Introduction
Acute respiratory distress syndrome (ARDS), a common
cause of death in intensive care units (1,2), is a
devastating clinical syndrome characterized by non-cardiogenic
pulmonary edema, respiratory distress and hypoxemia (3–5). There
are ~190,000 newly diagnosed ARDS cases in the United States each
year (6). Although a great deal of
progress has been made in ARDS management in previous years, the
mortality rate caused by ARDS is still as high as 35–50% (7,8), while
the survivors suffer from significant physical and psychological
impairments (3–5).
Hypercoagulation and fibrinolysis inhibition,
existing either systemically or locally, are important
characteristics in the pathogenesis of ARDS (9,10).
However abnormalities of coagulation and fibrinolysis are more
obvious in the alveolar space than in systemic circulation,
demonstrated by much higher levels of tissue factor (TF) and
plasminogen activator inhibitor (PAI) in airspaces than in the
blood, and the numerous fibrin deposits seen in the alveolar
compartment in ARDS (11–14). Coagulation and fibrinolysis
dysfunctions in local pulmonary tissue lead to reduced lung
compliance, diffusion dysfunction, and disruption of V/Q ratios,
resulting in refractory hypoxemia, very small lung volumes (baby
lungs) and even pulmonary fibrosis. Lung epithelial cells have been
confirmed to be the primary source of TF, contributing 60–70% of
the total lung TF (15) and alveolar
epithelial cells (AEC II) have been shown to express a large number
of TF and PAI-1 when stimulated (16,17),
indicating that AEC II has a pivotal role in regulating coagulation
and fibrinolysis in airspaces, via expressing TF and PAI-1.
However, until now, the specific regulatory mechanism of AEC II
still remains to be elucidated.
Nuclear factor (NF)-κB is an evolutionarily
conserved family of DNA binding proteins involved in
transcriptional regulation of a number of gene products, including
inflammation and apoptosis. Under normal conditions, NF-κB is
sequestered in the cytoplasm, bound by members of the IkB family of
inhibitor proteins, which include IkBα, IkBβ and IkBε. IκB kinase
(IKK) complex, containing IKKα, IKKβ and IKKγ, are important
molecules that initiate the NF-κB cascade activation by promoting
IκB phosphorylation (18) and
liberating NF-κB from the combined state. NF-κB activation could be
initiated by canonical and non-canonical pathway, among which the
former is the main form, and IKKβ is the essential upstream signal
in activation of NF-κB canonical pathway. Moreover, studies have
shown that IKKβ is a major kinase that activates NF-κB activation
induced by proinflammatory cytokines and IKKβ may be more important
in NF-κB activation pathway than IKKα (19–21). The
NF-κB pathway has been shown to play a key role in inflammatory
processes, angiogenesis, immunity and apoptosis (22). In addition, previous studies in
vitro or in vivo have demonstrated that the NF-κB
pathway was also involved in regulating coagulation and
fibrinolytic factors (23–26). Ding et al (27) reported that inhibiting Rho kinase (an
upstream site of NF-κB signal pathway) significantly reduced the
lung tissue inflammatory response and lung TF and PAI-1 levels by
blocking the NF-κB pathway. Since IKKβ, just like Rho kinase is
also an essential upstream molecule of NF-κB pathway, the present
study speculated that adjusting IKKβ gene expression could impact
the expression of coagulation and fibrinolysis factors in
LPS-stimulated AEC II. To confirm this hypothesis,
IKKβ+/+ and IKKβ−/− AEC II models were first
set up using lentiviral vector cell transfection and then observed
whether coagulation, and fibrinolysis factors in LPS-stimulated AEC
II would be changed during IKKβ gene up- or downregulation.
Materials and methods
Cell culture and LPS stimulation
The cell line used for lentivirus vector
transfection in the experiment was the RLE-6TN cell line (The Cell
Bank of Xiangya Medical College; ACE II cell line from rats). This
cell line was grown in M199 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum
(Hyclone; SH30070.03), penicillin (10,000 U/ml) and streptomycin
(10,000 U/ml) (Hyclone; SV 30010). Cells were cultured in an
incubator at 37°C and 5% CO2. The cells in the control
group were not manipulated. The cells in short-hairpin
(sh)-negative control (NC) group were infected using a negative
control viral plasmid, while the cells in sh-IKKβ group were
infected by IKKβ shRNA interference (20 µl of virus solution per
well) (RNAi) virus. Cells in the NC group were infected by empty
pcDNA3.1 virus (Hunan Fenghui Biotechnology Co., Ltd; 0 µl of virus
solution per well) and cells in the IKKβ group were infected with
the pcDNA3.1-IKKβ overexpression virus. Cells except the control
group were all stimulated with LPS at a concentration of 50 µg/ml
for 24 h.
Construction of IKKβ+/+ and
IKKβ−/− model by virus transfection
Based on the IKKβ gene (NM_053355), the shRNA
sequence was designed (Table I) for
the IKKβ gene and a negative control was designed that was verified
by BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) to have no
interference effect on other genes. Then based on the small
interfering (si)RNA sequences, complementary single-stranded DNA
was designed (Table II).
 | Table I.The short hairpin RNA sequence for
the IKKβ gene and the sequence of negative control. |
Table I.
The short hairpin RNA sequence for
the IKKβ gene and the sequence of negative control.
| Negative |
5′-GCCTTATTTCTATCTTACGtt-3′ |
| siRNA |
5′-GCACAATCAGGTGACAGGTtt-3′ |
| Table II.The shRNA sequence for the
complementary single-stranded DNA. |
Table II.
The shRNA sequence for the
complementary single-stranded DNA.
| IKKβ | F:
5′-GTACCTCGCACAATCAGGTGACAGGTTCAAGAGACCTGTCACCTGATTGTGCTTTTTGGAAA-3′ |
|
|
R-5′AGCTTTTCCAAAAAGCACAATCAGGTGACAGGTCTCTTGAACCTGTCACCTGATTGTGCGAG-3′ |
| NC | F:
5′-GTACCTCGCCTTATTTCTATCTTACGTCAAGAGCGTAAGATAGAAATAAGGCTTTTTGGAAA-3′ |
|
| R:
5′-AGCTTTTCCAAAAAGCCTTATTTCTATCTTACGCTCTTGACGTAAGATAGAAATAAGGCGAG-3′ |
The lentivirus vector plasmids used in this study
are pcDNA3.1+ and pLKO.1 (Tiangen Biotech Co., Ltd), among which
the interference and overexpression sequence were constructed in
pLKO.1 and in pcDNA3.1+ plasmid respectively. Plasmids were
amplified in Escherichia coli., followed by the lentivirus
packaging. RLE-6TN cells (1×106/well, 20 µl of virus
solution per well) were infected with lentivirus which carried
IKKβ-shRNA and IKKβ overexpression, by which the stable expression
of sh-IKKβ and IKKβ overexpression was obtained. RLE-6TN cells
infected by the virus alone was used as control.
