Introduction
Pulmonary arterial hypertension (PAH) is
hemodynamically characterized by a progressive elevation in
pulmonary vascular resistance and the loss of pulmonary arterial
compliance, giving rise to excessive right ventricular (RV)
afterload. This ultimately results in abnormal vascular remodeling,
RV hypertrophy and heart failure (1). In addition to the abnormal
proliferation of pulmonary artery endothelial and smooth muscle
cells, it has been revealed that factors contributing to the
progressive narrowing of the distal pulmonary arterioles include
inflammation, in situ thrombosis and an imbalance in the
expression of various endothelial vasoactive mediators; this
includes the reduced production of nitric oxide (NO) and
prostacyclin, and the increased production of endothelin (ET)-1.
Therapies targeting the prostacyclin, ET-1 or NO pathways have
resulted in substantially improved outcomes in patients with PAH
(2). However, current treatment
strategies remain insufficient, with substantial hemodynamic and
functional impairments that cause considerable morbidity (3). Therefore, novel therapeutic approaches
are urgently required.
The β3-adrenergic receptor (β3-AR), first identified
in 1989, has been demonstrated to serve an important function in
heart failure, hypertension, obesity, diabetes and coronary artery
disease, which is independent of the stimulation effects of the β1-
and β2-ARs (4,5). Unlike β1- and β2-AR, which produce
positive chronotropic and inotropic effects upon stimulation, β3-AR
imparts a marked reduction in cardiac contractility by activating
endothelial NO synthase (eNOS), resulting in the subsequent release
of NO from cardiac myocytes (6,7).
Upregulation of β3-AR has been observed in the myocytes of animal
heart failure models in addition to patients with heart failure
(8,9). Nevertheless, the β3-AR responses have
been revealed to vary considerably between species (10), and the efficacy of β3-AR
pharmacotherapy may depend on a number of factors, including the
severity of heart failure and the therapeutic time interval
(11,12).
β3-AR activation is able to influence the
vasodilation of specific blood vessels in humans and animal models
(13–15). However, conflicting results have
proposed the antagonism of β3-AR as a potential preventative
strategy for the development of heart failure (9,16). Due
to the lack of evidence for the existence of β3-AR in the pulmonary
artery (17,18), few studies have reported a
β3-adrenergic response in PAH. Indeed, emerging technologies were
at the forefront of this research area when a rat RNA-Seq
transcriptomic BodyMap across 11 organs confirmed the expression of
β3-AR in rat adrenal, thymus, heart and lung tissues (19). An additional study revealed that
β3-AR was expressed in the human pulmonary artery (20), and that the β3-AR agonist BRL37344
reduced pulmonary vascular resistance and improved RV performance
in a porcine chronic pulmonary hypertension model. A further study
indicated that nebivolol, a β3-adrenergic agonist, reduced the
overexpression of growth and inflammatory mediators in pulmonary
vascular cells harvested from patients with PAH (21). However, BRL37344 and nebivolol are
not selective β3-AR agonists, therefore their effects may result
from the stimulation of alternative β-ARs (22,23).
Apart from a limited number of studies using β3-AR antagonists to
block the effect of β3-AR agonists (24,25), no
studies have been reported to investigate the antagonism of β3-AR
alone in PAH.
The present study established a rat PAH model, which
was treated with the selective β3-AR antagonist, SR59230A, to
investigate the functional involvement of β3-AR in hemodynamic and
morphological impairment in PAH, and identify novel therapeutic
targets. The generation of two isoforms of NOS following β3-AR
inhibition were also investigated, investigating the potential
signaling pathways of β3-AR in PAH.
Materials and methods
Animals
In total, 12 male Sprague-Dawley rats (weight,
250–300 g; age, 8 weeks) were purchased from the Second Affiliated
Hospital of Harbin Medical University (Harbin, China). The present
study was ethically approved by the Harbin Medical University
Committee on Animal Care (26) and
was performed in adherence with the National Institutes of Health
Guidelines on the Use of Laboratory Animals (27). The rats were randomly assigned to
three groups receiving: i) An equal volume of solvent (vehicle, 1
ml/kg body weight); ii) a single subcutaneous injection of
monocrotaline (MCT; 80 mg/kg body weight; Sigma-Aldrich; Merck
KGaA) to induce PAH within 4 weeks; or iii) a single injection of
MCT and injections of SR59230A (2 mg/kg body weight; Sigma-Aldrich;
Merck KGaA) every other day for 4 weeks. Injections of SR59230A or
an equal volume of solvent were administered via the tail vein of
the rat.
In vivo Doppler echocardiography
Subsequent to 4 weeks of MCT administration, the
rats were anesthetized using isoflurane. The isoflurane vaporizer
was adjusted to 3–5% for induction and 1–3% for maintenance. The
animals were stabilized in the supine position, and M mode echo
recordings from the parasternal long-and short axes were collected.
All measurements were subsequently analyzed using Vevo 770 software
(Fujifilm VisualSonics Inc.).
Tissue fixation, embedding and
sectioning
Following the in vivo Doppler
echocardiograph, the rats were euthanized by continuous isoflurane
exposure (5%). Following dissection, rat RV and left lung tissues
were immediately fixed in 4% formaldehyde solution for 24 h at room
temperature. The tissues were dehydrated by immersion in increasing
concentrations of ethanol (from 70 to 90%, and then from 90 to 100%
ethanol). Each dehydration step lasted for 1 h. The dehydrating
agent was subsequently cleared by incubation in xylene for 2 h at
room temperature prior to paraffin embedding. Paraffin was
typically heated to 60°C and then allowed to harden overnight, and
the tissues were sectioned to 8 µm using a microtome.
Histological
characterization-hematoxylin and eosin staining
Tissue sections were deparaffinized through three
changes of xylene and hydrated using distilled water. Subsequent to
rinsing, the slides were stained with hematoxylin for 2 min, rinsed
again, dipped in ammonia hydroxide solution (0.2%) until a blue
color change was observed, and washed once more. The slides were
then counterstained with eosin for 10 min and dehydrated with 95%
ethanol, followed by three changes of 100% ethanol. The dehydrated
slides were then cleared with three changes of xylene and mounted
onto a coverslip with Neutral Canada balsam. All the staining
solutions were incubated with the samples at room temperature.
Immunohistochemistry (IHC) and
semi-quantitative analysis
Tissue sections were incubated three times in xylene
(5 min each at room temperature), followed by two washes each with
100, 95 and 80% ethanol for 10 min, and two washes in distilled
water for 5 min each. Following deparaffinization and rehydration,
antigen retrieval was performed using a pressure cooker. First, the
appropriate antigen retrieval buffer (10 mM sodium citrate, 0.05%
Tween-20, pH 6.0) was added to the pressure cooker. Once the
boiling temperature was reached (95-100°C), the sections were then
submerged using forceps and the pressure cooker lid was secured.
The cooker as allowed to reach maximum pressure prior to
depressurization, and was subsequently filled with cold water for
10 min. The sections were then transferred to phosphate-buffered
saline (PBS) prior to incubation with a blocking buffer [1X PBS, 5%
goat serum (cat. no. ab7481; Abcam) and 0.3% Triton™ X-100] at room
temperature for 30 min. Subsequently, the sections were incubated
with an anti-β3-AR primary antibody (cat. no. ab94506; Abcam;
working concentration, 10 µg/ml) overnight at 4°C. The sections
were washed three times in PBS for 5 min each, and incubated with a
secondary antibody anti rabbit (cat. no. PV-9001; dilution,
1:2,000; Beijing Zhongshanjinqiao Biotechnology Co., Ltd.) in the
dark at 37°C for 20 min. Following three additional washes in PBS,
the sections were incubated in 3,3′-diaminobenzidine
(DAB)-peroxidase substrate solution for 5–10 sec (cat. no.
ZLI-9017; Beijing Zhongshanjinqiao Biotechnology Co., Ltd.), and
washed in PBS once again. The sections were then counterstained
with hematoxylin for 2 min at room temperature, dehydrated through
two changes of 95% ethanol and three changes of 100% ethanol,
cleared with three washes in xylene and mounted on a coverslip with
Neutral Canada balsam.
