Introduction
Infection and inflammation of the urogenital tract
caused by microbiological factors accounts for >12% of cases of
male infertility, while sexually transmitted infections are the
leading cause of female infertility (1). For years, microbiological studies have
focused on how infection and inflammation of the urogenital tract
induce infertility, but little attention has been paid to the
possible impact of the normal reproductive microbiome on
infertility (2).
The vaginal microbiota of healthy females consists
of a wide variety of anaerobic and aerobic bacterial genera (two to
five genera at any one time) (3,4), and the
vaginal microbiomes have been classified as Lactobacillus (L.)
crispatus type, L. gasseri type, L. iners type
and L. jensenii type, based on the most dominant species of
Lactobacillus present (4,5).
Depletion of Lactobacillus has been associated with several
adverse conditions, including ectopic pregnancy, pelvic
inflammatory disease and infertility (3,4).
In previous studies, Anaerococcus,
Corynebacterium, Gardnerella, Lactobacillus, Prevotella,
Pseudomonas, Streptococcus and Veillonella have been
identified as the most abundant bacteria in semen (6–10), and
the predominance of Anaerococcus, Prevotella, Pseudomonas
and Lactobacillus are typically associated with semen health
and fertility (6–9). Levels of infertility are predicted to
increase in the future (7), and
infectious agents, including bacteria (Escherichia coli in
particular), yeasts and viruses may directly impair sperm motility
(12). However, the role of other
types of pathogenic bacteria or vaginal probiotic bacteria on sperm
motility has remained largely unexplored.
In the present study, vaginal secretions from 60
healthy human females were used to isolate vaginal bacteria. These
bacteria were used to evaluate bacterial capacity to adhere to
sperm, to study bacterial effect on human sperm function and to
study the potential role of Lactobacillus in fertility.
Materials and methods
Sample collection and treatment
A total of 100 reproductive-aged females were
recruited for the present study between June 2016 and November
2016. After health screening 60 of these females were selected for
collection of vaginal secretions. Subjects with pathogen or HIV
infection were excluded. The subjects were known to be free of
hysteromyoma, adenomyosis, endometriosis, salpingemphraxis vaginal
inflammation, severe pelvic adhesion, any acute inflammation,
cancer and endocrine as well as autoimmune disorders. None of the
subjects reported any recent use of hormones, antibiotics or
vaginal medications. The subjects had not received any cervical
treatment within one week and were instructed not to perform any
douching within 5 days or sexual activity within 48 h. None of the
subjects were pregnant, lactating or during menses at the time of
sampling.
Samples of total semen were collected from donors in
The Second Affiliated Hospital of Nanchang University, China (n=10;
age, 22–38 years; without leukocytospermia) between October 2016
and December 2016 after informed consent was obtained. Donors with
normal sperm parameters according to the 2010 World Health
Organization criteria, 5th edition (13) (seminal volume, sperm count,
progressive motile spermatozoa, motile spermatozoa, non-motile
spermatozoa and sperm morphology) were recruited. Subjects with
abnormal karyotype and those who had suffered injury to the
genitals were excluded. Prior to sampling, the donors washed the
glans of their penises using soap and water. Semen was obtained by
masturbation, ejaculated into a sterile collection tube and
incubated at 37°C for 25–45 min for liquefaction. Basic semen
parameters (including seminal volume and sperm count) and
leukocytospermia were measured immediately.
Microbial diversity in sperm, vaginal
fluid and co-cultures
Vaginal fluid from all 60 donors were pooled
together in a 50 ml tube. Bacteria were isolated from the mixture
of vaginal fluids using a plate separation method and identified
using 16S ribosomal (r)RNA gene sequencing, as previously described
(13).
