Introduction
Osteoarthritis (OA) is a common and costly disease
in modern society. It is the leading reason for medical
consultation in the elderly and affects >30% of people >60
years old in the USA (1,2). OA was once considered an articular
cartilage disease; however, numerous studies investigating the
structures at the osteochondral junction and the
pathogenesis/symptoms of OA indicate that it is a ‘whole joint
disease’. OA is a pathology with the characteristics of decreased
articular cartilage thickness, sclerosis of subchondral bone,
formation of osteophytes and alteration of the synovial fluid
components (2,3).
Bone tissue is dynamically shaped and repaired. The
delicate balance between bone resorption and bone formation is
important to maintain normal bone structure and function (4,5).
Previous studies have suggested that osteoclast-mediated bone
resorption of mineralized cartilage at the interface of subchondral
bone and cartilage is an early initiating pathology in the
progression of OA, and bisphosphonate can significantly improve the
cartilage damage in the early stages (6). Osteoclast-mediated resorption plays a
crucial role in OA and may be an initiator during OA development
(7,8). Excessive activation of osteoclasts is
considered to be the primary mechanism that leads to joint
diseases, including OA, cartilaginous degeneration and osteoporosis
(9,10). Thus, inhibition of osteoclastogenesis
is likely to be chondroprotective and prevent resorption of
cartilage in OA.
Osteoclasts are derived from monocyte/macrophage
precursor cells, near to or at the bone surface, and are a
tissue-specific macrophage polykaryon (4). Receptor activator of nuclear factor-κB
(RANK) ligand (RANKL) and monocyte/macrophage colony-stimulating
factor are haematopoietic factors essential for osteoclastogenesis
and bone reabsorption. Once stimulated in the presence of these two
cytokines, osteoclast precursors move towards the bone surface and
begin to differentiate (11,12). The binding of RANKL with its
receptor, RANK, induces osteoclast differentiation and activation
through numerous signaling cascades, including inhibitor of NF-κB
kinase (IKK), p38, proto-oncogene tyrosine-protein kinase (c-Src)
and JNK pathways. Following osteoclast differentiation, various
osteoclastogenesis associated transcription factors, such as NF-κB,
c-Fos and nuclear factor of activated T cells 1 (NFATc1), are
activated (13,14).
Protocatechuic acid (PCA), also known as
3,4-dihydroxybenzoic acid, is an abundantly and naturally
distributed phenolic acid (15). It
has been reported to have antioxidant, antibacterial,
anti-inflammatory and anti-cancer activity (16–19).
Experimental findings demonstrate the promising anti-inflammatory
and analgesic activity of PCA in rats and mice, including in a
Freund's adjuvant-induced arthritis rat model (19). PCA has recently been found to inhibit
the differentiation of osteoclasts and induce apoptosis of mature
osteoclasts by regulating oxidative stress and inflammation
(20). The present study was
designed to investigate the positive effects of PCA on anterior
cruciate ligament (ACL) transection (ACLT)-induced OA. Furthermore,
an attempt was made to understand the underlying mechanisms of
action behind the antiarthritic activity of PCA.
Materials and methods
Experimental animal model and drug
treatment
A total of 72 male Wistar rats (age, 8 weeks;
weight, 245–255 g) obtained from Shanghai SLAC Laboratory Animal
Co., Ltd. were used in the present study. Rats were housed under
controlled conditions (25±2°C; 70% humidity; 12 h light/dark cycle)
with free access to water and food. The experimental protocols were
approved by the Animal Experimental Ethical Committee of the
General Hospital of Ningxia Medical University and were carried out
on the basis of relevant national and international guidelines.
ACLT was generated as previously described (21). Rats were operated on under
anesthesia, consisting of an intraperitoneal injection of mixed
acepromazine (1.25 mg/kg), xylazine (6.25 mg/kg) and ketamine (38
mg/kg) (22). Incisions were made on
the medial side of the right knee joint and the medial side of the
patellar tendon with a para-patellar skin incision, successively.
The patella was dislocated laterally to gain entry to the joint
space, then the ACL was transected when the knee was flexed.
Complete transection of the ligament was confirmed by a positive
anterior drawer test (23). Then,
the joint was washed with sterile saline to minimize the
accompanied inflammatory response. Finally, the joint capsule was
closed using 3-0 absorbable thread and the skin was sutured with
5-0 nylon thread. Rats in the sham group underwent arthrotomy
rather than the full ACLT (21).
All rats were randomly allocated to three groups:
Group 1, Sham (arthrotomy); group 2, ACLT; and group 3, ACLT with
PCA. Rats within the ACLT with PCA group were administered PCA (50
mg/kg) daily by gavage for 4 consecutive weeks from the day of
surgery. The dose of PCA used in the experiment was based upon our
own pilot data (Zhang et al; unpublished). Euthanasia was
performed on all rats at 1, 2 or 4 weeks post-surgery (19).
ELISA
Protein was extracted from knee joints according to
Nielsen et al (24) at week
0, 1, 2 and 4 after operation. Knee joints were isolated after
euthanasia as described above, then immediately frozen in liquid
nitrogen and stored at −80°C for use. The tibia and femur were cut
3 mm from the joint, producing samples weighing 500–700 mg. These
samples were frozen in liquid nitrogen again, and transferred to a
Bessman tissue pulverizer (Spectrum Chemical Manufacturing Corp.).
The samples were crushed with 3 ml extraction buffer, consisting of
50 mM Tris-HCl buffer (pH 7.4), 0.1% Triton X-100, 0.1 M NaCl, 10
Mm (GM6001; Enzo Life Sciences, Inc.) and 1 tablet/10 ml buffer of
Complete Mini EDTA-free protease inhibitor cocktail (Roche
Diagnostics). Tissues were homogenized twice for 30 sec using an
OMNI homogenizer (Omni International, Inc.) at speed level 4, then
the samples were centrifuged for 10 min at 1,700 × g and 4°C,
supernatants were obtained and centrifuged at 10,000 × g and 4°C.
Finally, the supernatants were stored at −20°C until use.
Rat-specific commercially available ELISA kits
(Elabscience®) were used to evaluate the levels of
C-terminal telopeptide of type I collagen (CTX)-I (cat no.
E-EL-R1456) and CTX–II (cat no. E-EL-R2554) in protein extracts
collected from the knee, according to the manufacturer's
instructions (24).
Specimen preparation and histological
analysis
Knee joints were isolated 4 weeks after surgery. The
knee samples were fixed in 4% formaldehyde in PBS for 48 h (pH 7.0;
4°C) and decalcified for 24 h at 37°C with 22.5% formic acid and
340 mM sodium citrate. Specimens were embedded in paraffin after
demineralization. Blocks were trimmed and the articular cartilage
was exposed. Sections were collected at intervals of 0, 100 and 200
µm. A total of 10 5 µm-thick sections were collected at each
interval. These sections were treated with Safranin-O/Fast Green
staining as previously described (25).
