Introduction
Type 2 diabetes mellitus (T2D), commonly recognized
as a collection of prolonged metabolic disorders, is a public
health issue with increasing prevalence worldwide (1). Ectopic lipid deposition in skeletal
muscle and the liver may lead to insulin resistance and diabetes
(2,3). Adipose tissue is the largest organ used
in humans for lipid storage and mobilization based on energy
requirements: It contains several types of cells, including mature
adipocytes. As an endocrine organ, adipose tissue contributes to
the complex regulatory homeostasis of energy intake, and to the
metabolism of glucose and lipids (4). An increasing number of studies have
identified that adipose tissue dysfunction may cause metabolic
syndrome, such as T2D, atherosclerosis, cardiovascular disease and
even cancer (5,6). Consequently, adipocytes have emerged as
a possible pharmacological target in T2D (7).
Adipocyte differentiation is complex with the
process orchestrated by a cascade of transcription factors and
other regulatory proteins. Nuclear receptor peroxisome
proliferator-activated receptor-γ (PPARγ) and CCAAT/enhancer
binding protein-α (C/EBPα) are activated during adipocyte
differentiation. PPARγ acts together with retinoid X receptor to
induce differentiation by hormonal stimulation (8,9). In the
early stages of adipocyte differentiation, C/EBPβ and C/EBPδ are
both expressed earlier than C/EBPα. C/EBPβ and C/EBPδ induce C/EBPα
and PPARγ expression, and then together with PPARγ and C/EBPα
stimulate the expression of the downstream adipose-specific genes
involved in glucose uptake, adipose phenotype, and lipid
accumulation (10,11).
Signaling pathways such as PI3K/Akt, Wnt/β-catenin
and mitogen-activated protein kinase signaling mediate the
adipogenic transcriptional cascade involved in adipogenesis
(12). PI3K/Akt signaling is
important in PPARγ and C/EBPα upregulation and adipogenesis
(13). Fibroblasts lacking Akt
activation, or when Akt activation is inhibited, cannot
differentiate from preadipocytes into adipocytes (14). By contrast, Akt activation can
promote embryonic fibroblast differentiation into mature adipocytes
even if other adipogenic factors are absent (15).
Recently, dietary phytochemicals that have
beneficial effects on obesity and T2D by regulating adipocyte
differentiation have become a highly attractive topic, as natural
products have lower risks of adverse effects compared to synthetic
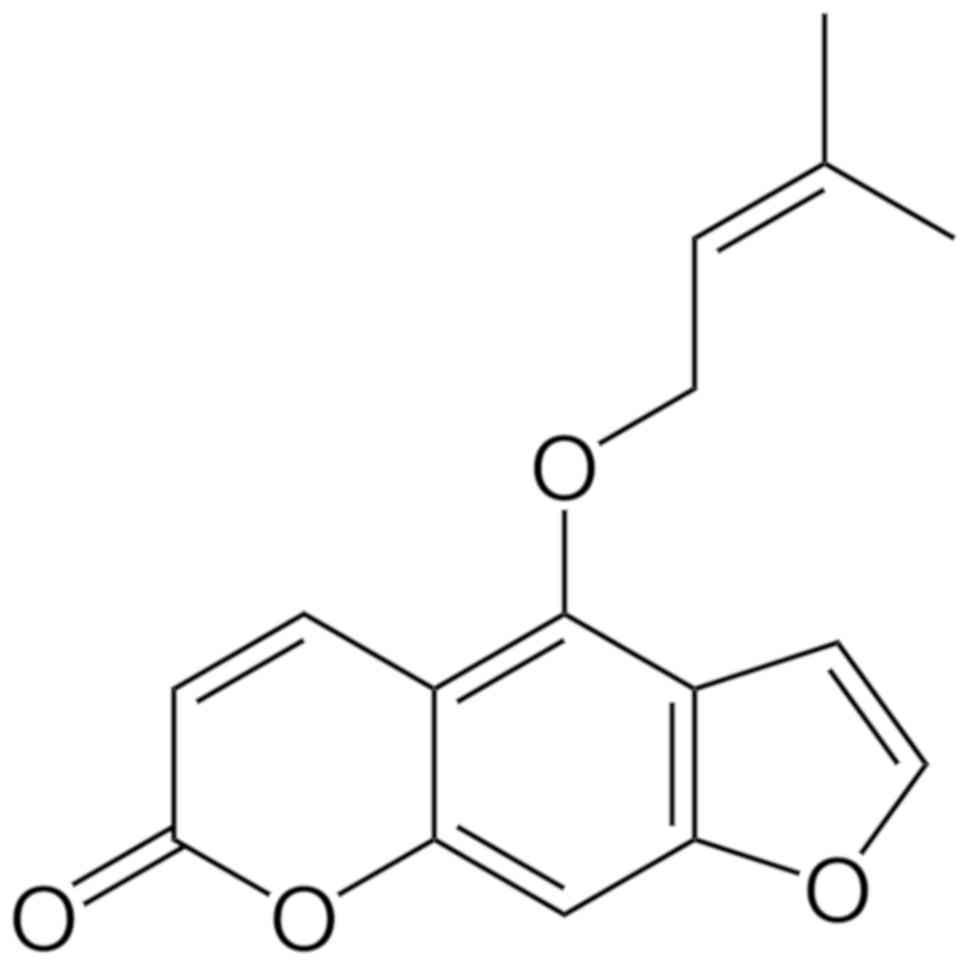
drugs (16). Isoimperatorin (ISOIM;
Fig. 1) is included in the
6,7-furanocoumarin family of compounds and is the main effective
element in the Umbelliferae family (17). Heracleum, Angelica dahurica,
Chinese angelica, coastal glehnia root and Peucedanum
ostruthium are members of the Umbelliferae family, which are
all widely used as traditional medicines in many countries
(18). ISOIM displays
anti-inflammatory (19),
anti-hypertension (20), analgesic
(19), anti-cancer (21), and hepatoprotective properties
(22). Additionally, dietary
furocoumarin imperatorin, an ISOIM isomer, increases glucagon-like
peptide secretion, reducing blood sugar in rodents by activating G
protein-coupled bile acid receptor 1 (23). In addition, lipodystrophic patients
have an adipose tissue triglyceride storage defect that causes
ectopic lipid accumulation, leading to the development of severe
insulin resistance (2,3). Furthermore, increased fat capacity
storage in adipose with low fat mobilization leads to the expansion
of fat mass, and may also be considered the best means of storing
lipids in a harmless compartment (3).
