Introduction
Periodontitis is a chronic inflammatory disease that
affects the integrity of the periodontium, which is composed of the
periodontal ligament (PDL), gingiva and alveolar bone (1). Periodontitis leads to tooth loss and
may also induce systemic disorders, including adverse pregnancy
outcomes, atherosclerosis, rheumatoid arthritis, aspiration
pneumonia or cancer (2–4). Periodontitis affects approximately
45–75% of adults worldwide (1–4). In
developing countries, such as China, poor overall oral health has
led to periodontitis becoming a commonly diagnosed disorder in
stomatology departments (5).
Periodontitis is generally treated using systemic antibiotics;
however long-term use is common and more severe disorders, such as
the development of resistance, are inevitable (6). Therefore, the identification of a novel
therapeutic target for the treatment of periodontitis is
required.
The abnormal increased proliferation of periodontal
ligament stem cells (PDLSCs) has been demonstrated to be associated
with the pathogenesis of periodontitis (7,8). It has
been reported that fibroblast growth factor 2 (FGF2) promotes the
proliferation and differentiation of human PDLSCs (8). This suggests that FGF2 may be involved
in the pathogenesis of periodontitis. In addition, FGF2 protein
expression and secretion has been revealed to be positively
regulated by long non-coding RNA (lncRNA) metastasis-associated
lung adenocarcinoma transcript 1 (MALAT1) (9), which is a well-characterized lncRNA in
cancer biology (10,11). In the current study, the involvement
of MALAT1 and FGF2 in periodontitis was assessed and the results
demonstrated that MALAT1 promoted the expression of FGF2 and the
proliferation of PDLSCs, which serve a role in periodontitis.
Materials and methods
Patients
All patients included in the current study were
diagnosed and treated at The Stomatological Hospital of Southern
Medical University (Guangzhou, China) from January 2017 to January
2018. All patients were treated with antibiotics in combination
with pocket reduction surgery. The diagnostic criteria for
periodontitis and the exclusion and inclusion criteria for the
present study were essentially the same as described by Zheng et
al (7). The only difference
between the studies is that fish patients in the study by Zheng
et al did not consume a diet rich in fish. Disease-free
third molars and premolars were collected during orthodontic
treatment from 12 patients (8 male; 4 female), and diseased teeth
were collected during tooth extraction from 12 patients (7 males; 5
females) with generalized chronic periodontitis. All patients
included in the current study were 30–40 years of age. The present
study was approved by the Ethics Committee of The Stomatological
Hospital of Southern Medical University. All participants were
informed of the experimental protocol and signed informed
consent.
Cell culture and transfection
PDL tissues were obtained from the central root of
healthy and periodontitis-affected teeth. PDL tissues were cut into
pieces (1 mm3) and cultured in a six-well plate filled
with α-MEM (Sigma-Aldrich; Merck KGaA) containing 100 µM/l ascorbic
acid, 0.292 mg/ml glutamine, 10% FBS (Sigma-Aldrich; Merck KGaA)
and 100 U/ml penicillin/streptomycin. Cells were subsequently
cultured in an incubator at 37°C and 5% CO2.
To overexpress MALAT1, full-length MALAT1 cDNA was
digested using NheI restriction enzymes and amplified using
PCR and inserted into a NheI-linearized pEGFPC3 vector
(Clontech Laboratories, Inc.) to generate a MALAT1 expression
vector. Primer sequences were as follows: Forward,
5′-CGCAGCCTGCAGCCCGAGAC-3′ and reverse,
5′-CCCAATCAAGATTTTTTTATTC-3′. PCRs were carried out using Pfu DNA
Polymerase kit (Promega Corporation) with the following
thermocycling conditions: 95°C for 5 min, followed by 35 cycles of
95°C for 10 sec, 55°C for 10 sec and 72°C for 90 sec. To silence
MALAT1 expression, MALAT1 small interfering RNA (siRNA,
5′-CCACUUACUGCUUUAAGUUGG-3′) or negative control siRNA
(5′-UUCUCCGAACGUGUCACGUdTdT-3′) were designed and synthesized by
Sangon Biotech Co., Ltd. A total of 10 nM vectors or 50 nM siRNAs
were transfected into 4×105 cells using Lipofectamine
2000® reagent (cat. no. 11668-019; Invitrogen; Thermo
Fisher Scientific, Inc.). Control cells were untransfected and
negative control cells were transfected with empty vectors or
negative control siRNA. Subsequent experiments were performed at 24
h after transfection.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from PDLSCs using TRIzol
reagent (Thermo Fisher Scientific, Inc.). cDNA was synthesized
using a SuperScript III Reverse Transcriptase kit (Thermo Fisher
Scientific, Inc.) according to the following thermocycling
conditions: 25°C for 5 min, 51°C for 20 min and 75°C for 15 min.
