Introduction
CD4+CD25+forkhead box
(Fox)P3+ T-regulatory (Treg) cells are an important
subset of immune cells that may inhibit the activation,
proliferation and cytokine secretion of other types of immune cell.
Treg cells have an indispensable role in driving autoimmune
tolerance and maintaining the immune balance in vivo
(1–3). Abnormal Treg cell functions are widely
involved in the occurrence and development of numerous diseases
(4–6), and immunotherapy to recover the number
and/or function of Treg cells is a good optional treatment for such
diseases.
Immunotherapy with transplanted Treg cells has been
used in autoimmune diseases and other immune-associated diseases,
including type-1 diabetes mellitus, systemic lupus erythematosus
(SLE) and graft vs. host disease (GVHD) (7–13).
Culturing sufficient numbers of Treg cells in vitro is the
foundation of Treg-based immunotherapy, and maintaining the stable
inhibitory function of Treg cells in vivo is pivotal for
successful treatment (8,9). However, the stability and inhibitory
function of Treg cells in the internal inflammatory environment
requires further systematic investigation.
The internal environment of patients with autoimmune
diseases is complex and there may be inflammation or elevated
levels of inflammatory cytokines, including tumour necrosis
factor-α, interleukin-1 (IL-1), interleukin-6 (IL-6),
interleukin-23 (IL-23) and interferon-γ (IFN-γ) expressed in human
atherosclerotic plaques (14,15);
interleukin-17 (IL-17), IFN-γ, IL-6 and IL-23 expressed in type 1
diabetes mellitus (16); IL-1β and
IL-17 expressed in SLE (17); and
IL-6 expressed in GVHD (9,18). IL-6 is the critical cytokine that
mediates inflammation (18,19). As mentioned above, IL-6 is highly
expressed in autoimmune diseases and GVHD (9,14–16,18),
and the inflammatory environment in vivo may be simulated by
adding IL-6.
In the present study, the possible inflammatory
environment was simulated in vitro by using recombinant
human (rh)IL-6 to observe the absolute number, stability, activity
and inhibitory function of Treg cells. The present study lays a
foundation for Treg cell-based immunotherapy in various
diseases.
Materials and methods
Samples
A total of eight healthy blood donors were recruited
from Shaanxi Provincial People's Hospital Affiliated to Xi'an
Medical University (Xi'an, China); the male/female ratio was 4:4,
and the average age was 27.8±1.3 years. A total of 40 ml sterile
peripheral venous blood samples (including heparin to prevent
clotting) were collected from all healthy blood donors. The study
was approved by the Ethics Committee of Xi'an Medical University
(Xi'an, China; approval no. XYLS2019131). According to the
principle of informed consent, all healthy blood donors signed
consent forms prior to collection of the peripheral blood samples.
All of the experiments in this study were performed in accordance
with the guidelines for blood sample collection approved by the
Institutional Ethics Committee of Xi'an Medical University.
Isolation of peripheral blood
mononuclear cells (PBMCs)
PBMCs were isolated from the samples via
density-gradient centrifugation. First, 20 ml of Lymphoprep™
(Axis-Shield) was added to each centrifuge tube, and then, 20 ml of
the individual peripheral blood sample diluted with an equal volume
of PBS was slowly added. After centrifugation for 20 min at 500 × g
under room temperature, the centrifuge tubes were gently removed
and the monocyte suspension was isolated and washed with PBS. After
the erythrocytes were lysed with FACS lysis solution (BD
Biosciences), the isolated PBMCs were washed with PBS and then
resuspended in PBS and counted.
Sorting of Treg cells and T-effector
(Teff) cells
After 4×107 PBMCs were resuspended in
RPMI 1640 Media supplemented with 10% fetal bovine serum and 100
U/ml penicillin and 100 mg/ml streptomycin (All Gibco; Thermo
Fisher Scientific, Inc.), peridinin chlorophyll (PerCP)-conjugated
anti-CD4 (cat. no. 347324, BD Biosciences) and allophycocyanine
(APC)-conjugated anti-CD25 antibodies (cat. no. 555434, BD
Biosciences) were added both at a dilution of 1:5 with the final
concentration of PBMCs at 1×106/100 µl. Another
1×106 PBMCs were resuspended in media mentioned above,
PerCP-conjugated Mouse IgG1 κ Isotype Control (cat. nos. 559425, BD
Biosciences) and APC-conjugated Mouse IgG1 κ Isotype Control (cat.
no. 555751, BD Biosciences) were added at a dilution of 1:5 with
the final concentration of PBMCs at 1×106/100 µl. After
incubation with the antibodies at 4°C protected from light for 20
min, the PBMCs were washed with PBS and resuspended in fully
supplemented RPMI 1640 Medium. After filtration with a cell
strainer (Falcon; BD Biosciences), the stained PBMCs were processed
with a BD FACS Aria II (BD Biosciences) and the
CD4+CD25high Treg cells and
CD4+CD25− Teff cells were sorted.
CD25− cells were defined using the isotype control
antibody and CD25high cells were defined according to a
previous reported protocol (20).
In vitro culture of Treg cells
rhIL-2 and anti-CD3/CD28 beads
(Dynabeads® Human T-Expander CD3/CD28 beads; Life
Technologies; Thermo Fisher Scientific, Inc.) are necessary for the
culturing of Treg cells in vitro, since IL-2 signals are
required for the maintenance of lineage stability, survival and
suppressor function in mature Tregs, whilst CD3 and CD28 signaling
is crucial for the survival and proliferation of Treg cells
(21–23). Freshly sorted Treg cells were
cultured (1×105/well) in a 96-well U-bottomed plate in
the presence of 100, 300 or 500 U/ml rhIL-2 (HumanZyme; ProteinTech
Group, Inc.) and anti-CD3/CD28 beads (Treg cells/beads ratio,
0.5:1, 1:1 or 2:1), with or without 25 ng/ml rhIL-6 (HumanZyme)
under 37°C in a humidified atmosphere containing 5% CO2.
The cell concentration was 1×106/ml and 100 µl/well. The
Treg cell population was counted after culturing for 3, 5, 7, 10
and 14 days, the medium was replaced after culturing for 3, 5, 7
and 10 days with RPMI 1640 Media supplemented with 10% fetal bovine
serum, 100 U/ml penicillin, 100 mg/ml streptomycin and
corresponding concentrations of rhIL-2 and rhIL-6.
Stability and activity of Treg
cells
Treg cells were collected after culturing for 3, 5,
7, 10 and 14 days. After the cells were washed and resuspended in
PBS, PerCP-conjugated anti-CD4 and APC-conjugated anti-CD39 (cat.
no. 560239; BD Biosciences) were added at a dilution of 1:5 with
final concentration of cells at 1×106/100 µl. The cells
were then incubated with the antibodies at 4°C while protected from
light for 20 min. The cells were then washed twice with PBS and
treated for 30 min with 1 ml fixation/permeabilization buffers and
washed three times with 2 ml permeabilization buffer (cat. no.