The isolated bacteria were co-cultured with purified
and capacitated sperm at 37°C and 5% CO2 for 2 h and the
initial sperm: bacteria ratio was 1:10; the non-adhered bacteria
were removed by washing three times using PBS. Subsequently, the
mixed samples were centrifuged at 400 × g for 5 min at room
temperature, and the supernatant was removed. Extraction of
bacterial DNA from the sperm [sperm mixture (SM) group; repeated 3
times], vaginal fluid (vaginal fluid mixture (VM) group; repeated 3
times) and co-cultures [mixture of the bacteria adhered to sperm
(AM) group; repeated 3 times] was performed using a TIANamp Genomic
DNA kit (Tiangen Biotech Co., Ltd) combined with bead-beating
(using 0.5 mm diameter glass beads; Bio Spec Products, Inc.) by
vortex mixer (model, MXF; SCILOGEX, LLC) according to the
manufacturer's protocol. The concentration and quality of extracted
genomic DNA was tested prior to sequencing using a NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc.). Extracted
genomic DNA was used as the template to amplify the V4 region of
16S rRNA genes using the forward barcoded 515F/806R primer pair
(GenBank accession no. PRJNA517276) (13). PCR, pyrosequencing of the PCR
amplicons and quality control of raw data were performed as
described previously (13).
Paired-end reads from the original DNA fragments were merged using
Fast Length Adjustment of Short Reads when there was a certain
overlap with the read generated from the opposite end of the same
DNA fragment and paired-end reads were assigned to each sample
according to the unique barcodes (13).
Sequence analysis was performed with UPARSE software
version 7.0.100 (http://drive5.com/uparse/) using the
UPARSE-operational taxonomic units (OTU) and UPARSE-OTUref
algorithms (http://www.drive5.com/usearch/manual/uparseotu_algo.html).
Pre-existing Perl scripts were used to analyse alpha and beta
diversity (within and among sequences respectively). Sequences with
≥97% similarity were assigned to the same OTUs. A sequence was
picked as representative for each OTU and the Ribosomal Database
Project classifier (http://rdp.cme.msu.edu/; Michigan State University)
was used to annotate taxonomic information (14). Weighted UniFrac distance analysis was
performed using the Quantitative Insights Into Microbial Ecology
(QIIME) software package version 1.9.1 (http://qiime.org/; QIIME Development Team) and cluster
analysis was then performed. Differentially abundant species were
identified and subjected to Kyoto Encyclopedia of Genes and Genomes
(KEGG, https://www.genome.jp/kegg/; Kanehisa
Laboratories) pathway analysis and they were characterised for
their metabolic capacity by Phylogenetic Investigation of
Communities by Reconstruction of Unobserved States, version 1.0.0
(http://picrust.github.io/picrust/;
The PICRUSt project) (13,15). Unweighted Pair-Group Method with
Arithmetic Mean method (UPGMA, https://www.sequentix.de/gelquest/help/upgma_method.htm;
SequentiX) was used to build a phylogenetic tree and the LEfSe
(Linear discriminant analysis effect size) method was used to
analyse the bacteria with significant differences among AM, SM and
VM groups (16).
Real time-quantitative PCR
DNA extraction was as described above. The primers
for PCR were designed using Primer 5.0, (http://www.premierbiosoft.com/; Premier Biosoft
International) and real-time PCR amplification was performed using
an ABI 7900HT Fast Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.) (13). The
PCR mixture contained 10 µl SYBR® Primer Ex Taq II
(Takara Biotechnology, Co., Ltd), 0.4 µl ROX Reference Dye (50X;
Takara Biotechnology, Co., Ltd), 1.0 µl template DNA, 0.8 µl each
of the primers (final concentration, 0.4 µM) and 7 µl milli-Q
H2O. The amplification was programmed to start at 95°C
for 10 min, followed by 40 cycles of denaturation at 95°C for 30
sec, annealing at 60°C for 30 sec and extension at 72°C for 30 sec.
Relative levels (fold change) of the target bacteria were analysed
using the 2−∆∆Cq method using data from the comparative
quantification cycle (Cq) (16). The
primers are listed in Table I.
Enterococcus was used as a control species due to its
similar numbers across all groups.
 | Table I.Primers for analysis of microbiota
via PCR. |
Table I.