The present study investigated the underlying
molecular mechanism by which ISOIM regulates the differentiation of
3T3-L1 adipocytes and the accumulation of lipids. These finding may
contribute to the development of novel drugs that ameliorate
diabetes.
Materials and methods
Materials
ISOIM (≥98% purity;
C16H14O4) was obtained from Wuhan
Jonk Biological Technology Co., Ltd. and maintained in 100 mM stock
solutions in dimethyl sulfoxide then stored at −20°C.
Cell culture and differentiation
3T3-L1 fibroblasts were purchased from the Stem Cell
Bank, Chinese Academy of Sciences and were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) containing 10% fetal bovine serum (FBS; Zeta Life, Inc.) and
1% streptomycin/penicillin under a humidified 5% CO2
incubator at 37°C. Two days after reaching confluence, the culture
medium was switched to differentiation medium: DMEM containing 10%
FBS and 0.5 mM isobutylmethylxanthine, 1 µM dexamethasone and 10
µg/ml insulin (all Sigma-Aldrich; Merck KGaA) for 2 days at 37°C.
Then, cells were maintained in differentiation medium II: DMEM
containing 10% FBS and 10 µg/ml insulin. The medium was replaced
every 2 days. To observe the impact of ISOIM on adipogenesis, the
cells were treated with 20 µM ISOIM, which was included in each
medium used, from day 0 to 6 during differentiation.
Cell Counting Kit-8 (CCK-8)
Cell viability was determined using CCK-8 kit
(Vazyme) according to the manufacturer's protocol. Cells
(1×104 per well) were seeded in a 96-well plate and
incubated overnight at 37°C, before being treated with 0–60 µM
ISOIM for 48 h at 37°C. Then, 10 µl CCK-8 solution was added to
each well and the plate was incubated at 37°C for 2 h. A microplate
reader (Thermo Fisher Scientific, Inc.) was used to measure the
absorbance at 450 nm.
Oil red O staining
Following the six-day induction of 3T3-L1 fibroblast
differentiation, the accumulation of lipids was visualized using
lipid-specific oil red O staining (Sigma-Aldrich; Merck KGaA). The
cells were washed twice with PBS then were fixed for 30 min at room
temperature in 4% formalin. The cells were washed with 60%
isopropanol and incubated with oil red O working solution (0.5 g
oil red O in 100 ml isopropanol diluted with water in a 3:2 ratio
then filtered) at room temperature for 30 min. Samples were washed
twice in water then imaged with and inverted microscope (Primovert;
Carl Zeiss Microscopy, LLC) at a magnification of ×200. Oil red O
stain was extracted from the cells using 60% isopropanol and the
absorbance measured at 490 nm using a microplate reader (Multiskan
FC; Thermo Fisher Scientific, Inc.).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from cells using RNAiso Plus
Reagent (Takara Bio, Inc.). A total of 500 ng total RNA was reverse
transcribed to complementary DNA using a PrimeScript RT Reagent Kit
(Takara Bio, Inc.) with the following thermocycling parameters:
37°C for 15 min and 85°C for 5 sec. A SYBR Green kit (Takara Bio,
Inc.) was used for qPCR, with measurement using an iQ5 real-time
qPCR detection system (Bio-Rad Laboratories, Inc.). The
thermocycling conditions were as follows: Initial denaturation at
94°C for 5 min; 40 cycles of 94°C for 10 sec, 60°C for 30 sec, 72°C
for 30 sec; and a final elongation at 72°C for 10 min. The internal
control was β-actin. mRNA levels were quantified using the
comparative threshold cycle 2−ΔΔCq method (24). The primer sequences were as follows:
C/EBPβ forward, 5′-AAGCTGAGCGACGAGTACAAGA-3′ and reverse,
5′-GTCAGCTCCAGCACCTTGTG-3′; PPARγ forward,
5′-CCAAGAATACCAAAGTGCGATCA-3′ and reverse,
5′-CCCACAGACTCGGCACTCAAT-3′; C/EBPα forward,
5′-CTGATTCTTGCCAAACTGAG-3′ and reverse,
5′-GAGGAAGCTAAGACCCACTAC-3′; sterol regulatory element-binding
transcription factor 1c (SREBP1c) forward,
5′-GGAGCCATGGATTGCACATT-3′ and reverse, 5′-GGCCCGGGAAGTCACTGT-3′;
adipocyte protein 2 (aP2) forward, 5′-AAGAAGTGGGAGTGGGCTTTG-3′ and
reverse, 5′-CTCTTCACCTTCCTGTCGTCTG-3′; fatty acid synthase (FAS)
forward, 5′-ATCAAGGAGGCCCATTTTGC-3′ and reverse,
5′-TGTTTCCACTTCTAAACCATGCT-3′; diacylglycerol
O-acyltransferase 2 (DGAT2) forward,
5′-CCTTCCTGGTGCTAGGAGTG-3′ and reverse, 5′-CCAGTCAAATGCCAGCCA-3′;
adiponectin forward, 5′-AAAGGGCTCAGGATGCTACTG-3′ and reverse,
5′-TGGGCAGGATTAAGAGGAACA-3′; adipose triglyceride lipase (ATGL)
forward, 5′-TTCGCAATCTCTACCGCCTC-3′ and reverse,
5′-AAAGGGTTGGGTTGGTTCAG-3′; hormone-sensitive lipase (HSL) forward,
5′-GCTGGGCTGTCAAGCACTGT-3′ and reverse, 5′-GTAACTGGGTAGGCTGCCAT-3′;
and β-actin forward, 5′-GTCCCTGACCCTCCCAAAAG-3′ and reverse,
5′-GCTGCCTCAACACCTCAACCC-3′.