The IHC images were obtained (Olympus, BX51; light
microscope; magnification, ×200) and analyzed using the FIJI
analysis software (ImageJ win64; National Institutes of Health,
Bethesda, MD, USA). Run Image > Color > Color Deconvolution
was performed and ‘H DAB’ from the Vectors pull-down was selected
as the stain. The three image windows (colors 1, 2 and 3)
corresponded to hematoxylin, DAB and residual (which should be
close to white if the separation is optimal), respectively. DAB
positivity was quantified in units of intensity and normalized to
the Sham group.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the frozen lung tissue
samples (50–100 mg) using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) and isolated using the
RNeasy® Plus Mini kit (Qiagen GmbH). The RNA quality and
concentration were measured using a Nanodrop 2000 (NanoDrop
Technologies; Thermo Fisher Scientific, Inc.). Total RNA (2 µg) was
reverse transcribed into cDNA using the High Capacity cDNA Reverse
Transcription kit (Invitrogen; Thermo Fisher Scientific, Inc.). The
RT reaction was incubated at 25°C for 10 min, at 37°C for 2 h and
at 85°C for 5 min. The mRNA expression levels of the β3-AR, eNOS
and inducible NOS (iNOS) were determined by RT-qPCR. The
thermocycling conditions were as follows: Initial denaturation at
95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec, 55°C for
10 sec and 72°C for 15 sec. qPCR was performed using a reaction
mixture containing SYBR® Green dye (Invitrogen; Thermo
Fisher Scientific, Inc.) on a StepOnePlus PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Using these systems,
relative alterations in mRNA levels were calculated according to
the ΔΔCq formula (28). β-actin was
used as an internal control. The primer sequences are listed in
Table I.
 | Table I.Reverse transcription-polymerase
chain reaction primers sequences. |
Table I.
Reverse transcription-polymerase
chain reaction primers sequences.
| Gene | Primers |
|---|
| β3-AR | F:
AGAACTCACCGCTCAACAGG |
|
| R:
CAGAAGTCAGGCTCCTTGCTA |
| eNOS | F:
GAAGACTGAGACTCTGGCCC |
|
| R:
CCGTGGGGCTTGTAGTTGAC |
| iNOS | F:
GGCTGAGTACCCAAGCTGAG |
|
| R:
ATTGTGGCTCGGGTGGATTT |
| β-Actin | F:
CACTTTCTACAATGAGCTGCG |
|
| R:
CTGGATGGCTACGTACATGG |
Western blotting
The lung tissues were homogenized and total protein
was extracted using ice-cold radioimmunoprecipitation assay buffer
with protease inhibitor mix (Sigma-Aldrich; Merck KGaA). Protein
was quantified using a bicinchoninic acid protein assay kit (Thermo
Fisher Scientific, Inc.) Equivalent amounts of protein (50 ug/well)
were separated using SDS-PAGE with a 12% gel, and the proteins were
transferred to polyvinylidene difluoride membranes, which were
blocked using Tris-buffered saline with TBS-Tween (0.1% Tween-20)
with 5% non-fat milk for 1 h. The membranes were then incubated
with primary antibodies against eNOS (cat. no. ab5589; 1:1,000;
Abcam) and iNOS (cat. no. ab15323; 1/250; Abcam) at 4°C overnight
and washed 3 times with TBST prior to incubation with horseradish
peroxidase (HRP)-conjugated secondary antibodies (cat. no. ab6721;
1:2,000; Abcam) at room temperature for 1 h. β-actin was used as an
internal control (cat no. ab115777; 1:200; Abcam). The blots were
washed and developed using a HRP substrate solution (EMD
Millipore), and the ChemiDoc XRS+ system (Bio-Rad Laboratories,
Inc.) was used for imaging and FIJI analysis software (ImageJ
win64; National Institutes of Health) was used for image processing
and analysis. Each experiment was repeated in triplicate.
Statistical analysis
All data were analyzed using GraphPad Prism 6
(GraphPad Software, Inc.) and expressed as the mean ± standard
error of the mean. One-way analysis of variance was used to compare
the differences between groups, and P<0.05 was considered to
indicate a statistically significant difference.
Results
β3-AR antagonist SR59230A improves RV
performance in an MCT-induced PAH rat model
Echocardiography was performed on each rat and the
heart function was evaluated in all groups 4 weeks subsequent to
MCT administration; representative M-mode echocardiographic images
(Fig. 1A) reveal the RV function and
structure in the three groups. Significant increases in RV basal
diameter (P<0.001), RV wall thickness (RVWT; P<0.001) and E/A
ratio [the ratio of peak velocity flow in early diastole (the E
wave) to peak velocity flow in late diastole caused by atrial
contraction (the A wave)] (P<0.01), and a significant decrease
in the RV ejection fraction (EF) (P<0.01) were observed in
MCT-injected rats compared with the sham group, indicating that the
MCT-induced PAH model had been successfully established. The MCT
rats that had received SR59230A (MCT+SR) exhibited a significant
decrease in RV basal diameter, E/A ratio (P<0.01) and RVWT
(P<0.05), with considerable recovery of the EF (P<0.01)
compared with the MCT-induced PAH rats (Fig. 1B-E). In addition, SR59230A treatment
significantly reduced the left atrial diameter compared with the
MCT group (P<0.001), which was significantly higher (P<0.001)
in the MCT-induced PAH model compared with the sham group (Fig. 1F). Furthermore, compared with the
sham rats, MCT-induced PAH rats displayed slight, but significant
(P<0.01) decreases in left ventricular internal diameter (LVID)
systole (LVIDs), LVID diastole (LVIDd), left ventricular posterior
wall thickness at end-diastole (LVPWd) and interventricular septal
end diastole (IVSd), all of which were slightly recovered following
SR59230A administration (P<0.05; Fig.
1G-I).
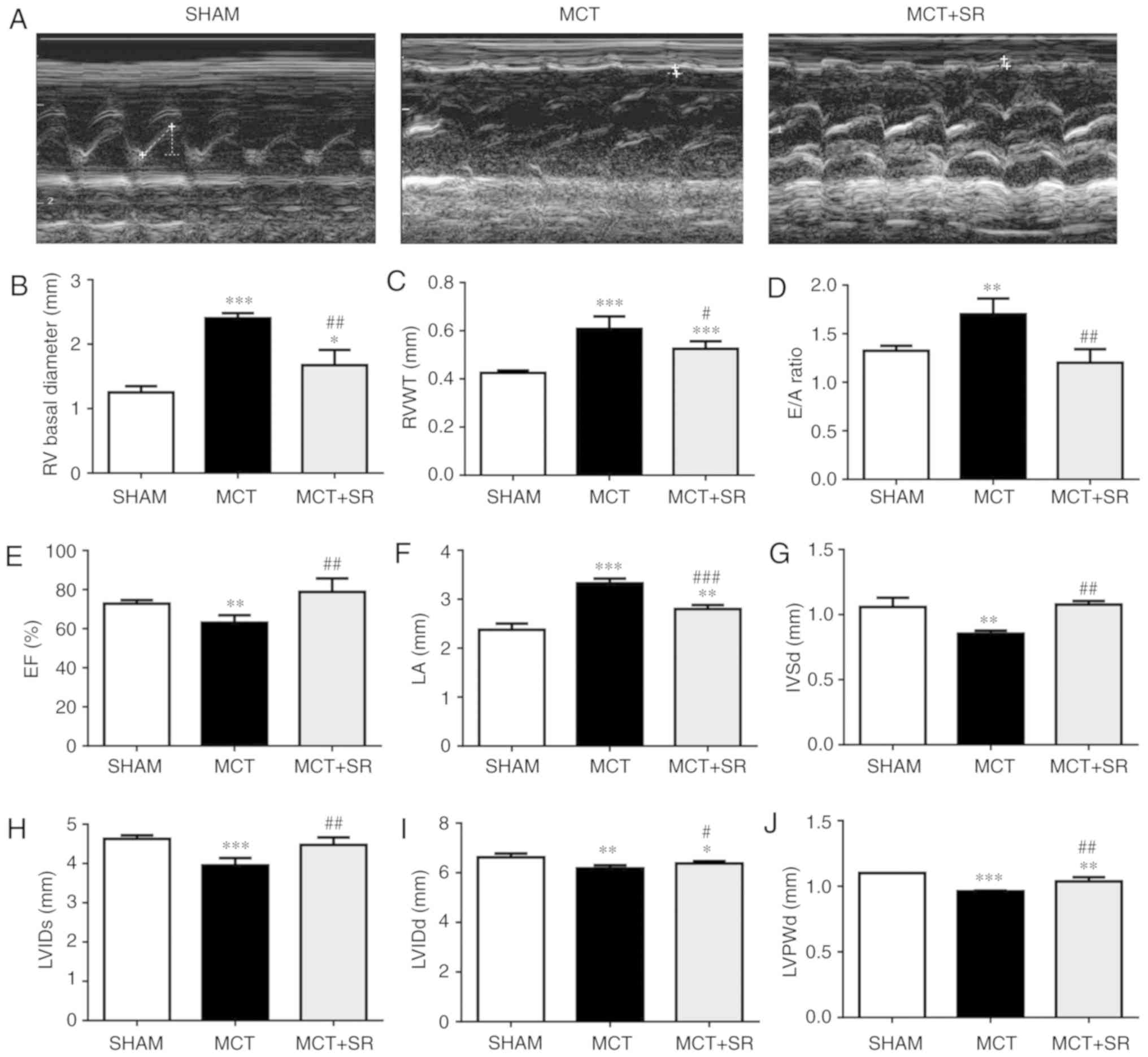 | Figure 1.β3-AR antagonist SR59230A improves
right ventricular performance in MCT-induced PAH. (A)
Representative M-mode echocardiographic images were taken at the
midventricular level (parasternal long axis). Quantification of (B)
basal RV diameter, (C) RVWT, (D) E/A ratio, (E) right ventricular
EF, (F) LA diameter, (G) IVSd, (H) LVIDs, (I) LVIDd and (J) LVPWd
from sham, MCT and MCT+SR rats 4 weeks following MCT
administration. N=4 per group. *P<0.05, **P<0.01 and
***P<0.001 vs. Sham. #P<0.05,
##P<0.01 and ###P<0.001 vs. the MCT
group. β3-AR, β3-adrenergic receptor; PAH, pulmonary artery
hypertension; RV, right ventricular; RVWT, right ventricular wall
thickness; EF, ejection fraction; LA, left atrial; IVSd,
Interventricular septal end diastole; LVIDs, left ventricular
internal diameter end systole; LVIDd, left ventricular internal
diameter end systole; LVPWd, left ventricular posterior wall
thickness at end-diastole; MCT, monocrotaline; MCT+SR, MCT +
SR59230A. |
SR59230A reduces MCT-induced RV
hypotrophy
Histological examination of hematoxylin and
eosin-stained RV sections revealed that MCT administration resulted
in considerable alterations to the myocardial structure in rats
(Fig. 2). RV samples from healthy
control rats exhibited the cardiac syncytia of myocardial fibers
with central nuclei, while RV from the MCT-treated rats exhibited
contracture cardiomyocyte damage, hypereosinophilia, wavy
arrangement of the myofibers and cardiomyocyte apoptosis. Moderate
macroscopic hypertrophy and reduced inflammatory cell infiltration
were observed in sections from the MCT+SR59230A treatment group
compared with the MCT-induced PAH group.