Reverse transcription-quantitative
(RT-q)PCR assay
The mRNA expression of IKKβ, p65, TF and PAI-1 was
detected by qPCR. GAPDH was used as internal reference. Briefly,
cells were collected after 48 h of virus transfection and total RNA
was extracted using Trizol® (Takara Bio, Inc.; cat. no.
9108), and then the concentration of the total mRNA was assessed
using the NanoDrop2000 Spectrophotometer (NanoDrop Technologies;
Thermo Fisher Scientific, Inc.). The A260/A280 ratio of the
extracted RNA was adjusted to be 1.8–2.0, then reverse
transcription was performed on 2 µg RNA with oligo (dT) primers in
20 µl reactions using the RevertAid First Strand cDNA Synthesis kit
(Thermo Fisher Scientific, Inc.; K1622) according to the
manufacturer's protocol. Primers were designed according to the
sequence of IKKβ gene of rat in the NCBI gene database (https://www.ncbi.nlm.nih.gov/gene/84351). The primer
sequences used were as follows: GADPH forward,
5′-GGGAAACCCATCACCATCTT-3′ and reverse,
5′-CCAGTAGACTCCACGACATACT-3′; IKKβ forward,
5′-GTGACATAGCATCGGCTCTTAG-3′ and reverse,
5′-CTCTCCTTGCTGTAGGACAATG-3′; NF-κB p65 forward,
5′-CATGCGTTTCCGTTACAAGTG-3′ and reverse,
5′-CCCGTGTAGCCATTGATCTT-3′; TF forward, 5′-CCTCCAGGGAAAGCGTTTAAT-3′
and reverse, 5′-GTGTAGGTATAGTTGGTGGGTTTC-3′; PAI-1 forward,
5′-GCCACCAACTTCGGAGTAAA-3′ and reverse,
5′-GTAGGGAGAGAAGACCACATTTC-3′. PCR amplification was performed
using the cDNA as template. The temperature protocol was as
follows: 95°C for 10 min, heating for 95°C for 5 sec, 60°C for 1
min for 40 cycles, 95°C for 15 sec, 60°C for 1 min and 95°C for 15
sec. The reaction system was set up as follows: SYBR Green Mix
(cat. no. RR820A; Takara Bio, Inc.) 10 µl, forward primer and
reverse primer 0.4 µl respectively, cDNA template 2 µl,
ddH2O 7.2 µl, which were made up into a system
containing 20 µl reagents. The dissolution and amplification curve
of the genes were recorded following the gene amplification. The
specificity of the reaction was evaluated and the Cq value was
calculated according to the dissolution and amplification curve,
respectively. Expression of target genes was calculated using the
2−∆∆Cq method, ∆∆Cq=(Cq, target-Cq,
GAPDH) sample-(Cq, target-Cq,
GAPDH) control (28).
Western blotting
After being transfected with virus and then being
treated with LPS for 24 h, the cells were washed with cold PBS. The
total protein was extracted with RIPA (Hunan Fenghui Biotechnology
Co., Ltd). Briefly, concentration of protein was measured with a
BCA assay kit according to the manufacturer's protocol. An equal
amount of protein (30 mg of the protein solution) from each sample
was resolved in Tris-glycine 10% SDS-PAGE. Protein bands were
blotted onto nitrocellulose membranes. At the end of the membrane
transferring, the membrane was soaked from the bottom to the top
with TBS, and then transferred to a dish containing the blocking
solution (blocking solution: 5% skim milk powder diluted with TBST
solution), and shaken for 3 h at room temperature on a shaker.
The membrane was incubated for 24 h with the
antibody of rabbit anti-rat IKKβ (1:1,000; cat. no. ab124957;
Abcam), p-IKKβ (1:1,000; cat. no. ab194519; Abcam), p65 (1:1,000;
cat. no. ab16502; Abcam), p-p65 (1:1,000; cat. no. ab86299; Abcam),
IκBα (1:1,000; cat. no. ab32518; Abcam), p-IκBα (Cell Signaling
Technology, Inc; 1:1,000; cat. no. 9241; Abcam), TF (1:1,000; cat.
no. ab151748; Abcam) and PAI-1 (1:1,000; cat. no. ab66705; Abcam)
at 4°C. The secondary antibody (horseradish peroxidase-conjugated
goat ant-rabbit immunoglobulin; 1:5,000; cat. no. ZB-2301;
ZSGB-BIO) was added and incubated with horseradish blocking
solution for 10 min at room temperature, the membrane
chemiluminescence detection system (EMD Millipore). Relative band
densities were quantified by Image J software 1.4.3 (National
Institutes of Health).
Detection of TF procoagulant
activity
TF procoagulant activity was performed using split
RLE-6TN cells for a one-step recalcification clot time assay
(29).
ELISA assay
Cell supernatants were harvested and stored at
−80°C. Thrombin antithrombin (TAT) (Cusabio Biotech Co., Ltd; cat.
no. CSB-E08432r), antithrombin III (ATIII) (Cusabio Biotech Co.,
Ltd; cat. no. CSB-E13885r), procollagen III propeptide (PIIIP)
(Cusabio Biotech Co., Ltd; cat. no. CSB-E08096r), thrombomodulin
(TM) (Cusabio Biotech Co., Ltd; cat. no. CSB-E07939r) and PAI-1
(Cloud-Clone Corp; cat. no. SEA532Ra) levels in cell supernatants
were determined by ELISA according to the manufacturer's
protocol.
Immunofluorescence
Briefly, cell each group was fixed at room
temperature with 4% formaldehyde in PBS for the first 30-min and
then permeabilized with 0.5% Triton X-100 for another 30-min,
followed by a third blocking step of 30 min with 1% bovine serum
albumin. After that, these cells were incubated with primary rabbit
antibody against rat p65 and IKKβ (1:100; cat. no. ab16502; Abcam)
overnight at 4°C. And next, they were incubated with fluorescein
isothiocyanate-labeled secondary antibody (OriGene Technologies,
Inc.) for 1 h at room temperature. Each step was followed with
5-min of washes in PBS three times. The prepared specimens were
counterstained with DAPI for 10 min at room temperature and
observed with a fluorescence microscope (Carl Zeiss AG) and were
captured under an original magnification of ×20.
Statistical analysis
Data are expressed as mean ± standard deviation.