Western blot analysis
Cells were lysed in radioimmunoprecipitation lysis
buffer (Appligen) containing protease inhibitor cocktail (Beijing
ComWin Biotech Co., Ltd.). Protein concentrations were determined
using a bicinchoninic acid protein assay kit (Beijing ComWin
Biotech Co., Ltd.). Proteins (30 µg/lane) were separated via
SDS-PAGE on a 10% gel, and transferred to polyvinylidene difluoride
membranes (EMD Millipore; Merck KGaA). Membranes were blocked for
1.5 h at room temperature using 5% non-fat milk in Tris-buffered
saline containing 0.1% Tween. Membranes were then incubated with
primary antibodies against PPARγ (cat. no. 2430), ATGL (cat. no.
2138), HSL (cat. no. 4107), Akt (cat. no. 9272), phosphorylated
Akt-pS473 (cat. no. 9271) and β-actin (cat. no. 4970; all Cell
Signaling Technology, Inc.) and C/EBPα (cat. no. ab40764; all
1:1,000; Abcam) at 4°C overnight. Following washing, the membranes
were incubated with horseradish peroxidase-conjugated anti-rabbit
immunoglobulin G secondary antibodies (cat. no. 7074; 1:3,000; Cell
Signaling Technology, Inc.) for 1.5 h at room temperature. An
enhanced chemiluminescent peroxidase substrate (EMD Millipore;
Merck KGaA) was used to visualize the protein bands. ImageJ
software (1.8.0_112 version; National Institutes of Health) was
used to quantify band density. The expression level of the protein
was calculated using the ratio of the target protein intensity to
β-actin.
Statistical analysis
Each experiment was performed three times
independently unless stated otherwise. Statistical analysis was
performed using SPSS v18.0 (SPSS, Inc.) and GraphPad Prism 6
(GraphPad Software, Inc.). Quantitative data were analyzed using
Student's t-test and one-way analysis of variance followed by
Tukey's post hoc test for multiple group comparisons. Data were
presented as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Cytotoxic effects of ISOIM on 3T3-L1
preadipocytes
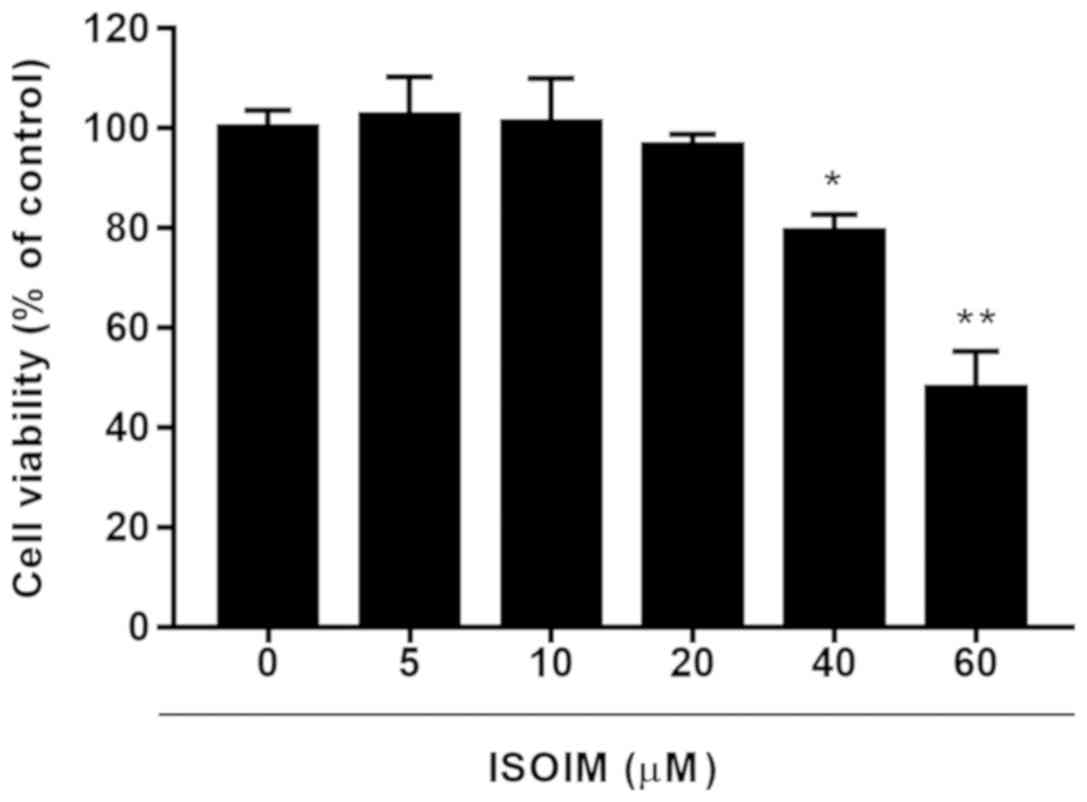
To select a subtoxic ISOIM dose for later
adipogenesis study, ISOIM cytotoxicity to 3T3-L1 fibroblasts was
evaluated. ISOIM inhibited cell viability in a dose-dependent
manner. At 40 and 60 µM, ISOIM significantly decreased cell
viability to 79.2±3.50 and 47.7±7.56% of the control, respectively
(Fig. 2). To avoid obvious ISOIM
cytotoxicity, <40 µM ISOIM was selected for subsequent
experiments to detect the impact on lipid accumulation.
