75:523–529. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dollins AB, Zhdanova IV, Wurtman RJ, Lynch
HJ and Deng MH: Effect of inducing nocturnal serum melatonin
concentrations in daytime on sleep, mood, body temperature and
performance. Proc Natl Acad Sci USA. 91:1824–1828. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Iguchi H, Kato KI and Ibayashi H:
Age-dependent reduction in serum melatonin concentrations in
healthy human subjects. J Clin Endocrinol Metab. 55:27–29. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sack RL, Lewy AJ, Erb DL, Vollmer WM and
Singer CM: Human melatonin production decreases with age. J Pineal
Res. 3:379–388. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Haimov I, Laudon M, Zisapel N, Souroujon
M, Nof D, Shlitner A, Herer P, Tzischinsky O and Lavie P: Sleep
disorders and melatonin rhythms in elderly people. BMJ.
309:1671994. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhdanova I, Yu L, Lopez-Patino M, Shang E,
Kishi S and Guelin E: Aging of the circadian system in zebrafish
and the effects of melatonin on sleep and cognitive performance.
Brain Res Bull. 75:433–441. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nikaido Y, Aluru N, McGuire A, Park YJ,
Vijayan MM and Takemura A: Effect of cortisol on melatonin
production by the pineal organ of tilapia, Oreochromis mossambicus.
Comp Biochem Physiol A Mol Integr Physiol. 155:84–90. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Larson ET, Winberg S, Mayer I, Lepage O,
Summers CH and Øverli Ø: Social stress affects circulating
melatonin levels in rainbow trout. Gen Comp Endocrinol.
136:322–327. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roseboom PH, Coon SL, Baler R, McCune SK,
Weller JL and Klein DC: Melatonin synthesis: Analysis of the more
than 150-fold nocturnal increase in serotonin N-acetyltransferase
messenger ribonucleic acid in the rat pineal gland. Endocrinology.
137:3033–3045. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Coon SL, Bégay V, Deurloo D, Falcón J and
Klein DC: Two arylalkylamine N-acetyltransferase genes mediate
melatonin synthesis in fish. J Biol Chem. 274:9076–9082. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Munck A and Náray-Fejes-Tóth A:
Glucocorticoids and stress: Permissive and suppressive actions. Ann
N Y Acad Sci. 746:115–130. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harris A and Seckl J: Glucocorticoids,
prenatal stress and the programming of disease. Horm Behav.
59:279–289. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tongjaroenbuangam W, Ruksee N, Mahanam T
and Govitrapong P: Melatonin attenuates dexamethasone-induced
spatial memory impairment and dexamethasone-induced reduction of
synaptic protein expressions in the mouse brain. Neurochem Int.
63:482–491. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Benyassi A, Schwartz C, Ducouret B and
Falcón J: Glucocorticoid receptors and serotonin
N-acetyltransferase activity in the fish pineal organ. Neuroreport.
12:889–892. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Meneses-Santos D, Buonfiglio DDC,
Peliciari-Garcia RA, Ramos-Lobo AM, Souza DDN, Carpinelli AR,
Carvalho CRO, Sertie RAL, Andreotti S, Lima FB, et al: Chronic
treatment with dexamethasone alters clock gene expression and
melatonin synthesis in rat pineal gland at night. Nat Sci Sleep.
10:203–215. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Van den Berg R, Haenen G, Van den Berg H
and Bast A: Transcription factor NF-kappaB as a potential biomarker
for oxidative stress. Br J Nutr. 86 (Suppl 1):S121–S127. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Baydas G, Ozer M, Yasar A, Koz S and Tuzcu
M: Melatonin prevents oxidative stress and inhibits reactive
gliosis induced by hyperhomocysteinemia in rats. Biochemistry
(Mosc). 71 (Suppl 1):S91–S95. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gilad E, Wong HR, Zingarelli B, Virág L,
O'Connor M, Salzman AL and Szabó C: Melatonin inhibits expression
of the inducible isoform of nitric oxide synthase in murine
macrophages: Role of inhibition of NFkappaB activation. FASEB J.
12:685–693. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kumar N, Misra P, Dube A, Bhattacharya S,
Dikshit M and Ranade S: Piper betle Linn. a maligned Pan-Asiatic
plant with an array of pharmacological activities and prospects for
drug discovery. Cur Sci. 99:922–932. 2010.
|
|
31
|
Dwivedi V and Tripathi S: Review study on
potential activity of Piper betle. J Pharmacogn Phytochem. 3:93–98.
2014.
|
|
32
|
Alam B, Akter F, Parvin N, Pia RS, Akter
S, Chowdhury J, Sifath-E-Jahan K and Haque E: Antioxidant,
analgesic and anti-inflammatory activities of the methanolic
extract of Piper betle leaves. Avicenna J Phytomed. 3:112–125.