SYBR® Green Real-Time PCR Master Mixes (Thermo Fisher
Scientific, Inc.) was used to prepare all PCR reactions. Reaction
conditions were as follows: 95°C for 50 sec, followed by 40 cycles
of 95°C for 15 sec and 58°C for 20 sec. The following primers were
used: MALAT1 forward, 5′-CAGTGGGGAACTCTGACTCG-3′, and reverse,
5′-GTGCCTGGTGCTCTCTTACC-3′; FGF2 forward,
5′-GCACUGAAACGAACUGGGCAGUAU-3′, and reverse,
5′-UAUACUGCCCAGUUCGUUUCAGUGC-3′; β-actin forward,
5′-GACCTCTATGCCAACACAGT-3′, and reverse,
5′-AGTACTTGCGCTCAGGAGGA-3′. This experiment was performed in
triplicate and MALAT1 expression was normalized to the β-actin
endogenous control using the 2−∆∆Cq method (12).
Cell proliferation assay
Following confirmation of MALAT1 overexpression in
PDLSCs compared with the controls, PDLSCs were subsequently
collected and single cell suspensions were prepared with a final
cell density of 3×104 cells/ml. Each well of a 96-well
plate was filled with 3×103 cells in 0.1 ml cell
suspension. Cells were cultured in an incubator at 37°C and 5%
CO2 for 24, 48, 72 and 96 h, before 10 µl Cell Counting
kit-8 (CCK-8) solution was added at the end of each time point.
Cells were cultured at 37°C for an additional 4 h and a
Fisherbrand™ accuSkan™ GO UV/Vis Microplate Spectrophotometer
(Thermo Fisher Scientific, Inc.) was used to measure optical
density (OD) values at 450 nm. Cell proliferation was normalized to
the sample with the highest OD value at 96 h, which was set to
100%.
Western blot analysis
Subsequent to transfection and RT-qPCR analysis,
PDLSCs were collected and mixed with RIPA buffer (Thermo Fisher
Scientific, Inc.) to extract total protein, and protein
concentrations were measured using a BCA kit (Thermo Fisher
Scientific, Inc.). Protein samples (30 µg/lane) were denatured and
separated by 10% SDS-PAGE. Following gel transfer, PVDF membranes
were then blocked with 5% skimmed milk at room temperature for 2 h.
Samples were then incubated with rabbit anti-human primary
antibodies against FGF2 (1:1,300; cat. no. ab8880; Abcam) and GAPDH