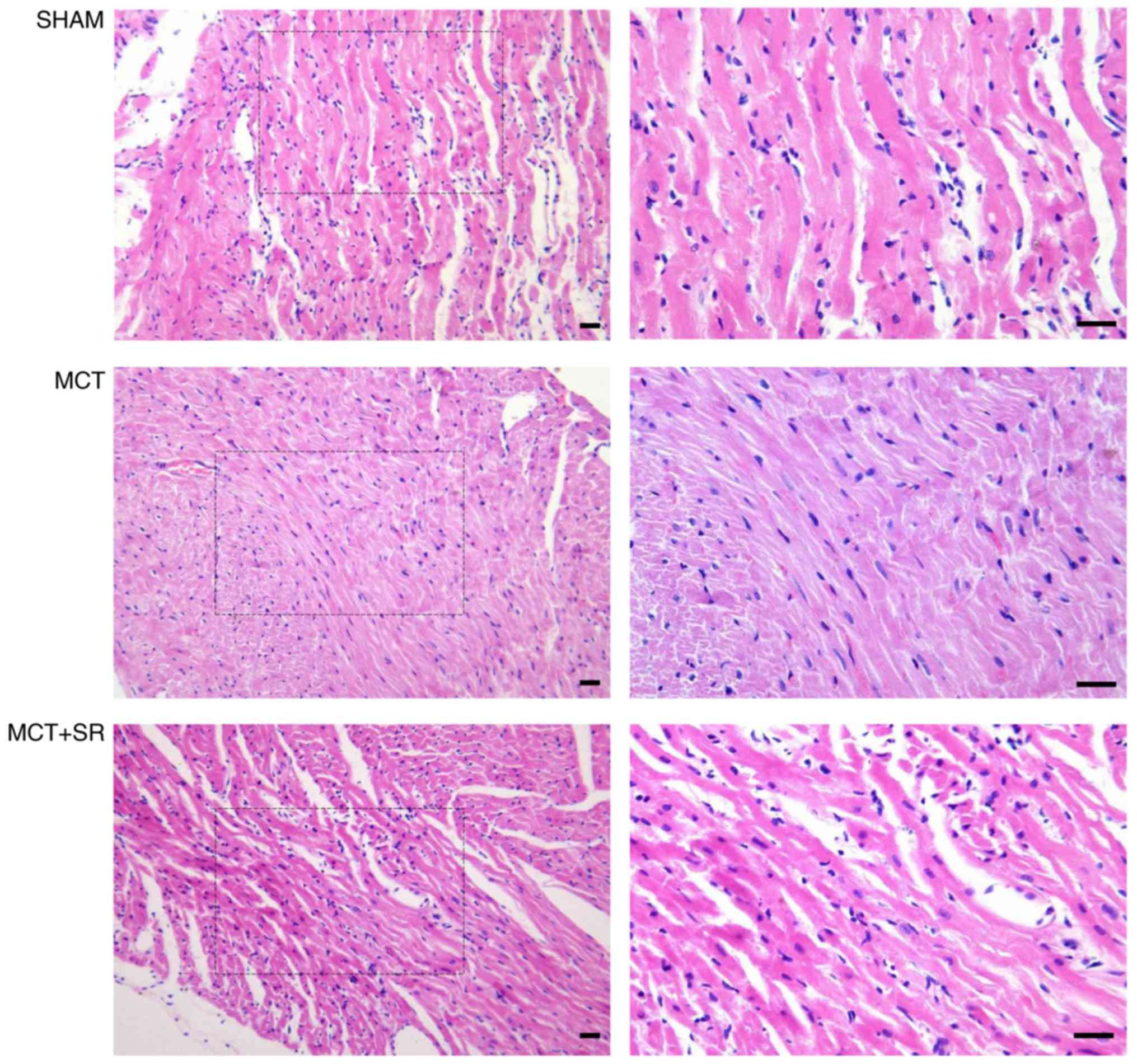 | Figure 2.Histological evaluation of right
ventricular sections of the PAH rats. Representative images of the
right ventricle stained with hematoxylin and eosin. Left, original
magnification, ×200; right, original magnification, ×400, magnified
images of the boxed areas; bar, 20 µm. Contracture of
cardiomyocytes, hypereosinophilia, wavy arrangement of myofibers
and cardiomyocyte apoptosis were observed in MCT-induced PAH
samples. MCT+SR samples exhibited reduced myocardial disarray. PAH,
pulmonary artery hypertension; MCT, monocrotaline; MCT+SR, MCT +
SR59230A. |
SR59230A reduces MCT-induced alveolar
expansion and inflammatory infiltration
Severe pulmonary vascular remodeling was observed in
the lung sections of the MCT-induced PAH rats. Compared with the
control group, samples from the MCT group exhibited emphysematous
expansion of the alveoli, discontinuous endothelium in the
pulmonary arterioles, smooth muscle cell hypotrophy and
disarrangement, thickening of the vessel wall, a narrowed vascular
lumen and dense inflammatory infiltration in a perivascular and
peribronchial distribution. Following SR59230A administration,
reduced emphysematous expansion of the alveoli and inflammatory
infiltration were observed in a perivascular and peribronchial
distribution compared with the MCT group. However, MCT-induced
pulmonary artery smooth muscle cell hypotrophy and disarrangement
did not exhibit obvious improvement following SR59230A treatment
(Fig. 3).
