Introduction
Lung cancer is one of the most lethal types of
cancer, accounting for ~18% of cancer-associated mortalities
worldwide (1). Lung cancer is
divided into two major subtypes, namely small-cell lung cancer and
non-small-cell lung cancer (NSCLC) (2). NSCLC accounts for >80% of all lung
cancer cases and its prognosis is relatively poor due to its highly
aggressive nature (3,4). It is crucial to elucidate the precise
molecular mechanisms underlying NSCLC progression in order to
identify novel biomarkers and therapeutic targets for the treatment
of patients with NSCLC.
microRNAs (miRNAs or miRs) are small, non-coding,
single-stranded RNA molecules that are involved in the regulation
of gene expression in eukaryotes (5). miRNA deregulation is frequently
observed in human diseases, including different types of cancer
(6,7). During cancer initiation and
progression, miRNAs serve an important role through downregulation
of oncogenes and/or tumor suppressors, thereby affecting tumor cell
proliferation, metastasis and apoptosis (8–10). In
NSCLC, several miRNAs have been found to be deregulated and
demonstrated to promote cancer progression or drug resistance via
repression of target genes (11,12). The
expression of several of those miRNAs in tumor tissues, plasma and
exosomes has been considered a promising diagnostic and therapeutic
biomarker for patients with NSCLC (13). miR-448 has been identified as a tumor
suppressor in several cancer types, including breast,
hepatocellular and colorectal cancer (14–16). A
recent study reported that low expression of miR-448 predicted poor
prognosis of lung squamous cell carcinoma (SCC), a subtype of
NSCLC, and it suppressed SCC cell proliferation and metastasis via
targeting DCLK1 (17).
Silent mating-type information regulation 2
homologue 1 (sirtuin1; SIRT1) is a member of the sirtuin family
that acts as an NAD-dependent protein deacetylase (18). Through regulating the acetylation
level of target proteins, such as p53, SIRT1 participates in a
number of cellular events that are implicated in cancer progression
(19,20). Accumulating evidence indicates that
overexpression of SIRT1 promotes cancer progression, including
hepatocellular carcinoma and breast cancer (21,22). In
NSCLC, overexpression of SIRT1 has been associated with poor
prognosis (23). However, the
regulatory mechanism of SIRT1 in NSCLC remains unknown.
In the present study, a significant reduction of
miR-448 levels and elevation of SIRT1 mRNA levels were observed in
tumor samples from patients with NSCLC. In vitro experiments
demonstrated that enhanced expression of miR-448 inhibited the
proliferation and migration of A549 cells. Additionally,
overexpression of miR-448 decreased SIRT1 mRNA and protein levels,
thus inhibiting epithelial-mesenchymal transition (EMT). Activation
of SIRT1 partially rescued miR-448-induced cell growth arrest. The
findings of the present study suggest a pivotal role for miR-448 in
the inhibition of NSCLC progression via targeting SIRT1.
Materials and methods
Clinical specimens
Tumor tissues and adjacent normal tissues were
collected from 25 patients (17 males and 8 females; aged 35–65
years) with NSCLC undergoing surgery at Changyi People's Hospital
(Weifang, China) between September 2, 2014 and May 4, 2016. None of
the patients received chemotherapy or radiotherapy prior to
surgery. Written informed consent was obtained from all
participants and the clinical experiments were approved by the
Ethics Committee of Changyi People's Hospital. All specimens were
immediately stored at −80°C until use in subsequent
experiments.
Cell lines and reagents
293T cells and the human NSCLC cell line A549 were
purchased from American Type Culture Collection (Manassas, VA, USA)
and used within 6 months. A549 cells and 293T cells were cultured
in Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific) in a 37°C
incubator with 5% CO2.
miR-negative control (NC) mimics and miR-448 mimics
were purchased from Shanghai GenePharma Co., Ltd. (Shanghai,
China). The sequences were as follows: miR-NC mimics,
5′-UCGCUUGGUGCAGGUCGGGAA-3′; miR-448 mimics,
5′-UUGCAUAUGUAGGAUGUCCCAU-3′. Transfection of 50 nM miR-NC mimics
or miR-448 mimics was performed with Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. After 48 h, cells were subjected to
experiments.
Resveratrol was obtained from Sigma-Aldrich; Merck
KGaA (Darmstadt, Germany) and dissolved in dimethyl sulfoxide to a
concentration of 50 mM for storage. For the resveratrol + miR-448
mimics group, cells were firstly transfected with 50 nM miR-448
mimics using Lipofectamine 2000, then resveratrol was diluted and
added into the culture medium at a final concentration of 20 µM for
SIRT1 activation for 48 h at 37°C. The DMSO + miR-448 mimics and
DMSO + miR-NC mimics group was transfected with 50 nM miR-448
mimics miR-NC mimics using Lipofectamine 2000 followed by treatment
with 20 µM DMSO for 48 h at 37°C.
Western blotting
Whole proteins were extracted from A549 cells using
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China). Pierce BCA Protein Assay kit
(Pierce; Thermo Fisher Scientific, Inc.) was used to determine the
concentration of lysates according to the manufacturer's protocol.
In brief, 20 µg protein lysates were loaded into each lane of 8%
gels, separated by SDS-PAGE for 30 min and transferred onto
polyvinylidene fluoride (PVDF) membranes. The PVDF membranes were
blocked with 5% non-fat milk for 1 h at room temperature and
incubated with the indicated primary antibodies overnight at 4°C.