Primers for analysis of microbiota
via PCR.
| Target
bacteria | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| Bacteroides |
GGTGTCGGCTTAAGTGCCAT |
CGGA(C/T)GTAAGGGCCGTGC |
|
Bifidobacterium |
TCGCGTC(C/T)GGTGTGAAAG |
CCACATCCAGC(A/G)TCCAC |
| Enterococcus |
CCCTTATTGTTAGTTGCCATCAT |
ACTCGTTGTACTTCCCATTGT |
|
Enterobacteriaceae |
CATGACGTTACCCGCAGAAGAAG |
CTCTACGAGACTCAAGCTTGC |
| Clostridium
perfringens |
CGCATAACGTTGAAAGATGG |
CCTTGGTAGGCCGTTACCC |
| Lactobacillus |
CACCGCTACACATGGAG |
AGCAGTAGGGAATCTTCCA |
| Fusobacterium
spp. |
CCCTTCAGTGCCGCAGT |
GTCGCAGGATGTCAAGAC |
Adherence assay
Single bacterial strains or the bacterial mixture
isolated from vaginal fluid were co-cultured with sperm in HEPES
saline (135 mM NaCl, 5 mM KCl, 1 mM MgSO4, 2 mM
CaCl2, 20 mM HEPES, 5 mM glucose, 10 mM lactic acid and
1 mM Na-pyruvate at pH 7.4) at 37°C with 5% CO2. After 2
h of incubation, cultures were centrifuged at a speed of 400 × g
for 5 min at room temperature, to separate sperm from any
non-adhered bacteria. The remaining sperm were washed with sterile
PBS and centrifuged at 400 × g for 5 min at room temperature and
this procedure was repeated 4 times. The cells were then fixed with
methanol for 30 min, Gram-stained and examined microscopically
(Olympus BX63 optical microscope; Olympus Corp.). An adherence
index, defined as the number of adherent bacteria per 100 sperm,
was determined from 20 random microscopic fields. Each adherence
assay was performed in triplicate. Control bacterial strains
(purchased from BeNa Culture Collection; Beijing Beina Chuangian
Biotechnology Institute) of Salmonella typhimurium, Escherichia
coli O157:H7, Group β-H streptococcus, Candida albicans,
Salmonella enteritidis, Shigella flexneri, Staphylococcus aureus,
Listeria monocytogenes and Pediococcus acidilactici were
also used for adhesion assays.
Determination of sperm motility
Sperm motility was examined after incubation with
bacteria using a computer-assisted sperm analysis (CASA) system
(WLJY-9,000; WeiLi Co., Ltd.). Parameters associated with
progressive motility (PR), progressive straight-line motility
(PRA), progressive non-linear motility (PRB), total sperm motility
(TM) and average path velocity (VAP) were recorded. A minimum of
200 sperm cells were analysed for each assay.
Sperm penetration of a viscous
medium
The ability of human sperm to penetrate viscous
media is a comprehensive indicator for the evaluation of sperm
motility in the viscous environment of the female reproductive
tract. In the present study, methylcellulose solution was used to
mimic the viscous environment encountered in the female
reproductive tract (17).
Methylcellulose (1% w/v) was dissolved in human tubal fluid (HTF)
medium and introduced into 7.5-cm flattened capillary tubes with
1.0 mm inner diameter (Elite Medical Co., Ltd.). One end of the
tube was sealed with plasticine. Human sperm was incubated in HTF
medium for 1 h at 37°C and 5% CO2, as sperm motility was
commonly observed to increase to hyperactivity after incubation.
Next, the open ends of the capillary tubes were inserted into the
incubation mixture. After 1 h, the tubes were removed, wiped and
the penetration of the methylcellulose by the sperm was analysed
using the CASA system (WLJY-9,000; WeiLi Co., Ltd.). Three fields
(×10) were counted at 1 and 2 cm from the base of the tube and the
average number of cells per field were calculated. The cell numbers
were normalised to the values of untreated controls.