(1:1,300; cat. no. ab8245; Abcam) at 4°C overnight. Following that,
membranes were further incubated with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibodies
(1:1,300; cat. no. ab6721; Abcam) at 24°C for 2 h. Signal
development was performed using ECL™ Detection Reagents GE
Healthcare (Sigma-Aldrich; Merck KGaA). Signals were detected using
a MYECL™ Imager (Thermo Fisher Scientific, Inc.). ImageJ software
(version 1.34; National Institutes of Health) was used to normalize
FGF2 expression to the GAPDH endogenous control.
Statistical analysis
GraphPad Prism 6 software (GraphPad Software, Inc.)
was used for statistical analysis. Data are expressed as the mean ±
standard deviation. A Student's t-test was used to perform
statistical comparisons between two groups, and one-way ANOVA
followed by a Fisher's least significant difference test was used
to perform comparisons among multiple groups. Pearson's correlation
coefficient was used for correlational analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
MALAT1 and FGF2 mRNA are upregulated
in PDLSCs derived from periodontitis-affected teeth
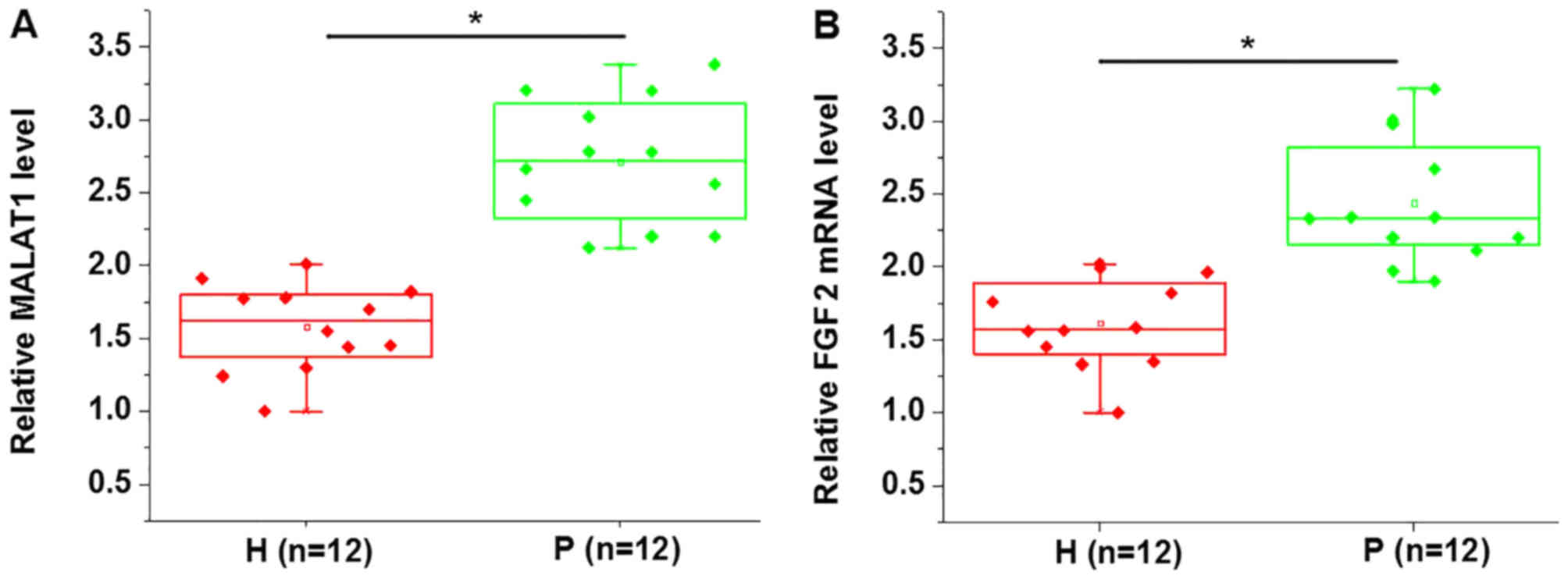
MALAT1 and FGF2 mRNA expression in PDLSCs derived
from periodontitis-affected and healthy teeth was detected using
RT-qPCR analysis. As presented in Fig.
1, the expression of MALAT1 (Fig.
1A) and FGF2 mRNA (Fig. 1B) was
significantly upregulated in PDLSCs derived from
periodontitis-affected teeth when compared with PDLSCs derived from
healthy teeth.
MALAT1 and FGF2 mRNA expression are
positively correlated in PDLSCs derived from healthy and
periodontitis-affected patients
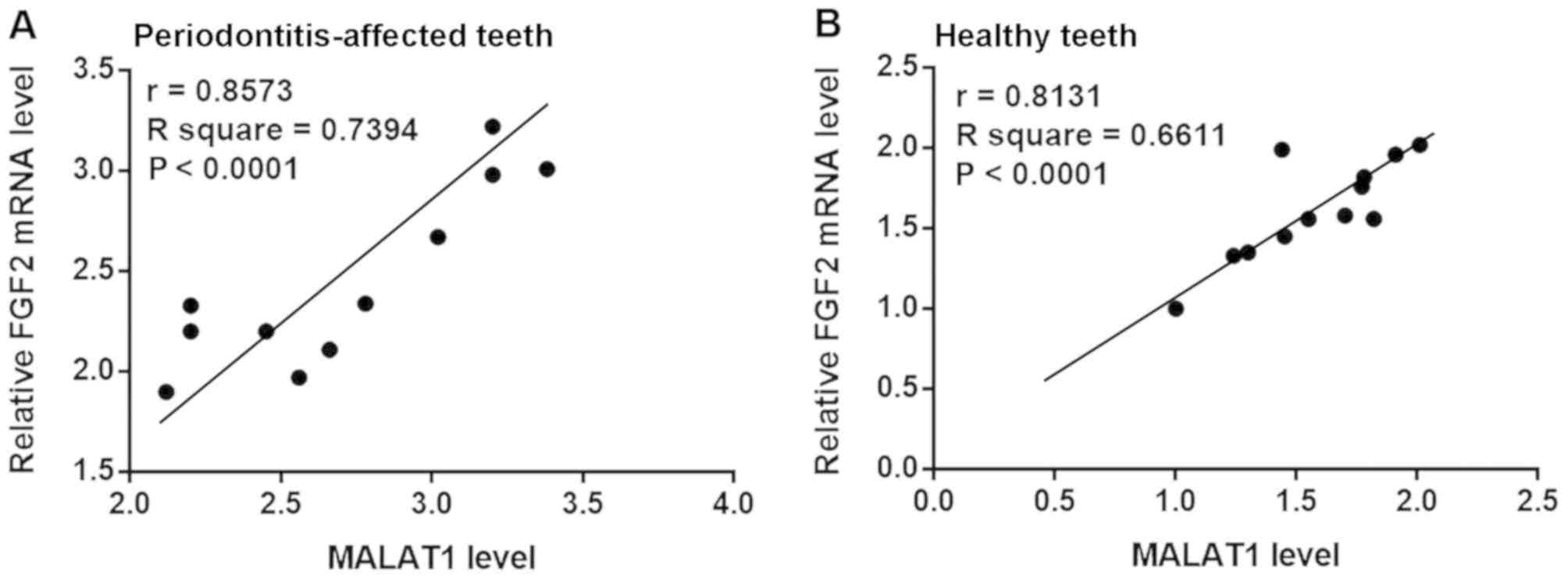
Pearson's correlation coefficient analysis of MALAT1
and FGF2 mRNA expression in PDLSCs derived from
periodontitis-affected and healthy teeth was performed. As
presented in Fig. 2, a significant
positive correlation between the expression of MALAT1 and FGF2 mRNA
in PDLSCs derived from periodontitis-affected teeth (Fig. 2A) and healthy teeth (Fig. 2B) was observed.