00-5523-00; Foxp3/transcription factor staining buffer set;
eBioscience; Thermo Fisher Scientific, Inc.). Finally, the cells
were washed twice with PBS and incubated at 4°C in the dark for 30
min with PE-conjugated anti-FoxP3 antibody (cat. no. 12-4777-42;
eBioscience) at a dilution of 1:20. APC-conjugated Mouse IgG2b κ
Isotype Control (cat. no. 555745; BD Biosciences) were used at a
dilution of 1:5 and PE-conjugated Mouse IgG1 kappa Isotype Control
(P3.6.2.8.1; cat. no. 12-4714-82; eBioscience) were used at a
dilution of 1:20 in the staining procedures. After being washed
twice with PBS and filtered using a cell strainer, the stained
cells were processed using BD FACSCalibur (BD Biosciences) and the
proportions of CD4+FoxP3+ Treg cells and
CD39+CD4+FoxP3+ active Treg cells
were analysed.
Detection of inhibitory function of
Treg cells
The inhibitory function of Treg cells was measured
by detecting the suppression of Teff cell proliferation in the
presence of Treg cells. Freshly sorted Teff cells were labelled
with 5,6-carboxyfluorescein succinimidyl ester (CFSE; CellTrace™
CFSE Cell Proliferation Kit; Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. The labelled cells
(1×105/well) were cultured with or without sorted Treg
cells (5×104/well) in a 96-well U-bottomed plate in the
presence of anti-CD3/CD28 beads (Teff cells/beads, 1:1), without or
with 25 ng/ml rhIL-6. rhIL-2 was not added, as the Teff cells are
able to secrete IL-2 after activation with anti-CD3/CD28 beads
(24). The Teff cell proliferation
was determined by flow cytometry after Teff cells were cultured for
5 days. The proliferation curves of the Teff cells were fitted, and
the Teff-cell proliferation were expressed as the response (R; the
proportion of the precursor sample pool that responded to
stimulation by dividing), doubling time (Td; the time required for
the average responding T cell to achieve a single cell division)
and the proliferative capacity (Cp) of the responding T cells in
each sample. The methodology for calculating R, Td and Cp from the
proliferation curves was described in detail in previous studies
(24–26).
Statistical analysis
All flow cytometry results of the present study were
analysed with FlowJo software version 7.6.1 (FlowJo LLC).
Calculations were performed using GraphPad Prism software, version
6 (GraphPad Software, Inc). Values are expressed as the mean ±
standard error of the mean, the significance of differences between
two groups were calculated using paired t-test and Student's
unpaired 2-tailed t-te#st. The significance of differences among
three groups was calculated using One-way ANOVA followed by Tukey's
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
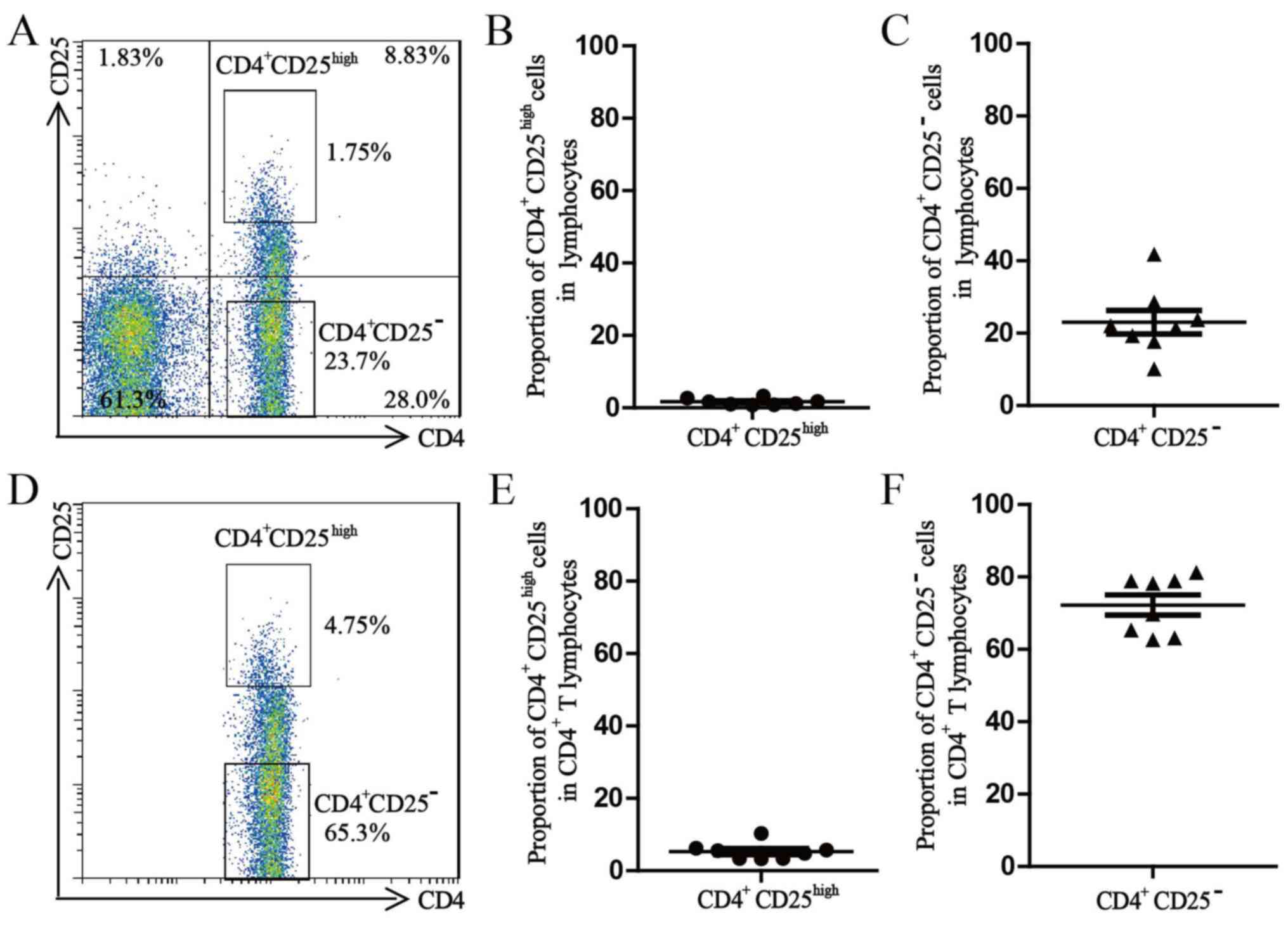
Sorting of Treg cells and Teff
cells
After extracellular staining with PerCP-conjugated
anti-CD4 and APC-conjugated anti-CD25, the freshly isolated PBMCs
were processed with a BD FACS Aria II to sort the Treg cells and
Teff cells (98.0% purity for each subset). The average proportion
of CD4+CD25high Treg cells in the lymphocyte
population was 1.7±0.3% and the average proportion of
CD4+CD25high Treg cells in the
CD4+ T-lymphocyte population was 5.3±0.8%. The average
proportion of CD4+CD25− Treg cells in the
lymphocyte population was 23.1±3.3% and the average proportion of
CD4+CD25high Treg cells in the
CD4+ T-lymphocyte population was 72.2±2.8% (Fig. 1).