On the next day, the PVDF membranes were incubated with the
secondary antibodies for 1 h at room temperature and developed with
enhanced chemiluminescence substrate (EMD Millipore, Billerica, MA,
USA). Antibodies against SIRT1 (cat. no. 8469), vimentin (cat. no.
5741) and E-cadherin (cat. no. 14472; all 1:1,000) were supplied by
Cell Signaling Technology, Inc. (Danvers, MA, USA). Antibodies
against GAPDH (cat. no. G8795; 1:5,000) were purchased from
Sigma-Aldrich; Merck KGaA. Horseradish peroxidase-conjugated
anti-mouse (cat. no. SA00001-1) and anti-rabbit (cat. no.
SA00001-2; both 1:10,000) secondary antibodies were purchased from
ProteinTech Group, Inc. (Chicago, IL, USA). Images were captured
with ImageQuant300 (GE Healthcare, Chicago, IL) and analyzed using
ImageJ software (version 1.8.0; National Institutes of Health,
Bethesda, MD, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from A549 cells, as well as
NSCLC tissues and adjacent normal tissues using TRIzol®
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The
concentration of RNA was measured with NanoDrop™ 2000 (Thermo
Fisher Scientific, Inc.).
To measure miR-448 levels, RNA was
reverse-transcribed into cDNA using a TaqMan MicroRNA Reverse
Transcription kit (Thermo Fisher Scientific, Inc.). RT-PCR was
performed using a TaqMan MicroRNA Assay kit (Thermo Fisher
Scientific, Inc.) on the CFX-96 module (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). U6 served as an internal control for miR-448
expression. For the detection of SIRT1, vimentin and E-cadherin
mRNA levels, RNA was reverse-transcribed into cDNA using
PrimeScript RT Master Mix (Takara Bio, Inc., Otsu, Japan). GAPDH
served as an internal control. The thermocycling conditions for
mRNA and miRNA analyses were: Pre-denaturing at 95°C for 30 sec,
and 35 cycles of denaturing at 95°C for 10 sec, annealing and
elongation at 60°C for 10 sec. The relative expression of gene was
calculated using the 2−ΔΔCq method (24). The primer sequences were: Stem loop:
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGATGGGA-3′; miR-448-forward,
5′-TCGGCAGGTTGCATATGTAGGA-3′ and reverse, 5′-CTCAACTGGTGTCGTGGA-3′;
U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′; Vimentin forward,
5′-GACGCCATCAACACCGAGTT-3′ and reverse,
5′-CTTTGTCGTTGGTTAGCTGGT-3′; E-cadherin-forward,
5′-CGAGAGCTACACGTTCACGG-3′ and reverse,
5′-GGGTGTCGAGGGAAAAATAGG-3′; SIRT1 forward,
5′-TAGCCTTGTCAGATAAGGAAGGA-3′ and SIRT1 reverse,
5′-ACAGCTTCACAGTCAACTTTGT-3′; GAPDH forward,
5′-CCTGCACCACCAACTGCTTA-3′ and reverse,
5′-GGCCATCCACAGTCTTCTGAG-3′.
Dual luciferase reporter assay
To further investigate whether miR-448 directly
regulates SIRT1 expression in A549 cells, the online bioinformatics
tool TargetScan (www.targetscan.org) was used to predict miR-448
potential binding sites in SIRT1. The 3′ untranslated region (UTR)
of SIRT1 mRNA was amplified from cDNA of 293T cells and ligated
into the pGL3 plasmid (Promega Corporation, Madison, WI, USA) to
construct pGL3-SIRT1-3′UTR-wild-type (WT). pGL3-SIRT1-3′UTR-mutant
(Mut) was obtained by introducing 2 site mutations into miR-448
potential binding sites using a Quicksite mutation kit (Agilent
Technologies, Inc., Santa Clara, CA, USA). pGL3-SIRT1-3′UTR-WT or
pGL3-SIRT1-3′UTR-Mut was co-transfected with Renilla plasmid
into A549 cells using Lipofectamine 2000, followed by transfection
of miR-NC mimics or miR-448 mimics. Following 48 h, a Dual
Luciferase Reporter Assay system (Promega Corporation) was used to
measure the relative luciferase activity of each well. In each
group, firefly luciferase activity was normalized to Renilla
luciferase activity. Subsequently, the relative luciferase activity
of control group was normalized to 1 and was compared with the
treatment group.
Cell proliferation assay
The proliferation of A549 cells was measured using a
Cell Counting kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). Subsequently, 10 µl CCK-8 solution was added into
each well and sustained for 2 h. Immediately following, and 24, 48,
72 and 96 h after the incubation, the medium containing CCK-8 was
transferred into a clean 96-well plate and the absorbance at 450 nm
of each well was detected with a microplate reader (Bio-Rad
Laboratories, Inc.).
Wound-healing assay
The cell migration ability was measured using a
wound-healing assay. Briefly, 2×106 cells were plated in
a 6-well plate and cultured until reaching 100% confluence. A wound
was made by scratching the middle area of each well using a 10-µl
pipette. After washing with PBS, DMEM without FBS was added into
each well and images of the wound areas were captured immediately
using an inverted microscope at a magnification of ×40. After the
cells were cultured for 40 h, images of wound areas were captured
again. The relative migrated areas were calculated using Image
Pro-Plus version 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Statistical analysis
All data in the present study were analyzed using
GraphPad Prism 7.0 (GraphPad Software, Inc., La Jolla, CA, USA) and
the results are expressed as the mean + standard deviation.