PDLSCs derived from
periodontitis-affected teeth reveal a higher cell proliferation
rate than PDLSCs derived from healthy teeth
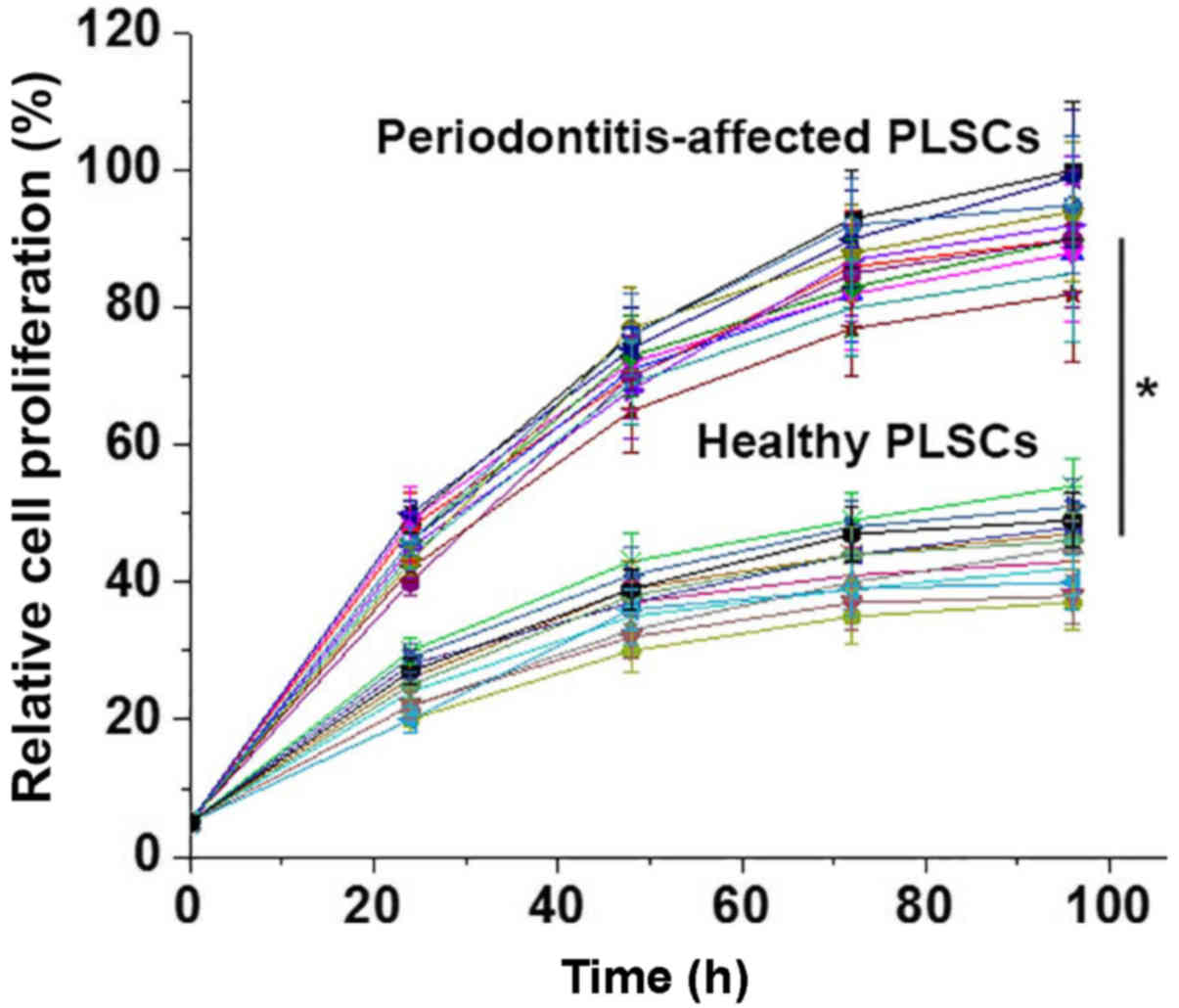
A CCK-8 kit was used to detect the proliferation of
PDLSCs derived from periodontitis-affected teeth and PDLSCs derived
from healthy teeth. As presented in Fig.