Statistical significance was determined using one-way analysis of
variance (ANOVA) and Student-Newman-Keuls method (SPSS 17.0; SPSS,
Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
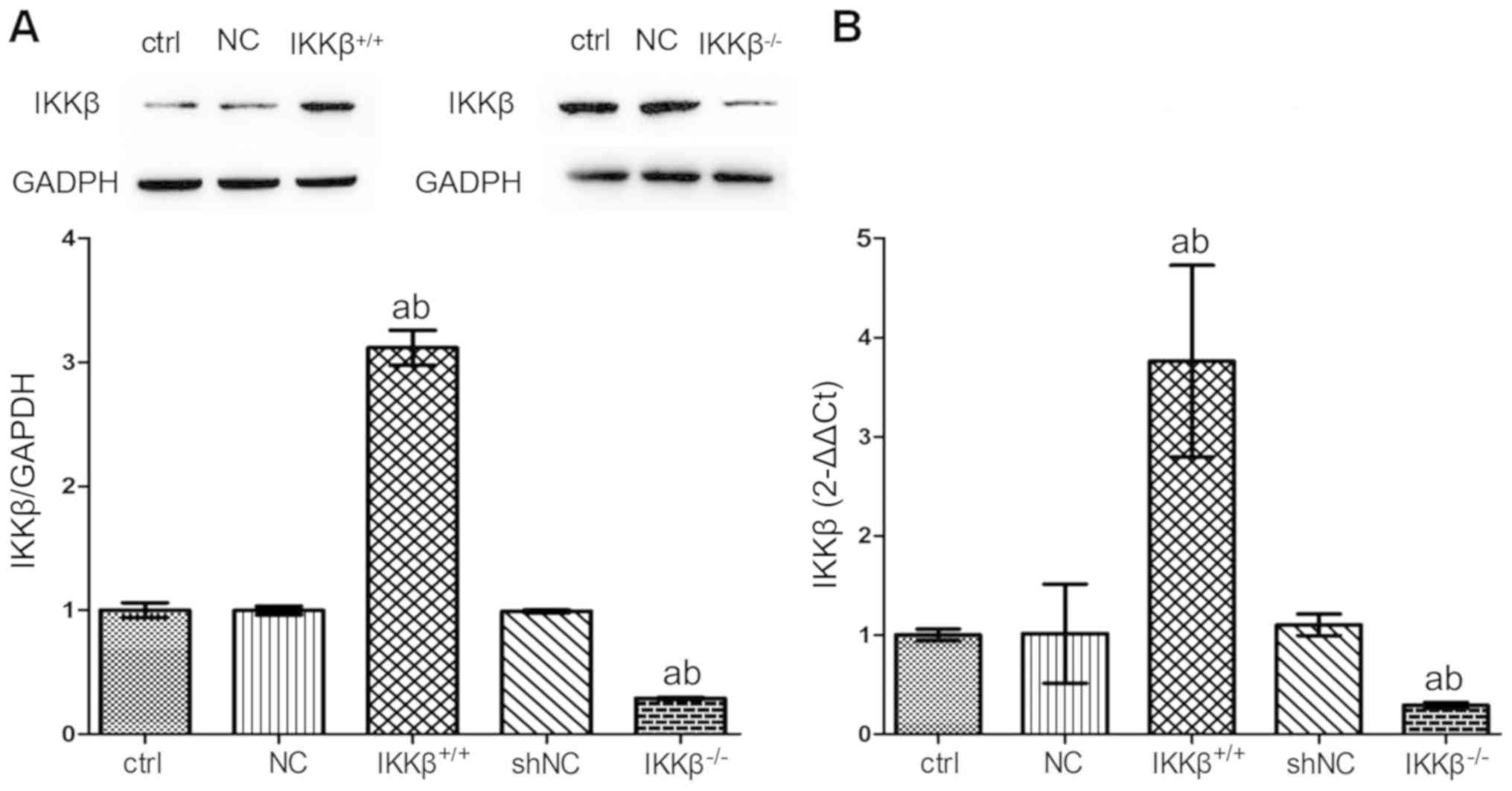
IKKβ+/+ and
IKKβ−/− cell models are replicated successfully by virus
transfection
The IKKβ gene overexpression and low expression
models were set up by the virus transfection technique. The protein
and mRNA levels of IKKβ were screened to verify the success of the
transfection. Expression of mRNA and protein in IKKβ+/+
cells were significantly increased, and the expression in
IKKβ−/− cells were significantly decreased in wild type
(WT) cells, respectively (P<0.05; Fig. 1), indicating the success of
transfection.
Genetic changes in IKKβ can affect the
expression of TF and PAI-1 in LPS-stimulated AEC II
To observe the impact of IKKβ gene level on
expression of TF and PAI-1 in condition of LPS injury, cells were
stimulated with different levels of the IKKβ gene using LPS for 24
h. Results showed that expression of TF and PAI-1, either in mRNA
or in protein, were all significantly upregulated in WT cell
following 24-h of LPS stimulation (P<0.05). In
IKKβ+/+ cells, however, the expression was further
enhanced by LPS stimulation, while TF and PAI-1 expression in
IKKβ−/− cells was inhibited in spite of LPS stimulation
(Fig. 2).
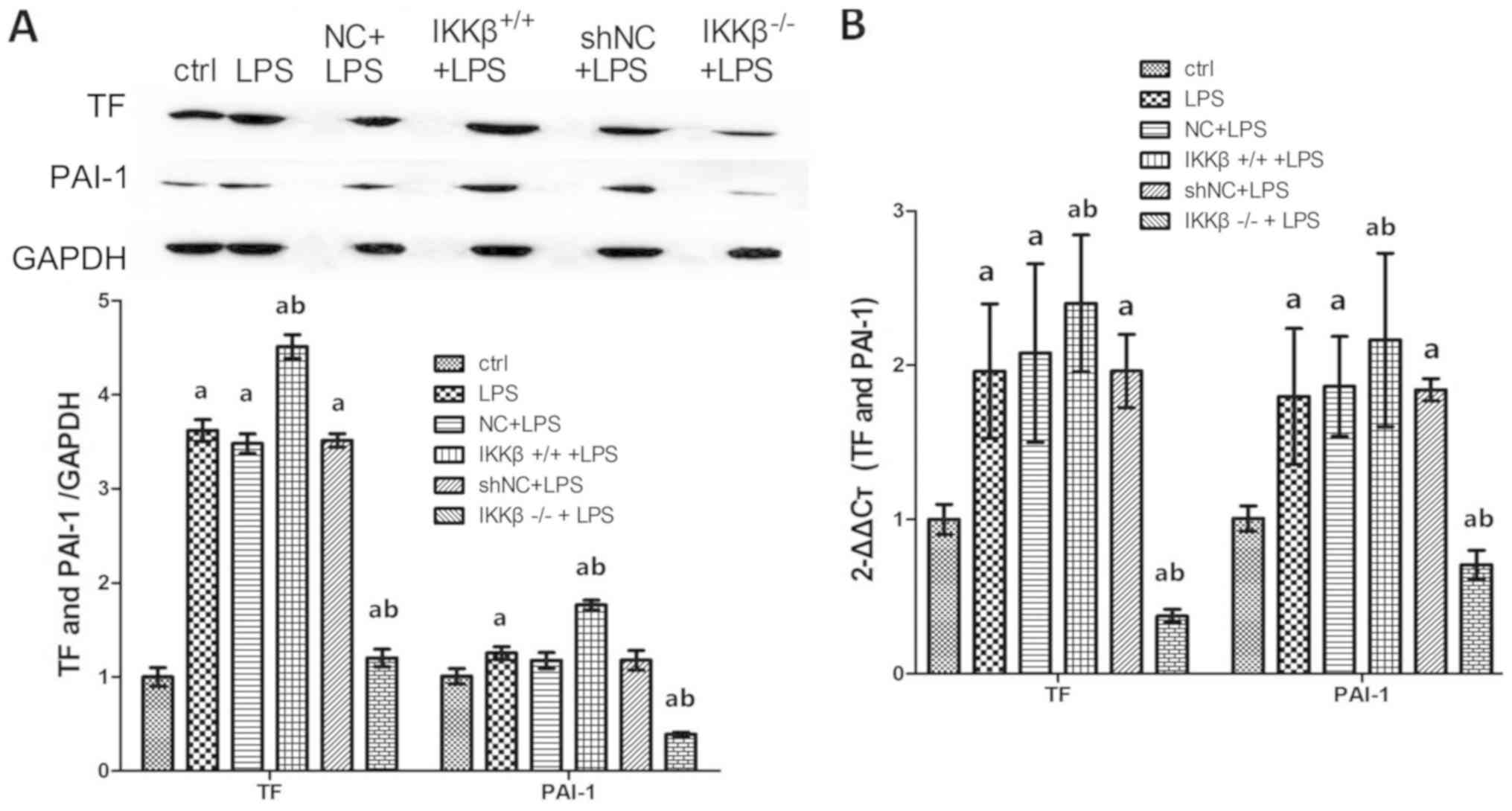 | Figure 2.Over-expression of IKKβ promotes,
low-expression of IKKβ inhibits the expression of TF and PAI-1 in
LPS-stimulated AECII. (A) Western blotting was performed to measure
TF and PAI-1protein expression in cells, GAPDH was used as an
internal control for protein reference. Some bands appear obscured
as some unnecessary groups, which were added at the beginning of
the experiment, were removed. (B) Reverse
transcription-quantitative-PCR was performed to analyze expression
TF and PAI-1 mRNA in cells. Each bar represents the mean ± standard
deviation of 3 groups of cells. aP<0.05 vs. ctrl.