ISOIM enhances 3T3-L1 cell lipid
accumulation
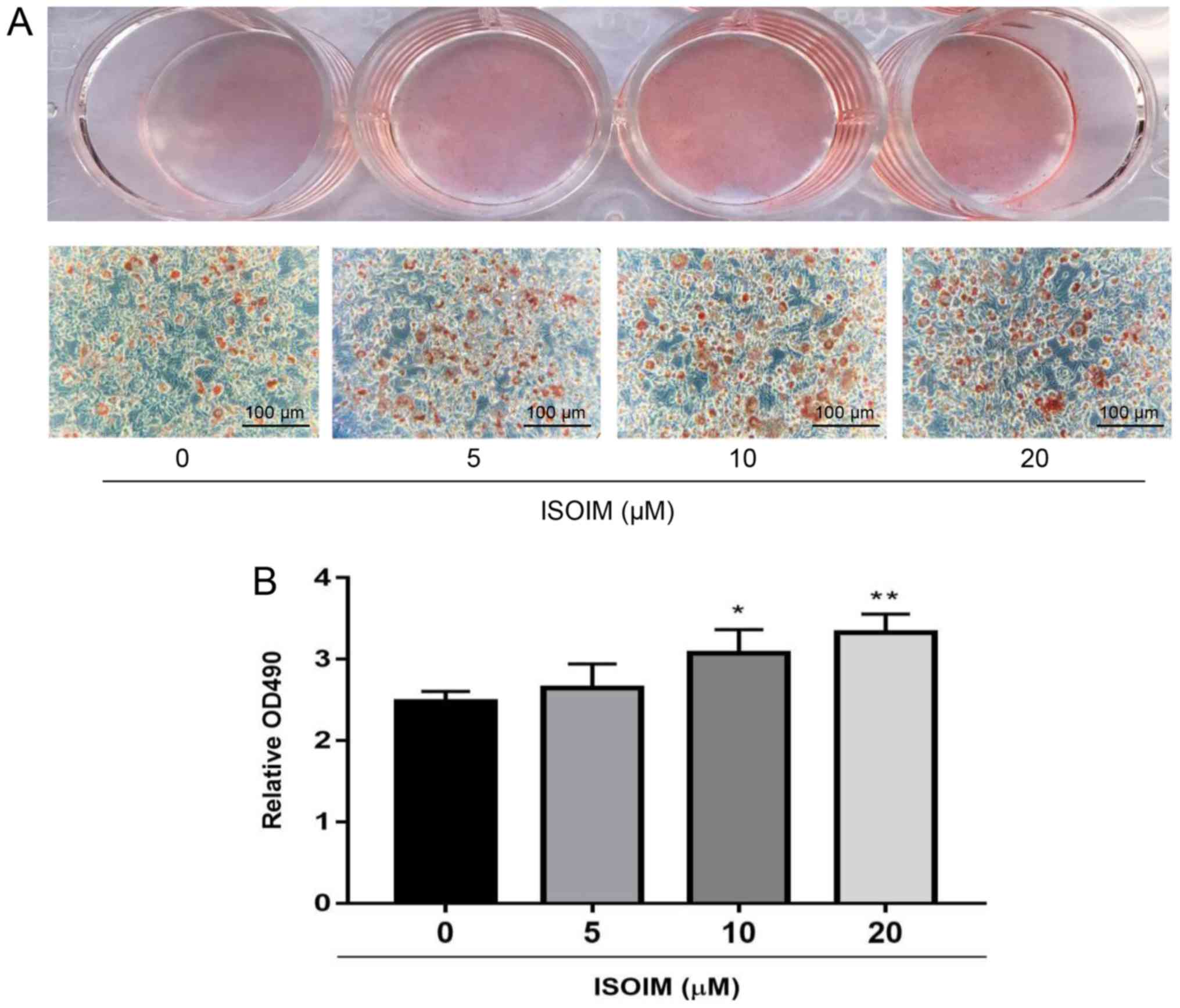
To investigate the possible effect of ISOIM on
adipogenesis, 3T3-L1 fibroblasts were treated with 0, 5, 10 or 20
µM ISOIM for 6 days in the differentiation mediums. ISOIM promoted
3T3-L1 adipocyte differentiation (Fig.
3A). The cells were treated with isopropanol to release the oil
red O stain into the solution, which represented the cytoplasmic
accumulation of lipid droplets, with 10 and 20 µM ISOIM absorbance
significantly increased to 24.0 and 34.3% relative to the control.
However, the 5 µM group and the control were not significantly
different (Fig. 3B).
ISOIM increases adipogenesis-related
mRNA and protein expression in differentiated adipocytes
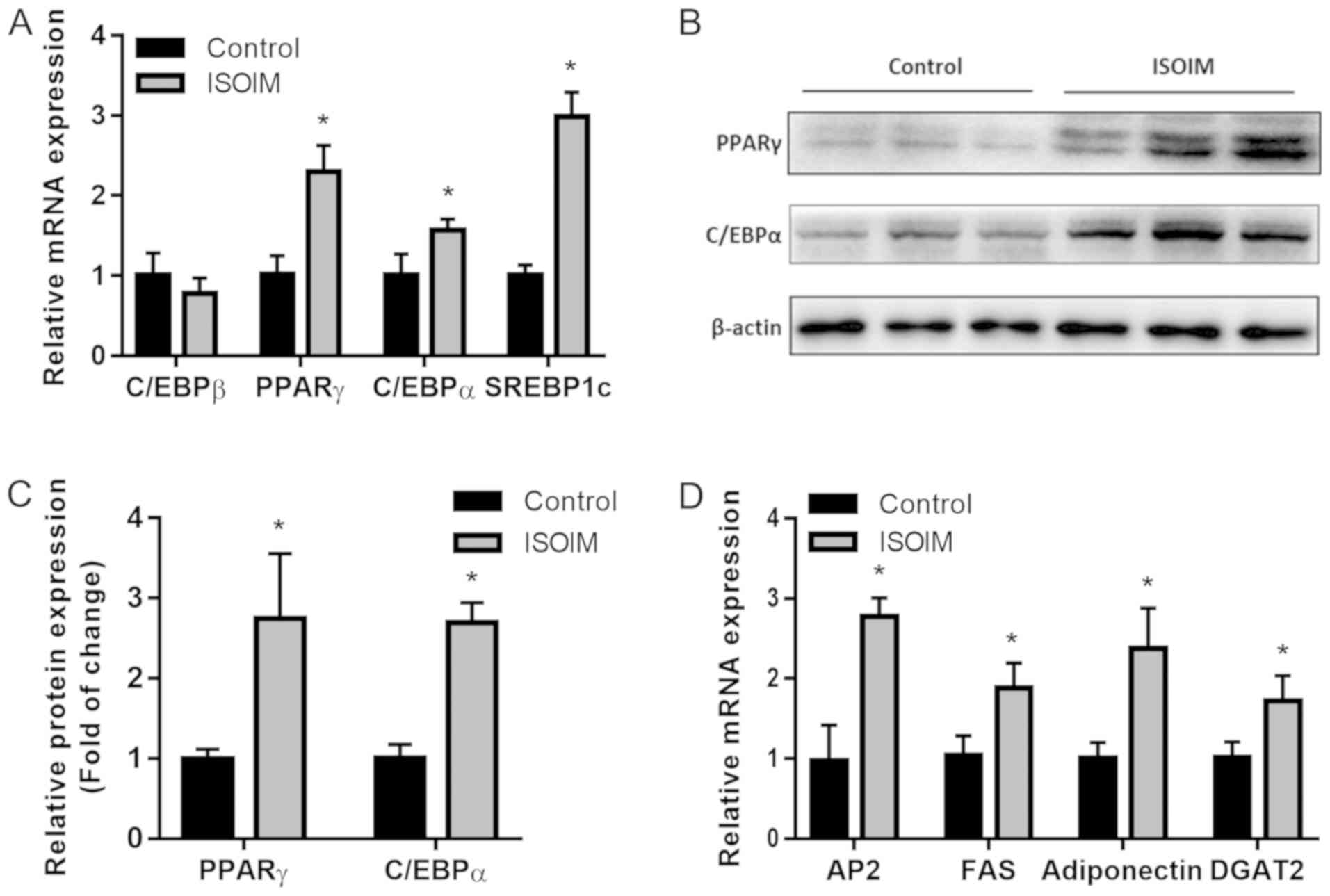
To examine ISOIM adipogenic efficacy during
differentiation, mRNA and protein expression of the genes related
to adipogenesis were measured in 3T3-L1 fibroblasts treated with or
without 20 µM ISOIM for 6 days. Compared with the control,
ISOIM-treated differentiated adipocytes had higher PPARγ, C/EBPα
and SREBP1c mRNA levels, but not C/EBPβ mRNA levels (Fig. 4A). In accordance with the