Statistical analysis between two groups was performed using
Student's t-test, and the differences between normal tissues and
tumor tissues were analyzed by paired Student's t-test. The
differences among three different groups were analyzed using
one-way analysis of variance followed by Tukey's analysis. The
correlation analysis was carried out using Pearson Correlation
analysis. P<0.05 was considered to indicate statistically
significant differences.
Results
Expression levels of miR-448 and SIRT1
in tumor tissues and normal tissues from patients with NSCLC
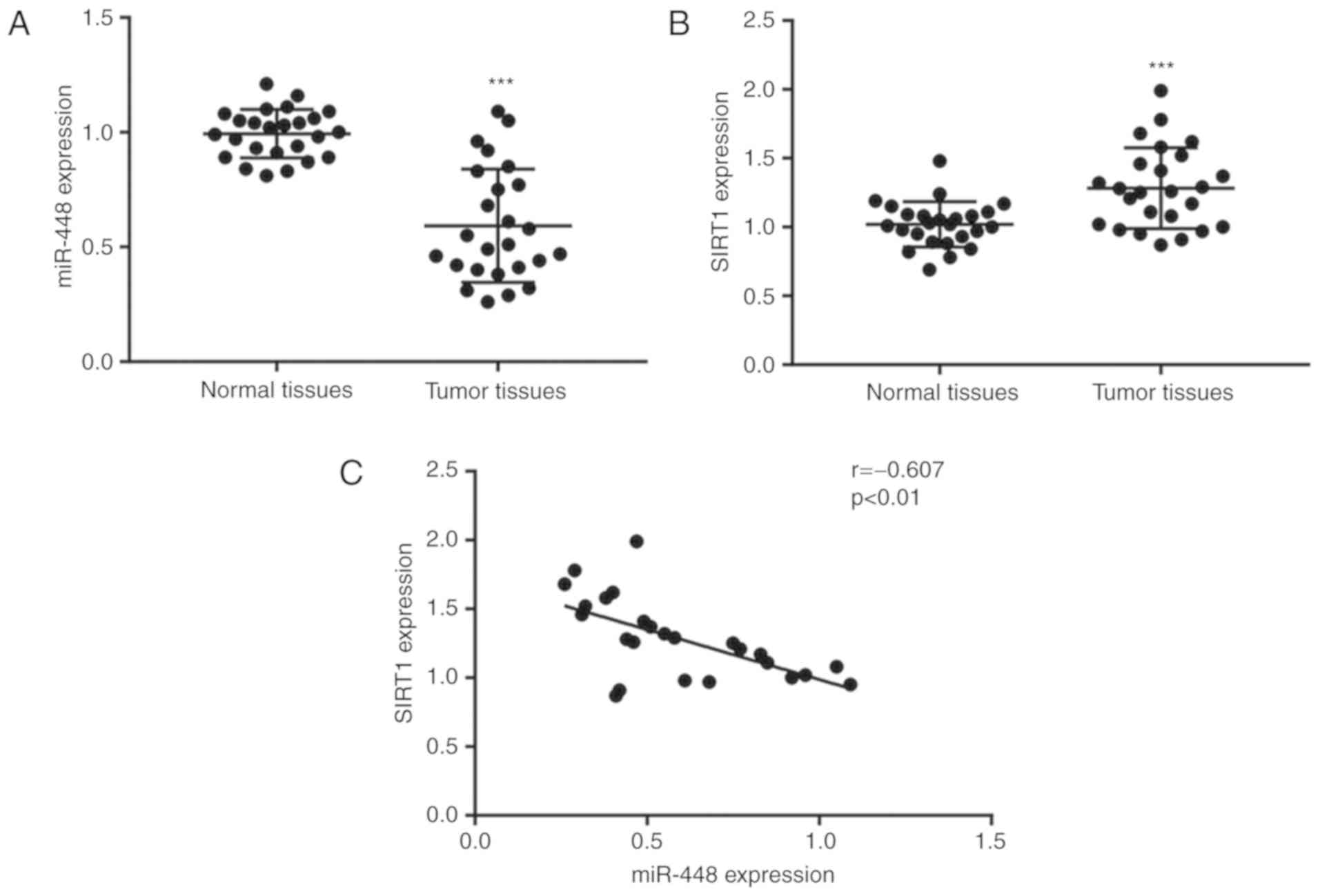
To determine the expression levels of miR-448 and
SIRT1 in NSCLC, tumor tissues and matched normal tissues from 25
patients with NSCLC were used for RT-qPCR. Compared with the normal
tissues, the expression of miR-448 was significantly lower, while
the SIRT1 mRNA levels were significantly higher in tumor tissues
(Fig. 1A and B). Their different
expression pattern suggested a potential association between the
two. Pearson's correlation analysis of miR-448 levels and SIRT1
mRNA levels revealed a strong negative correlation between their
expressions in NSCLC tumor tissues (Fig.
1C), suggesting a potential regulatory association between
them.