3, the proliferation rates of the 12 PDLSCs samples derived
from periodontitis-affected teeth were significantly higher than
those of the 12 cases of PDLSCs derived from healthy teeth.
MALAT1 promotes the proliferation of
PDLSCs derived from periodontitis-affected and healthy teeth
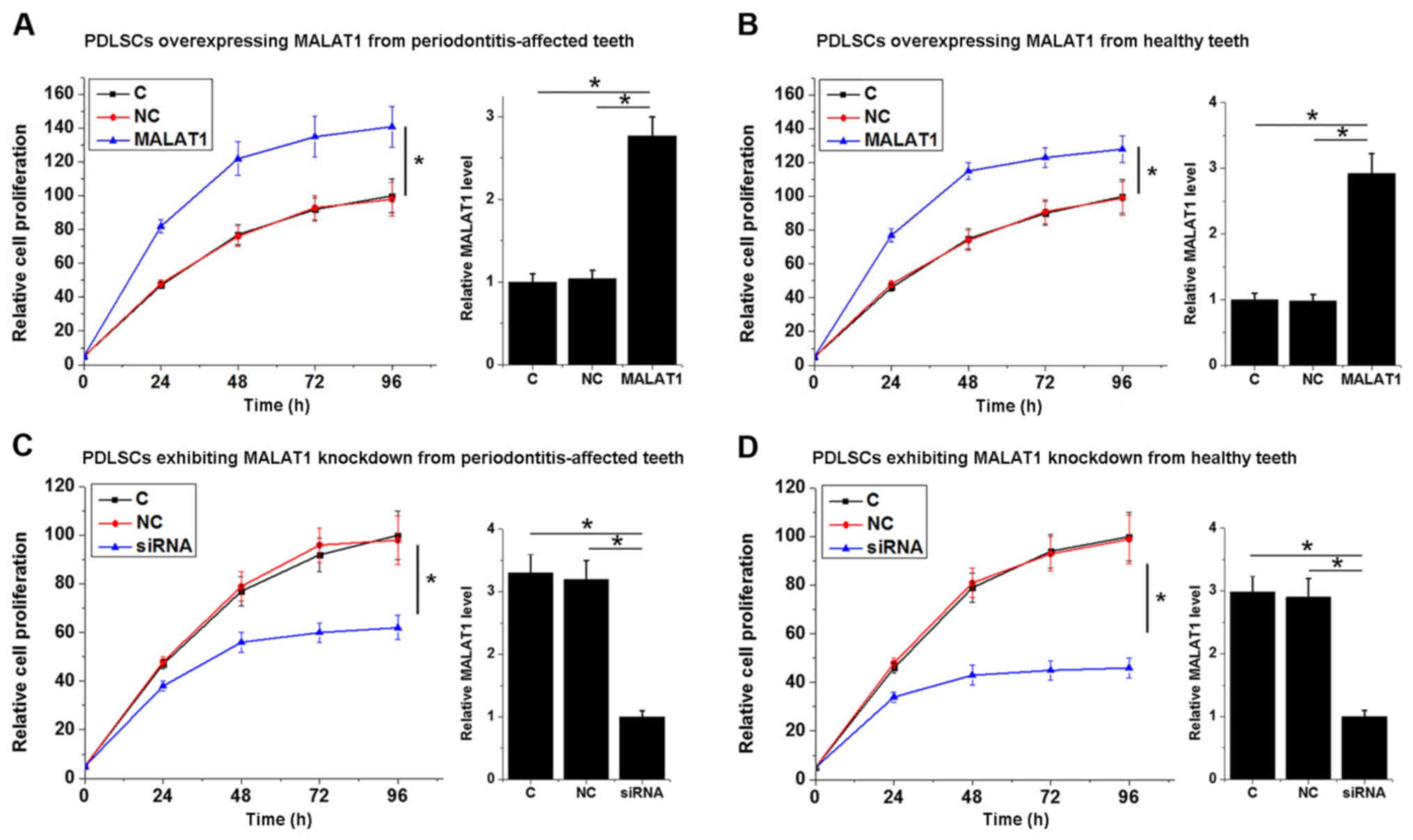
To investigate the involvement of MALAT1 in the
proliferation of PDLSCs, a MALAT1 expression vector was transfected
into each of the 12 PDLSC samples derived from
periodontitis-affected teeth and the PDLSCs derived from 12 healthy
teeth samples. The results shown in Fig.