Optimum concentration of rhIL-2 and
anti-CD3/CD28 beads for culture of Treg cells
The absolute counts of Treg cells cultured with 100,
300 or 500 U/ml rhIL-2 for 14 days in the presence of anti-CD3/CD28
beads (Treg cells/beads, 1:1) in vitro are presented in
Fig. 2A. The number of Treg cells
was 1×105/well on day 0, except for the experiment for
detection of inhibitory function. The number of Treg cells
increased steadily until reaching a plateau ~day 10 after culturing
in vitro. There were no differences among the counts of Treg
cells cultured with 100, 300 or 500 U/ml rhIL-2 for 3 or 5 days.
There was a significant difference between the counts of Treg cells
cultured with 100 and 300 U/ml rhIL-2 for 7, 10 or 14 days. There
was no difference between the counts of Treg cells cultured with
300 and 500 U/ml rhIL-2 for 7, 10 or 14 days.
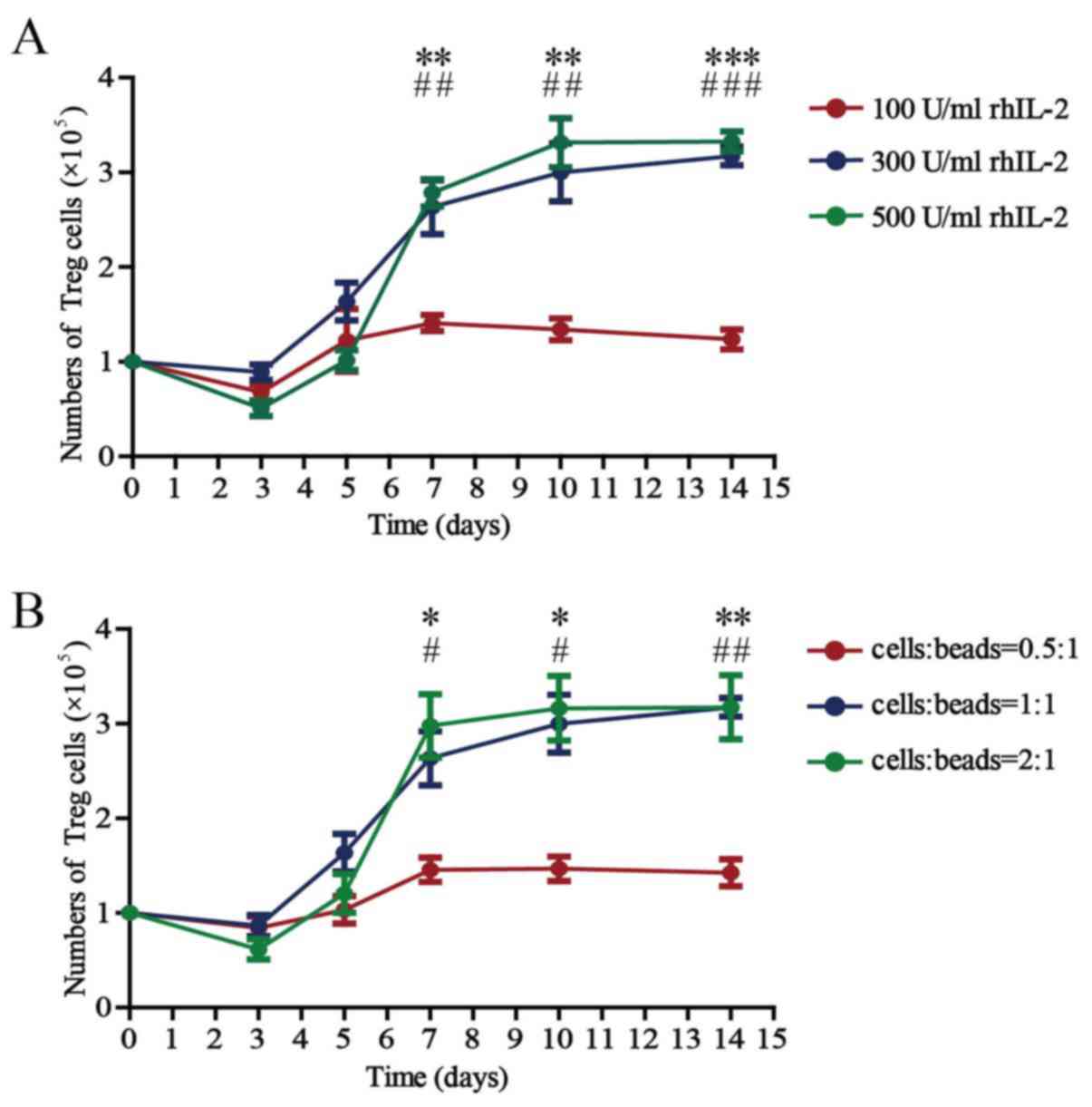 | Figure 2.Number of Treg cells cultured in the
presence of different concentrations of rhIL-2 and anti-CD3/CD28
beads. (A) Number of all Treg cells after 3, 5, 7, 10 and 14 days
of culture in the presence of anti-CD3/CD28 beads (Treg
cells/beads, 1:1) and rhIL-2 (100, 300 or 500 U/ml). Values are
expressed as the mean ± standard error of the mean (n=8).
**P<0.01 and ***P<0.0001, 100 vs. 300 U/ml rhIL-2 on the
indicated days and ##P<0.01 and
###P<0.0001, 100 vs. 500 U/ml rhIL-2 on the indicated
days. (B) Number of all Treg cells after 3, 5, 7, 10 and 14 days of
culture in the presence of rhIL-2 (300 U/ml) and anti-CD3/CD28
beads (Treg cells/beads, 0.5:1, 1:1 or 2:1). Values are expressed
as the mean ± standard error of the mean (n=8). *P<0.05 and
**P<0.01, cells:beads 0.5:1 vs. 1:1 on the indicated days and
#P<0.05 and ##P<0.01, cells/beads 0.5:1
vs. 2:1 on the indicated days. rhIL, recombinant human interleukin;
Treg cells, T-regulatory cells. |
The absolute counts of the Treg cells cultured for
14 days with anti-CD3/CD28 beads (Treg cells/beads, 0.5:1, 1:1 or
2:1) in the presence of 300 U/ml rhIL-2 in vitro are
presented in Fig. 2B. The Treg cell
counts also increased with culture time until plateauing ~day 10.
There were no differences among the counts of Treg cells cultured
in vitro, whether the ratio of cells to beads was 0.5:1, 1:1
or 2:1 during culture for 3 or 5 days. There was a significant
difference between the counts of Treg cells cultured with beads at
a 0.5:1 cell-to-bead ratio and those cultured with beads at a 1:1
cell-to-bead ratio at 7, 10 or 14 days. There was no difference in
the counts between Treg cells cultured with beads at a 1:1 ratio
and those cultured with beads at a 2:1 ratio at 7, 10 or 14
days.