bP<0.05 vs. LPS. LPS, lipopolysaccharide; AEC II,
alveolar epithelial cell type II; ctrl, control; TF, tissue factor;
PAI-1, plasminogen activator inhibitor; NC, negative control; sh,
shorthairpin. |
Different IKKβ gene results in
different secretions of ATIII, TAT, PAI-1, TM and PIIIP from
LPS-stimulated AEC II
To further determine the secretion of coagulation
and fibrinolysis related molecules from LPS-treated cells with
different IKKβ gene levels, the concentrations of ATIII, TAT, TM,
PIIIP and PAI-1 were measured. The results of the present study
showed that LPS induction significantly promoted secretions of TAT,
TM, PIIIP and PAI-1 from WT cell compared with WT cells induced by
saline (P<0.05). More secretions of these molecules were
obtained from IKKβ+/+ cells under LPS treatment, but
secretions of TAT, TM, PIIIP and PAI-1 in IKKβ−/− cells
decreased under LPS stimulation compared with WT cells as well as
IKKβ+/+cells. The ATIII level, however, demonstrated the
opposite change when compared with changes of TAT, TM, PIIIP and
PAI-1 (Fig. 3).
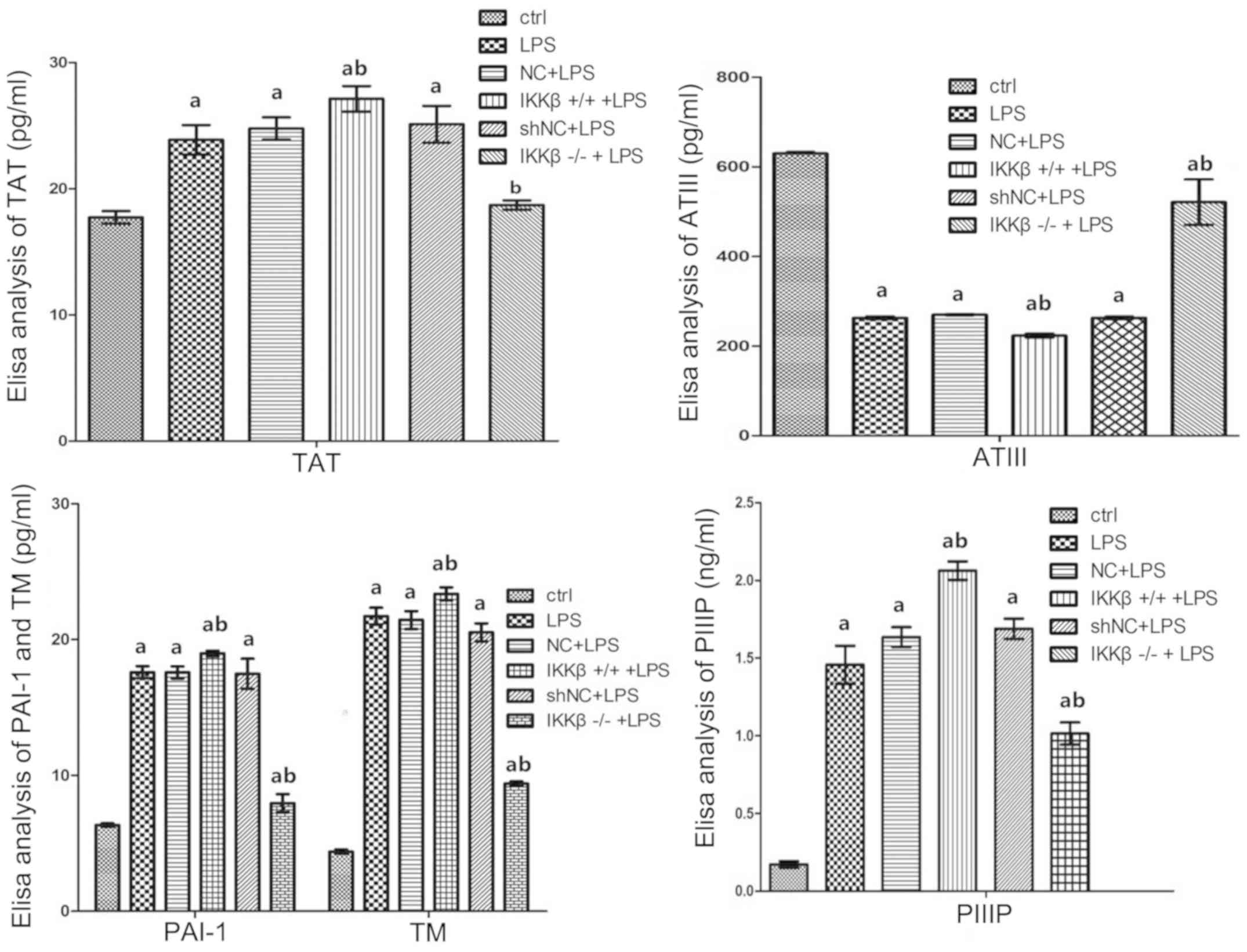 | Figure 3.Over-expression of IKKβ promotes
secretions of TAT, TM, PIIIP and PAI-1, and inhibits ATIII
production, while low-expression has inverse effects on these
molecules from LPS-stimulated AEC II. Bar graphs of concentrations
about the indicators above as assessed by enzyme-linked
immunosorbent assays in cell-supernatant of each group. Values are
presented as the mean ± standard deviation; aP<0.05