Total RNA preparation and reverse
transcription-quantitative PCR (RT-qPCR)
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to extract total RNA from knee
tissues following the manufacturer's instructions. cDNA was
generated by reverse transcription with equal quantities of RNA
using the SuperScript Reverse Transcriptase kit (Thermo Fisher
Scientific, Inc.). The PCR was performed under the following
conditions: 10 min at 95°C, 55 cycles of 15 sec at 95°C and 1 min
at 60°C and a final 2 min at 50°C, were performed with an MX3000P
(Stratagene; Agilent Technologies, Inc.). The comparative
2−ΔΔCq method was used to calculate the relative
quantification (26). All qPCR
primer sequences were as follows: c-Src forward,
5′-GGACAGTGGCGGATTCTACAT-3′ and reverse,
5′-GGGTCTGAGGCTTGGATGTG-3′; β3-integrin forward,
5′-TACTCTGCCTCCACCACCAT-3′ and reverse,
5′-TTTCCCGTAAGCATCAACAA-3′); MMP-9 forward,
5′-GCAGAGGCATACTTGTACCG-3′ and reverse,
5′-TGATGTTATGATGGTCCCACTTG-3′; IL-6 forward,
5′-GGCCCTTGCTTTCTCTTCG-3′ and reverse, 5′-ATAATAAAGTTTTGATTATGT-3′.
Total cDNA was amplified and analyzed using SYBR Green PCR Master
Mix (Thermo Fisher Scientific, Inc.) in a Fast Real-time PCR 7500
System (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
original Cq values were adjusted to GAPDH (3,10).
Tartrate-resistance acid phosphatase
(TRAP) staining
RAW264.7 cells (1×106 cells/well) were
cultured in α-MEM (Sigma-Aldrich; Merck KGaA) at 37°C supplemented
with 10% FBS (Dalian Meilun Biology Technology Co., Ltd.) and RANKL
(100 ng/ml) to induce differentiation (27). These cells were treated with 8 mM PCA
for 2 h and continuously treated with RANKL (100 ng/ml) for 4 days.
The cells were collected and washed with PBS, then fixed in
formalin for 10 min at room temperature and permeabilized in
ethanol/acetone (1:1) for 1 min at room temperature. The number of
TRAP-positive multinucleated cells (MNCs) was observed and images
captured using a light microscope at ×100 magnification (Leica
Microsystems GmBH) (20,28).
Functional bone resorption pit
assay
The resorption pit assay was conducted following the
methods described by Lu et al (28). RAW264.7 cells (1×105
cells/well) were seeded at 37°C on a 24-well Corning Osteo Assay
Surface well plate (Corning, Inc.), and were pretreated with PCA (8
mM) for 2 h followed by the addition of 100 ng/ml RANKL. After 5
days of incubation, all remaining cells were lysed by NaOH (1 M)
for 10 min at 4°C, then the wells were washed twice with PBS. The
numbers of bone lacuna were observed under a phase contrast
inverted microscope (BH-2; Olympus Corporation) and analyzed with
Metamorph imaging analysis software version 2.5 at ×400
magnification (Molecular Devices, LLC).
Western blotting
RAW264.7 cells were pretreated with PCA (8 mM) for 2
h at room temperature and then cultured with RANKL (100 ng/ml) for
1 h at 37°C [for CTX–I, CTX–II, c-Src, interleukin (IL)-6,
mitogen-activated protein kinases (MAPKs) and phosphorylated
(p)-Akt expression] or for 3 h at 37°C [for p65, cyclooxygenase
(COX)-2 and NFATc1 expression]. RIPA lysis buffer (Sangon Biotech
Co., Ltd.) was used to isolate the whole cell protein and a Nuclear
Extraction kit (cat. no. Ab113474; Abcam) was used to isolate the
nuclear extracts. The bicinchoninic acid method was used to
determine protein concentration. Proteins (100 µg) were separated
by 10% SDS-PAGE and transferred to PVDF membranes. The membranes
were blocked with 5% powdered non-fat milk for 1 h at room
temperature and were then incubated with antibodies against CTX–I
(1:500; cat. no. CEA665Ra; Wuhan Uscn Business Co., Ltd.), LCTX–II
(1:500; cat. no. CEA686Ra; Wuhan Uscn Business Co., Ltd.), MAPK
(1:250; cat. no. ab151279; Abcam), Akt (1:250; cat. no. ab151279;
Abcam), p-Akt (1:500; cat. no. ab38449; Abcam), COX-2 (1:1,000;
cat. no. ab179800; Abcam), NFATc1 (1:1,000; cat. no. ab207215;
Abcam), Lamin A/C (1:1,000; cat. no. ab108922, Abcam), β-actin
(1:1,000; cat. no. ab8226; Abcam), c-Src (1:500; cat. no. ab207462;
Abcam), IL-6 (1:1,000; cat. no. ab208113; Abcam), NF-κB p65
(1:1,000; cat. no. ab207297; Abcam) overnight at 4°C. The membranes
were further incubated with a 1:10,000 of goat anti-rabbit IgG
(H+L) horseradish peroxidase conjugate (cat. no. A2028; Beyotime
Institute of Biotechnology) as a secondary antibody for 2 h at room
temperature. The immunoreactivity was visualized using an ECL
system (Amersham; GE Healthcare). The relative intensity of the
signal produced by the western blotting bands was analyzed using
ImageJ software (version 1.4.0.; National Institutes of Health)
(20).