Statistical analysis
Data analysis was performed using Graphpad Prism
version 7.0 (GraphPad Software, Inc.). Values are expressed as the
mean ± standard deviation. Differences between the controls and the
various samples were assessed by one-way analysis of variance
followed by Dunnett's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Molecular identification of the
vaginal bacteria adhered to sperm
A total of 100 females donated their vaginal fluids
and 60 samples were identified as healthy samples using clinical
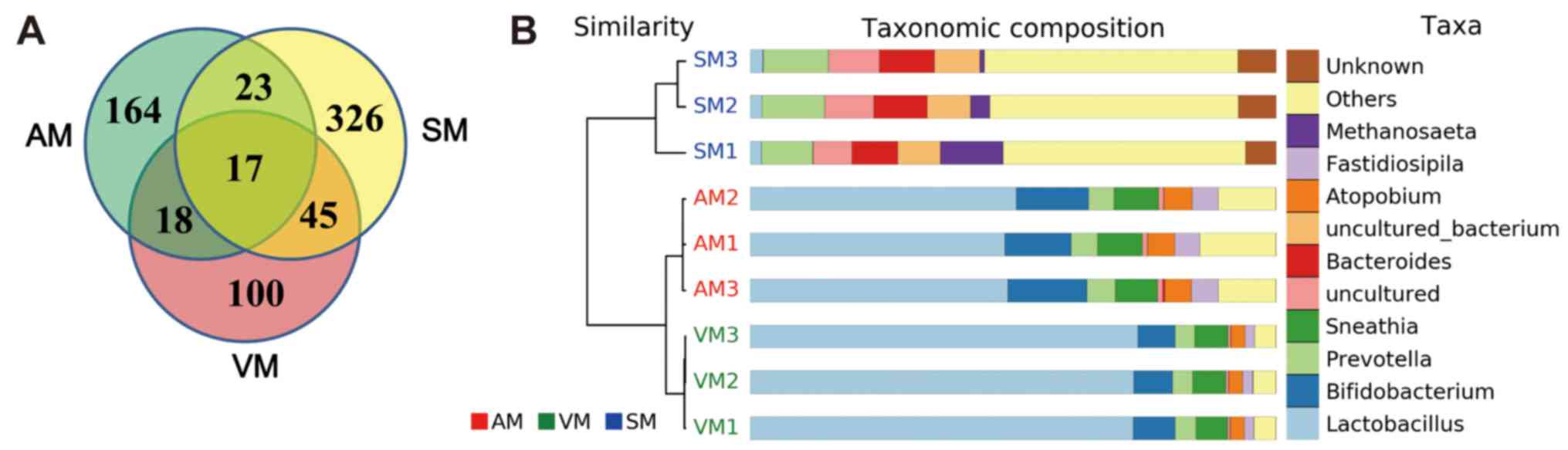
examinations. As presented in Fig.
1A, 222, 411 and 180 OTUs were identified from the groups AM,
SM and VM, respectively. Only 17 OTUs were determined to be common
OTUs among all groups, and Lactobacillus, Enterococcus and
Prevotella accounted for three, one and three of the common
OTUs, respectively (Table II).
Analysis of the top 10 genus populations using the UPGMA method
indicated that the microbial populations in groups AM and VM were
highly similar (Fig. 1B).
Lactobacillus accounted for 49 and 73% of the total OTUs in
the AM group and VM group, respectively, but only 2.3% of the total
OTUs in the SM group (Fig. 1B).
| Table II.Taxonomy of 17 common OTUs among the
three groups (vaginal bacteria, sperm bacteria and their
combination). |
Table II.
Taxonomy of 17 common OTUs among the
three groups (vaginal bacteria, sperm bacteria and their
combination).
| OTU ID | Taxonomy (at genus