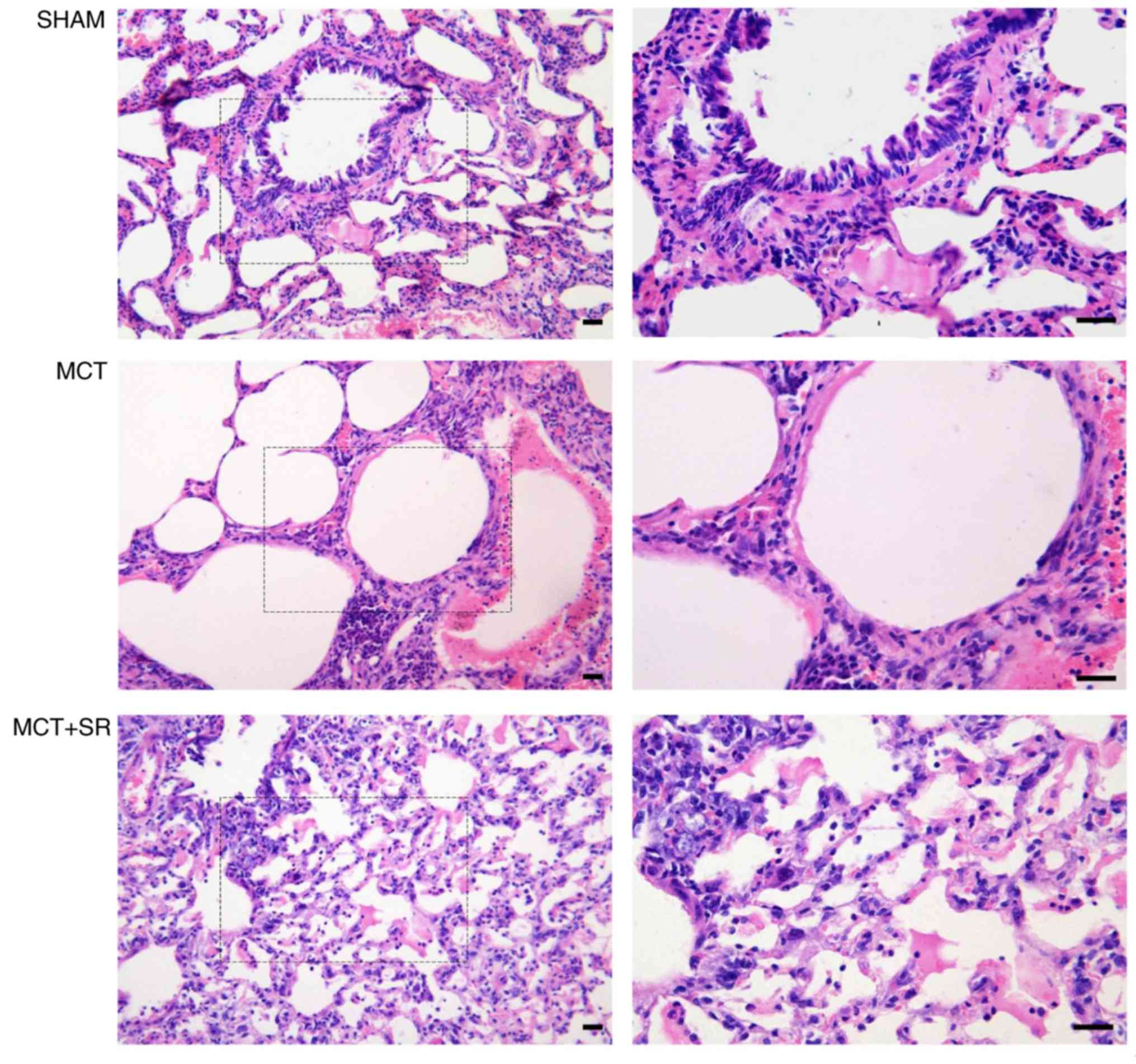 | Figure 3.Histological evaluation of lung
sections from PAH rats. Representative images of the left lung
stained with hematoxylin and eosin. Left, original magnification,
×200; right, original magnification, ×400, magnified images of the
boxed areas; bar, 20 µm. The diameters of the pulmonary alveoli
were increased upon MCT administration, with remodeled bronchioles
and vessels surrounded by a dense infiltration of inflammatory
cells, which were alleviated in the MCT+SR group. PAH, pulmonary
artery hypertension; MCT, monocrotaline; MCT+SR, monocrotaline +
SR59230A. |
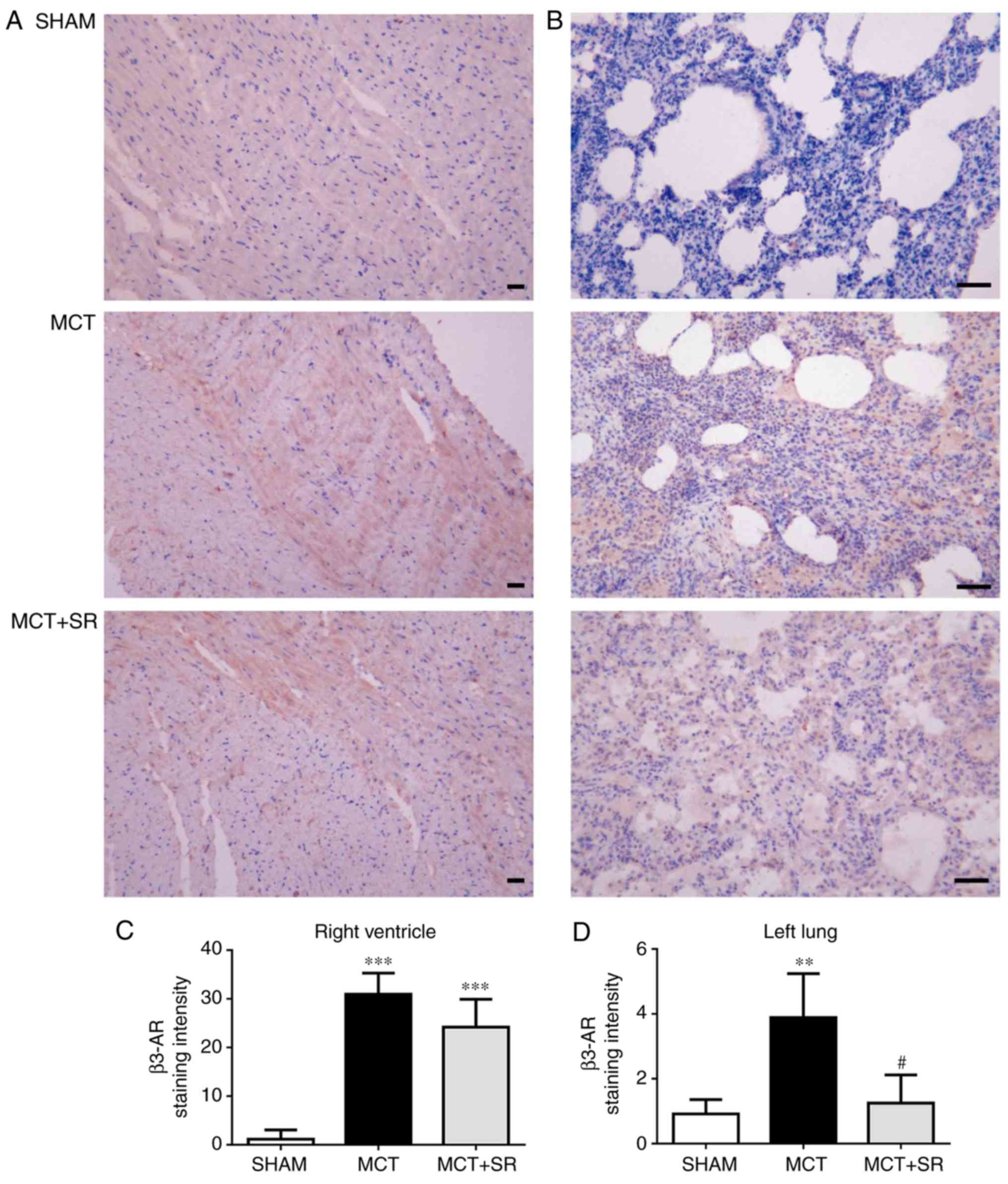
SR59230A suppresses the increase of
β3-AR in the lung tissue, but not the RV
To investigate the expression level of β3-AR in the
lung and heart tissues, IHC staining of β3-AR was conducted in
samples from all three groups. In MCT-induced PAH rats, a
significant elevation in β3-AR expression levels was revealed in
the RV and lung tissues compared with the sham group (P<0.01).
Following 4 weeks of SR59230A treatment, no significant change in
β3-AR was observed in the RV (Fig. 4A
and C), but a significant reduction was observed in the left
lung (P<0.05; Fig. 4B and D).