vs. the ctrl, bP<0.05 vs. the LPS group as determined
by one-way analysis of variance. TAT, thrombin-antithrombin; ATIII,
antithrombin III; TM, thrombomodulin; PIIIP, procollagen III N
terminal peptide; ctrl, control; AEC II, alveolar epithelial cell
type II; LPS, lipopolysaccharide; NC, negative control; sh, short
hairpin. |
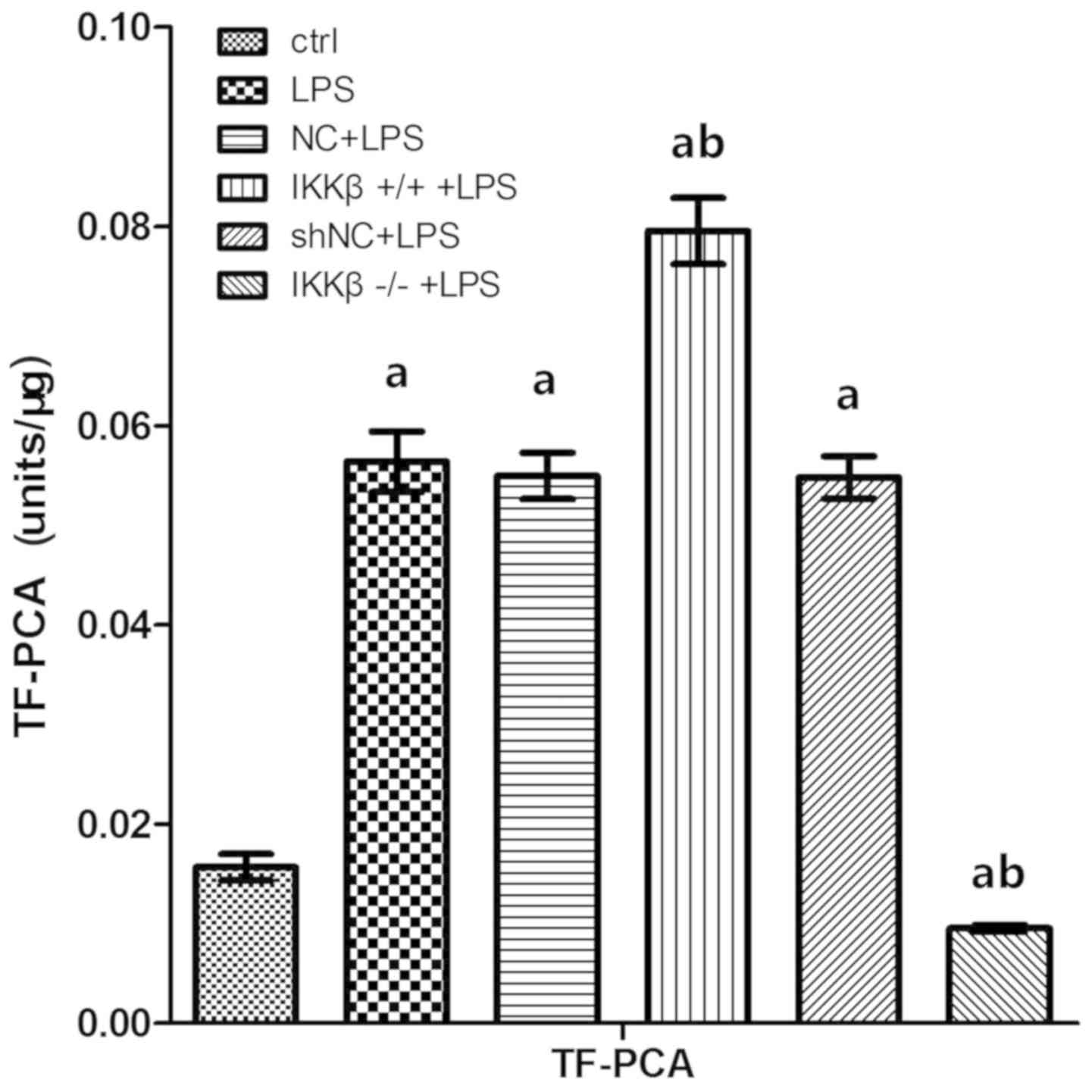
IKKβ gene level impacts LPS-induces TF
procoagulant activity (TF-PCA) in AEC II
TF-PCA in LPS-stimulated AEC II was examined using
ELISA. The results showed that LPS stimulation resulted in
increased TF-PCA in WT cells and TF-PCA was further enhanced in
IKKβ+/+ cells, and but was significantly inhibited in
IKKβ−/− cells as compared with in WT cells (P<0.05;
Fig. 4).
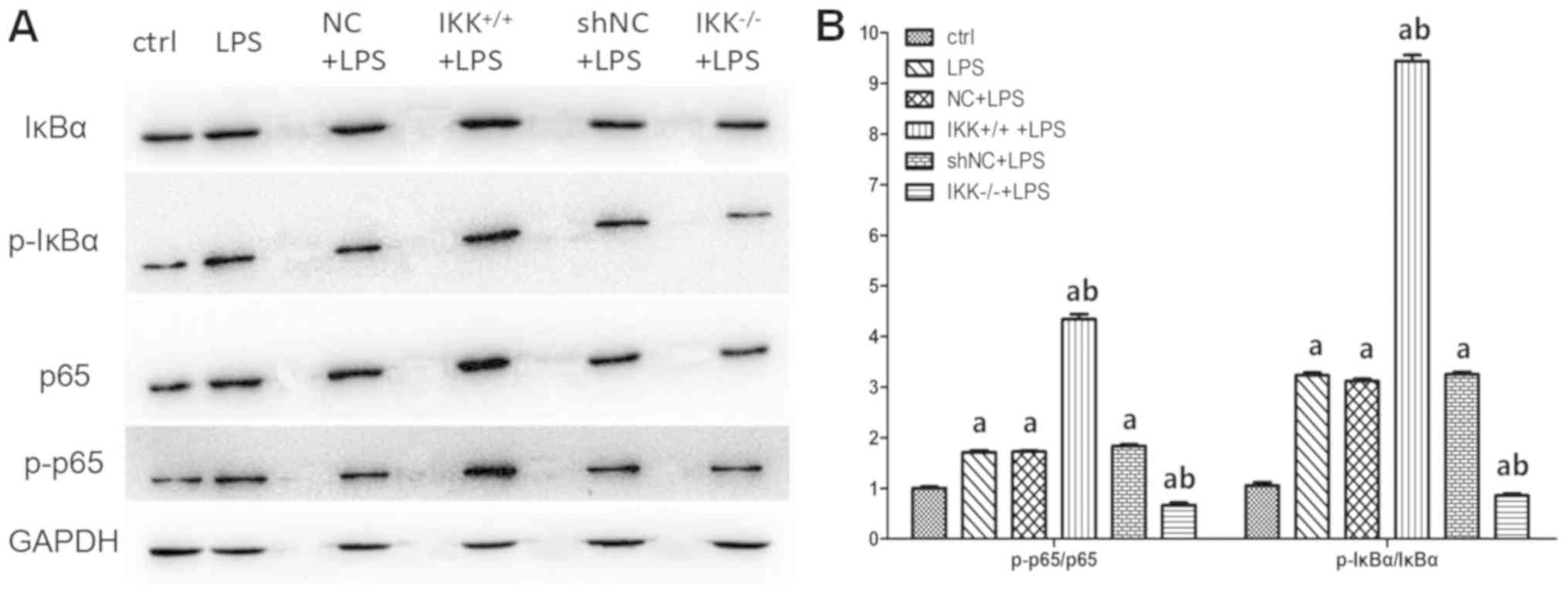
Conditional up-/downregulation of IKKβ
gene affects the NF-κB canonical signaling pathway in AEC II
To explore the mechanism by which the IKKβ gene
impacts coagulation and fibrinolysis related molecules in AEC II
under the condition of LPS treatment, the role of IKKβ gene on
certain important molecules in the NF-κB canonical signaling
pathway in LPS-induced AEC II was observed. The results
demonstrated that conditional IKKβ gene upregulation significantly
improved expression of p65, p-p65, IkB and p-IkB induced by LPS
(P<0.05). However, the expression of these molecules were
significantly inhibited in LPS-treated AEC II if IKKβ gene was
downregulated beforehand (P<0.05; Fig. 5).