aforementioned findings, western blot analysis demonstrated that
ISOIM increased PPARγ (2.7 fold) and C/EBPα (2.6 fold) protein
expression, two important adipogenesis transcriptional factors, in
comparison to the control (Fig. 4B and
C). Moreover, mRNA expression of PPARγ target genes and
differentiation markers of late-stage adipocytes including aP2 (2.8
fold), FAS (1.9 fold), adiponectin (2.4 fold) and DGAT2 (1.7 fold)
increased relative to the control (Fig.
4D). Results suggested that ISOIM promoted 3T3-L1 cell
adipogenesis by upregulating PPARγ and C/EBPα.
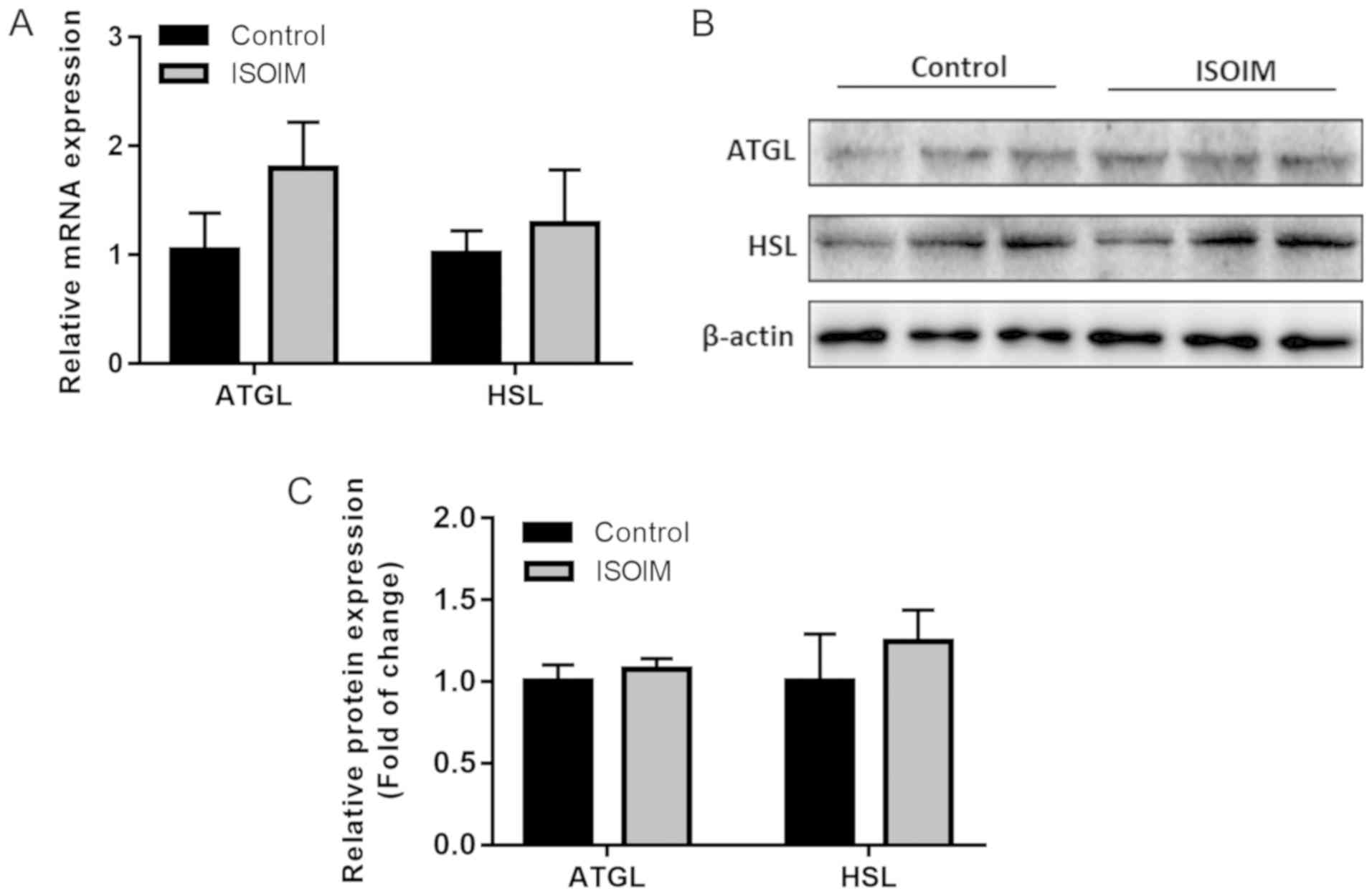
ISOIM does not significantly affect
lipolysis-related mRNA and protein expression in differentiated
adipocytes
The study assessed whether ISOIM accelerated
adipogenesis by downregulating lipolysis-related genes. mRNA
expression of the genes encoding the lipolytic enzymes ATGL and HSL
increased slightly but not significantly compared to the control
(Fig. 5A). Similarly, there was no
marked difference in the ATGL and HSL protein levels between the
ISOIM and control groups (Fig. 5B and
C). These findings demonstrated that ISOIM did not affect
lipolysis.