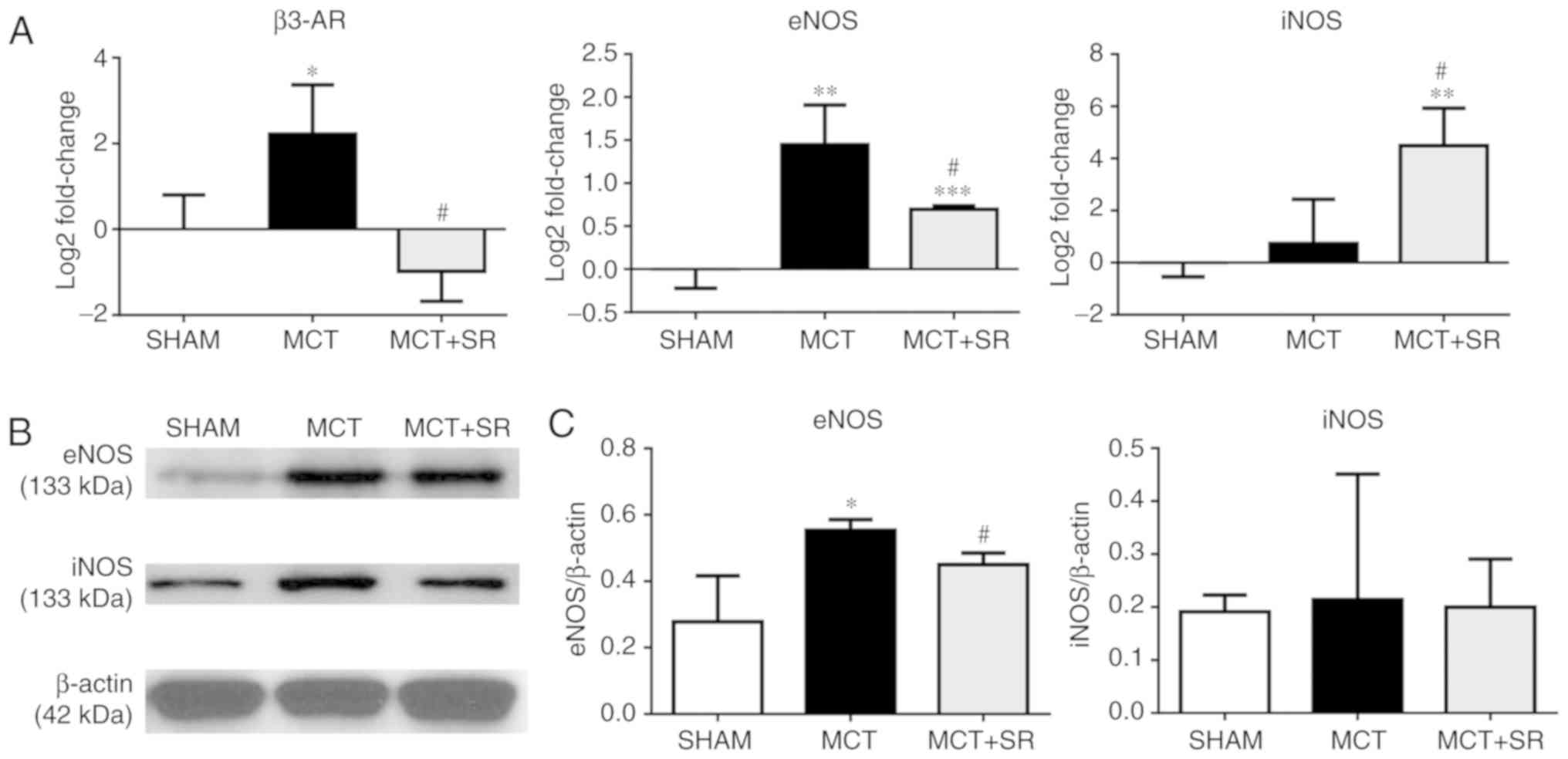
SR59230A inhibits eNOS induction in
the lung
To determine whether β3-AR is involved in regulating
the NO signaling pathway in MCT-induced PAH, the expression levels
of β3-AR, eNOS and iNOS in the lung were determined. RT-qPCR
revealed a significant increase in the expression level of eNOS and
an upregulation in β3-AR in the MCT-induced PAH group compared with
the sham group (P<0.05), which was consistent with the IHC data.
Four weeks of SR59230A treatment significantly reversed the
increase in eNOS expression level (P<0.05); though iNOS
expression was not significantly altered in the MCT-induced PAH
model compared with the sham, there was a significant induction
following SR59230A administration (P<0.05; Fig. 5A). At the protein level, western
blotting revealed that the eNOS expression level was similarly
increased (P<0.05), but to a lesser degree than the change in
gene expression (P<0.05). However, no significant change in iNOS
expression levels were observed in the SR59230A-treated group
(Fig. 5B and C).
Discussion
The present study initially investigated the
cardio-protective effects of a β3-AR antagonist in PAH. Treatment
with the β3-AR antagonist SR59230A improved RV performance in
MCT-induced PAH rats, and alleviated the structural impairment and
inflammatory cell infiltration to the RV and lung. Furthermore, the
presence of β3-AR in rat lung tissues was confirmed, in addition to
the observed overexpression of β3-AR in the RV and lung in PAH.
Long-term administration of SR59230A significantly reduced the
expression level of β3-AR in the lung compared with the group
without SR59230 treatment, but not the RV. Furthermore, the
production of eNOS in the PAH lung was associated with changes in
the expression level of β3-AR, which was inhibited by SR59230A.
MCT is a pyrrolizidine alkaloid. Its bioactive
metabolite causes selective damage to the vascular endothelium of
the lung vessels, resulting in an increase in vascular resistance
and consequently resulting in high pulmonary arterial pressure
(29). The increase in RV-afterload
not only induced right ventricular dilation and failure (as
indicated by a marked elevation in RV basal diameter, RVWT, E/A
ratio and a marked decrease in EF), but also affected the structure
and function of the left heart, characterized by reduced LVIDs,
LVIDd and LVPWd. In the present study, it was demonstrated that
long-term treatment with SR59230A significantly suppressed the
MCT-induced increase in RV basal diameter and RVWT. Also, right
ventricular EF was recovered and the E/A ratio, a recommended
parameter to evaluate RV diastolic function (30), was revealed to approach the normal
level following SR59230A therapy. These results contradict previous
reports, which revealed the beneficial effects of the β3-AR agonist
in a number of pulmonary hypertension models (20,21).
These controversies may be partly due to the differences between
experimental models; one study used pigs with chronic PH
established by the surgical banding of the main pulmonary vein
(20), whilst another established a
rat PAH model using a lower dose of MCT (60 mg/kg) (21). In addition, it should be noted that
the majority of previous studies used non-selective β3-AR agonists;
therefore the possibility that the observed beneficial effects may
be caused by other β-adrenergic functions (including β2-AR
stimulation) should not be excluded (20,22).
Conversely, one other study identified the involvement of β1/β2-AR
in the development of PAH, and that the α1/β1/β2-AR blocker
carvedilol improved RV function and prevented maladaptive
myocardial remodeling in rats with severe experimentally-induced
PAH. However, concerning side effects, including a large decrease
in heart rate, remained (31).
Considering that, to the best of our knowledge, no β3-AR antagonist
had ever been studied alone in PAH, the present study utilized
SR59230A treatment and was the first to reveal the
cardio-protective effect of a β3-AR antagonist on an MCT-induced
PAH model in vivo.
In contrast to a number of other classical β-AR
antagonists, SR59230A has a higher affinity for β3-AR, while it has
also been identified to have a similar affinity for other β-ARs
(32). By contrast to β1/2AR, β3-AR
was revealed to be activated at higher concentrations of
catecholamines, and preferentially activated in situations of high
adrenergic tone, including chronic heart failure, sepsis and
diabetes (33,34). The results revealed an increase in
β3-AR expression level in the heart in MCT-induced PAH, though this
elevation did not change substantially upon SR59230A treatment.
Various reports have indicated that β3-AR antagonists blocked β3-AR
activation, therefore preventing the suppression of cardiac
contraction and reversing the progression of cardiac dysfunction
(35,36). However, only one previous study
evaluated the β3-AR expression levels in β3-AR antagonist
application (37). In a rat model of
isoproterenol-induced heart failure, SR59230A ameliorated cardiac
function resulting in reduced β3-AR expression levels. Combined
with these results, the results of the present study suggest that
the cardio-protective effects of SR59230A in PAH may not be due to
its direct effect in controlling the induction of β3-AR in the
heart. Alternatively, a higher level of β3-AR expression in PAH may
increase the binding affinity of SR59230A, therefore failing to
activate the β3-AR response in the heart (38,39).