The above results demonstrate that the absolute
count of the Treg cells is significantly affected by the
concentration of rhIL-2 and anti-CD3/CD28 beads; 300 U/ml rhIL-2
and a 1:1 cell/bead ratio are the optimal concentrations when the
Treg cells are cultured in vitro, and
3.17±0.10×105 cells were obtained after 14 days of
culture.
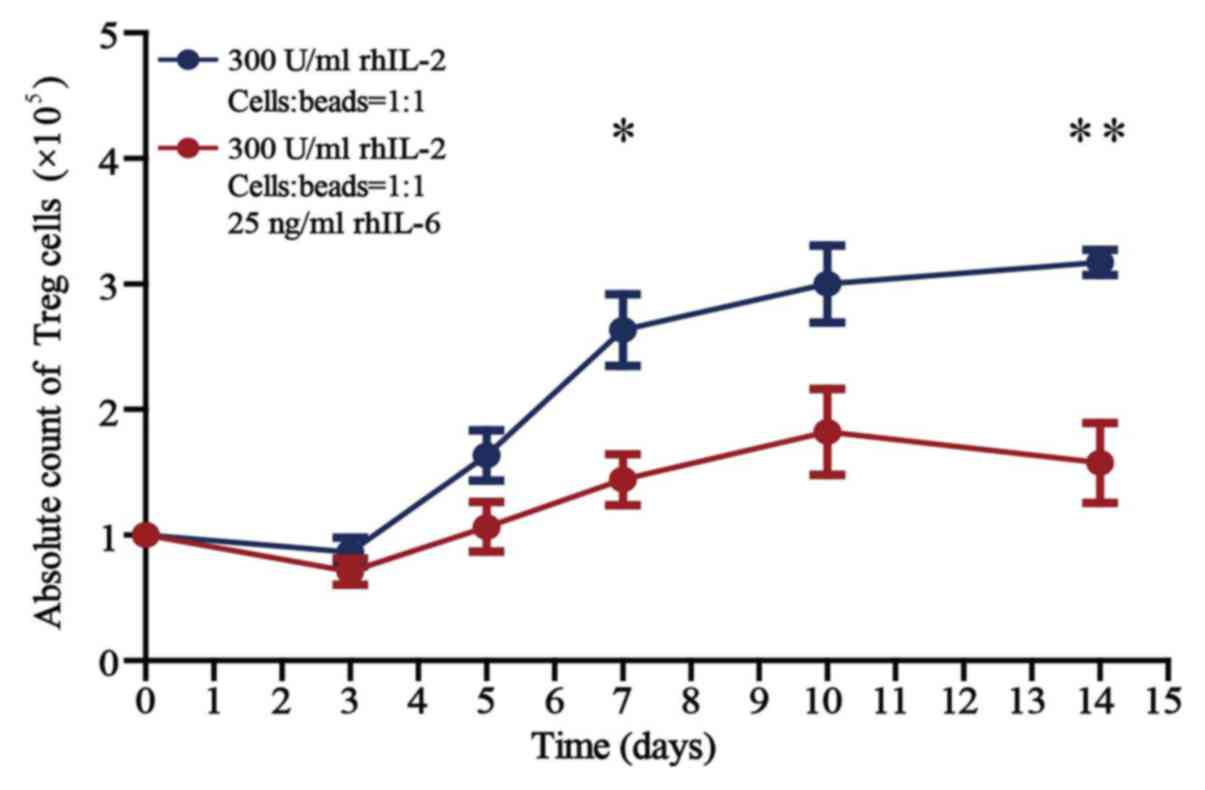
The Treg cell number is reduced under
inflammatory conditions
The absolute counts of Treg cells cultured for 14
days in the presence of rhIL-2 (300 U/ml) and anti-CD3/CD28 beads
(Treg cells/beads, 1:1), without or with 25 ng/ml rhIL-6 in
vitro are presented in Fig. 3.
There was no difference in the counts of Treg cells cultured
without and with 25 ng/ml rhIL-6 when cultured for <5 days in
vitro. There was a significant difference in the counts of Treg
cells cultured without and with 25 ng/ml rhIL-6 when cultured for
>7 days (P<0.05). The number of Treg cells cultured in the
presence of 25 ng/ml rhIL-6 for 14 days was reduced by 49.7% when
compared with that of cells cultured without rhIL-6.
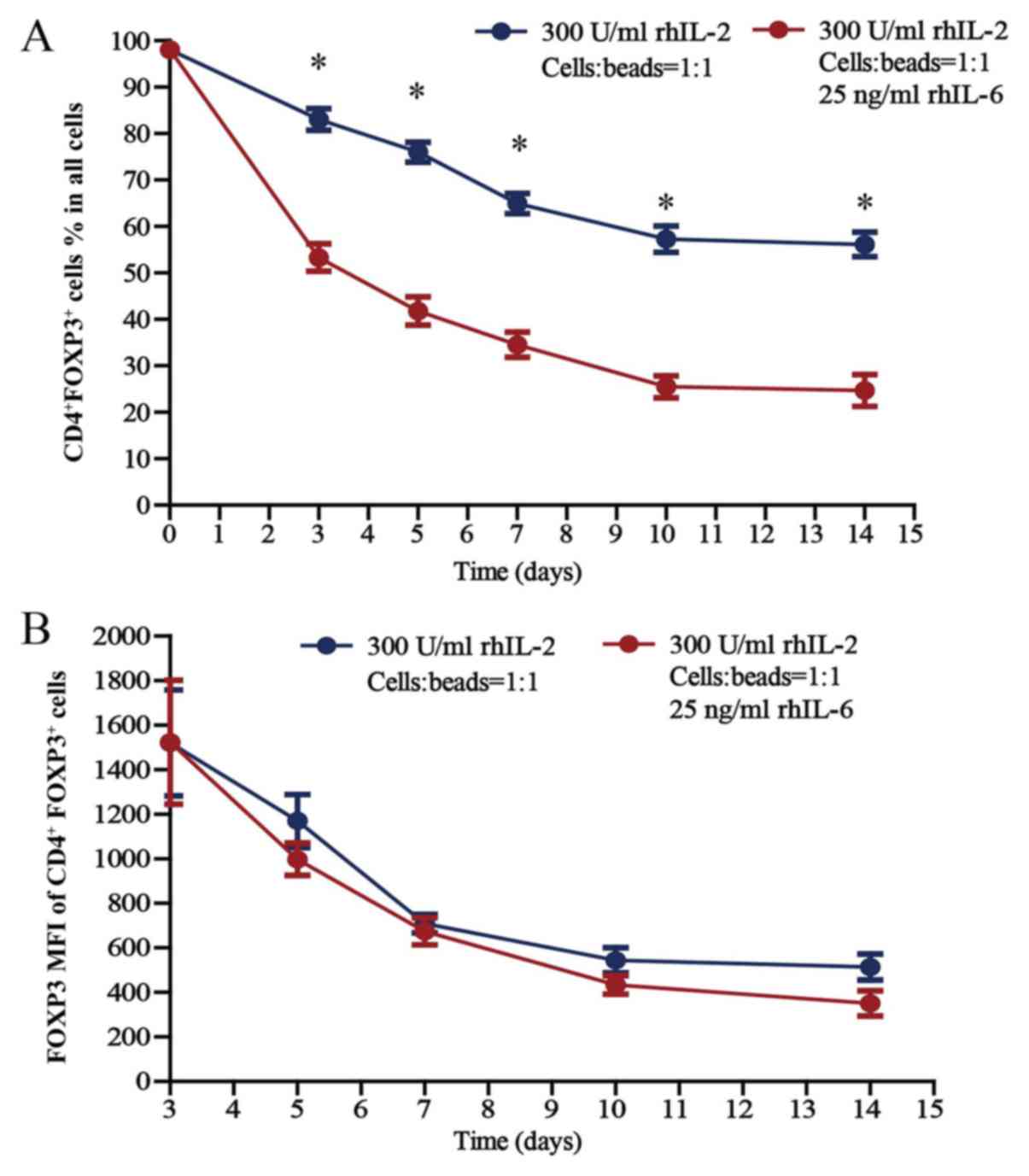
The stability of Treg cells declines
under inflammatory conditions
Treg cells were cultured (300 U/ml rhIL-2; Treg
cells/beads, 1:1; without or with 25 ng/ml rhIL-6) for 14 days
in vitro; the proportion of CD4+FoxP3+
Treg cells within the total cell population of the well (thereafter
referred to as total cell population) and the FoxP3 mean
fluorescence intensity (MFI) of CD4+FoxP3+
Treg cells are presented in Fig. 4A and
B, respectively. The proportion of
CD4+FoxP3+ Treg cells was 98.0% on day 0, as
the purity for sorted Treg cells was 98.0%. Regarding the
proportion of CD4+FoxP3+ Treg cells within
the total cell population, there was a significant difference in
the proportion of CD4+FoxP3+ Treg cells
cultured without and with 25 ng/ml rhIL-6 when cultured for 3, 5,
7, 10 and 14 days (P<0.05). In terms of the FoxP3 MFI of
CD4+FoxP3+ Treg cells, there was no