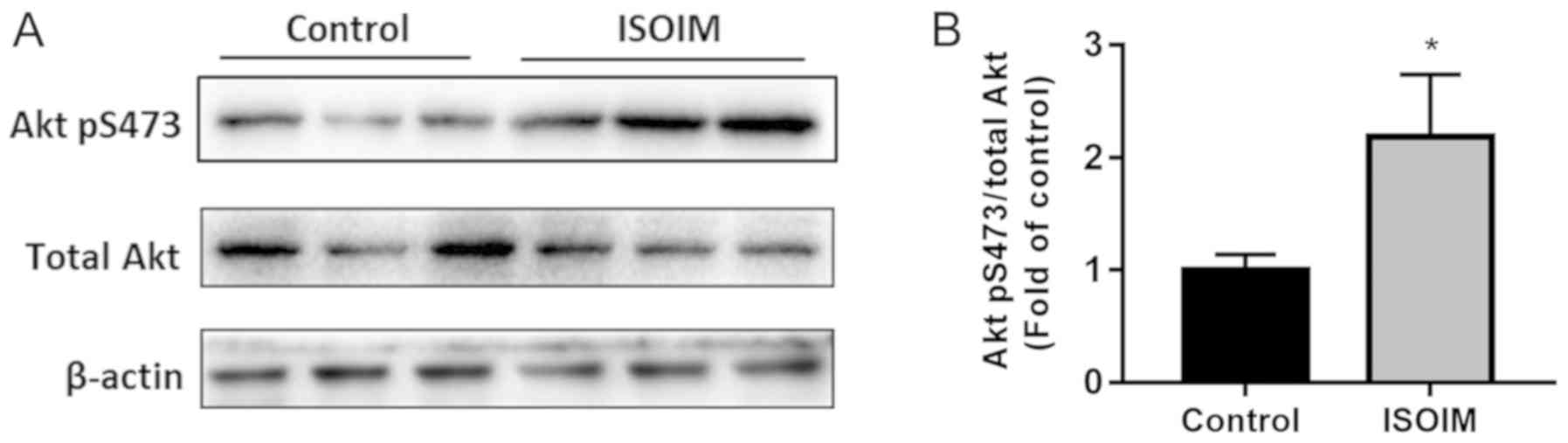
Effect of ISOIM on the Akt signaling
pathway
Akt has an important role in adipocyte
differentiation, which affects lipid metabolism through the insulin
pathway (13). To define the
underlying molecular mechanism of ISOIM-induced adipocyte
differentiation, differentiated 3T3-L1 adipocytes were treated with
ISOIM and Akt protein phosphorylation was detected. In addition to
inducing the differentiation of 3T3-L1 cells with insulin, 3T3-L1
cells were treated with 20 µM ISOIM for 6 days. ISOIM increased Akt
phosphorylation significantly (2.2 fold) compared with the control
(Fig. 6A and B). These results
suggest that ISOIM increased Akt phosphorylation during adipocyte
differentiation.
Discussion
ISOIM has anti-inflammatory (19), anti-hypertension (20), analgesic (19), anti-cancer (21) and hepatoprotective properties
(22). In the present study, ISOIM
promoted adipogenesis in 3T3-L1 cells in a dose-dependent manner,
and significantly increased lipid accumulation in the cells.
Mechanistically, ISOIM stimulated activation of the insulin
signaling pathway by phosphorylating Akt, critical for PPARγ and
C/EBPα expression and transcriptional activity. This may have led
to enhanced expression of PPARγ, C/EBPα, aP2, FAS, adiponectin and
DGAT2 genes, which are involved in the differentiation of 3T3-L1
adipocytes.
Adipogenesis is complex and regulated by a cascade
of transcription factors and other regulatory proteins (25). Previous studies have indicated that
C/EBPβ is pivotal in regulating adipogenesis, and its activation is
essential for initiating mitotic clonal expansion in adipocyte
differentiation (26–28). Following a 16–20 h delay, C/EBPβ
activates PPARγ and C/EBPα expression coordinately through
regulatory elements of C/EBP in their respective genes' proximal
promoters (29,30). However, the present study determined
that ISOIM did not regulate C/EBPβ mRNA expression in mature
adipocytes. PPARγ is the adipocyte differentiation master
regulator, during which it modulates the expression of several
genes, whereas C/EBPα adipogenic activity relies on PPARγ being
present. The terminally differentiated state is maintained by this
transcription cascade via the induction of high expression levels
of PPARγ and C/EBPα in a positive feedback loop (31–33). The
present study determined that ISOIM increased PPARγ and C/EBPα mRNA
and protein expression significantly, indicating that ISOIM can
enhance adipocyte differentiation effectively by upregulating PPARγ
and C/EBPα, rather than C/EBPβ.
For the downstream gene products of adipogenesis,
SREBP1c binds to the FAS promoter to promote lipogenesis whilst
also increasing aP2 expression levels in adipocytes (34). aP2 expression is found primarily in
mature adipose tissue, which regulates lipid and glucose metabolism
(35). Long-chain fatty acids are
catalyzed and synthesized from acetyl-coenzyme A (CoA) by FAS, a
multifunctional lipogenesis enzyme (36). Adipocytes secrete adiponectin, which
modulates glucose regulation and fatty acid oxidation (37,38).
DGAT2 catalyzes triglyceride formation from diacylglycerol and
acyl-CoA, considered the terminal and sole committed step in the
synthesis of triglycerides, and is essential for adipogenesis
(39). The present study determined
that ISOIM significantly increased SREBP1c, aP2, FAS, adiponectin
and DGAT2 expression, which would likely mean enhanced fatty acid
transport and synthesis of lipids.
Lipolysis affects lipid accumulation in adipocyte
differentiation. A lipolytic enzyme cascade catalyzes triglyceride
hydrolysis in adipocytes, where ATGL and HSL form the first and
second steps (40). Therefore, the
effect of ISOIM on lipolysis in 3T3-L1 adipocytes was investigated.