Statistical analysis
Results in the present study are presented as the
mean ± SEM of three independent experiments. Data comparisons
between groups and within groups were analyzed using either
unpaired or paired Student's t-test and one-way ANOVA with Tukey's
post-hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
PCA relieves ACLT-induced OA
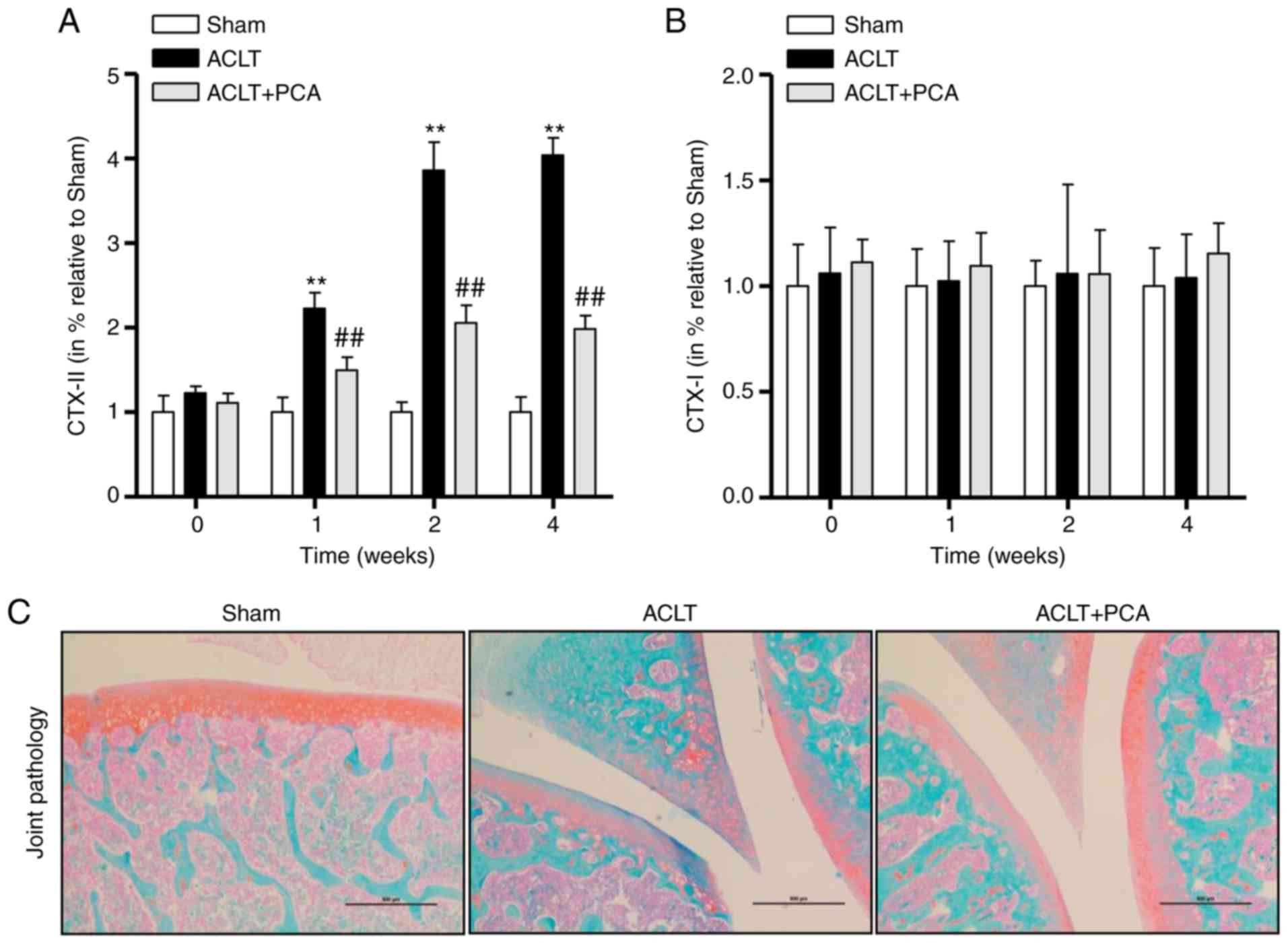
To confirm the effect of PCA on collagen degradation
and synthesis, two osteoarthritic markers, CTX–I and CTX–II, were
detected within whole knee joints from the sham, ACLT and ACLT with
PCA-treated groups at weeks 0, 1, 2 and 4 after the operation. The
levels of CTX–II in the ACLT group were significantly raised 1–4
weeks after surgery compared to the sham control, while this effect
was significantly reversed in the ACLT-PCA group (Fig. 1A). This suggested that the rat model
of ACLT was successfully established and that PCA had an
anti-osteoarthritic effect (Fig.
1A). By contrast, concentrations of CTX–I were not altered
during the period after surgery in any group (Fig. 1B). The Safranin-O/Fast Green staining
demonstrated decreased proteoglycans in the superficial and even
the middle zone, with clefts extending into the middle layers after
ACLT surgery. However, the severity of degenerative features in the
PCA treatment group was much milder than that in ACLT group,
demonstrating that PCA relieved the ACLT-induced degenerative
changes in the articular cartilage (Fig.
1C). Collectively, these results indicated that PCA has
potential for treating ACLT-induced OA.
PCA decreases the activation of
osteoclasts in ACLT rats
To further identify the therapeutic effects of PCA
for treating ACLT-induced OA, the mRNA expression of
osteoclast-related genes in knee tissues were analyzed by RT-qPCR.
Compared with the sham group, the expression levels of c-Src,
β3-integrin, matrix metalloproteinase (MMP)-9 and IL-6 were
significantly increased at the mRNA level after ACLT surgery
(Fig. 2A-D). However, PCA treatment
effectively reserved these abnormalities. Furthermore, it was also
found that the protein levels of CTX–I, CTX–II, c-Src and IL-6 were
raised after ACLT surgery compared to the sham-operated group
(Fig. 2E). PCA treatment
significantly reversed the increased levels of CTX–I, CTX–II, c-Src
and IL-6 compared to the ACLT group (Fig. 2E). These results demonstrated that
PCA can decrease ACLT-induced osteoclastogenesis in
vivo.