difference between the cells cultured without and with 25 ng/ml
rhIL-6, whether cultured for 3, 5, 7, 10 or 14 days in
vitro.
Regarding the proportion of
CD4+FoxP3+ cells within the total cell
population cultured in the presence of rhIL-2, which gradually
declined over the 14-day culture period, the proportion of Treg
cells cultured with rhIL-6 decreased much faster compared with the
proportion of Treg cells cultured without rhIL-6 (Fig. 4A). Among the Treg cells cultured
without rhIL-6, there were always certain amounts of cells
expressing FoxP3, although the proportion declined gradually, and
the proportion of CD4+FoxP3+ Treg cells
within the total cell population was 56.2±2.7% after Treg cells
were cultured for 14 days. However, FoxP3 expression decreased more
rapidly in the Treg cells cultured with rhIL-6, and the proportion
of CD4+FoxP3+ Treg cells within the total
cell population was 24.7±3.4% after Treg cells were cultured for 14
days, which suggests a lower stability of Treg cells in the
inflammatory environment and that increased numbers of Treg cells
should be used in treatments considering the complexity of the
internal environments of patients.
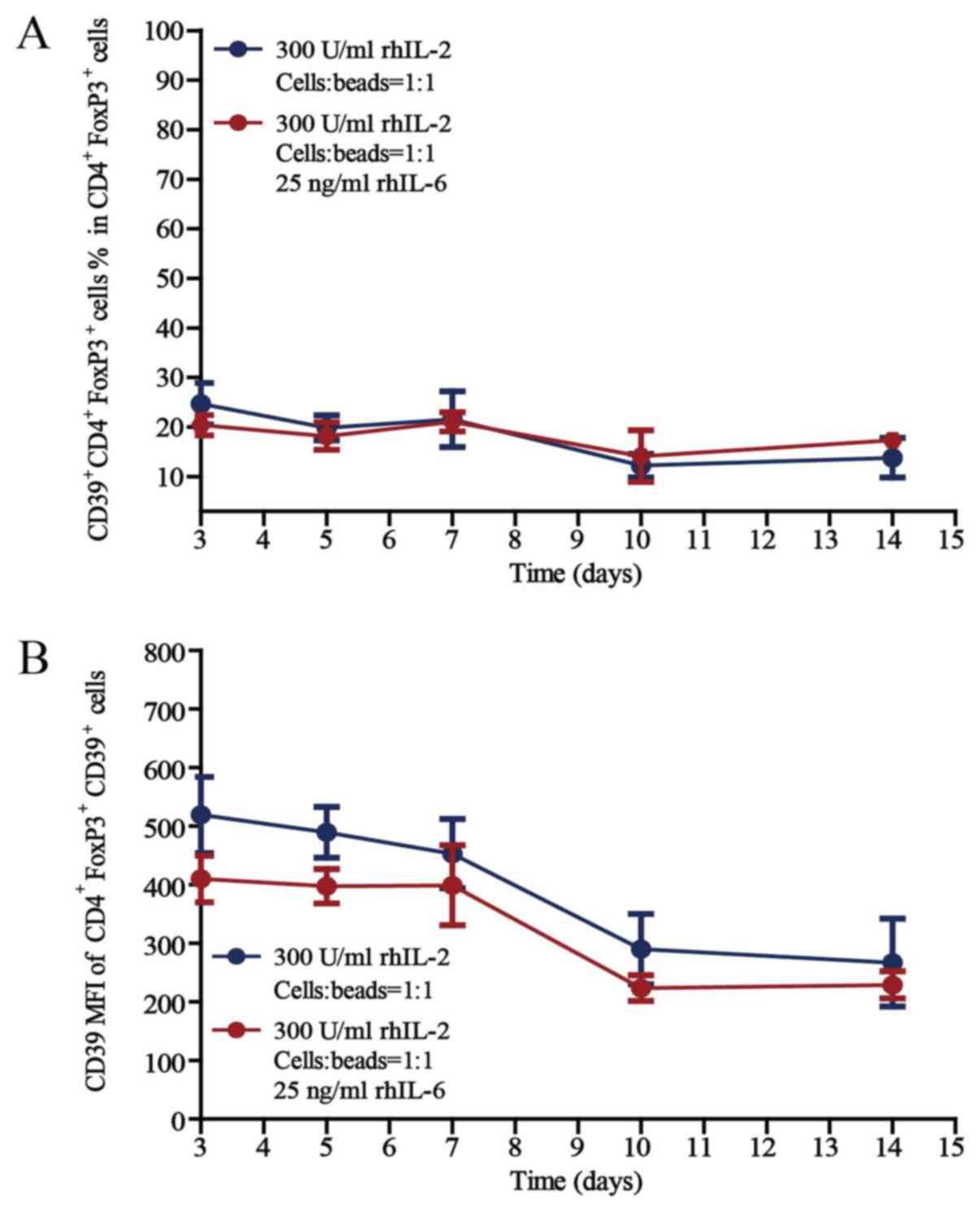
The activity of Treg cells remains
unchanged under inflammatory conditions
The activity of Treg cells was then investigated by
detecting CD39 expression, as the expression of CD39 on T cells
reflects high activity (27). The
proportions of CD39+CD4+FoxP3+
active Treg cells in the CD4+FoxP3+ cell
population after culture for 3, 5, 7, 10 or 14 days in the presence
of rhIL-2 (300 U/ml) and anti-CD3/CD28 beads (Treg cells/beads,
1:1), without or with 25 ng/ml rhIL-6, are provided in Fig. 5. Regarding the proportion of
CD39+CD4+FoxP3+ Treg cells in the
CD4+FoxP3+ cell population and the CD39 MFI
of CD39+CD4+FoxP3+ Treg cells,
there was no difference between the cells cultured without rhIL-6
and with 25 ng/ml rhIL-6, whether cultured for 3, 5, 7, 10 or 14
days in vitro.