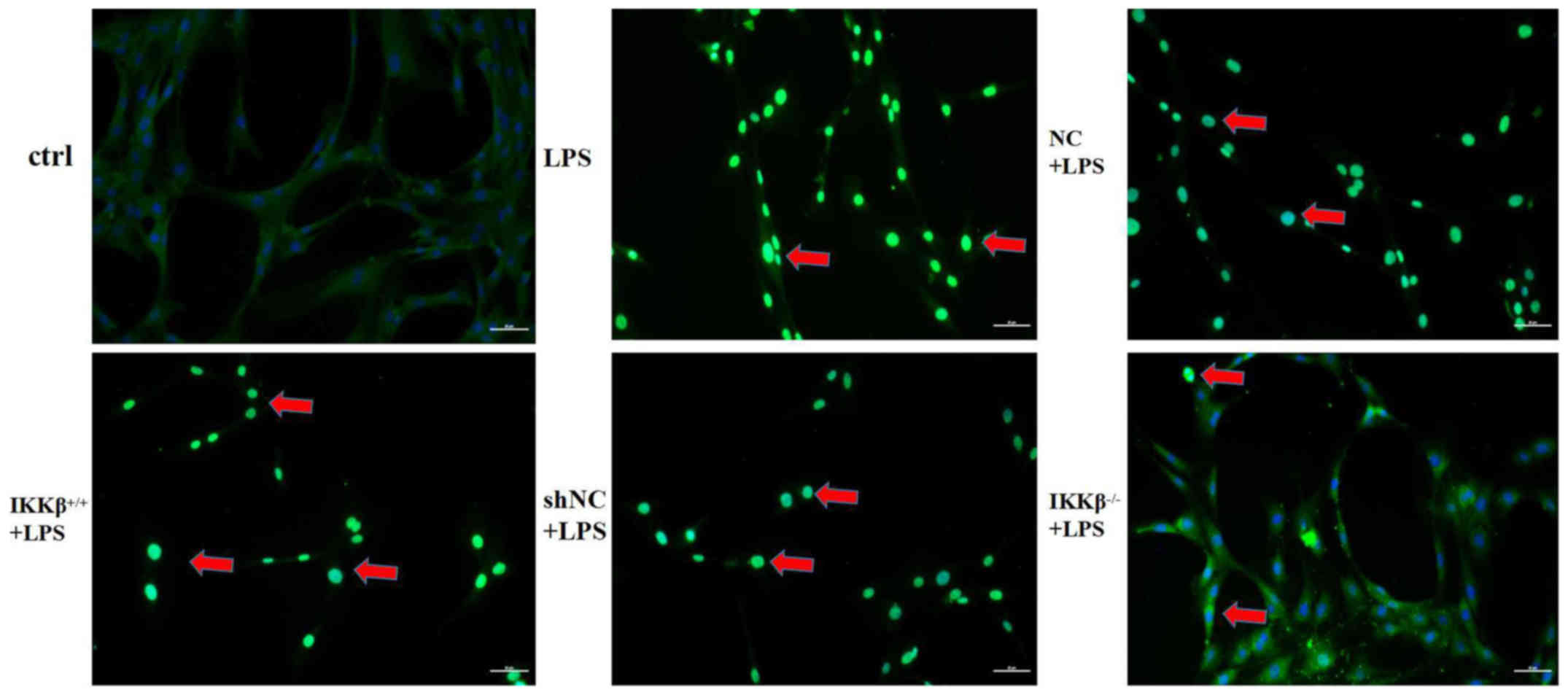
The present study also determined the translocation
ability of p65 from the cytoplasm into the nucleus using an
immunofluorescence assay in the LPS-stimulation state. The staining
results indicated that LPS stimulation resulted in a marked
increase of p65 fluorescent staining in nucleus, indicating
enhanced p65 translocation from the cytoplasm into the nucleus. p65
fluorescent staining was seen to have an additional enhancement in
IKKβ+/+ cells but it was weakened in IKKβ−/−
cells with LPS stimulation (Fig.
6).
Discussion
In biological research, the virus has become an
effective carrier for delivering DNA or RNA into cells.
Lentiviruses can efficiently introduce the target gene or RNAi into
human or animal primary cells or cell lines, so that conditional
up-/ downregulation of the target gene could be set up (30,31). In
the present study, up-/downregulation of IKKβ expression was
achieved through lentivirus transfection therapy. The results of
the present study showed that, as compared with WT cell, a higher
IKKβ and a lower IKKβ gene expression were achieved by target gene
or RNAi transfection respectively, indicating that the
IKKβ+/+ and IKKβ−/− cell models were
successfully set up (32).
Previous studies have shown that the
hypercoagulability and fibrinolysis inhibition in the alveolar
space in ARDS is more prominent than in the systemic vascular
network because the TF levels in the bronchoalveolar lavage fluid
of patients with ARDS are significantly increased compared with in
the plasma (9,10,32,33).