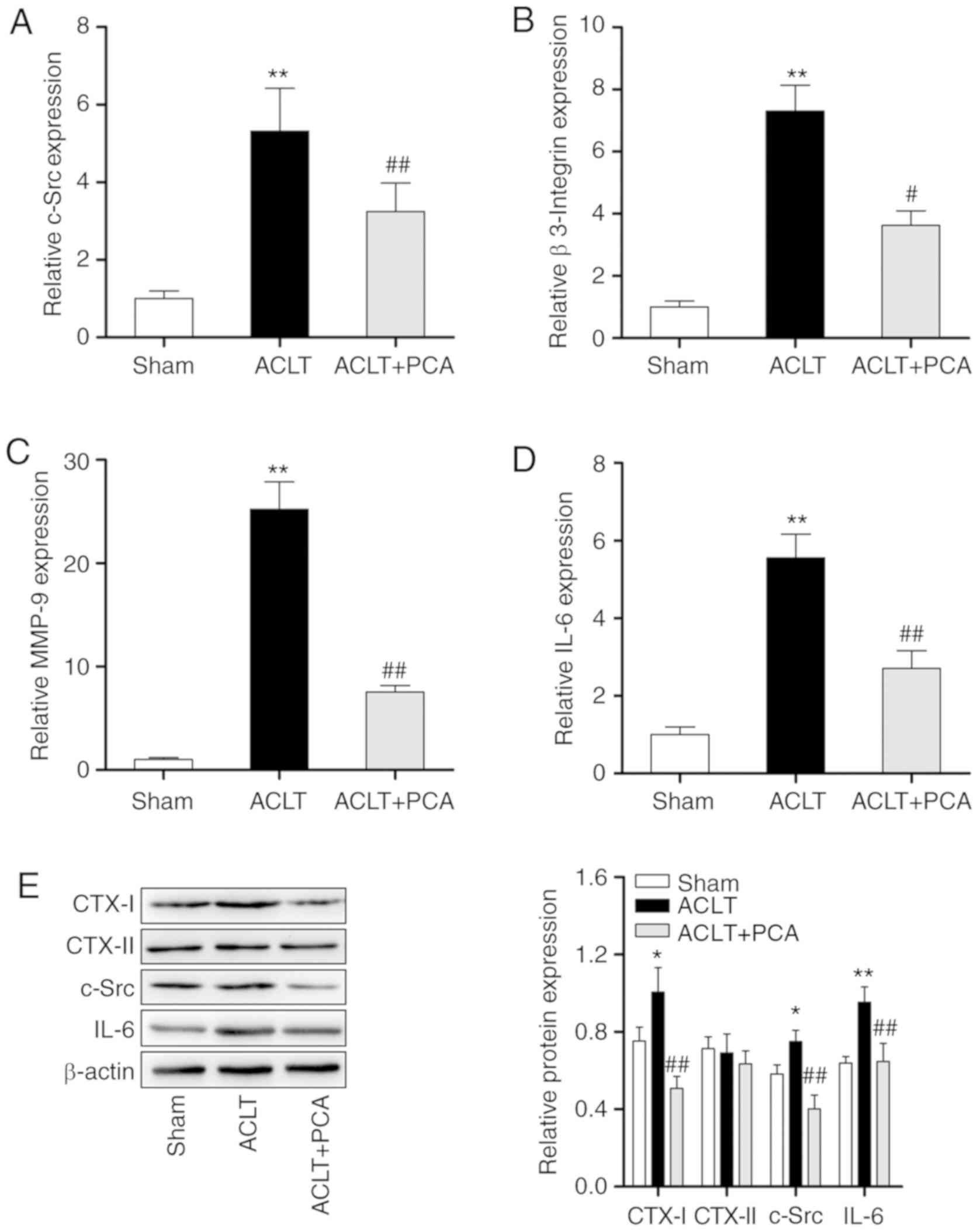 | Figure 2.PCA decreases the activation of
osteoclast in ACLT rats. The relative expression levels of (A)
c-Src, (B) β-3 Integrin, (C) MMP-9 and (D) IL-6 in knee tissue
samples were evaluated through reverse transcription-quantitative
PCR. GAPDH was used as a reference. (E) The protein expressions of
CTX–I, CTX–II, c-Src and IL-6 in knee tissue samples were evaluated
through western blotting. β-actin was used as a loading control.
*P<0.05, **P<0.01 vs. sham; #P<0.05,
##P<0.01 vs. ACTL. ACLT, anterior cruciate ligament
transection; c-Src, proto-oncogene tyrosine-protein kinase; MMP,
matrix metalloproteinase; PCA, protocatechuic acid. |
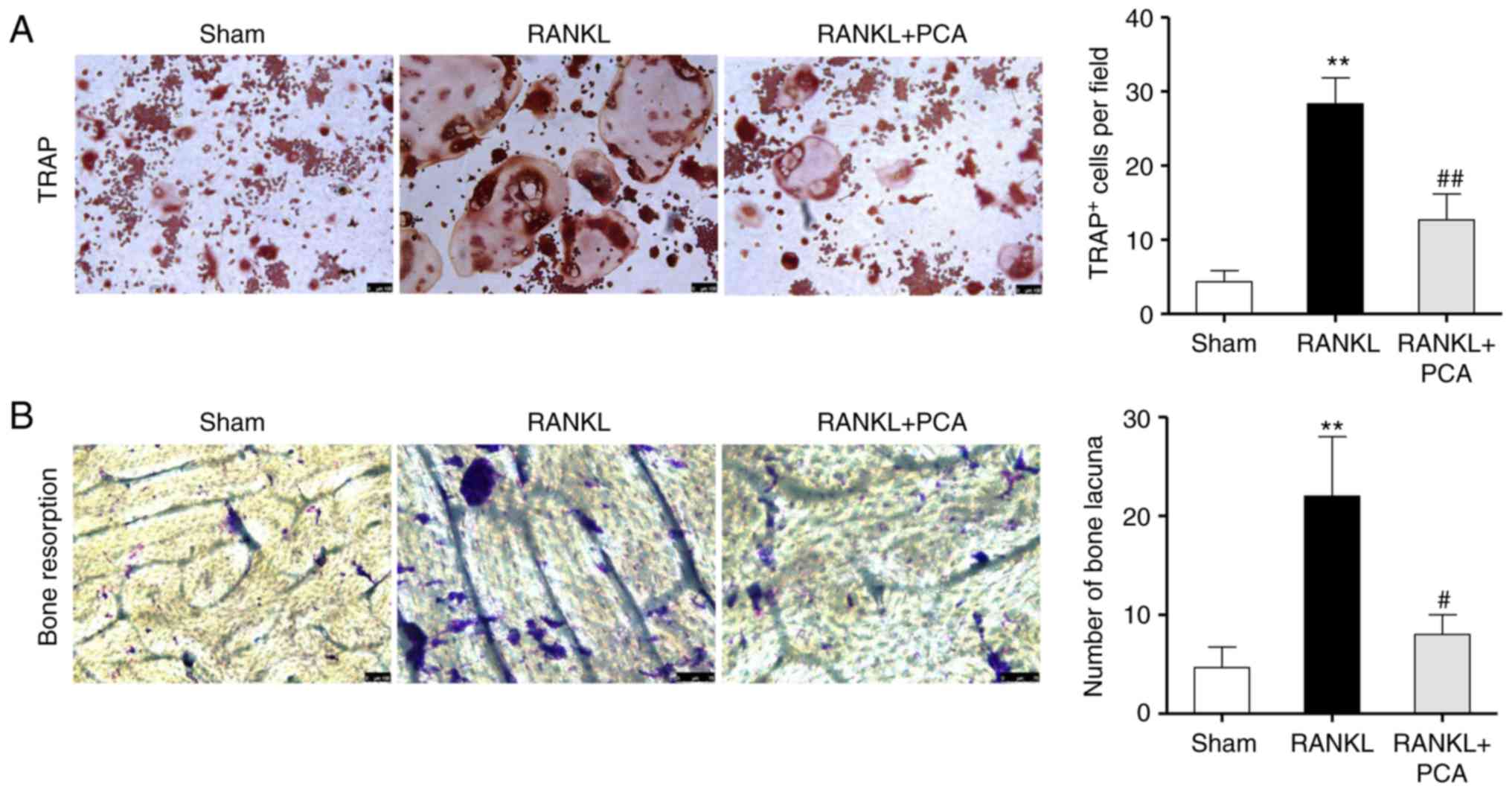
PCA suppresses RANKL-induced
osteoclast differentiation and the bone resorbing activity of
RAW264.7 cells
To observe the effect of PCA on osteoclast
differentiation, murine macrophage RAW264.7 cells were incubated in
the presence of RANKL with or without PCA. The effect of PCA on
osteoclast differentiation was evaluated using TRAP staining.