4 are representative data. Cell proliferation was detected
using a CCK-8 assay. As presented in Fig. 4, MALAT1 overexpression significantly
promoted the proliferation of PDLSCs derived from
periodontitis-affected teeth (P<0.05; Fig. 4A) and healthy teeth (P<0.05;
Fig. 4B). By contrast, MALAT1
knockdown significantly inhibited the proliferation of PDLSCs
derived from periodontitis-affected teeth (P<0.05; Fig. 4C) and healthy teeth (P<0.05;
Fig. 4D). Transfections were
confirmed by qPCR and data were presented on the right part of each
section. The data revealed that the expression level of MALAT1 was
significantly upregulated after the transfection of expression
vectors and was significantly downregulated after the transfection
of siRNAs (P<0.05; Fig. 4).
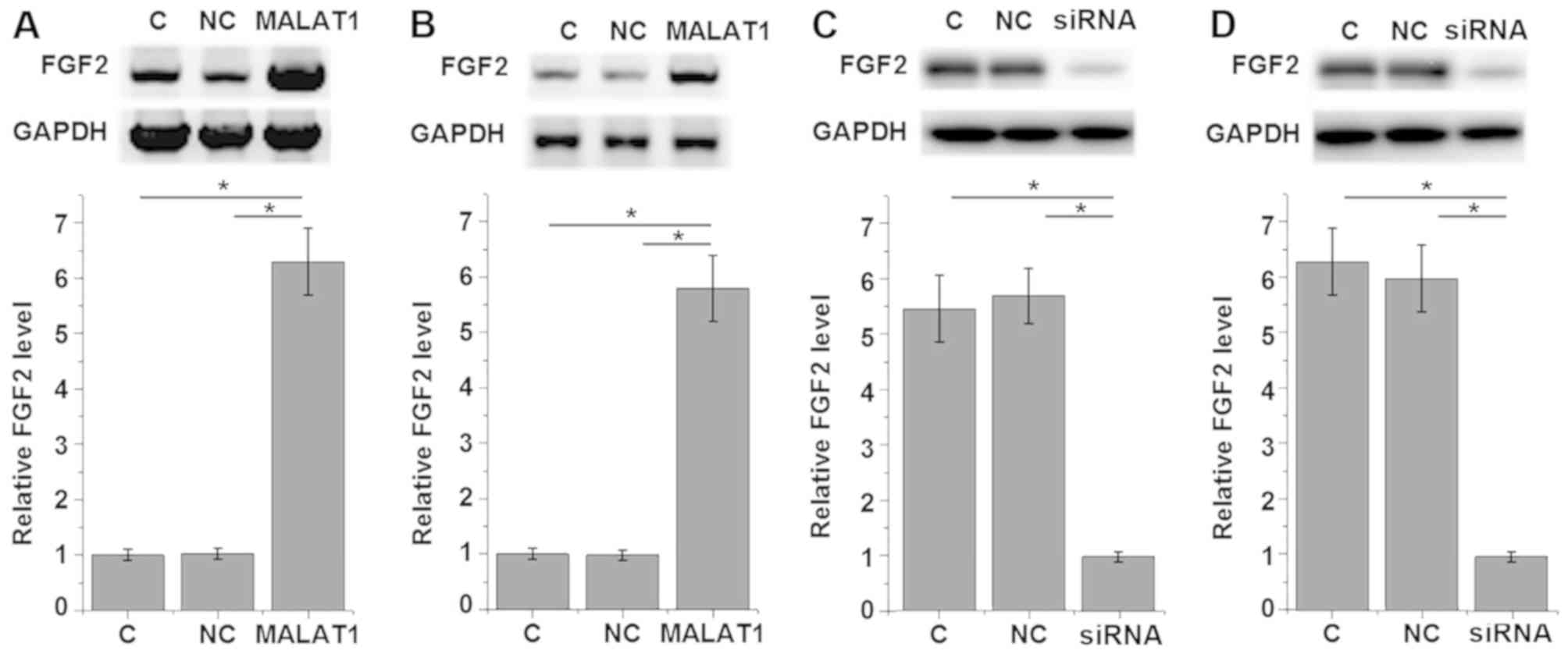
MALAT1 upregulates FGF2 protein
expression in PDLSCs derived from healthy individuals and patients
with periodontitis
To further investigate the interaction between
MALAT1 and FGF2 in PDLSCs, a MALAT1 expression vector was
transfected into each of the 12 PDLSCs samples derived from
periodontitis-affected teeth and the PDLSCs derived from 12 healthy
teeth samples. The results shown in Fig.