Considering the differences between experimental models, it is
necessary to further investigate the association between β3-AR
expression levels and β3-adrenergic effects in the heart under PAH
conditions. On the other hand, numerous studies have revealed that
SR59230A possesses partial or full β3-AR-agonist properties
(40,41), but in the present study, no evidence
was identified to suggest that SR59230A exhibited β3-AR agonistic,
as opposed to antagonistic, properties.
In the present study, SR59230A administration
significantly downregulated β3-AR expression levels in the lung
compared with the group without SR59230 treatment, but not the
heart; this is consistent with the belief that the factors
regulating β3-AR vary between species, tissues and diseases stages
(4). In addition, the results also
revealed that in MCT-induced PAH, alveolar and vascular endothelial
damage were accompanied by the overexpression of β3-AR, and that
SR59230A treatment inhibited β3-AR expression in the lung,
alleviating the expansion of the alveoli and inhibiting
inflammatory infiltration to the perivascular and peribronchial
area. These results suggest that the increase in β3-AR expression
levels in PAH may be responsible for the progressive impairment of
alveolar structure and chronic inflammation. Inflammatory
infiltration has frequently been observed surrounding remodeled
vessels, including plexiform lesions in human PAH (42,43),
with an elevated level of pro-inflammatory cytokines including
interleukin-6 (IL-6) in circulation and in the lung tissue;
specifically, β3-AR stimulation induced IL-6 release from
adipocytes (44,45), and β2/3-AR may be responsible for the
increase in plasma IL-6 observed during sympathetic activation and
in obesity, as opposed to macrophages (46). However, there is currently no
evidence to confirm the association between the elevation of IL-6
and the β3 adrenergic responses in experimental models or patients
with PAH. Notably, the alleviation of lung remodeling and the
reduction of inflammatory infiltration to the pulmonary vasculature
following SR59230A therapy may influence RV chronic pressure
overload and improve RV function (47,48).
Endothelial-derived NO is a recognized mediator of
angiogenesis in pulmonary circulatory vessels. However, the
function of NO in regulating pulmonary vascular tone remains
unclear (49). Studies have reported
that disruption of the eNOS gene and NO administration were
associated with altered pulmonary arterial pressure in chronically
hypoxic conditions (50,51). Additionally, iNOS deficiency
contributed to increased basal pulmonary vascular tone, whereas
deficiency of neuronal NOS (nNOS) had no such effect (52). In particular, β3-AR was demonstrated
to modulate NO signaling via eNOS, nNOS and iNOS in the heart and
vasculature under either pathological conditions or pharmacological
stimulation (5,53,54).
Furthermore, another study demonstrated that SR59230A inhibited
nebivolol-induced NO in isolated heart tissues, suggesting a
potential function for β3-AR in regulating iNOS-dependent NO
production (54). The dysfunction of
eNOS in endothelial cells may indicate an important source for the
production of reactive oxygen species (ROS) in pulmonary
hypertension, so-called eNOS uncoupling, which has been observed
between β3-AR and eNOS in the failing human myocardium (55,56). In
order to investigate the effects of β3-AR antagonism in PAH (via
NOS regulation), the expression levels of eNOS and iNOS in the lung
were determined in the present study. The results illustrated a
marked increase in the eNOS expression level at the transcriptional
and translational level in an MCT-induced PAH model compared with
the sham group, and a significant decrease in eNOS following
SR59230A administration compared with no administration group. The
change in eNOS expression level in PAH was consistent with a
previous study reporting an increase in eNOS-encoding mRNA in the
MCT rat artery (57), but
inconsistent with another study reporting no change in MCT-induced
PAH (58). These results questioned
whether elevated eNOS expression was a compensatory response to
satisfy the requirement of NO during PAH, or a sign of oxidative
stress (59).
Dissociation between the expression of eNOS and NO
was reported in MCT-induced PAH rats (57). Increased ROS, an altered redox state
and elevated oxidant stress have been indicated in the lung and RV
of various PAH animal models, in addition to MCT toxicity (60). Furthermore, a more recent study
revealed that a deficiency in tetrahydrobiopterin, an essential
cofactor for eNOS coupling, may be associated with MCT-induced PAH
rats (61). In addition, SR59230A
may inhibit inflammatory infiltration in the perivascular and
peribronchial regions, whereas inflammatory cell recruitment has
been known to accelerate the accumulation of ROS and lung
functional impairment (62).
Therefore, the results of the present study suggested that eNOS
induction in PAH may result in the generation of superoxides as
opposed to NO. β3-AR antagonists may alleviate progressive lung
injury by inhibiting inflammatory infiltration and preventing
oxidative stress by regulating the generation of uncoupled eNOS
(55).
In addition, no significant change in iNOS was
demonstrated in PAH rats compared with the sham control in the
present study, which is inconsistent with a previous study in a
chronic hypoxia-induced PAH model; this identified a transient
induction of iNOS in the pulmonary vascular wall at the initial
stages of hypoxia, which returned to more characteristically low
levels 20 days later (63). As
anticipated, the present study indicated that SR59230A induced a
marked increase in the expression level of iNOS mRNA, but not
protein. Since changes in gene expression cannot be accounted for
by transcription alone, this highlighted that SR59230A itself may
be involved in the post-transcriptional or translational regulation
of iNOS, which requires further investigation. Additional
limitations include the fact that MCT-induced PAH rats may not
represent the entire spectrum of PAH models, as only a single time
point was observed. However, the present study aimed to evaluate
the effect of a β3-AR antagonist on RV function, as the
cardio-protective effect of β3-AR agonists and other drug have
previously been studied (21,64). RV
basal diameter, RVWT, E/A ratio and EF were selected to evaluate RV
structure and function under PAH conditions, but due to technical
limitations, the pulmonary artery pressure value, Tricuspid Annular
Plane Systolic Excursion and fractional area change values
(65) were not obtained. Therefore,
it is necessary in future studies to include these parameters.
Furthermore, phospho-eNOS and phospho-iNOS antibodies were not
included in the western blotting. This may have indicated whether
the translocation or phosphorylation of eNOS was involved in the
different mechanisms of β3-AR-stimulated eNOS activation, according
to the associated anatomical region (66,67).
Therefore, further studies evaluating the eNOS activation pathway
in the lung are warranted. Last but not least, since only MCT
induced PAH models were studied in this manuscript, the effects of
β3-AR blockage in other PAH models, for example, in hypoxia rats,
remain unknown. Future studies on different PAH models are required
to understand the involvement of the β3-AR response in the
initiation and development of PAH.
In conclusion, SR59230A, a selective β3-AR
antagonist, exerted beneficial effects on RV performance in a rat
model of MCT-induced PAH. Also, inhibiting β3-AR with SR59230A may
alleviate structural changes and inflammatory infiltration by
reducing oxidative stress in the lung.
Acknowledgements
Not applicable.
Funding
The present study was supported by Harbin Medical
University (grant no. 2013SYYRCYJ06).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
JS and WL designed the experiment. JS performed
research and analyzed the data. XD, JC and JY contributed to the
interpretation of the results. JS, JLC and WL wrote the manuscript.
The final version of the manuscript has been read and approved by
all authors and each author believes that the manuscript represents
honest work.
Ethics approval and consent to
participate
All experimental procedures were ethically approved
by the Harbin Medical University Committee on Animal Care and were
performed in adherence with the National Institutes of Health
Guidelines on the Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
β3-AR
|
β3 adrenergic receptor
|
|
EF
|
ejection fraction
|
|
eNOS
|
endothelial nitric oxide synthase
|
|
IHC
|
immunohistochemistry
|
|
iNOS
|
inducible nitric oxide synthase
|
|
IVSd
|
interventricular septal end
diastole
|
|
LVIDd
|
left ventricular internal diameter end
diastole
|
|
LVIDs
|
left ventricular internal diameter end
systole
|
|
LVPWd
|
left ventricular posterior wall
thickness at end-diastole
|
|
MCT
|
monocrotaline
|
|
PAH
|
pulmonary arterial hypertension
|
|
PH
|
pulmonary hypertension
|
|
RV
|
right ventricle
|
|
RVWT
|
right ventricular wall thickness
|
References
|
1
|
Xu D, Guo H, Xu X, Lu Z, Fassett J, Hu X,
Xu Y, Tang Q, Hu D, Somani A, et al: Exacerbated pulmonary arterial
hypertension and right ventricular hypertrophy in animals with loss
of function of extracellular superoxide dismutase. Hypertension.