Microscopic assessment showed that abundant TRAP + MNCs were formed
in the culture as a response to RANKL on day 4 (Fig. 3A), whereas the PCA treatment reduced
the number of RANKL-stimulated TRAP + MNCs cells (P<0.01).
Consistent with the function of preventing osteoclastic
differentiation, PCA disrupted the pit-forming activity of
RANKL-stimulated osteoclasts. Compared to the group treated with
RANKL alone, a notable reduction in resorption pit area and number
of bone lacuna was observed in the RANKL + PCA group (Fig. 3B). Together, these data demonstrated
that PCA suppresses osteoclastogenesis and osteoclast function
in vitro. Thereafter, the inhibitory effects of PCA on
osteoclastogenesis and bone resorption were further validated in
vitro.
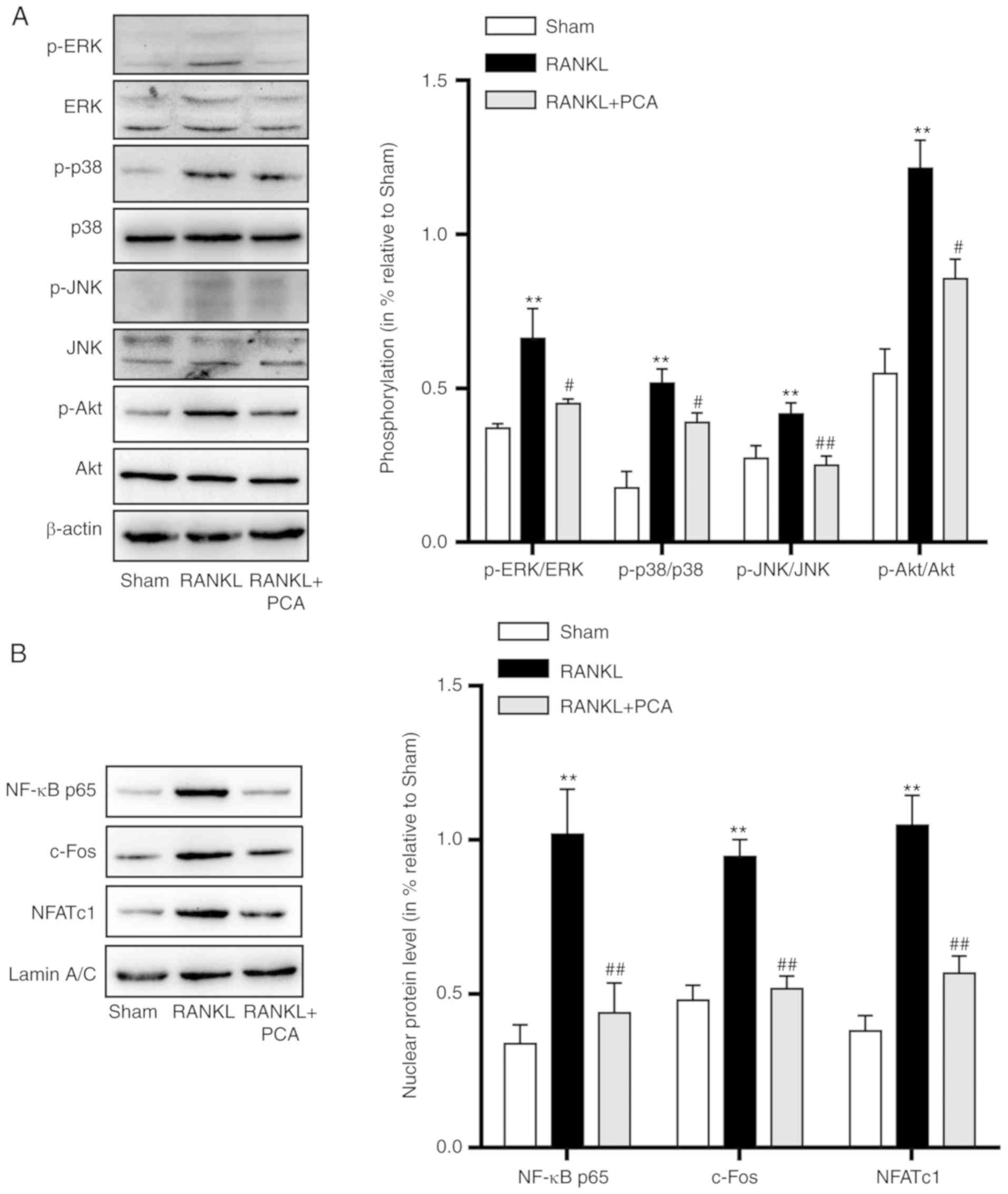
PCA inhibited the RANKL-induced
activation of MAPK, Akt and NF-κB signaling
In osteoclast precursor cells, the binding of RANKL
to its precursor RANK triggers downstream signaling pathways,
including MAPKs and Akt signaling pathways which are essential for
osteoclast differentiation and activation. In order to investigate
the signaling pathways that PCA affects, the expression of MAPKs,
p-Akt and the three critical nuclear factors involved in
osteoclastogenesis (NF-κB p65, c-Fos and NFATc1) were evaluated.
The results showed that there was not only a significant increase
in the proportion of phosphorylated of MAPKs (ERK, p-38 and JNK)
and Akt compared to their unphosphorylated versions, but also
increased protein expression levels of NF-κB p65, c-Fos and NFATc1
(Fig. 4). However, pretreatment with
PCA in the present of RANKL led to downregulation in the
phosphorylation levels of MAPKs and Akt. Similarly, the downstream
nuclear factors (NF-κB p65, c-Fos and NFATc1), crucial for
osteoclast differentiation and activation were also downregulated
(Fig. 4). These results indicated
that PCA suppressed the activation of the MAPK, Akt and NF-κB
signaling pathways that were induced by RANKL during
osteoclastogenesis.
Discussion
OA is a prevalent form of arthritic disease, which
causes pain and dysfunction principally in the knee and hip joints.
OA affects 18% of women and 9.6% of men over the age of 60
(29). Patients are burdened with
the high medical cost of substantial use of medications. At
present, the optimal therapy for OA is a combination of
non-pharmacological and pharmacological therapies. Pharmacological
therapies are essential to relieve pain and recover joint function
for most patients (2). Nonsteroidal
anti-inflammatory drugs, corticosteroids and hyaluronan are used
clinically to improve the symptoms of OA, but they fail to reverse
cartilage damage and they also exhibit a wide range of adverse
effects, including cardiovascular, renal and gastrointestinal side
effects (30). As such, more
effective therapies with milder adverse effects need to be
developed.