Overexpression of miR-448 suppressed
the proliferation and migration of A549 cells
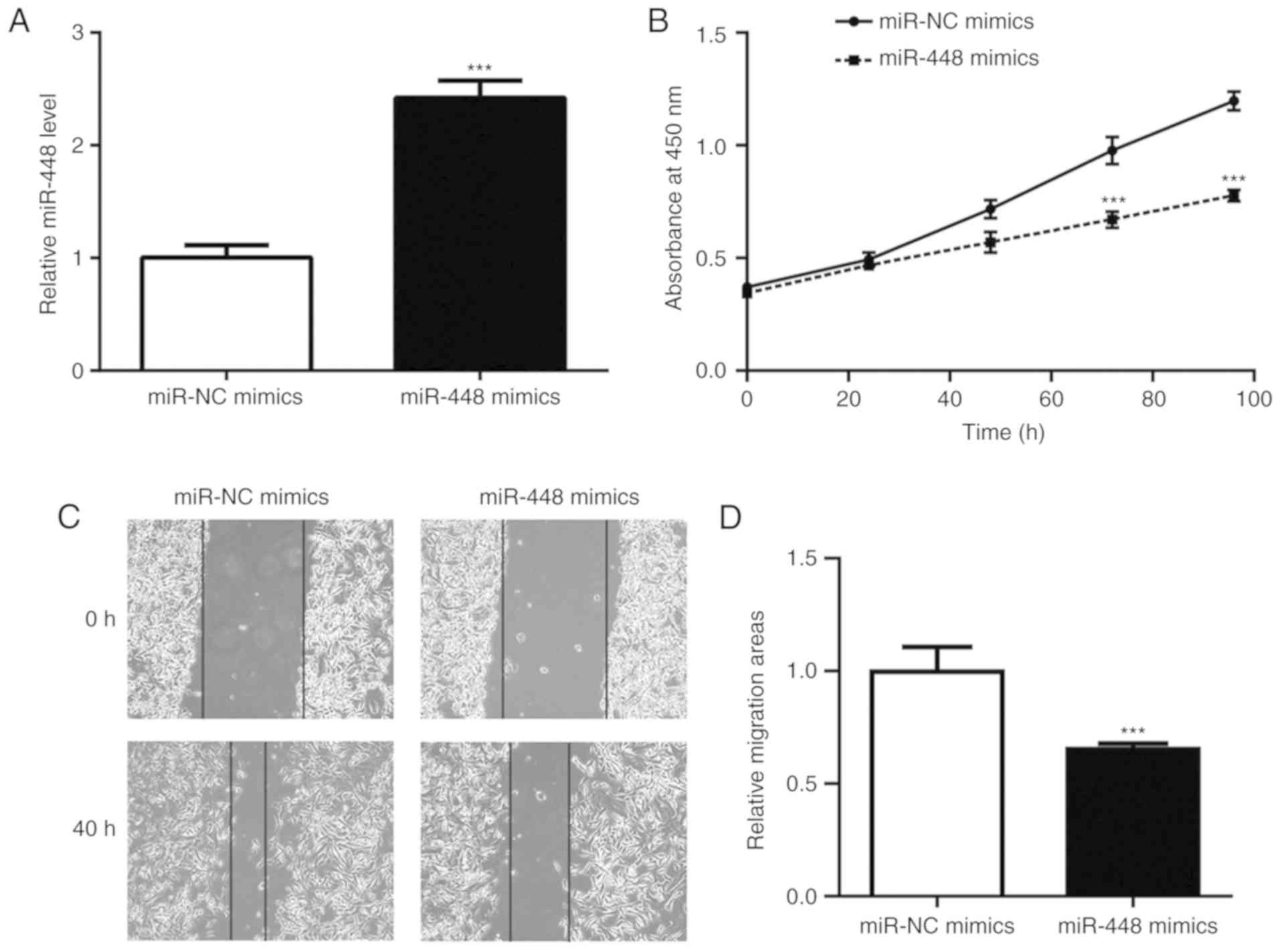
To investigate the role of miR-448 in NSCLC cells,
the A549 NSCLC cell line was used to perform the associated
functional studies and experiments. Transfection of miR-448 mimics
markedly elevated the miR-448 level in A549 cells (Fig. 2A). Overexpression of miR-448
significantly reduced cell viability and wound healing, indicating
its role in the inhibition of cell proliferation and cell migration
(Fig. 2B-D).