Since TF is a key coagulation factor that initiates the extrinsic
coagulation pathway, it plays an important regulatory role in
abnormal coagulation of ARDS (34),
while PAI-1 is a key factor regulating fibrinolysis inhibition
(35,36). Therefore, TF and PAI-1 were
respectively selected as the coagulation and fibrinolysis factor in
this experiment. The present data showed that both TF and PAI-1 are
highly expressed in WT rat AEC II under LPS stimulation, either at
the mRNA or protein level, which indicates the dysfunction in
coagulation and fibrinolysis under LPS stimulation.
In addition to TF and PAI-1, there are other factors
associated with coagulation and fibrinolysis, such as TAT, ATIII,
PIIIP and TM. TAT, a complex of thrombin and anti-thrombin,
reflects coagulation state, and ATIII is an anticoagulant substance
in the body. TM is a glycoprotein and has anticoagulation
characteristics, whose increase indicates a hypercoagulable state.
PIIIP is an important marker of tissue fibrosis and a high level of
PIIIP is often associated with inhibition of fibrinolysis. The
results of the present study demonstrated that WT cells secreted a
larger amount of TAT, TM and PIIIP and PAI-1, but just produced a
little of ATIII after being induced by LPS compared with being
treated by saline. Combined with the expression changes of TF and
PAI-1, all the changes of these factors indicated hypercoagulation
and fibrinolysis inhibition mediated by LPS-stimulated AEC IIs.
IKKβ is a crucial molecule located at upstream of
the NF-κB canonical signaling pathway which was thought to be
involved in regulating coagulation and fibrinolysis in ARDS
(23–26), and AEC II contributes to most of
alveolar coagulation and fibrinolysis abnormalities by expressing
some important factors such as TF and PAI-1 (16,17,36,37), so
the present study speculated that IKKβ would adjust the expression
of coagulation and fibrinolysis factors. To test this hypothesis,
IKKβ+/+ and IKKβ−/− AEC cell models were set
up first, and then the role of IKKβ on these factors was explored.
The results of the present study demonstrated that under LPS
stimulation, overexpression of IKKβ (IKKβ+/+ cell) not
only boosted TF and PAI-1 expression, but also promoted the
secretion of TAT, TM, PIIIP and PAI-1, and inhibited ATIII
production from the LPS treated AEC II. When IKKβ was downregulated
(IKKβ−/− cells), however, the expression level or amount
of production of all these factors was completely reversed. From
the results of the present study, it is reasonable to think that
IKKβ was involved in the regulation of AEC II-mediated coagulation
and fibrinolysis abnormalities in some pathological circumstances
such as ARDS.
The NF-κB cascade mainly comprises two divergent
signaling pathways; the classical canonical pathway and
non-canonical pathway (38). In the
canonical pathway of NF-κB, stimuli such as LPS stimulates the IκB
kinase (IKKα, IKKβ and IKKγ) and activation of IKKs results in
phosphorylation of the IκBs proteins and degradation of IκBs
follows. Degradation of IκBs proteins reveals nuclear localization
sequences of NF-κB, leading to the rapid translocation of active
NF-κB p65 into the nucleus, where they bind to κB binding sites in
the promoters of target genes, promoting the transcription of these
genes. In an alternative pathway of NF-κB, the non-canonical
pathway however, it is p100 rather than p65 that is activated
(38), although IKKβ is also
involved in the regulation. The figures from the present study
showed that the expression of IκB, p-IκB, p65 and p-p65 changed
with the variation of IKKβ gene level under LPS treatment.
Furthermore, conditional up- or downregulation of IKKβ could
enhance or inhibit p65 translocation from cytoplasm into nucleus,
demonstrated by p65 fluorescent staining in the nucleus. Therefore,
it is estimated that NF-κB canonical cascade is at least one of the
mechanisms by which IKKβ regulates procoagulant and fibrinolytic
inhibitory factors in LPS-stimulated AEC II.
There are some limitations in the present
experiment. First, the condition of this experiment in vitro
is relatively simple as a single cell is assessed and not an entire
body, and it does not necessarily represent the real situation in
the whole body. Second, because of possible toxicity of the virus
and the reagent itself, the technology used could not be used in a
whole body study. Finally, LPS is just one of the causes of ARDS
(bacterial infection), so these results do not stand for ARDS
caused by other causes such as aspiration or severe acute
pancreatitis.
In conclusion, the present study has demonstrated
that upregulation of IKKβ enhanced hypercoagulation and
fibrinolysis inhibition, while conditional deletion of IKKβ has a
completely reversed role on coagulation and fibrinolysis related
factors in LPS-stimulated AEC II. The NF-κB canonical signaling
pathway is at least one of the mechanisms by which IKKβ impacts the
coagulation and fibrinolysis function in LPS-induced AEC II. IKKβ
is expected to be a new target for prevention of coagulation and
fibrinolysis abnormalities in ARDS.
Acknowledgements
Not applicable.
Funding
The funding supporting the present study was
received from The Science and Technology Supportive Plan Project of
Guizhou Province [grant no. (2017) 2876], The Major Research
Project of Innovation Group in Education Department of Guizhou
Province [grant no. (2016) 034] and The Science and Technology
Innovation Project of Overseas Students in Guizhou [grant no.
(2016) 19].
Availability of data and materials
The datasets used and/or analyzed during the preent
study are available from the corresponding author on reasonable
request.
Authors' contributions
BL performed the whole experimental operation,
finished the statistical analysis and wrote the primary manuscript.
YWa, YWu, YC, HQ and HY were mainly responsible for the data
collection. FS conceived the whole design of the study, analyzed
the data and organized the final manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The current study was approved by the Ethics
Committee of Animal and Cell Laboratory of Guizhou Medical
University.
Patient consent for publication
Not applicable.
Conflicts of interest
The authors declare no conflict of interest.
References
|
1
|
Piantadosi CA and Schwartz DA: The acute
respiratory distress syndrome. Ann Intern Med. 141:460–470. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ware LB and Matthay MA: The acute
respiratory distress syndrome. New Eng J Med. 342:1334–1349. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kangelaris KN, Calfee CS, May AK, Zhuo H,
Matthay MA and Ware LB: Is there still a role for the lung injury
score in the era of the berlin definition ARDS? Ann Intensive Care.
4:42014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Khemani RG, Wilson DF, Esteban A and
Ferguson ND: Evaluating the berlin definition in pediatric ARDS.
Intensive Care Med. 39:2213–2216. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thompson BT and Matthay MA: The berlin
definition of ARDS versus pathological evidence of diffuse alveolar
damage. Am J Respir Crit Care Med. 187:675–677. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xiao M, Zhu T, Zhang W, Wang T, Shen YC,
Wan QF and Wen FQ: Emodin ameliorates LPS-induced acute lung
injury, involving the inactivation of NF-κB in mice. Int J Mol Sci.
15:19355–19368. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gotts JE and Matthay MA: Treating ARDS:
New hope for a tough problem. Lancet Respir Med. 2:84–85. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ferguson ND, Fan E, Camporota L, Antonelli
M, Anzueto A, Beale R, Brochard L, Brower R, Esteban A, Gattinoni
L, et al: The Berlin definition of ARDS: An expanded rationale,
justification, and supplementary material. Intensive Care Med.