PCA is a phenolic compound on which a variety of
research work has been carried out. Recently, a study demonstrated
that PCA has promising anti-inflammatory and analgesic activities
in Freund's adjuvant-induced arthritis, because of its antioxidant
and membrane-stabilizing properties (19). Another study showed that PCA
suppresses osteoclast differentiation induced by RANKL in RAW264.7
cells by regulating oxidative stress and inflammation, and that two
osteoclastogenesis-related transcription factors, NF-κB and Nrf-2,
are involved (20). PCA has also
been proven to induce apoptosis in mature osteoclasts (20). Importantly, the activation of
osteoclasts plays an important role in the initiation and
progression of OA. Park et al (31) found that PCA could attenuate
osteoclastogenesis by regulating JNK/c-Fos/NFATc1 signaling and
preventing inflammatory bone loss in mice. Considering the
aforementioned observations, this present study attempted to
determine whether PCA can attenuate OA by suppressing
osteoclastogenesis mediated by the osteoclastogenesis-associated
MAPK and Akt signaling pathways, as well as critical nuclear
factors.
In this present study, it was demonstrated that PCA
exerts an anti-arthritic effect on ACLT-induced OA rats, a model
that has been widely applied to investigate OA. Many biomarkers
such as collagens, chondroitin sulfate and aggrecan are used to
evaluate OA (32,33). CTX–II is a primary component of
articular cartilage and accounts for 90–95% of collagen content,
acting in the formation of the fibrillar structure to maintain the
tensile strength of cartilage (33).
Thus, CTX–II has been considered as a crucial biomarker for OA in
many studies (34). This present
research revealed that ALCT surgery substantially raised CTX–II
levels in the knee joints compared to the levels in the sham group.
Notably, PCA inhibited the degradation of CTX–II induced by ACLT
surgery. These results suggested that PCA could regulate the
expression of CTX–II during the development of OA. Safranin-O/Fast
Green staining demonstrated reduced proteoglycan loss and milder
degenerative changes in articular cartilage with PCA treatment.
However, consistent with another study (24), the levels of CTX–I were not changed
by ACLT or PCA administration. Based on these results, PCA appears
to exert a chondroprotective effect on OA induced by ACLT by
correcting the metabolism of proteoglycans as well as by inhibiting
the degradation of CTX–II, and possibly even promoting the
synthesis of CTX–II in the cartilage.
Osteoclast activation plays a crucial role in the
progression of OA. Inhibiting osteoclast activity prevents bone
resorption and can prevent the progression of OA (9). Thus, the expression of
osteoclast-related genes in knee tissues was examined in this
present study. ACLT enhanced the expression of c-Src, β3-integrin
and IL-6 at both the mRNA and protein level. Additionally, Dong
et al (35) reported that the
absence of MMP-9 restrained the movement of osteoclasts and further
affected the functions of osteoclasts. The present study also found
that ACLT upregulated the expression of MMP-9. PCA treatment
effectively corrected these abnormalities, demonstrating that PCA
can reduce osteoclastogenesis in ALCT-induced OA, which may account
for the antiarthritic activity of PCA.
To further confirm that PCA suppresses
osteoclastogenesis, RAW264.7 osteoclast precursor cells were
incubated with PCA and their differentiation induced by RANKL.
These data demonstrated that PCA effectively inhibits osteoclast
differentiation, reflected by the decrease in TRAP+ MNCs when the
cells were treated with both RANKL and PCA, compared to RANKL
alone. Osteoclasts are essential for bone resorption which may be
enhanced by excessive osteoclastogenesis. A previous study
indicated that osteoclast bone resorption is critical in
pathological bone diseases including OA (3), and inhibition of osteoclasts has been
proven to prevent pain and cartilage degradation in a degenerative
joint disease rat model (7,8). In this present study, it was also found
that PCA was chondroprotective to the ACLT-induced OA. Thus, it was
further speculated that PCA inhibits osteoclast bone resorption.
The results from this present study confirmed that PCA
significantly decreased the resorption pit area and the number of
bone lacuna compared to RANKL treatment alone. Altogether, these
in vitro results demonstrated that PCA suppresses
osteoclastogenesis and osteoclast function. This conclusion is
similar to that of Wu et al (20), who demonstrated that PCA suppresses
osteoclast differentiation by negatively regulating
osteoclast-related genes and oxidative stress through two crucial
transcription factors, NF-κB and Nrf-2 (20).
RANK/RANKL/osteoprotegerin signaling is an important
mediator of osteoclast differentiation and activation (36,37). TNF
receptor associated factor 6 (TRAF6) is a key adaptor that
assembles signaling proteins and triggers the expression of
osteoclast-specific genes, resulting in osteoclast differentiation
and activation (38,39). Among those known signaling cascades
involved in osteoclast differentiation and activation, MAPK
signaling is critical for osteoclast differentiation and has been
relatively well characterized (40).
c-Src plays an important role in osteoclast activation, by binding
to TRAF6 and delivering RANK-induced signaling to PI3K and Akt,
promoting cell survival, cytoskeletal rearrangement and cellular
motility (38,39,41).
Many downstream molecules of PI3K/Akt, such as mTOR, protein
tyrosine kinase 2β and CBL, are known to induce osteoclast
differentiation, survival and bone resorption (40,42,43). The
NF-κB pathway is also involved in osteoclast activation and the
subsequent bone resorption. Upon RANKL stimulation, IKK
phosphorylates p65, which is then translocated to the nucleus
(44). NF-κB regulates the
expression of c-Fos (45) and NFATc1
(46) in RANKL-mediated
osteoclastogenesis. MAPKs also regulate the RANKL-induced
activation of AP-1 through c-Fos and NFATc1. The two factors work
synergistically to promote the expression of terminal
osteoclastogenesis-related genes (44). Consistent with a previous study
(20), the results of this present
study demonstrated that all three MAPK families, ERK, JNK and p38,
as well as Akt, were activated by RANKL in osteoclast precursors.
The downstream nuclear factors p65, c-Fos and NFATc1, essential for
osteoclastogenesis, were also activated. Nevertheless, PCA
effectively inhibited the phosphorylation of MAPKs and Akt induced
by RANK signaling. Likewise, the critical downstream factors p65,
c-Fos and NFATc1 were suppressed by PCA.
There are several limitations related to the current
study. For example, PCA can promote the proliferation and
phenotypic maintenance of rabbit articular chondrocytes (47). However, the effect of PCA on
chondrocytes has not been assessed in the ACLT-induced OA rat
model. Additionally, the balance between osteoblasts and
osteoclasts is important for the progression of OA. It has been
proven that PCA also can promote the proliferation and
differentiation of primary rat osteoblasts cultured in vitro
(48). This present study aimed to
determine the effects of PCA on osteoclasts in OA; the effects of
PCA on osteoblasts in OA will be the focus of future studies. It is
planned for future research to use MAPK, Akt or NF-κB agonists for
rescue experiments in osteoclasts after PCA treatment. Changes in
related phenotypes and biomarkers, such as osteoclast proliferation
and differentiation will be examined in further in vitro
experiments.