miR-448 repressed EMT through
downregulation of SIRT1
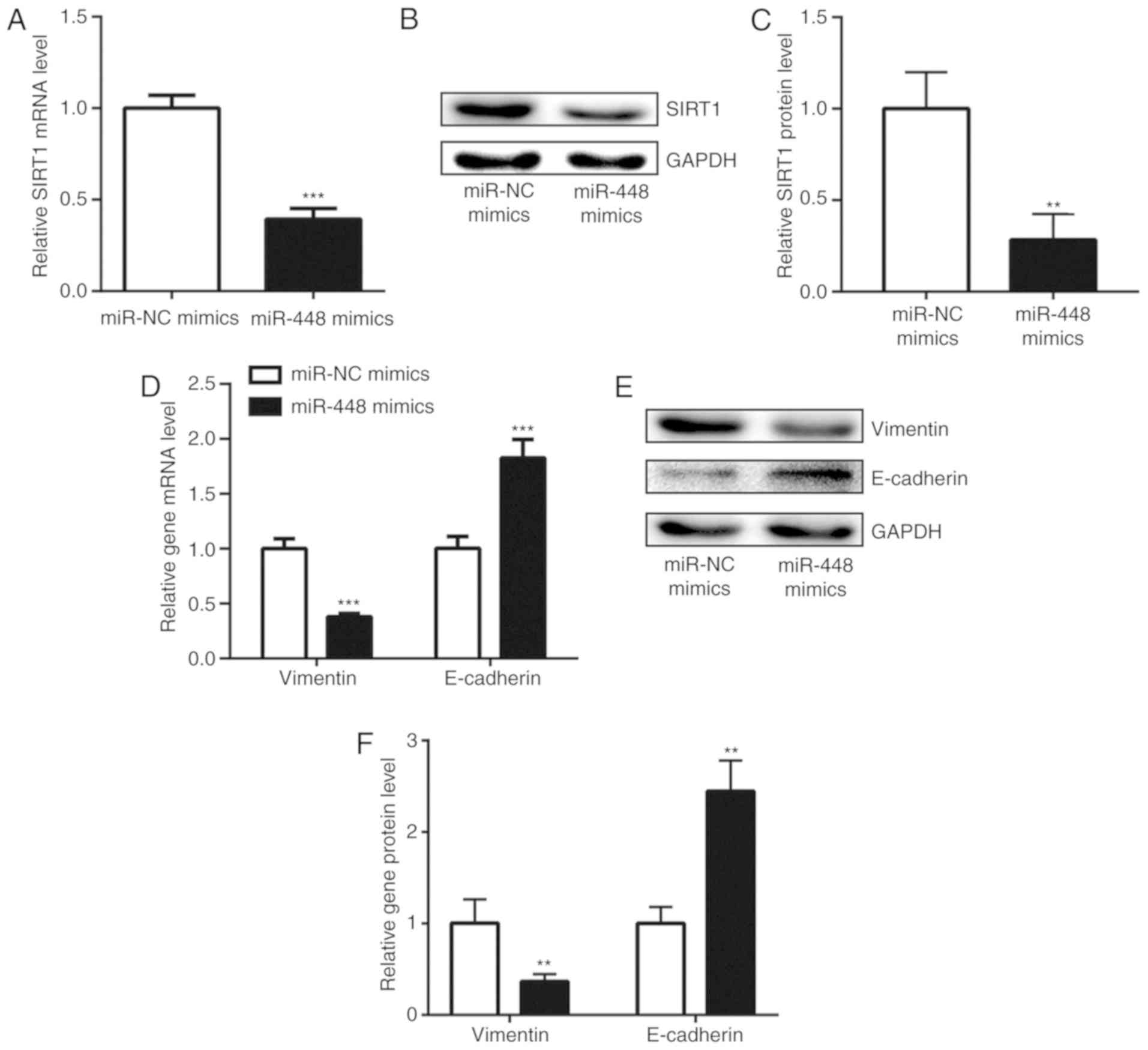
The negative correlation between miR-448 and SIRT1
mRNA levels indicated a regulatory association between miR-448 and
SIRT1. Consistent with the correlation analysis, enhanced
expression of miR-448 significantly decreased SIRT1 mRNA level in
A549 cells (Fig. 3A). Additionally,
the protein level of SIRT1 was also reduced with miR-448
overexpression (Fig. 3B and C).
SIRT1 was previously reported to promote NSCLC cell proliferation
and metastasis via inducing EMT (25). Increase of miR-448 levels
downregulated vimentin and upregulated E-cadherin expression at
both the mRNA and protein levels in A549 cells (Fig. 3D-F), suggesting that miR-448 may
inhibit EMT through repression of SIRT1 expression.
SIRT1 is a direct target of
miR-448
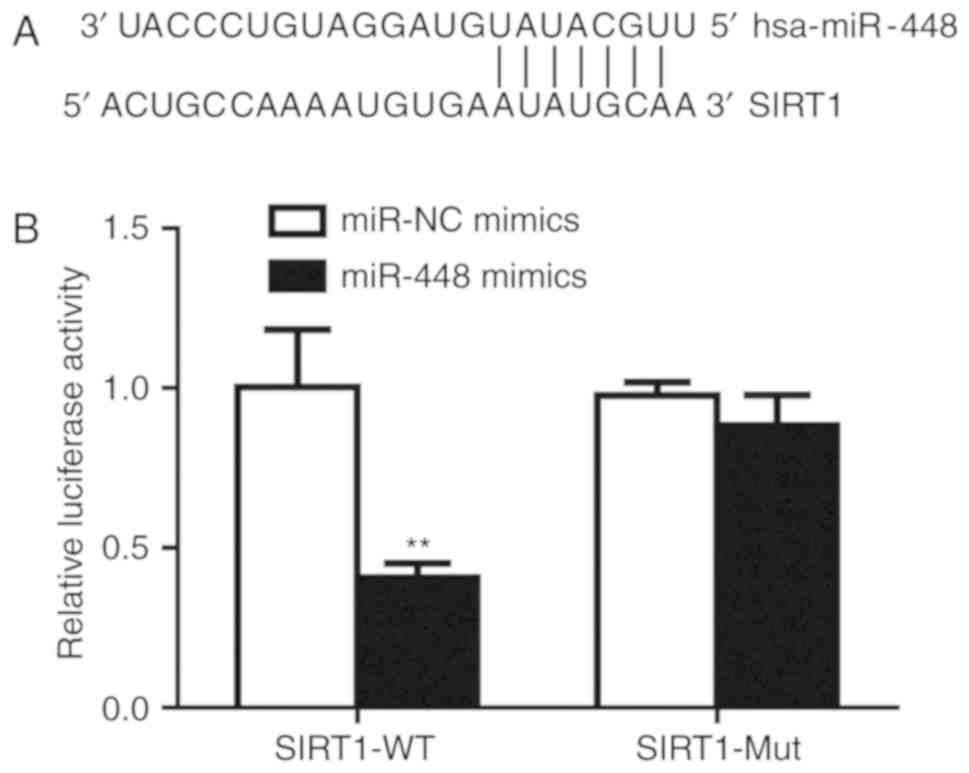
It is observed that there was an miR-448 binding
site in the 3′UTR of SIRT1 mRNA (Fig.