5 are representative data. FGF2 expression was detected using
western blot analysis. As presented in Fig. 5, MALAT1 overexpression significantly
upregulated FGF2 expression in PDLSCs derived from
periodontitis-affected teeth (P<0.05; Fig. 5A) and healthy teeth (P<0.05;
Fig. 5B) when compared with the
control and negative control group. By contrast, MALAT1 knockdown
significantly downregulated FGF2 expression in PDLSCs derived from
periodontitis-affected teeth (P<0.05; Fig. 5C) and healthy teeth (P<0.05;
Fig. 5D) when compared with their
respective control and negative control groups.
Discussion
The present study demonstrated that lncRNAs serve
pivotal roles in cancer biology and lncRNA MALAT1 was revealed to
be associated with the pathogenesis of periodontitis. The results
also revealed that lncRNA MALAT1 may promote the development of
periodontitis by upregulating the expression of FGF2 and promoting
the proliferation of PDLSCs.
The onset and development of periodontitis is
accompanied by changes in the expression of a large variety of
lncRNAs, indicating their association with the pathogenesis of this
disease (13). However, studies
assessing the expression and functionality of specific lncRNAs in
periodontitis are required. In a recent study, Wang et al
(14) demonstrated that lncRNA-POIR
functions as a positive regulator of osteogenic differentiation,
which is impaired in periodontitis, indicating a protective role of
lncRNA-POIR in periodontitis (14).
In an additional study, the ANRIL lncRNA was demonstrated to
participate in the pathogenesis of periodontitis via interacting
with multiple signaling pathways (15). MALAT1 is a well-characterized
oncogenic lncRNA, which exhibits upregulated expression in a
variety of human cancer types (10,11). In
the present study, MALAT1 expression was revealed to be
significantly upregulated in PDLSCs derived from
periodontitis-affected teeth, suggesting an involvement of MALAT1
in periodontitis.
FGF2 expression stimulates periodontal regeneration,
indicating its potential clinical importance as a therapeutic
target in novel periodontitis treatments (16,17). The
current study observed a significant upregulation of FGF2 mRNA
expression in PDLSCs derived from periodontitis-affected teeth when
compared with PDLSCs derived from healthy teeth. This result
indicates the potential involvement of FGF2 in the pathogenesis of
periodontitis. FGF2 interacts with lncRNAs through certain
mediators (such as miRNAs) to achieve its biological functions,
such as the regulation of vascular endothelial cell apoptosis
(18). In a previous study, the
MALAT1 lncRNA was demonstrated to be positively correlated with
FGF2 protein expression and secretion in thyroid cancer (9). Similarly, MALAT1 and FGF2 mRNA
expression was revealed to be positively correlated in PDLSCs
derived from both periodontitis-affected and healthy teeth in the
present study. In addition, MALAT1 overexpression was associated
with upregulation of FGF2 protein in PDLSCs derived from
periodontitis-affected and healthy teeth. These results suggest
that MALAT1 is a positive regulator of FGF2 in periodontitis, and
the regulatory effects of MALAT1 on FGF2 are unlikely to be
mediated by periodontitis-specific factors.
PDLSCs possess immunomodulatory properties through
the secretion of a variety of cytokines, including interleukin
(IL)-6, IL-8 and monocyte chemotactic protein 1 (19). It has been previously reported that
abnormally accelerated PDLSC proliferation is associated with the
pathogenesis of periodontitis (8).
In the current study, a significantly higher proliferation rate was
observed in all 12 PDLSC samples derived from
periodontitis-affected teeth when compared with the 12 PDLSC
samples derived from healthy teeth. MALAT1 was also revealed to be
a likely activator of PDLSCs based on the observation that MALAT1
overexpression led to the significantly increased proliferation of
PDLSCs derived from both periodontitis-affected and healthy teeth.
However, the molecular mechanisms underlying this effect is
currently unknown and future studies should aim to elucidate this
mechanism. In addition, the present study is limited by a small
sample size. Therefore, future studies that include a larger sample
size are required to validate the results of the current study. In
conclusion, MALAT1 and FGF2 are upregulated in periodontitis. In
addition, MALAT1 may serve a role in periodontitis by promoting
FGF2 expression and the proliferation of PDLSCs.
Acknowledgements
Not applicable.
Funding
The current study received financial support from
Natural Science Foundation of Tibet Autonomous Region (grant no.
XZ2017ZR-ZYZ37) and The Scientific Research Project of Southern
Medical University Stomatological Hospital (grant no.