In conclusion, this present study suggested that PCA
can efficiently attenuate ALCT-induced OA. PCA not only suppressed
the formation of osteoclasts, but also interrupted osteoclast bone
resorption. It was found that osteoclastogenesis-associated
signaling, including MAPK, Akt and NF-κB signaling, as well as the
critical nuclear factors induced by RANKL, were suppressed by PCA.
Thus, the anti-arthritic effect of PCA is primarily associated with
the inhibition effect of osteoclastogenesis. Therefore, it was
indicated that PCA has the potential to be developed as a new
therapy for OA and other diseases associated with excessive
osteoclast differentiation and activation.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed in this study are
available from the corresponding author on reasonable request.
Authors' contributions
HY conceived and designed the study. JZ and BF
performed the experiments and collected data. XC and DC were
responsible for statistical analysis and literature research. JZ,
BF and XC were major contributors in writing the manuscript and
data interpretation. HY had full access to all data in the study
and took responsibility for the integrity of the data and the
accuracy of the data analysis. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The experimental protocols were approved by the
Animal Experimental Ethical Committee of the General Hospital of
Ningxia Medical University and were carried out on the basis of
relevant national and international guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lluch Girbés E, Nijs J, Torres-Cueco R and
López Cubas C: Pain treatment for patients with osteoarthritis and
central sensitization. Phys Ther. 93:842–851. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Clouet J, Vinatier C, Merceron C,
Pot-vaucel M, Maugars Y, Weiss P, Grimandi G and Guicheux J: From
osteoarthritis treatments to future regenerative therapies for
cartilage. Drug Discov Today. 14:913–925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Suri S and Walsh DA: Osteochondral
alterations in osteoarthritis. Bone. 51:204–211. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boyle WJ, Simonet WS and Lacey DL:
Osteoclast differentiation and activation. Nature. 423:337–342.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rodan GA and Martin TJ: Therapeutic
approaches to bone diseases. Science. 289:1508–1514. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koh YH, Hong SH, Kang HS, Chung CY, Koo
KH, Chung HW, Cha JH and Son KR: The effects of bone turnover rate
on subchondral trabecular bone structure and cartilage damage in
the osteoarthritis rat model. Rheumatol Int. 30:1165–1171. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Strassle BW, Mark L, Leventhal L, Piesla
MJ, Jian Li X, Kennedy JD, Glasson SS and Whiteside GT: Inhibition
of osteoclasts prevents cartilage loss and pain in a rat model of
degenerative joint disease. Osteoarthritis Cartilage. 18:1319–1328.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bagi CM, Berryman E, Zakur DE, Wilkie D
and Andresen CJ: Effect of antiresorptive and anabolic bone therapy
on development of osteoarthritis in a posttraumatic rat model of
OA. Arthritis Res Ther. 17:3152015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Siebelt M, Waarsing JH, Groen HC, Müller
C, Koelewijn SJ, de Blois E, Verhaar JA, de Jong M and Weinans H:
Inhibited osteoclastic bone resorption through alendronate
treatment in rats reduces severe osteoarthritis progression. Bone.
66:163–170. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao C, Liu Q and Wang K: Artesunate
attenuates ACLT-induced osteoarthritis by suppressing
osteoclastogenesis and aberrant angiogenesis. Biomed Pharmacother.
96:410–416. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yasuda H, Shima N, Nakagawa N, Yamaguchi
K, Kinosaki M, Mochizuki S, Tomoyasu A, Yano K, Goto M, Murakami A,
et al: Osteoclast differentiation factor is a ligand for
osteoprotegerin/osteoclastogenesis-inhibitory factor and is
identical to TRANCE/RANKL. Proc Natl Acad Sci USA. 95:3597–3602.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lacey DL, Timms E, Tan HL, Kelley MJ,
Dunstan CR, Burgess T, Elliott R, Colombero A, Elliott G, Scully S,
et al: Osteoprotegerin ligand is a cytokine that regulates
osteoclast differentiation and activation. Cell. 93:165–176. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao Q, Wang X, Liu Y, He A and Jia R:
NFATc1: Functions in osteoclasts. Int J Biochem Cell Biol.
42:576–579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takayanagi H, Kim S, Koga T, Nishina H,
Isshiki M, Yoshida H, Saiura A, Isobe M, Yokochi T, Inoue J, et al:
Induction and activation of the transcription factor NFATc1 (NFAT2)
integrate RANKL signaling in terminal differentiation of
osteoclasts. Dev Cell. 3:889–901. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kakkar S and Bais S: A review on
protocatechuic acid and its pharmacological potential. ISRN
Pharmacol. 2014:9529432014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li XC, Wang XZ, Chen DF and Chen SZ:
Antioxidant activity and mechanism of protocatechuic acid in vitro.
Funct Foods Health Dis. 1:232–244. 2011.
|
|
17
|
Mahadevan N, Shival i and Kamboj P:
Hibiscus sabdariffa Linn.-An overview. Nat Prod Radiance. 8:77–83.
2009.
|
|
18
|
Tanaka T, Tanaka T and Tanaka M: Potential
cancer chemopreventive activity of protocatechuic acid. J Exp Clin
Med. 3:27–33. 2011. View Article : Google Scholar
|
|
19
|
Lende AB, Kshirsagar AD, Deshpande AD,
Muley MM, Patil RR, Bafna PA and Naik SR: Anti-inflammatory and
analgesic activity of protocatechuic acid in rats and mice.
Inflammopharmacology. 19:255–263. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu YX, Wu TY, Xu BB, Xu XY, Chen HG, Li XY
and Wang G: Protocatechuic acid inhibits osteoclast differentiation
and stimulates apoptosis in mature osteoclasts. Biomed
Pharmacother. 82:399–405. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Galois L, Etienne S, Grossin L,
Watrin-Pinzano A, Cournil-Henrionnet C, Loeuille D, Netter P,
Mainard D and Gillet P: Dose-response relationship for exercise on
severity of experimental osteoarthritis in rats: A pilot study.
Osteoarthritis Cartilage. 12:779–786. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Welberg LA, Kinkead B, Thrivikraman K,
Huerkamp MJ, Nemeroff CB and Plotsky PM:
Ketamine-xylazine-acepromazine anesthesia and postoperative
recovery in rats. J Am Assoc Lab Anim Sci. 45:13–20.