4A). Subsequently, the dual luciferase reporter assay was
performed to validate their association. Transfection of miR-448
mimics significantly decreased relative luciferase activity in A549
cells transfected with SIRT1-3′UTR-WT, but not SIRT1-3′UTR-Mut,
suggesting that miR-448 can bind to SIRT1-3′UTR-WT (Fig. 4B). Therefore, these results confirmed
that miR-448 directly binds to the 3′UTR of SIRT1 mRNA to inhibit
its expression in A549 cells.
Elevation of SIRT1 partially rescued
miR-448 mimics-induced cell growth arrest
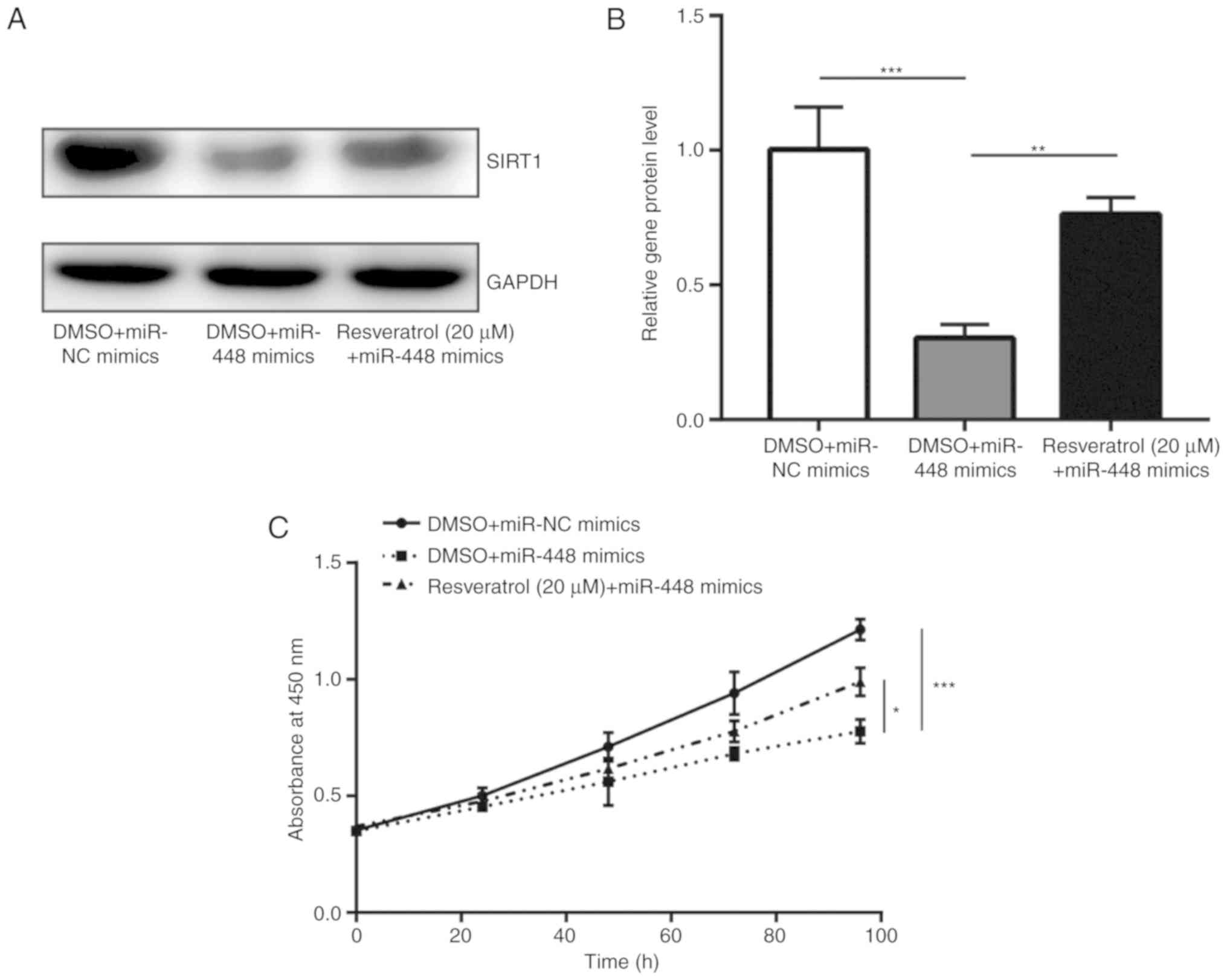
Resveratrol treatment was previously demonstrated to
induce SIRT1 expression and promote cell proliferation in A549
cells (26). In the present study,
it was observed that treatment with resveratrol was able to
partially recover SIRT1 expression, which was downregulated by
miR-448 overexpression (Fig. 5A and
B). Notably, resveratrol partially reversed the cell growth
inhibition induced by miR-448 mimics in A549 cells (Fig. 5C). These data indicated that miR-448
mainly relies on regulation of SIRT1 to suppress NSCLC
progression.