2013.PubMed/NCBI
|
|
33
|
Swati B and Madhusudhanan N:
Antidepressant activity of ethanolic extract of piper betle leaves
in mice. Cur Res Neuroscience. 2:11–16. 2012. View Article : Google Scholar
|
|
34
|
Hoo JY, Kumari Y, Shaikh MF, Hue SM and
Goh BH: Zebrafish: a versatile animal model for fertility research.
BioMed Res Int. 2016:97327802016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Levin ED: Zebrafish assessment of
cognitive improvement and anxiolysis: Filling the gap between in
vitro and rodent models for drug development. Rev Neurosci.
22:75–84. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Idda ML, Bertolucci C, Vallone D, Gothilf
Y, Sánchez-Vázquez FJ and Foulkes NS: Chapter 3-Circadian clocks:
Lessons from fish. Prog Brain Res. 199:41–57. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kazimi N and Cahill GM: Development of a
circadian melatonin rhythm in embryonic zebrafish. Brain Res Dev
Brain Res. 117:47–52. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhdanova IV: Sleep and its regulation in
zebrafish. Rev Neurosci. 22:27–36. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ganguly S, Mula S, Chattopadhyay S and
Chatterjee M: An ethanol extract of Piper betle Linn. mediates its
anti-inflammatory activity via down-regulation of nitric oxide. J
Pharm Pharmacol. 59:711–718. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen Y, Xu Z, Wu D, Li J, Song C, Lu W and
Huang J: Luciferase reporter gene assay on human 5-HT receptor:
Which response element should be chosen? Sci Rep. 5:80602015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Graham FL, Smiley J, Russell W and Nairn
R: Characteristics of a human cell line transformed by DNA from
human adenovirus type 5. J Gen Virol. 36:59–74. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kundap UP, Kumari Y, Othman I and Shaikh
M: Zebrafish as a model for epilepsy-induced cognitive dysfunction:
A pharmacological, biochemical and behavioral approach. Front
Pharmacol. 8:5152017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cuesta A, Cerezuela R, Esteban MA and
Meseguer J: In vivo actions of melatonin on the innate immune
parameters in the teleost fish gilthead seabream. J Pineal Res.
45:70–78. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zlotos DP: Recent progress in the
development of agonists and antagonists for melatonin receptors.
Curr Med Chem. 19:3532–3549. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pandi-Perumal SR, Srinivasan V, Spence DW
and Cardinali DP: Role of the melatonin system in the control of
sleep: Therapeutic implications. CNS Drugs. 21:995–1018. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zawilska JB and Sadowska M: Prolonged
treatment with glucocorticoid dexamethasone suppresses melatonin
production by the chick pineal gland and retina. Pol J Pharmacol.
54:61–66. 2002.PubMed/NCBI
|
|
47
|
López-Patiño MA, Gesto M, Conde-Sieira M,
Soengas JL and Míguez JM: Stress inhibition of melatonin synthesis
in the pineal organ of rainbow trout (Oncorhynchus mykiss) is
mediated by cortisol. J Exp Biol. 217:1407–1416. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Villafuerte G, Miguel-Puga A, Rodríguez
EM, Machado S, Manjarrez E and Arias-Carrión O: Sleep deprivation
and oxidative stress in animal models: A systematic review. Oxid
Med Cell Longev. 2015:2349522015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Durmer JS and Dinges DF: Neurocognitive
consequences of sleep deprivation. Semin Neurol. 25:117–129. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Pagel J and Parnes BL: Medications for the
treatment of sleep disorders: An overview. Prim Care Companion J
Clin Psychiatry. 3:118–125. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Appelbaum L, Vallone D, Anzulovich A, Ziv
L, Tom M, Foulkes N and Gothilf Y: Zebrafish
arylalkylamine-N-acetyltransferase genes-targets for regulation of
the circadian clock. J Mol Endocrinol. 36:337–347. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Klein DC: Arylalkylamine
N-acetyltransferase: ‘The Timezyme’. J Biol Chem. 282:4233–4237.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Pandi-Perumal SR, Trakht I, Srinivasan V,
Spence DW, Maestroni GJ, Zisapel N and Cardinali DP: Physiological
effects of melatonin: Role of melatonin receptors and signal
transduction pathways. Prog Neurobiol. 85:335–353. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Claustrat B, Brun J and Chazot G: The
basic physiology and pathophysiology of melatonin. Sleep Med Rev.
9:11–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Emet M, Ozcan H, Ozel L, Yayla M, Halici Z
and Hacimuftuoglu A: A review of melatonin, its receptors and
drugs. Eurasian J Med. 48:135–141. 2016. View Article : Google Scholar : PubMed/NCBI
|