38:1573–1582. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sebag SC, Bastarache JA and Ware LB:
Therapeutic modulation of coagulation and fibrinolysis in acute
lung injury and the acute respiratory distress syndrome. Curr Pharm
Biotechnol. 12:1481–1496. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ozolina A, Sarkele M, Sabelnikovs L,
Skesters A, Jaunalksne I, Serova J, Ievins T, Bjertnaes LJ and
Vanags I: Activation of coagulation and fibrinolysis in acute
respiratory distress syndrome: A prospective pilot study. Front Med
(Lausanne). 3:642016.PubMed/NCBI
|
|
11
|
Idell S, James KK, Levin EG, Schwartz BS,
Manchanda N, Maunder RJ, Martin TR, McLarty J and Fair DS: Local
abnormalities in coagulation and fibrinolytic pathways predispose
to alveolar fibrin deposition in the adult respiratory distress
Syndrome. J Clin Invest. 84:695–705. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hasday JD, Bachwich PR, Lynch JP III and
Sitrin RG: Procoagulant and plasminogen activator activities of
bronchoalveolar fluid in patients with pulmonary sarcoidosis. Exp
Lung Res. 14:261–278. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chapman HA, Allen CL and Stone OL:
Abnormalities in pathways of alveolar fibrin turnover among
patients with interstitial lung disease. Am Rev Respir Dis.
133:437–443. 1986.PubMed/NCBI
|
|
14
|
Chapman HA, Bertozzi P and Reilly JJ Jr:
Role of enzymes mediating thrombosis and thrombolysis in lung
disease. Chest. 93:1256–1263. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shaver CM, Grove BS, Putz ND, Clune JK,
Lawson WZ, Carnahan RH, Mackman N, Ware LB and Bastarache JA:
Regulation of alveolar procoagulant activity and permeability in
direct acute lung injury by lung epithelial tissue factor. Am J
Respir Cell Mol Biol. 53:719–727. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bastarache JA, Wang L, Geiser T, Wang Z,
Albertine KH, Matthay MA and Ware LB: The alveolar epithelium can
initiate the extrinsic coagulation cascade through expression of
tissue factor. Thorax. 62:608–616. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Osterholzer JJ, Christensen PJ, Lama V,
Horowitz JC, Hattori N, Subbotina N, Cunningham A, Lin Y, Murdock
BJ, Morey RE, et al: PAI-1 promotes the accumulation of exudate
macrophages and worsens pulmonary fibrosis following type II
alveolar epithelial cell injury. J Pathol. 228:170–180. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Karin M: How NF-kappaB is activated: The
role of the IkappaB kinase (IKK) complex. Oncogene. 18:6867–6874.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gamble C, McIntosh K, Scott R, Ho KH,
Plevin R and Paul A: Inhibitory kappa B kinases as targets for
pharmacological regulation. Br J Pharmacol. 165:802–819. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kwon Y, Choi SK, Byeon S and Lee YH:
Involvement of inhibitor kappa B kinase 2 (IKK2) in the regulation
of vascular tone. Lab Invest. 98:1311–1319. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aupperle K, Bennett B, Han Z, Boyle D,
Manning A and Firestein G: NF-kappa B regulation by I kappa B
kinase-2 in rheumatoid arthritis synoviocytes. J Immunol.
166:2705–2711. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hayden MS and Ghosh S: Shared principles
in NF-kappaB signaling. Cell. 132:344–362. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gao MY, Chen L, Yang L, Yu X, Kou JP and
Yu BY: Berberine inhibits LPS-induced TF procoagulant activity and
expression through NF-κB/p65, Akt and MAPK pathway in THP-1 cells.
Pharmacol Rep. 66:480–484. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jeffers A, Owens S, Koenig K, Quaid B,
Pendurthi UR, Rao VM, Idell S and Tucker TA: Thrombin
down-regulates tissue factor pathway inhibitor expression in a
PI3K/Nuclear Factor-κB-dependent manner in human pleural
mesothelial cells. Am J Respir Cell Mol Biol. 52:674–682. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kebir DE, Damlaj A, Makhezer N and Filep
JG: Toll-like receptor 9 signaling regulates tissue factor and
tissue factor pathway inhibitor expression in human endothelial
cells and coagulation in mice. Crit Care Med. 43:e179–e189. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kitasato L, Yamaoka-Tojo M, Hashikata T,
Ishii S, Kameda R, Shimohama T, Tojo T and Ako J: Factor Xa in
mouse fibroblasts may induce fibrosis more than thrombin. Int Heart
J. 55:357–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ding R, Zhao D, Li X, Liu B and Ma X:
Rho-kinase inhibitor treatment prevents pulmonary inflammation and
coagulation in lipopolysaccharide-induced lung injury. Thromb Res.
150:59–64. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using Real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lesnik P, Rouis M, Skarlatos S, Kruth HS
and Chapman MJ: Up take of exogenous free cholesterol induces
upregulation of tissue factor express in human monocyte-derived
macrophages. Proc Natl Acad Sci USA. 89:10370–10374. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sakuma T, Barry MA and Ikeda Y: Lentiviral
vectors: Basic to Translational. Biochem J. 443:603–618. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu B, Tai A and Wang P: Immunization
delivered by lentiviral vectors for cancer and infectious diseases.
Immunol Rev. 239:45–61. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tanaka M, Funtes ME, Yamaguchi K, Durnin
MH, Dalrymple SA, Hardy KL and Goeddel DV: Embryonic lethality,
liver degeneration, and impaired NF-kappa B activation in
IKK-beta-deficient mice. Immunity. 10:421–429. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bastarache JA, Wang L, Geiser T, Wang Z,
Albertine KH, Matthay MA and Ware LB: The alveolar epithelium can
initiate the extrinsic coagulation cascade through expression of
tissue factor. Thorax. 62:608–616. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kasthuri RS, Glover SL, Boles J and
Mackman N: Tissue factor and tissue factor pathway inhibitor as key
regulators of global hemostasis: Measurement of their levels in
coagulation assays. Semin Thromb Hemost. 36:764–771. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu RM: Oxidative stress, plasminogen
activator inhibitor 1, and lung fibrosis. Antioxid Redox Signal.
10:303–319. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Prabhakaran P, Ware LB, White KE, Cross
MT, Matthay MA and Olman MA: Elevated levels of plasminogen
activator inhibitor-1 in pulmonary edema fluid are associated with
mortality in acute lung injury. Am J Physiol Lung Cell Mol Physiol.
285:L20–L28. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ling W, Bastarache JA, Wickersham N, Fang
X, Matthay MA and Ware LB: Novel role of the human alveolar
epithelium in regulating intra-alveolar coagulation. Am J Respir
Cell Mol Biol. 36:497–503. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cristina M: Nuclear factor-kappa-B
signaling in lung development and disease: One pathway, numerous
functions. Birth Defects Res A Clin Mol Teratol. 100:202–216. 2014.
View Article : Google Scholar : PubMed/NCBI
|