The proportion of active Treg cells in the
CD4+FoxP3+ cell population remained unchanged
during the 14 days of culture in vitro, even under
inflammatory conditions, which further proves the stability of the
Treg-cell subset and provides a basis for the clinical application
of Treg cells.
The inhibitory function of Treg cells
declines under inflammatory conditions
Finally, the suppressive function of Treg cells
cultured without or with 25 ng/ml rhIL-6 was investigated by
measuring their ability to suppress the Teff cell proliferation
in vitro. After Treg and Teff cells were sorted from the
PBMCs of healthy donors (n=8), the Teff cells were labelled with
CFSE and co-cultured with Treg cells (Teff cells/Treg cells, 2:1)
in the presence of anti-CD3/CD8 beads (Teff cells/beads, 1:1),
without or with 25 ng/ml rhIL-6. Teff-cell proliferation was
detected after culture for 5 days by gating on CFSE+
cells, and the results of the corresponding R, Td, and Cp
measurements were presented in Fig.
6 (24–26).
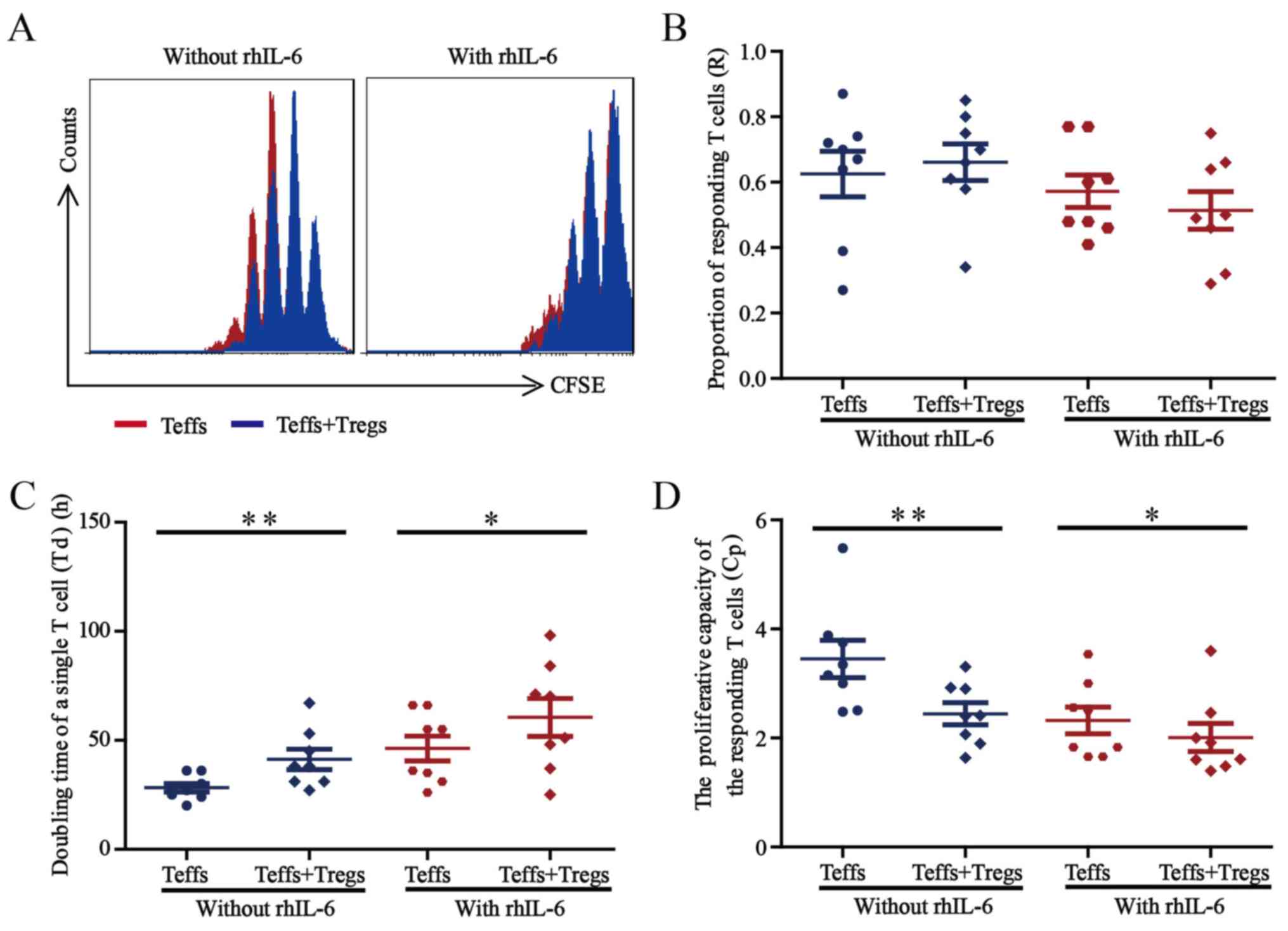 | Figure 6.Suppression of Treg cells under
inflammatory conditions. (A) Representative histograms indicating
the CFSE dilution from a sample cultured without rhIL-6 and a
sample cultured with 25 ng/ml rhIL-6. The proliferation curve of
Teff cells cultured alone (red) was overlaid with the curve for
Teff cells co-cultured with Treg cells (blue). (B-D) The sorted
Teff cells from healthy blood donors (n=8) were labelled with CFSE
and co-cultured with sorted Treg cells (Teff cells/Treg cells, 2:1)
in the presence of anti-CD3/CD28 beads (Teff cells/beads, 1:1),
without or with 25 ng/ml rhIL-6. After co-culture for 5 days,
Teff-cell proliferation was assessed by flow cytometry, and (B) the
proportion of responding T-cells, (C) doubling time of a single
T-cell and (D) proliferative capacity for the Teff cells were
calculated. Paired Student's t-test was used to compare differences
between groups without and with rhIL-6. Values are expressed as the
mean ± standard error of the mean (n=8). *P<0.05 and **P<0.01
compared the doubling time of a single T-cell and proliferative
capacity of Teff cells cultured for 5 days without vs. with Treg
cells. Treg cells, T-regulatory cells; Teff cells, T effector
cells; CFSE, 5,6-carboxyfluorescein succinimidyl ester; rhIL,
recombinant human interleukin. |
The representative overlaid proliferation curve
comparing Teff cells cultured alone and Teff cells co-cultured with
Treg cells are presented in Fig. 6A,
the proliferation of Teff cells was inhibited when co-cultured with
Treg cells, independent of rhIL-6 presence (Fig. 6A). As presented in Fig. 6B, the proportion of the precursor
sample pool that responded to stimulation by dividing (R-value) did
not change significantly when the Teff cells were co-cultured with
Treg cells, regardless of rhIL-6 presence. In terms of the time
required for the average responding T cell to achieve a single cell
division the (Td value) shown in Fig.