58:303–309. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Provencher S and Granton JT: Current
treatment approaches to pulmonary arterial hypertension. Can J
Cardiol. 31:460–477. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vachiery JL and Gaine S: Challenges in the
diagnosis and treatment of pulmonary arterial hypertension. Eur
Respir Rev. 21:313–320. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rozec B and Gauthier C:
Beta3-adrenoceptors in the cardiovascular system: Putative roles in
human pathologies. Pharmacol Ther. 111:652–673. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moens AL, Yang R, Watts VL and Barouch LA:
Beta 3-adrenoreceptor regulation of nitric oxide in the
cardiovascular system. J Mol Cell Cardiol. 48:1088–1095. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gauthier C, Tavernier G, Charpentier F,
Langin D and Le Marec H: Functional beta3-adrenoceptor in the human
heart. J Clin Invest. 98:556–562. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Niu X, Watts VL, Cingolani OH, Sivakumaran
V, Leyton-Mange JS, Ellis CL, Miller KL, Vandegaer K, Bedja D,
Gabrielson KL, et al: Cardioprotective effect of beta-3 adrenergic
receptor agonism: Role of neuronal nitric oxide synthase. J Am Coll
Cardiol. 59:1979–1987. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tavernier G, Toumaniantz G, Erfanian M,
Heymann MF, Laurent K, Langin D and Gauthier C: Beta3-Adrenergic
stimulation produces a decrease of cardiac contractility ex vivo in
mice overexpressing the human beta3-adrenergic receptor. Cardiovasc
Res. 59:288–296. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao Q, Zeng F, Liu JB, He Y, Li B, Jiang
ZF, Wu TG and Wang LX: Upregulation of β3-adrenergic receptor
expression in the atrium of rats with chronic heart failure. J
Cardiovasc Pharmacol Ther. 18:133–137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gauthier C, Langin D and Balligand JL:
Beta3-adrenoceptors in the cardiovascular system. Trends Pharmacol
Sci. 21:426–431. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bhadada SV, Patel BM, Mehta AA and Goyal
RK: β(3) Receptors: Role in cardiometabolic disorders. Ther Adv
Endocrinol Metab. 2:65–79. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rasmussen HH, Figtree GA, Krum H and
Bundgaard H: The use of beta3-adrenergic receptor agonists in the
treatment of heart failure. Curr Opin Investig Drugs. 10:955–962.
2009.PubMed/NCBI
|
|
13
|
Feng MG, Prieto MC and Navar LG:
Nebivolol-induced vasodilation of renal afferent arterioles
involves beta3-adrenergic receptor and nitric oxide synthase
activation. Am J Physiol Renal Physiol. 303:F775–F782. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rozec B, Serpillon S, Toumaniantz G, Sèze
C, Rautureau Y, Baron O, Noireaud J and Gauthier C:
Characterization of beta3-adrenoceptors in human internal mammary
artery and putative involvement in coronary artery bypass
management. J Am Coll Cardiol. 46:351–359. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pietri-Rouxel F and Strosberg AD:
Pharmacological characteristics and species-related variations of
beta 3-adrenergic receptors. Fundam Clin Pharmacol. 9:211–218.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moniotte S and Balligand JL: Potential use
of beta(3)-adrenoceptor antagonists in heart failure therapy.
Cardiovasc Drug Rev. 20:19–26. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pourageaud F, Leblais V, Bellance N,
Marthan R and Muller B: Role of beta2-adrenoceptors (beta-AR), but
not beta1-, beta3-AR and endothelial nitric oxide, in
beta-AR-mediated relaxation of rat intrapulmonary artery. Naunyn
Schmiedebergs Arch Pharmacol. 372:14–23. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tagaya E, Tamaoki J, Takemura H, Isono K
and Nagai A: Atypical adrenoceptor-mediated relaxation of canine
pulmonary artery through a cyclic adenosine monophosphate-dependent
pathway. Lung. 177:321–332. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu Y, Fuscoe JC, Zhao C, Guo C, Jia M,
Qing T, Bannon DI, Lancashire L, Bao W, Du T, et al: A rat RNA-Seq
transcriptomic BodyMap across 11 organs and 4 developmental stages.
Nat Commun. 5:32302014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
García-Aívarez A, Pereda D, Garcia-Lunar
I, Sanz-Rosa D, Fernández-Jiménez R, García-Prieto J, Nuño-Ayala M,
Sierra F, Santiago E, Sandoval E, et al: Beta-3 adrenergic agonists
reduce pulmonary vascular resistance and improve right ventricular
performance in a porcine model of chronic pulmonary hypertension.
Basic Res Cardiol. 111:492016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Perros F, Ranchoux B, Izikki M, Bentebbal
S, Happé C, Antigny F, Jourdon P, Dorfmüller P, Lecerf F, Fadel E,
et al: Nebivolol for improving endothelial dysfunction, pulmonary
vascular remodeling and right heart function in pulmonary
hypertension. J Am Coll Cardiol. 65:668–680. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pott C, Brixius K, Bundkirchen A, Bölck B,
Bloch W, Steinritz D, Mehlhorn U and Schwinger RH: The preferential
beta3-adrenoceptor agonist BRL 37344 increases force via
beta1-/beta2-adrenoceptors and induces endothelial nitric oxide
synthase via beta3-adrenoceptors in human atrial myocardium. Br J
Pharmacol. 138:521–529. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hicks A, McCafferty GP, Riedel E, Aiyar N,
Pullen M, Evans C, Luce TD, Coatney RW, Rivera GC, Westfall TD and
Hieble JP: GW427353 (solabegron), a novel, selective
beta3-adrenergic receptor agonist, evokes bladder relaxation and
increases micturition reflex threshold in the dog. J Pharmacol Exp
Ther. 323:202–209. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pankey EA, Edward JA, Swan KW, Bourgeois
CR, Bartow MJ, Yoo D, Peak TA, Song BM, Chan RA, Murthy SN, et al:
Nebivolol has a beneficial effect in monocrotaline-induced
pulmonary hypertension. Can J Physiol Pharmacol. 94:758–768. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gan RT, Li WM, Wang X, Wu S and Kong YH:
Effect of beta3-adrenoceptor antagonist on the cardiac function and
expression of endothelial nitric oxide synthase in a rat model of
heart failure. Zhongguo Wei Zhong Bing Ji Jiu Yi Xue. 19:675–678.
2007.(In Chinese). PubMed/NCBI
|
|
26
|
Wang Y, Li Z, Zhang Y, Yang W, Sun J, Shan
L and Li W: Targeting Pin1 protects mouse cardiomyocytes from
high-dose alcohol-induced apoptosis. Oxid Med Cell Longev.
2016:45289062016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guide for the Care and Use of Laboratory
Animals. th (ed). Washington (DC): 2011, PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schultze AE and Roth RA: Chronic pulmonary
hypertension-the monocrotaline model and involvement of the
hemostatic system. J Toxicol Environ Health B Crit Rev. 1:271–346.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rudski LG, Lai WW, Afilalo J, Hua L,
Handschumacher MD, Chandrasekaran K, Solomon SD, Louie EK and
Schiller NB: Guidelines for the echocardiographic assessment of the
right heart in adults: A report from the American Society of
Echocardiography endorsed by the European Association of
Echocardiography, a registered branch of the European Society of
Cardiology and the Canadian Society of Echocardiography. J Am Soc
Echocardiogr. 23:685-713–quiz 786–788. 2010. View Article : Google Scholar
|
|
31
|
Bogaard HJ, Natarajan R, Mizuno S, Abbate
A, Chang PJ, Chau VQ, Hoke NN, Kraskauskas D, Kasper M, Salloum FN
and Voelkel NF: Adrenergic receptor blockade reverses right heart
remodeling and dysfunction in pulmonary hypertensive rats. Am J
Respir Crit Care Med. 182:652–660. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Baker JG, Hill SJ and Summers RJ:
Evolution of β-blockers: From anti-anginal drugs to ligand-directed
signalling. Trends Pharmacol Sci. 32:227–234. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dessy C and Balligand JL: Beta3-adrenergic
receptors in cardiac and vascular tissues emerging concepts and
therapeutic perspectives. Adv Pharmacol. 59:135–163. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Strosberg AD: Structure and function of
the beta 3-adrenergic receptor. Annu Rev Pharmacol Toxicol.
37:421–450. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng HJ, Zhang ZS, Onishi K, Ukai T, Sane
DC and Cheng CP: Upregulation of functional beta(3)-adrenergic
receptor in the failing canine myocardium. Circ Res. 89:599–606.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Morimoto A, Hasegawa H, Cheng HJ, Little
WC and Cheng CP: Endogenous beta3-adrenoreceptor activation
contributes to left ventricular and cardiomyocyte dysfunction in
heart failure. Am J Physiol Heart Circ Physiol. 286:H2425–H2433.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gan RT, Li WM, Xiu CH, Shen JX, Wang X, Wu
S and Kong YH: Chronic blocking of beta 3-adrenoceptor ameliorates
cardiac function in rat model of heart failure. Chin Med J (Engl).