Results determined that there was a trend for increased ATGL and
HSL mRNA levels that was not statistically significant. Similarly,
ATGL and HSL protein levels did not significantly change, which
indicated that ISOIM does not regulate triglyceride hydrolysis
during 3T3-L1 adipocyte differentiation; thus ISOIM does not affect
lipid accumulation through triglyceride hydrolysis.
Finally, the signaling pathway involved in 3T3-L1
adipocyte differentiation was investigated. Akt has an essential
function in adipocyte differentiation into lipids (41). Akt deletion impairs the ability of
the 3T3-L1 cells to differentiate into adipocytes (15). In addition, PPARγ and C/EBPα
transcript upregulation activates Akt, thereby increasing 3T3-L1
cell glucose uptake and differentiation into adipocytes (13). Akt phosphorylation regulates several
biological processes (42), and is
essential in PPARγ expression induction (14). Numerous studies have reported that
Akt regulates PPARγ and adipocyte differentiation. Balakrishnan
et al (43) reported that
hyperglycemia in 3T3-L1 adipocytes is ameliorated by Moringa
concanensis Nimmo upregulation of PPARγ and C/EBPα via the Akt
signaling pathway. Conversely, Choe et al (44) reported that 3T3-L1 cell adipogenesis
is attenuated by water-extracted plum (Prunus salicina L.
cv. Soldam) via the PI3K/Akt signaling pathway. The present study
determined that compared with the control, ISOIM increased Akt
phosphorylation levels during the differentiation of 3T3-L1
preadipocytes. Therefore, the present results indicated that ISOIM
may enhance adipocyte differentiation and lipid accumulation by
enhancing PPARγ and C/EBPα expression via the Akt pathway.
To summarize, the present study demonstrated that
ISOIM increased the differentiation and accumulation of lipids of
3T3-L1 cells. Moreover, these results demonstrate that ISOIM may
have potential as a natural agent for the prevention and
improvement of diabetes.
Acknowledgements
The authors would like to acknowledge the
experimental condition and platform provided by Professor Gongshe
Yang's laboratory and Professor Jiangwei Wu's laboratory at
Northwest A&F University. They also wish to thank Professor
Jiangwei Wu, Dr Yongliang Wang and Dr Youlei Li for their helpful
discussions on experimental design.
Funding
The Discipline Construction Fund Project of Gansu
Agricultural University (grant no. GAU-XKJS-2018-054) supported the
present study.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
TJ and SG conceived and designed the experiments.
TJ, XS and ZY performed the experiments. TJ and XW analyzed the
data. TJ and SG drafted the manuscript. The final manuscript was
read and approved by all authors.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Petersen MC and Shulman GI: Mechanisms of
insulin action and insulin resistance. Physiol Rev. 98:2133–2223.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Samuel VT, Petersen KF and Shulman GI:
Lipid-induced insulin resistance: Unravelling the mechanism.
Lancet. 375:2267–2277. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Virtue S and Vidal-Puig A: Adipose tissue
expandability, lipotoxicity and the metabolic syndrome-an
allostatic perspective. Biochim Biophys Acta. 1801:338–349. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scherer PE: Adipose tissue: From lipid
storage compartment to endocrine organ. Diabetes. 55:1537–1545.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vatier C, Vantyghem M-C, Storey C, Jéru I,
Christin-Maitre S, Fève B, Lascols O, Beltrand J, Carel JC,
Vigouroux C and Bismuth E: Monogenic forms of lipodystrophic
syndromes: Diagnosis, detection, and practical management
considerations from clinical cases. Curr Med Res Opin. 35:543–552.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ghaben AL and Scherer PE: Adipogenesis and
metabolic health. Nat Rev Mol Cell Biol. 20:242–258. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nawrocki AR and Scherer PE: Keynote
review: The adipocyte as a drug discovery target. Drug Discov
Today. 10:1219–1230. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rosen ED and MacDougald OA: Adipocyte
differentiation from the inside out. Nat Rev Mol Cell Biol.
7:885–896. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rosen ED, Sarraf P, Troy AE, Bradwin G,
Moore K, Milstone DS, Spiegelman BM and Mortensen RM: PPAR gamma is
required for the differentiation of adipose tissue in vivo and in
vitro. Mol Cell. 4:611–617. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rosen ED, Walkey CJ, Puigserver P and
Spiegelman BM: Transcriptional regulation of adipogenesis. Genes
Dev. 14:1293–1307. 2000.PubMed/NCBI
|
|
11
|
Xavier MN, Winter MG, Spees AM, den
Hartigh AB, Nguyen K, Roux CM, Silva TM, Atluri VL, Kerrinnes T,
Keestra AM, et al: PPAR gamma-mediated increase in glucose
availability sustains chronic brucella abortus infection in
alternatively activated macrophages. Cell Host Microbe. 14:159–170.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cristancho AG and Lazar MA: Forming
functional fat: A growing understanding of adipocyte
differentiation. Nat Rev Mol Cell Biol. 12:722–734. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saltiel AR and Kahn CR: Insulin signalling
and the regulation of glucose and lipid metabolism. Nature.
414:799–806. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Peng XD, Xu PZ, Chen ML, Hahn-Windgassen
A, Skeen J, Jacobs J, Sundararajan D, Chen WS, Crawford SE, Coleman
KG and Hay N: Dwarfism, impaired skin development, skeletal muscle
atrophy, delayed bone development, and impeded adipogenesis in mice
lacking Akt1 and Akt2. Genes Dev. 17:1352–1365. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rochford JJ: Mouse models of lipodystrophy
and their significance in understanding fat regulation. Curr Top
Dev Biol. 109:53–96. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun NN, Wu TY and Chau CF: Natural dietary
and herbal products in anti-obesity treatment. Molecules.
21:13512016. View Article : Google Scholar
|
|
17
|
Wang S, Chen Q and He L: Development and
validation of a gas chromatography-mass spectrometry method for the
determination of isoimperatorin in rat plasma and tissue:
Application to the pharmacokinetic and tissue distribution study. J
Chromatogr B Analyt Technol Biomed Life Sci. 852:473–478. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi X, Liu M, Zhang M, Zhang K, Liu S,
Qiao S, Shi R, Jiang X and Wang Q: Identification of in vitro and
in vivo metabolites of isoimperatorin using liquid
chromatography/mass spectrometry. Food Chem. 141:357–365. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wijerathne CUB, Seo CS, Song JW, Park HS,
Moon OS, Won YS, Kwon HJ and Son HY: Isoimperatorin attenuates
airway inflammation and mucus hypersecretion in an
ovalbumin-induced murine model of asthma. Int Immunopharmacol.