6C, co-culture of Teff cells with Treg cells led to a
significantly prolonged Td (P<0.05) in a manner that was
independent of rhIL-6 presence. Regarding the proliferative
capacity (Cp value) showed in Fig.
6D, co-culture with Treg cells led to a significantly decreased
Cp (P<0.05) independent of rhIL-6 presence.
These results demonstrated that Treg cells exert
suppressive effects on Teff cells in the presence or absence of
rhIL-6. However, the inhibitory function of Treg cells may weaken
in the presence of rhIL-6, which further suggests that increased
numbers of Treg cells should be used for treatment considering the
complexity of the internal environment of the patient.
Discussion
As an important inhibitory immune cell subset in
vivo, Treg cells have great potential in the immunotherapy of
various diseases, including autoimmune diseases (7–10).
Considering the complex internal environment of patients, e.g.
inflammatory conditions, the stability and inhibitory function of
Treg cells under such conditions should be further
investigated.
Transplanting a sufficiently large number of Treg
cells is a key factor for successful immunotherapy of diseases,
including autoimmune diseases. Although Treg cells may be easily
cultured and amplified in vitro in the presence of IL-2 and
anti-CD3/CD8 beads (19,23,28,29), the
survival of Treg cells may be affected by inflammatory cytokines,
including IL-6, in vivo (30–33). In
the present study, the number of Treg cells cultured in the
presence of 25 ng/ml rhIL-6 for 14 days was reduced by 49.7% when
compared with that of cells cultured without rhIL-6, demonstrating
the influence of the environment on Treg cell survival. Therefore,
an increased number of Treg cells may be required for immunotherapy
in patients with autoimmune diseases.
A sufficient number of Treg cells is the foundation
of immunotherapy; more importantly, the stability and activity of
Treg cells are directly associated with the effectiveness of
treatment (34). In the present
study, 56.0 or 24.7% of Treg cells expressed the transcription
factor FoxP3 after in vitro culture for 14 days without or
with rhIL-6, respectively. These results indicate that Treg cells
are a relatively stable cell subset, but their stability declined
significantly under inflammatory conditions, further suggesting
that for immunotherapy, transplantation of increased numbers of
Treg cells is required.
The expression of CD39 on the Treg cell membrane
suggests increased activity of Treg cells (25,35). In
the present study, the proportion of CD39 expressed by
FoxP3+Treg cells did not change with culture time, even
in the presence of the inflammatory cytokine rhIL-6. These results
suggest that FoxP3+ Treg cells may maintain their
activity in an inflammatory environment. This result is also
consistent with that of a previous study, which indicated that
CD39+ Treg cells have a strong suppressive function even
in the presence of inflammatory cytokines in vitro (36).
Suppression of Teff cell proliferation is an
important aspect of the inhibitory function of Treg cells. In the
present study, Treg cells suppressed the proliferation of Teff
cells, which is consistent with the previous results regarding the
stability and activity of Treg cells cultured for 5 days. A
decreasing trend regarding Treg cell suppression was observed in
the presence of rhIL-6. The inhibitory function of Treg cells is
directly linked to the treatment effect of Treg cell-based
immunotherapy.
The present study systematically examined the
absolute count, stability, activity and inhibitory function of Treg
cells, and possible changes in Treg cell function in the presence
of cytokines that simulate inflammatory environments were
determined in vivo. To achieve good efficacy,
transplantation of increased numbers of Treg cells may be a
reasonable therapeutic approach for immunotherapy. The present
study provides an important experimental basis for the development
of Treg cell-based immunotherapy for various immune-associated
diseases, including autoimmune diseases.
Acknowledgements
The authors would like to thank The Youth Innovation
Team of Shaanxi Universities for their support.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81471415), the
Natural Science Basic Research Plan in Shaanxi Province of China
(program no. 2018JQ8043 and 2018ZDXM-SF-040), the Scientific
Research Fund of Shaanxi Provincial Education Department (grant no.
18JS104) and the Research Foundation of Xi'an Medical University
(grant nos. 12FZ28, 2017DOC14, 2017GJFY30 and 2016DOC28).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
HG, LX and XG conceived the project and designed the
present study. HG, RZ, FH and JL performed the experiments, data
analysis and wrote the manuscript. HG, XW and KL performed data
analysis and revised the manuscript. LX and XG supervised the work,
provided administrative support, performed data analysis and
proofread the paper.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Xi'an Medical University (Xi'an, China). According to the
principles of informed consent, all healthy blood donors provided
this prior to collection of peripheral blood samples. All of the
experiments of the present study were performed in accordance with
the relevant guidelines and regulations.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Togashi Y and Nishikawa H: Regulatory T
cells: Molecular and cellular basis for immunoregulation. Curr Top
Microbiol Immunol. 410:3–27. 2017.PubMed/NCBI
|
|
2
|
Vignali DA, Collison LW and Workman CJ:
How regulatory T cells work. Nat Rev Immunol. 8:523–532. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Collison LW, Chaturvedi V, Henderson AL,
Giacomin PR, Guy C, Bankoti J, Finkelstein D, Forbes K, Workman CJ,
Brown SA, et al: IL-35-mediated induction of a potent regulatory T
cell population. Nat Immunol. 11:1093–1101. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo H, Xun L, Zhang R and Gou X: Ratio of
CD147high/CD147low in
CD4+CD25+ T cells: A potential biomarker for
early diagnosis and prediction of response to therapy for
autoimmune diseases. Med Hypotheses. 115:1–4. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Talaat RM, Mohamed SF, Bassyouni IH and
Raouf AA: Th1/Th2/Th17/Treg cytokine imbalance in systemic lupus
erythematosus (SLE) patients: Correlation with disease activity.