120:2250–2255. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ursino MG, Vasina V, Raschi E, Crema F and
De Ponti F: The beta3-adrenoceptor as a therapeutic target: Current
perspectives. Pharmacol Res. 59:221–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hutchinson DS, Chernogubova E, Sato M,
Summers RJ and Bengtsson T: Agonist effects of zinterol at the
mouse and human beta(3)-adrenoceptor. Naunyn Schmiedebergs Arch
Pharmacol. 373:158–168. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sato M, Horinouchi T, Hutchinson DS, Evans
BA and Summers RJ: Ligand-directed signaling at the
beta3-adrenoceptor produced by
3-(2-Ethylphenoxy)-1-[(1,S)-1,2,3,4-tetrahydronapt-1-ylamino]-2S-2-propanol
oxalate (SR59230A) relative to receptor agonists. Mol Pharmacol.
72:1359–1368. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vrydag W and Michel MC: Tools to study
beta3-adrenoceptors. Naunyn Schmiedebergs Arch Pharmacol.
374:385–398. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Price LC, Wort SJ, Perros F, Dorfmüller P,
Huertas A, Montani D, Cohen-Kaminsky S and Humbert M: Inflammation
in pulmonary arterial hypertension. Chest. 141:210–221. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Steiner MK, Syrkina OL, Kolliputi N, Mark
EJ, Hales CA and Waxman AB: Interleukin-6 overexpression induces
pulmonary hypertension. Circ Res. 104:236–244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mottillo EP, Shen XJ and Granneman JG:
Beta3-adrenergic receptor induction of adipocyte inflammation
requires lipolytic activation of stress kinases p38 and JNK.
Biochim Biophys Acta. 1801:1048–1055. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tchivileva IE, Tan KS, Gambarian M,
Nackley AG, Medvedev AV, Romanov S, Flood PM, Maixner W, Makarov SS
and Diatchenko L: Signaling pathways mediating beta3-adrenergic
receptor-induced production of interleukin-6 in adipocytes. Mol
Immunol. 46:2256–2266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mohamed-Ali V, Flower L, Sethi J,
Hotamisligil G, Gray R, Humphries SE, York DA and Pinkney J:
Beta-adrenergic regulation of IL-6 release from adipose tissue: In
vivo and in vitro studies. J Clin Endocrinol Metab. 86:5864–5869.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Haddad F, Doyle R, Murphy DJ and Hunt SA:
Right ventricular function in cardiovascular disease, part II:
Pathophysiology, clinical importance and management of right
ventricular failure. Circulation. 117:1717–1731. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sydykov A, Mamazhakypov A, Petrovic A,
Kosanovic D, Sarybaev AS, Weissmann N, Ghofrani HA and Schermuly
RT: Inflammatory mediators drive adverse right ventricular
remodeling and dysfunction and serve as potential biomarkers. Front
Physiol. 9:6092018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Klinger JR, Abman SH and Gladwin MT:
Nitric oxide deficiency and endothelial dysfunction in pulmonary
arterial hypertension. Am J Respir Crit Care Med. 188:639–646.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Roberts JD Jr, Roberts CT, Jones RC, Zapol
WM and Bloch KD: Continuous nitric oxide inhalation reduces
pulmonary arterial structural changes, right ventricular
hypertrophy and growth retardation in the hypoxic newborn rat. Circ
Res. 76:215–222. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Steudel W, Ichinose F, Huang PL, Hurford
WE, Jones RC, Bevan JA, Fishman MC and Zapol WM: Pulmonary
vasoconstriction and hypertension in mice with targeted disruption
of the endothelial nitric oxide synthase (NOS 3) gene. Circ Res.
81:34–41. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fagan KA, Tyler RC, Sato K, Fouty BW,
Morris KG Jr, Huang PL, McMurtry IF and Rodman DM: Relative
contributions of endothelial, inducible and neuronal NOS to tone in
the murine pulmonary circulation. Am J Physiol. 277:L472–L478.
1999.PubMed/NCBI
|
|
53
|
Amour J, Loyer X, Le Guen M, Mabrouk N,
David JS, Camors E, Carusio N, Vivien B, Andriantsitohaina R,
Heymes C and Riou B: Altered contractile response due to increased
beta3-adrenoceptor stimulation in diabetic cardiomyopathy: The role
of nitric oxide synthase 1-derived nitric oxide. Anesthesiology.
107:452–460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Maffei A, Di Pardo A, Carangi R, Carullo
P, Poulet R, Gentile MT, Vecchione C and Lembo G: Nebivolol induces
nitric oxide release in the heart through inducible nitric oxide
synthase activation. Hypertension. 50:652–656. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
d'Uscio LV: eNOS uncoupling in pulmonary
hypertension. Cardiovasc Res. 92:359–360. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Napp A, Brixius K, Pott C, Ziskoven C,
Boelck B, Mehlhorn U, Schwinger RH and Bloch W: Effects of the
beta3-adrenergic agonist BRL 37344 on endothelial nitric oxide
synthase phosphorylation and force of contraction in human failing
myocardium. J Card Fail. 15:57–67. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Nakazawa H, Hori M, Ozaki H and Karaki H:
Mechanisms underlying the impairment of endothelium-dependent
relaxation in the pulmonary artery of monocrotaline-induced
pulmonary hypertensive rats. Br J Pharmacol. 128:1098–1104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Seta F, Rahmani M, Turner PV and Funk CD:
Pulmonary oxidative stress is increased in cyclooxygenase-2
knockdown mice with mild pulmonary hypertension induced by
monocrotaline. PLoS One. 6:e234392011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Karbach S, Wenzel P, Waisman A, Munzel T
and Daiber A: eNOS uncoupling in cardiovascular diseases-the role
of oxidative stress and inflammation. Curr Pharm Des. 20:3579–3594.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Demarco VG, Whaley-Connell AT, Sowers JR,
Habibi J and Dellsperger KC: Contribution of oxidative stress to
pulmonary arterial hypertension. World J Cardiol. 2:316–324. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Francis BN, Salameh M, Khamisy-Farah R and
Farah R: Tetrahydrobiopterin (BH4): Targeting endothelial nitric
oxide synthase as a potential therapy for pulmonary hypertension.
Cardiovasc Ther. 36:2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Dorfmüller P, Chaumais MC, Giannakouli M,
Durand-Gasselin I, Raymond N, Fadel E, Mercier O, Charlotte F,
Montani D, Simonneau G, et al: Increased oxidative stress and
severe arterial remodeling induced by permanent high-flow challenge
in experimental pulmonary hypertension. Respir Res. 12:1192011.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hampl V, BíbovBbová J, BanasovBbová A,
Uhlík J, Miková D, Hnilicková O, Lachmanová V and Herget J:
Pulmonary vascular iNOS induction participates in the onset of
chronic hypoxic pulmonary hypertension. Am J Physiol Lung Cell Mol
Physiol. 290:L11–L20. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang Y, Tian W, Xiu C, Yan M, Wang S and
Mei Y: Urantide improves the structure and function of right
ventricle as determined by echocardiography in
monocrotaline-induced pulmonary hypertension rat model. Clin
Rheumatol. 38:29–35. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kimura K, Daimon M, Morita H, Kawata T,
Nakao T, Okano T, Lee SL, Takenaka K, Nagai R, Yatomi Y and Komuro
I: Evaluation of right ventricle by speckle tracking and
conventional echocardiography in rats with right ventricular heart
failure. Int Heart J. 56:349–353. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Brixius K, Bloch W, Pott C, Napp A,
Krahwinkel A, Ziskoven C, Koriller M, Mehlhorn U, Hescheler J,
Fleischmann B and Schwinger RH: Mechanisms of beta
3-adrenoceptor-induced eNOS activation in right atrial and left
ventricular human myocardium. Br J Pharmacol. 143:1014–1022. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Brixius K, Bloch W, Ziskoven C, Bölck B,
Napp A, Pott C, Steinritz D, Jiminez M, Addicks K, Giacobino JP and
Schwinger RH: Beta3-adrenergic eNOS stimulation in left ventricular
murine myocardium. Can J Physiol Pharmacol. 84:1051–1060. 2006.
View Article : Google Scholar : PubMed/NCBI
|