level) |
|---|
| OTU11315 |
Lactobacillus |
| OTU14528 |
Gluconacetobacter |
| OTU15433 |
Lactobacillus |
| OTU37354 |
Ureaplasma |
| OTU46791 |
Leuconostoc |
| OTU46844 |
Enterococcus |
| OTU47147 |
Finegoldia |
| OTU51860 |
Porphyromonas |
| OTU56305 |
Bacillus |
| OTU71236 |
Prevotella |
| OTU74227 |
Streptococcus |
| OTU74838 |
Prevotella |
| OTU75162 |
Anaerococcus |
| OTU77493 |
Lactobacillus |
| OTU79751 |
Prevotella |
| OTU85858 |
Peptoniphilus |
| OTU96990 |
Mobiluncus |
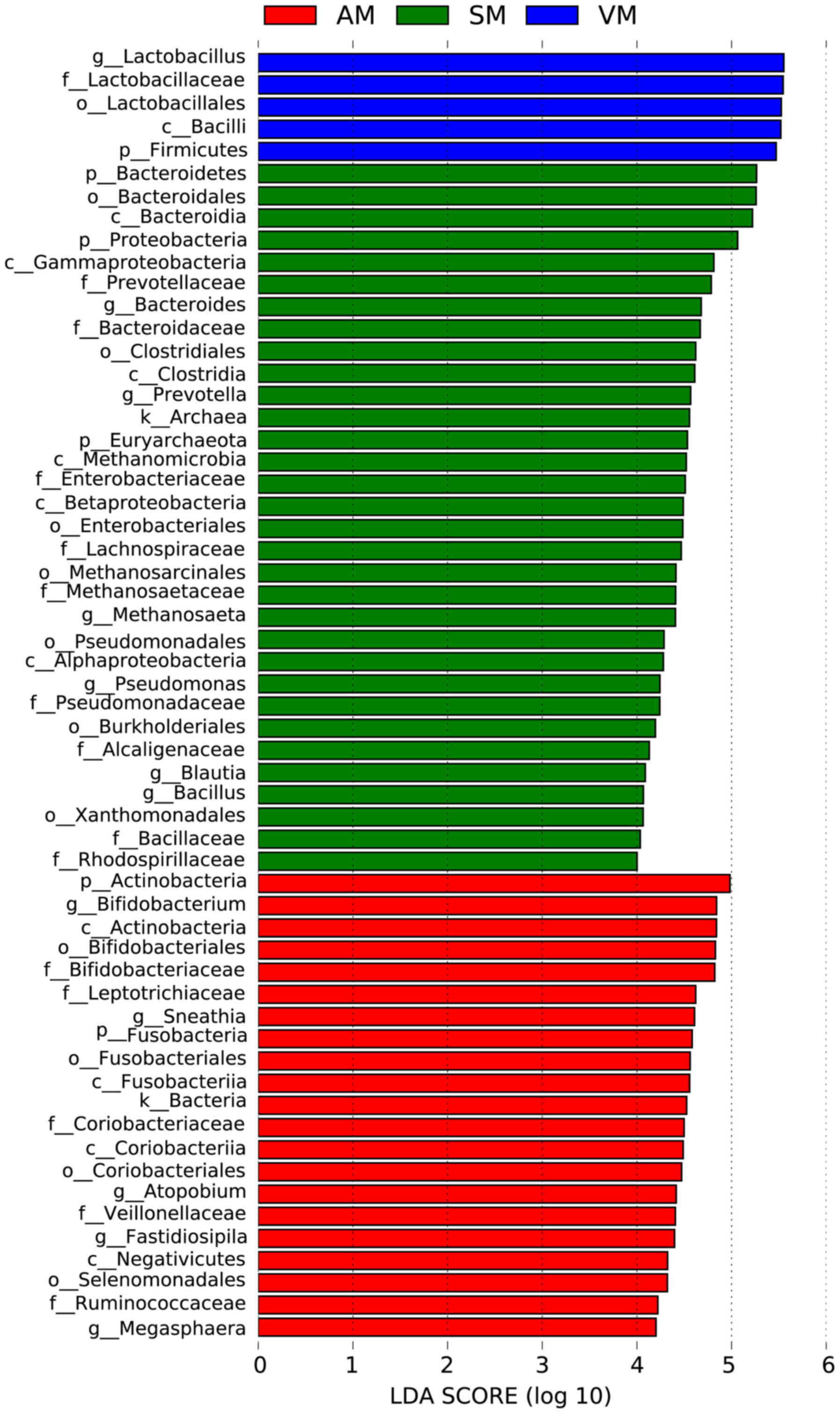
Real-time PCR analysis was performed to evaluate the
relative numbers of Enterococcus, Lactobacillus,
Fusobacterium spp., Bacteroides, Clostridium perfringens,
Bifidobacterium and Enterobacteriaceae in groups AM, SM
and VM, and the relatively stable genus of Enterococcus was
set as the control. As presented in Fig.
2, the numbers of Lactobacillus in the AM group and VM
group were 332- and 5917-fold higher than those of
Enterococcus, respectively, while the number of
Lactobacillus was 2.6-fold lower than that of
Enterococcus in the SM group. The numbers of
Enterobacteriaceae were 2.7-fold higher than those of
Enterococcus in the AM group, but 1.3- and 1.5-fold lower in
the SM group and the VM group, respectively. The prevalence of
Bacteroides, Prevotella, Methanosaeta, Pseudomonas, Blautia
and Bacillus strains was significantly higher in the SM
group than that in the AM group or the VM group (Fig. 3; P<0.05).