Cytokine. 72:146–153. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bluestone JA, Buckner JH, Fitch M,
Gitelman SE, Gupta S, Hellerstein MK, Herold KC, Lares A, Lee MR,
Li K, et al: Type 1 diabetes immunotherapy using polyclonal
regulatory T cells. Sci Transl Med. 7:315ra1892015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ou HX, Guo BB, Liu Q, Li YK, Yang Z, Feng
WJ and Mo ZC: Regulatory T cells as a new therapeutic target for
atherosclerosis. Acta Pharmacol Sin. 39:1249–1258. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Martin-Moreno PL, Tripathi S and
Chandraker A: Regulatory T cells and kidney transplantation. Clin J
Am Soc Nephrol. 13:1760–1764. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qiu R, Zhou L, Ma Y, Zhou L, Liang T, Shi
L, Long J and Yuan D: Regulatory T cell plasticity and stability
and autoimmune diseases. Clin Rev Allergy Immunol. Nov
17–2018.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Alvarez C, Rojas C, Rojas L, Cafferata EA,
Monasterio G and Vernal R: Regulatory T lymphocytes in
periodontitis: A translational view. Mediators Inflamm 2018.
78069122018.
|
|
11
|
Sakaguchi S, Sakaguchi N, Asano M, Itoh M
and Toda M: Pillars article: Immunologic self-tolerance maintained
by activated T cells expressing IL-2 receptor α-chains (CD25).
Breakdown of a single mechanism of self-tolerance causes various
autoimmune diseases. J. Immunol. 1995. J Immunol. 186:3808–3821.
2011.PubMed/NCBI
|
|
12
|
Jeffery HC, Braitch MK, Brown S and Oo YH:
Clinical potential of regulatory T cell therapy in liver diseases:
An overview and current perspectives. Front Immunol. 7:3342016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lam AJ, Hoeppli RE and Levings MK:
Harnessing advances in T regulatory cell biology for cellular
therapy in transplantation. Transplantation. 101:2277–2287. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ait-Oufella H, Taleb S, Mallat Z and
Tedgui A: Recent advances on the role of cytokines in
atherosclerosis. Arterioscler Thromb Vasc Biol. 31:969–979. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fatkhullina AR, Peshkova IO and Koltsova
EK: The role of cytokines in the development of atherosclerosis.
Biochemistry (Mosc). 81:1358–1370. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Badal D, Kumar R, Paul M, Dayal D,
Bhansali A, Bhadada SK, Kumar R and Sachdeva N: Peripheral blood
mononuclear cells of patients with latent autoimmune diabetes
secrete higher levels of pro- & anti-inflammatory cytokines
compared to those with type-1 diabetes mellitus following in vitro
stimulation with β-cell autoantigens. Indian J Med Res.
145:767–776. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yao Y, Wang JB, Xin MM, Li H, Liu B, Wang
LL, Wang LQ and Zhao L: Balance between inflammatory and regulatory
cytokines in systemic lupus erythematosus. Genet Mol Res. 15:2016.
View Article : Google Scholar
|
|
18
|
He X, Smeets RL, van Rijssen E, Boots AM,
Joosten I and Koenen HJ: Single CD28 stimulation induces stable and
polyclonal expansion of human regulatory T cells. Sci Rep.
7:430032017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fan MY, Low JS, Tanimine N, Finn KK,
Priyadharshini B, Germana SK, Kaech SM and Turka LA: Differential
roles of IL-2 signaling in developing versus mature tregs. Cell
Rep. 25:1204–1213.e4. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Baecher-Allan C, Brown JA, Freeman GJ and
Hafler DA: CD4+CD25high regulatory cells in human peripheral blood.
J Immunol. 167:1245–1253. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rubtsov YP, Niec RE, Josefowicz S, Li L,
Darce J, Mathis D, Benoist C and Rudensky AY: Stability of the
regulatory T cell lineage in vivo. Science. 329:1667–1671. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
de la Rosa M, Rutz S, Dorninger H and
Scheffold A: Interleukin-2 is essential for CD4+CD25+ regulatory T
cell function. Eur J Immunol. 34:2480–2488. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Levine AG, Arvey A, Jin W and Rudensky AY:
Continuous requirement for the TCR in regulatory T cell function.
Nat Immunol. 15:1070–1078. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guo H, Zheng M, Zhang K, Yang F, Zhang X,
Han Q, Chen ZN and Zhu P: Functional defects in CD4+ CD25high
FoxP3+ regulatory cells in ankylosing spondylitis. Sci Rep.
6:375592016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wells AD, Gudmundsdottir H and Turka LA:
Following the fate of individual T cells throughout activation and
clonal expansion. Signals from T cell receptor and CD28
differentially regulate the induction and duration of a
proliferative response. J Clin Invest. 100:3173–3183. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gudmundsdottir H, Wells AD and Turka LA:
Dynamics and requirements of T cell clonal expansion in vivo at the
single-cell level: Effector function is linked to proliferative
capacity. J Immunol. 162:5212–5223. 1999.PubMed/NCBI
|
|
27
|
Borsellino G, Kleinewietfeld M, Di Mitri
D, Sternjak A, Diamantini A, Giometto R, Höpner S, Centonze D,
Bernardi G, Dell'Acqua ML, et al: Expression of ectonucleotidase
CD39 by Foxp3+ Treg cells: Hydrolysis of extracellular ATP and
immune suppression. Blood. 110:1225–1232. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fontenot JD, Rasmussen JP, Gavin MA and
Rudensky AY: A function for interleukin 2 in Foxp3-expressing
regulatory T cells. Nat Immunol. 6:1142–1151. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X and Zheng Y: Regulatory T cell
identity: Formation and maintenance. Trends Immunol. 36:344–353.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Passerini L and Bacchetta R:
Forkhead-box-P3 gene transfer in human CD4(+) T conventional cells
for the generation of stable and efficient regulatory T cells,
suitable for immune modulatory therapy. Front Immunol. 8:12822017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ezzelarab MB: Regulatory T cells from
allo- to xenotransplantation: Opportunities and challenges.
Xenotransplantation. 25:e124152018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Min B: Heterogeneity and stability in
Foxp3+ regulatory T cells. J Interferon Cytokine Res. 37:386–397.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tanaka T, Narazaki M and Kishimoto T: IL-6
in inflammation, immunity, and disease. Cold Spring Harb Perspect
Biol. 6:a0162952014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wiesinger M, Stoica D, Roessner S, Lorenz
C, Fischer A, Atreya R, Neufert CF, Atreya I, Scheffold A,
Schuler-Thurner B, et al: Good manufacturing practice-compliant
production and lot-release of ex vivo expanded regulatory T cells
as basis for treatment of patients with autoimmune and inflammatory
disorders. Front Immunol. 8:13712017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fletcher JM, Lonergan R, Costelloe L,
Kinsella K, Moran B, O'Farrelly C, Tubridy N and Mills KH:
CD39+Foxp3+ regulatory T cells suppress pathogenic Th17 cells and
are impaired in multiple sclerosis. J Immunol. 183:7602–7610. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gu J, Ni X, Pan X, Lu H, Lu Y, Zhao J, Guo
Zheng S, Hippen KL, Wang X and Lu L: Human CD39(hi) regulatory T
cells present stronger stability and function under inflammatory
conditions. Cell Mol Immunol. 14:521–528. 2017. View Article : Google Scholar : PubMed/NCBI
|