Discussion
Aberrant expression of miRNA is frequent in cancer
cells. The dysregulation and tumor suppressor role of miR-448 has
been reported in various types of cancer (14–16).
However, the role of miR-448 in NSCLC remains controversial. A
previous study reported that miR-448 levels were significantly
higher in plasma samples from patients with NSCLC compared with
those from healthy volunteers, and that it may serve as a potential
biomarker (27). Another recent
study demonstrated that miR-448 was decreased in lung SCC tissues
and cell lines (17). In the present
study, it was observed that miR-448 was able to suppress the
progression of NSCLC through targeting SIRT1.
A previous meta-analysis revealed that high
expression of SIRT1 predicted poor overall survival for patients
with NSCLC (23). Several miRNAs
were reported to repress SIRT1 expression, thereby controlling
cancer cell behavior (28,29). Particularly, miR-138 was demonstrated
to suppress NSCLC cell growth, metastasis and autophagy via
targeting SIRT1 (24). Although a
previous study investigated miR-448 and SIRT1 in patients with
diabetes mellitus (30), their role
in cancer cells, particularly NSCLC cells, remains unknown. In
clinical samples, the expression of SIRT1 was higher in tumor
tissues and its levels were inversely correlated with miR-448
levels. In A549 cells, enhanced expression of miR-448 decreased
SIRT1 mRNA and protein levels. In addition, the present dual
luciferase reporter assay further supported that miR-448 could
directly bind to the SIRT1 3′UTR in A549 cells. miR-448 can also
bind to multiple target gene mRNAs such as EPHA7 and IGF1R in
cancer cells (16,31). It was observed that activation of
SIRT1 could reverse miR-448-induced cell growth inhibition. These
results collectively demonstrated that miR-448 mainly relies on the
regulation of SIRT1 to repress NSCLC progression.
Accumulating evidence has demonstrated that SIRT1 is
an inducer of EMT and promotes cancer progression (32,33). In
the present study, the expression of E-cadherin was increased while
that of vimentin was decreased in A549 cells transfected with
miR-448 mimics, suggesting that miR-448 reverses EMT by repressing
SIRT1 expression. These findings were consistent with those of
previous studies on breast cancer and hepatocellular carcinoma
cells, demonstrating that miR-448 inhibits EMT (15,34). As
EMT is an indicator of highly aggressive growth of cancer cells
(35), miR-448 may inhibit NSCLC
progression via reversing EMT. Taken together, the present findings
demonstrated that miR-448 might be a promising target for the
treatment of NSCLC.
Acknowledgements
Not applicable.
Funding
No funding received.
Availability of data and materials
The data and materials are available from the
corresponding author.
Authors' contributions
HQ, HW and DP designed the study and acquired the
data. HW collected the clinical samples. DP supervised the study,
and prepared and revised the manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Changyi People's Hospital.
Patient consent for publication
Written informed consents were provided by all
patients. The patients consented to the publication of data
presented in the manuscript.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Siegel RL and Jemal A: Lung
cancer statistics. Adv Exp Med Biol. 893:1–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Niklinski J, Niklinska W, Chyczewski L,
Becker HD and Pluygers E: Molecular genetic abnormalities in
premalignant lung lesions: Biological and clinical implications.
Eur J Cancer Prev. 10:213–226. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou J, Song XB, He H, Zhou Y, Lu XJ and
Ying BW: Prevalence and clinical profile of EGFR mutation in
non-small-cell lung carcinoma patients in Southwest China. Asian
Pac J Cancer Prev. 17:965–971. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu S, Zhao X, Wu S, Du R, Zhu Q, Fang H,
Zhang X, Zhang C, Zheng W, Yang J and Feng H: Overexpression of
B7-H3 correlates with aggressive clinicopathological
characteristics in non-small cell lung cancer. Oncotarget.
7:81750–81756. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mendell JT: MicroRNAs: Critical regulators
of development, cellular physiology and malignancy. Cell Cycle.
4:1179–1184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iorio MV and Croce CM: MicroRNAs in
cancer: Small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu T, Wu X, Chen T, Luo Z and Hu X:
Downregulation of DNMT3A by miR-708-5p inhibits lung cancer stem
cell-like phenotypes through repressing wnt/β-catenin signaling.
Clin Cancer Res. 24:1748–1760. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pang W, Tian X, Bai F, Han R, Wang J, Shen
H, Zhang X, Liu Y, Yan X, Jiang F and Xing L: Pim-1 kinase is a
target of miR-486-5p and eukaryotic translation initiation factor
4E, and plays a critical role in lung cancer. Mol Cancer.
13:2402014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Paliouras AR, Monteverde T and Garofalo M:
Oncogene-induced regulation of microRNA expression: Implications
for cancer initiation, progression and therapy. Cancer Lett.