2006.PubMed/NCBI
|
|
23
|
Nyska M, Amir H, Porath A and Dekel S:
Radiological assessment of a modified anterior drawer test of the
ankle. Foot Ankle. 13:400–403. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nielsen RH, Stoop R, Leeming DJ, Stolina
M, Qvist P, Christiansen C and Karsdal MA: Evaluation of cartilage
damage by measuring collagen degradation products in joint extracts
in a traumatic model of osteoarthritis. Biomarkers. 13:79–87. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang Z, Wei X, Gao J, Zhao Y, Zhao Y, Guo
L, Chen C, Duan Z, Li P and Wei L: Intra-Articular injection of
cross-linked hyaluronic acid-dexamethasone hydrogel attenuates
osteoarthritis: An experimental study in a rat model of
osteoarthritis. Int J Mol Sci. 17:4112016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mau LP, Cheng WC, Chen JK, Shieh YS,
Cochran DL and Huang RY: Curcumin ameliorates alveolar bone
destruction of experimental periodontitis by modulating osteoclast
differentiation, activation and function. J Funct Foods.
22:243–256. 2016. View Article : Google Scholar
|
|
28
|
Lu SH, Huang RY and Chou TC: Magnolol
ameliorates ligature-induced periodontitis in rats and
osteoclastogenesis: In vivo and in vitro study. Evid Based
Complement Alternat Med. 2013:6340952013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Woolf AD and Pfleger B: Burden of major
musculoskeletal conditions. Bull World Health Organ. 81:646–656.
2003.PubMed/NCBI
|
|
30
|
Farkouh ME, Greenberg JD, Jeger RV,
Ramanathan K, Verheugt FW, Chesebro JH, Kirshner H, Hochman JS, Lay
CL, Ruland S, et al: Cardiovascular outcomes in high risk patients
with osteoarthritis treated with ibuprofen, naproxen or
lumiracoxib. Ann Rheum Dis. 66:764–770. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Park SH, Kim JY, Cheon YH, Baek JM, Ahn
SJ, Yoon KH, Lee MS and Oh J: Protocatechuic acid attenuates
osteoclastogenesis by downregulating JNK/c-Fos/NFATc1 signaling and
prevents inflammatory bone loss in mice. Phytother Res. 30:604–612.
2016. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rousseau JC and Delmas PD: Biological
markers in osteoarthritis. Nat Clin Pract Rheumatol. 3:346–356.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Garnero P, Rousseau JC and Delmas PD:
Molecular basis and clinical use of biochemical markers of bone,
cartilage, and synovium in joint diseases. Arthritis Rheum.
43:953–968. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Garnero P, Piperno M, Gineyts E, Christgau
S, Delmas PD and Vignon E: Cross sectional evaluation of
biochemical markers of bone, cartilage, and synovial tissue
metabolism in patients with knee osteoarthritis: Relations with
disease activity and joint damage. Ann Rheum Dis. 60:619–626. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dong Z, Bonfil RD, Chinni S, Trindade
Filho JC, Bhagat S, Bhagat S, Fridman R and Cher ML: Enhanced
osteoclast motility due to overexpression of MMP-9 induced by
prostate cancer cells. Cancer Res. 64:773. 2004.PubMed/NCBI
|
|
36
|
Anderson DM, Maraskovsky E, Billingsley
WL, Dougall WC, Tometsko ME, Roux ER, Teepe MC, DuBose RF, Cosman D
and Galibert L: A homologue of the TNF receptor and its ligand
enhance T-cell growth and dendritic-cell function. Nature.
390:175–179. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hsu H, Lacey DL, Dunstan CR, Solovyev I,
Colombero A, Timms E, Tan HL, Elliott G, Kelley MJ, Sarosi I, et
al: Tumor necrosis factor receptor family member RANK mediates
osteoclast differentiation and activation induced by
osteoprotegerin ligand. Proc Natl Acad Sci USA. 96:3540–3545. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Franzoso G, Carlson L, Xing L, Poljak L,
Shores EW, Brown KD, Leonardi A, Tran T, Boyce BF and Siebenlist U:
Requirement for NF-kappaB in osteoclast and B-cell development.
Genes Dev. 11:3482–3496. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Grigoriadis AE, Wang ZQ, Cecchini MG,
Hofstetter W, Felix R, Fleisch HA and Wagner EF: C-Fos: A key
regulator of osteoclast-macrophage lineage determination and bone
remodeling. Science. 266:443–448. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee ZH and Kim HH: Signal transduction by
receptor activator of nuclear factor kappa B in osteoclasts.
Biochem Biophys Res Commun. 305:211–214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wong BR, Besser D, Kim N, Arron JR,
Vologodskaia M, Hanafusa H and Choi Y: TRANCE, a TNF family member,
activates Akt/PKB through a signaling complex involving TRAF6 and
c-Src. Mol Cell. 4:1041–1049. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kolbe L, Immeyer J, Batzer J, Wensorra U,
tom Dieck K, Mundt C, Wolber R, Stäb F, Schönrock U, Ceilley RI and
Wenck H: Anti-inflammatory efficacy of Licochalcone A: Correlation
of clinical potency and in vitro effects. Arch Dermatol Res.
298:23–30. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shui C, Riggs BL and Khosla S: The
immunosuppressant rapamycin, alone or with transforming growth
factor-beta, enhances osteoclast differentiation of RAW264.7
monocyte-macrophage cells in the presence of RANK-ligand. Calcif
Tissue Int. 71:437–446. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim WS, Kim HJ, Lee ZH, Lee Y and Kim HH:
Apolipoprotein E inhibits osteoclast differentiation via regulation
of c-Fos, NFATc1 and NF-κB. Exp Cell Res. 319:436–446. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yamashita T, Yao Z, Li F, Zhang Q, Badell
IR, Schwarz EM, Takeshita S, Wagner EF, Noda M, Matsuo K, et al:
NF-kappaB p50 and p52 regulate receptor activator of NF-kappaB
ligand (RANKL) and tumor necrosis factor-induced osteoclast
precursor differentiation by activating c-Fos and NFATc1. J Biol
Chem. 282:18245–18253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Takatsuna H, Asagiri M, Kubota T, Oka K,
Osada T, Sugiyama C, Saito H, Aoki K, Ohya K, Takayanagi H and
Umezawa K: Inhibition of RANKL-induced osteoclastogenesis by
(−)-DHMEQ, a novel NF-kappaB inhibitor, through downregulation of
NFATc1. J Bone Miner Res. 20:653–662. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Luo L, Wei Q, Liu L, Lin X, Lin C, Zheng
LI and Zhao J: Protocatechuic acid benefits proliferation and
phenotypic maintenance of rabbit articular chondrocytes: An study.
Exp Ther Med. 9:1865–1870. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chin A, Yang Y, Chai L, Wong RW and Rabie
AB: Effects of medicinal herb salvia miltiorrhiza on osteoblastic
cells in vitro. J Orthop Res. 29:1059–1063. 2011. View Article : Google Scholar : PubMed/NCBI
|