PY2018014)
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PC and MR designed the experiments. PC, YH and YW
performed the experiments. SL and HC collected and analyzed the
data. MR drafted manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The Stomatological Hospital of Southern Medical
University.
Patient consent for publication
Patients provide consent for the possible
publication of this paper.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hajishengallis G: Periodontitis: From
microbial immune subversion to systemic. Inflammation. Nat Rev
Immunol. 15:30–44. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lundberg K, Wegner N, Yucel-Lindberg T and
Venables PJ: Periodontitis in RA-the citrullinated enolase
connection. Nat Rev Rheumatol. 6:727–730. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kebschull M, Demmer RT and Papapanou PN:
‘Gum bug, leave my heart alone!’-epidemiologic and mechanistic
evidence linking periodontal infections and atherosclerosis. J Dent
Res. 89:879–902. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Madianos PN, Bobetsis YA and Offenbacher
S: Adverse pregnancy outcomes (APOs) and periodontal disease:
pathogenic mechanisms. J Periodontol. 84 (Suppl):S170–S180. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhang Q, Li Z, Wang C, Shen T, Yang Y,
Chotivichien S and Wang L: Prevalence and predictors for
periodontitis among adults in China, 2010. Glob Health Action.
7:245032014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cionca N: Use and misuse of systemic
antibiotics in periodontitis treatment. Oral Health Prev Dent.
15:305–306. 2017.PubMed/NCBI
|
|
7
|
Zheng W, Wang S, Wang J and Jin F:
Periodontitis promotes the proliferation and suppresses the
differentiation potential of human periodontal ligament stem cells.
Int J Mol Med. 36:915–922. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee JH, Um S, Jang JH and Seo BM: Effects
of VEGF and FGF-2 on proliferation and differentiation of human
periodontal ligament stem cells. Cell Tissue Res. 348:475–484.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang JK, Ma L, Song WH, Lu BY, Huang YB,
Dong HM, Ma XK, Zhu ZZ and Zhou R: LncRNA-MALAT1 promotes
angiogenesis of thyroid cancer by modulating tumor-associated
macrophage FGF2 protein secretion. J Cell Biochem. 118:4821–4830.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu JH, Chen G, Dang YW, Li CJ and Luo DZ:
Expression and prognostic significance of lncRNA MALAT1 in
pancreatic cancer tissues. Asian Pac J Cancer Prev. 15:2971–2977.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tian X and Xu G: Clinical value of lncRNA
MALAT1 as a prognostic marker in human cancer: Systematic review
and meta-analysis. BMJ Open. 5:e0086532015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zou Y, Li C, Shu F, Tian Z, Xu W, Xu H,
Tian H, Shi R and Mao X: lncRNA expression signatures in
periodontitis revealed by microarray: The potential role of lncRNAs
in periodontitis pathogenesis. J Cell Biochem. 116:640–647. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang L, Wu F, Song Y, Li X, Wu Q, Duan Y
and Jin Z: Long noncoding RNA related to periodontitis interacts
with miR-182 to upregulate osteogenic differentiation in
periodontal mesenchymal stem cells of periodontitis patients. Cell
Tissue Res. 7:e23272016.
|
|
15
|
Bochenek G, Häsler R, El Mokhtari NE,
König IR, Loos BG, Jepsen S, Rosenstiel P, Schreiber S and Schaefer
AS: The large non-coding RNA ANRIL, which is associated with
atherosclerosis, periodontitis and several forms of cancer,
regulates ADIPOR1, VAMP3 and C11ORF10. Hum Mol Genet. 22:4516–4527.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kitamura M, Akamatsu M, Machigashira M,
Hara Y, Sakagami R, Hirofuji T, Hamachi T, Maeda K, Yokota M, Kido
J, et al: FGF-2 stimulates periodontal regeneration: results of a
multi-center randomized clinical trial. J Dent Res. 90:35–40. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Murakami S: Periodontal tissue
regeneration by signaling molecule(s): What role does basic
fibroblast growth factor (FGF-2) have in periodontal therapy?
Periodontol. 56:188–208. 2000. View Article : Google Scholar
|
|
18
|
Lu W, Huang SY, Su L, Zhao BX and Miao JY:
Long noncoding RNA LOC100129973 suppresses apoptosis by targeting
miR-4707-5p and miR-4767 in vascular endothelial cells. Sci Rep.
6:216202016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Andrukhov O, Hong JS, Andrukhova O,
Blufstein A, Moritz A and Rausch-Fan X: Response of human
periodontal ligament stem cells to IFN-γ and TLR-agonists. Sci Rep.
7:128562017. View Article : Google Scholar : PubMed/NCBI
|