421:152–160. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fadejeva I, Olschewski H and Hrzenjak A:
MicroRNAs as regulators of cisplatin-resistance in non-small cell
lung carcinomas. Oncotarget. 8:115754–115773. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li S, Gao M, Li Z, Song L, Gao X, Han J,
Wang F, Chen Y, Li W, Yang J and Han X: Role of microRNAs in
metastasis of non-small cell lung cancer. Front Biosci (Landmark
Ed). 21:998–1005. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou Q, Huang SX, Zhang F, Li SJ, Liu C,
Xi YY, Wang L, Wang X, He QQ, Sun CC and Li DJ: MicroRNAs: A novel
potential biomarker for diagnosis and therapy in patients with
non-small cell lung cancer. Cell Prolif. 50:123942017. View Article : Google Scholar
|
|
14
|
Li QQ, Chen ZQ, Cao XX, Xu JD, Xu JW, Chen
YY, Wang WJ, Chen Q, Tang F, Liu XP and Xu ZD: Involvement of
NF-κB/miR-448 regulatory feedback loop in chemotherapy-induced
epithelial-mesenchymal transition of breast cancer cells. Cell
Death Differ. 18:16–25. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhu H, Zhou X, Ma C, Chang H, Li H, Liu F
and Lu J: Low expression of miR-448 induces EMT and promotes
invasion by regulating ROCK2 in hepatocellular carcinoma. Cell
Physiol Biochem. 36:487–498. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li B, Ge L, Li M, Wang L and Li Z: miR-448
suppresses proliferation and invasion by regulating IGF1R in
colorectal cancer cells. Am J Transl Res. 8:3013–3022.
2016.PubMed/NCBI
|
|
17
|
Shan C, Fei F, Li F, Zhuang B, Zheng Y,
Wan Y and Chen J: miR-448 is a novel prognostic factor of lung
squamous cell carcinoma and regulates cells growth and metastasis
by targeting DCLK1. Biomed Pharmacother. 89:1227–1234. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bordone L and Guarente L: Calorie
restriction, SIRT1 and metabolism: Understanding longevity. Nat Rev
Mol Cell Biol. 6:298–305. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ong ALC and Ramasamy TS: Role of
Sirtuin1-p53 regulatory axis in aging, cancer and cellular
reprogramming. Ageing Res Rev. 43:64–80. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu T, Liu PY and Marshall GM: The
critical role of the class III histone deacetylase SIRT1 in cancer.
Cancer Res. 69:1702–1705. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Xu S, Li J, Zheng L, Feng M, Wang X,
Han K, Pi H, Li M, Huang X, et al: SIRT1 facilitates hepatocellular
carcinoma metastasis by promoting PGC-1α-mediated mitochondrial
biogenesis. Oncotarget. 7:29255–29274. 2016.PubMed/NCBI
|
|
22
|
Zou Q, Tang Q, Pan Y, Wang X, Dong X,
Liang Z and Huang D: MicroRNA-22 inhibits cell growth and
metastasis in breast cancer via targeting of SIRT1. Exp Ther Med.
14:1009–1016. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Y, Wang T, Wang W, Hu J, Li R, He S
and Yang J: Prognostic and clinicopathological significance of
SIRT1 expression in NSCLC: A meta-analysis. Oncotarget.
8:62537–62544. 2017.PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ye Z, Fang B, Pan J, Zhang N, Huang J, Xie
C, Lou T and Cao Z: miR-138 suppresses the proliferation,
metastasis and autophagy of non-small cell lung cancer by targeting
Sirt1. Oncol Rep. 37:3244–3252. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu X, Shao K and Sun T: SIRT1 regulates
the human alveolar epithelial A549 cell apoptosis induced by
pseudomonas aeruginosa lipopolysaccharide. Cell Physiol Biochem.
31:92–101. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Powrozek T, Krawczyk P, Kowalski DM,
Kuźnar-Kamińska B, Winiarczyk K, Olszyna-Serementa M,
Batura-Gabryel H and Milanowski J: Application of plasma
circulating microRNA-448, 506, 4316, and 4478 analysis for
non-invasive diagnosis of lung cancer. Tumour Biol. 37:2049–2055.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guan Y, Rao Z and Chen C: miR-30a
suppresses lung cancer progression by targeting SIRT1. Oncotarget.
9:4924–4934. 2017.PubMed/NCBI
|
|
29
|
Li T, Ma J, Han X, Jia Y, Yuan H, Shui S
and Guo D: MicroRNA-320 enhances radiosensitivity of glioma through
down-regulation of sirtuin type 1 by directly targeting forkhead
box protein M1. Transl Oncol. 11:205–212. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Y, Wang DS, Cheng YS, Jia BL, Yu G,
Yin XQ and Wang Y: Expression of MicroRNA-448 and SIRT1 and
prognosis of obese type 2 diabetic mellitus patients after
laparoscopic bariatric surgery. Cell Physiol Biochem. 45:935–950.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu X, Yan L, Liu Y, Xian W, Wang L and
Ding X: MicroRNA-448 suppresses osteosarcoma cell proliferation and
invasion through targeting EPHA7. PLoS One. 12:e01755532017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun T, Jiao L, Wang Y, Yu Y and Ming L:
SIRT1 induces epithelial-mesenchymal transition by promoting
autophagic degradation of E-cadherin in melanoma cells. Cell Death
Dis. 9:1362018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ray U, Roy SS and Chowdhury SR:
Lysophosphatidic acid promotes epithelial to mesenchymal transition
in ovarian cancer cells by repressing SIRT1. Cell Physiol Biochem.
41:795–805. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ma P, Ni K, Ke J, Zhang W, Feng Y and Mao
Q: miR-448 inhibits the epithelial-mesenchymal transition in breast
cancer cells by directly targeting the E-cadherin repressor ZEB1/2.
Exp Biol Med (Maywood). 243:473–480. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|