49:67–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hwang YH, Yang HJ and Ma JY: Simultaneous
determination of three furanocoumarins by UPLC/MS/MS: Application
to pharmacokinetic study of Angelica dahurica radix after
oral administration to normal and experimental Colitis-induced
rats. Molecules. 22:E4162017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang HB, Gao HR, Ren YJ, Fang FX, Tian HT,
Gao ZJ, Song W, Huang SM and Zhao AF: Effects of isoimperatorin on
proliferation and apoptosis of human gastric carcinoma cells. Oncol
Lett. 15:7993–7998. 2018.PubMed/NCBI
|
|
22
|
Pokharel YR, Han EH, Kim JY, Oh SJ, Kim
SK, Woo ER, Jeong HG and Kang KW: Potent protective effect of
isoimperatorin against aflatoxin B1-inducible cytotoxicity in H4IIE
cells: Bifunctional effects on glutathione S-transferase and CYP1A.
Carcinogenesis. 27:2483–2490. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang LY, Cheng KC, Li Y, Niu CS, Cheng JT
and Niu HS: The dietary furocoumarin imperatorin increases plasma
GLP-1 levels in type 1-like diabetic rats. Nutrients. 9:E11922017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang QQ and Lane MD: Adipogenesis: From
stem cell to adipocyte. Annu Rev Biochem. 81:715–736. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang JW, Tang QQ, Vinson C and Lane MD:
Dominant-negative C/EBP disrupts mitotic clonal expansion and
differentiation of 3T3-L1 preadipocytes. Proc Natl Acad Sci USA.
101:43–47. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park M, Choi YA, Lee HG, Kim KI, Lim JS,
Lee MS, Oh KS and Yang Y: Dephosphorylation of
CCAAT/enhancer-binding protein beta by protein phosphatase 2A
containing B56 delta is required at the early time of adipogenesis.
Biochim Biophys Acta. 1841:1608–1618. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao H, Zhang S, Shan S, Sun C, Li Y, Wang
H, Yu S, Liu Y, Guo F, Zhai Q, et al: Ligand-dependent corepressor
(LCoR) represses the transcription factor C/EBP during early
adipocyte differentiation. J Biol Chem. 292:18973–18987. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tang QQ and Lane MD: Activation and
centromeric localization of CCAAT/enhancer-binding proteins during
the mitotic clonal expansion of adipocyte differentiation. Genes
Dev. 13:2231–2241. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morrison RF and Farmer SR: Role of
PPARgamma in regulating a cascade expression of cyclin-dependent
kinase inhibitors, p18(INK4c) and p21(Waf1/Cip1), during
adipogenesis. J Biol Chem. 274:17088–17097. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Weisiger RA: Cytosolic fatty acid binding
proteins catalyze two distinct steps in intracellular transport of
their ligands. Mol Cell Biochem. 239:35–43. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rosen ED, Hsu CH, Wang X, Sakai S, Freeman
MW, Gonzalez FJ and Spiegelman B: C/EBPalpha induces adipogenesis
through PPARgamma: A unified pathway. Genes Dev. 16:22–26. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gonzalez FJ: Getting fat: Two new players
in molecular adipogenesis. Cell Metab. 1:85–86. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mota de Sá P, Richard AJ, Hang H and
Stephens JM: Transcriptional regulation of adipogenesis. Compr
Physiol. 7:635–674. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang Y, Viscarra J, Kim SJ and Sul HS:
Transcriptional regulation of hepatic lipogenesis. Nat Rev Mol Cell
Biol. 16:678–689. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moseti D, Regassa A and Kim WK: Molecular
regulation of adipogenesis and potential anti-adipogenic bioactive
molecules. Int J Mol Sci. 17:E1242016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fan H, Dong W, Li Q, Zou X, Zhang Y, Wang
J, Li S, Liu W, Dong Y, Sun H and Hou Z: Ajuba preferentially binds
LXR alpha/RXR gamma heterodimer to enhance LXR target gene
expression in liver cells. Mol Endocrinol. 29:1608–1618. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen J, Zhou X, Wu W, Wang X and Wang Y:
FTO-dependent function of N6-methyladenosine is involved in the
hepatoprotective effects of betaine on adolescent mice. J Physiol
Biochem. 71:405–413. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cases S, Smith SJ, Zheng YW, Myers HM,
Lear SR, Sande E, Novak S, Collins C, Welch CB, Lusis AJ, et al:
Identification of a gene encoding an acyl CoA: Diacylglycerol
acyltransferase, a key enzyme in triacylglycerol synthesis. Proc
Natl Acad Sci USA. 95:13018–13023. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Watt MJ and Steinberg GR: Regulation and
function of triacylglycerol lipases in cellular metabolism. Biochem
J. 414:313–325. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu J and Liao K: Protein kinase B/AKT 1
plays a pivotal role in insulin-like growth factor-1 receptor
signaling induced 3T3-L1 adipocyte differentiation. J Biol Chem.
279:35914–35922. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Green CJ, Göransson O, Kular GS, Leslie
NR, Gray A, Alessi DR, Sakamoto K and Hundal H: Use of Akt
inhibitor and a drug-resistant mutant validates a critical role for
protein kinase B/Akt in the insulin-dependent regulation of glucose
and system A amino acid uptake. J Biol Chem. 283:27653–27667. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Balakrishnan BB, Krishnasamy K and Choi
KC: Moringa concanensis Nimmo ameliorates hyperglycemia in
3T3-L1 adipocytes by upregulating PPAR-gamma, C/EBP-alpha via Akt
signaling pathway and STZ-induced diabetic rats. Biomed
Pharmacother. 103:719–728. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Choe WK, Kang BT and Kim SO:
Water-extracted plum (Prunus salicina L. cv. Soldam)
attenuates adipogenesis in murine 3T3-L1 adipocyte cells through
the PI3K/Akt signaling pathway. Exp Ther Med. 15:1608–1615.
2018.PubMed/NCBI
|