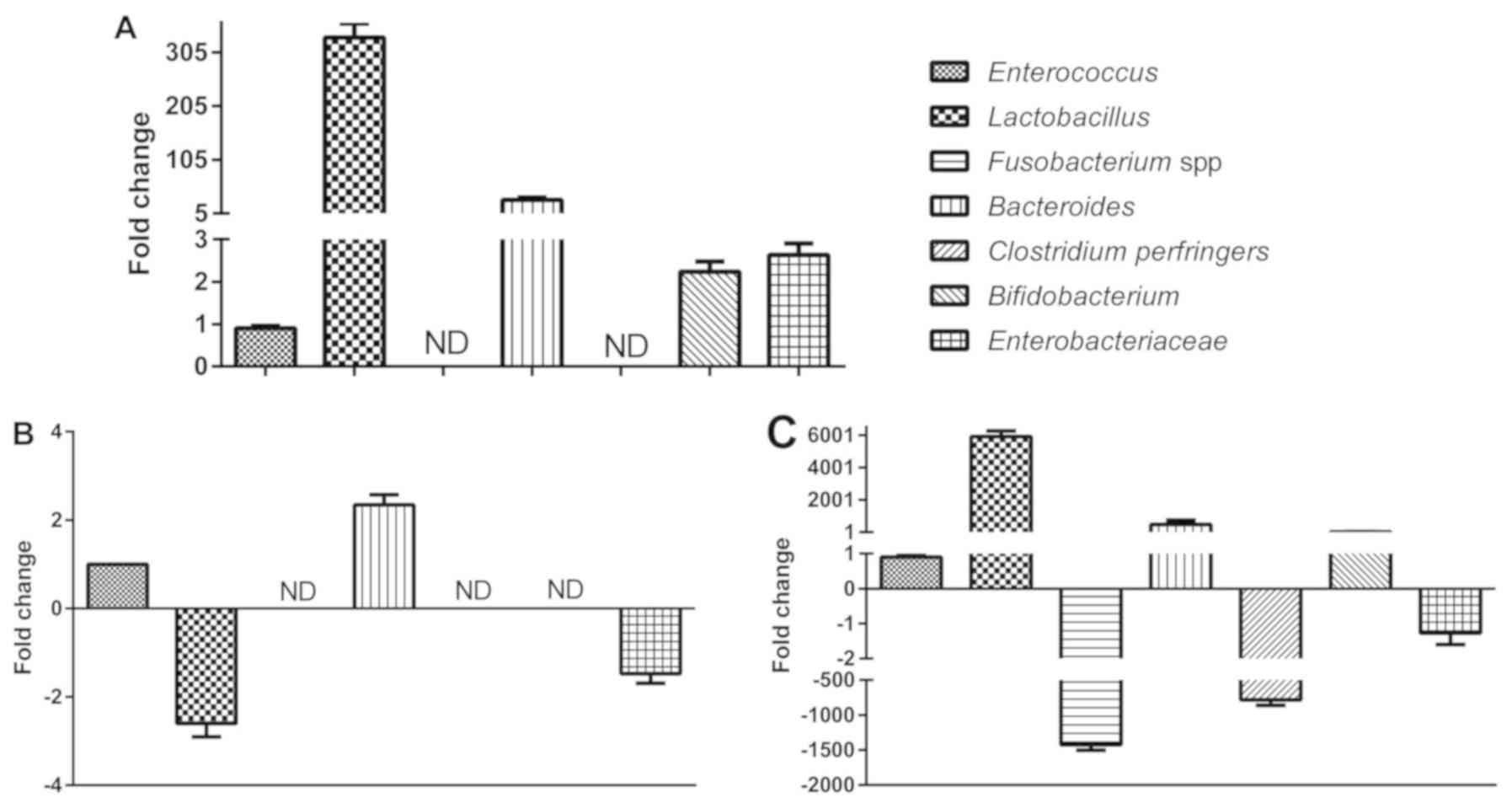 | Figure 2.PCR analysis of bacterial strains.
The relative amounts (fold change relative to Enterococcus)
of Lactobacillus, Fusobacterium spp., Bacteroides,
Clostridium perfringers, Bifidobacterium and
Enterobacteriaceae in the AM, SM and VM groups.
Enterococcus was selected as the reference strain due to its
conserved numbers between the groups (A) AM, (B) SM and (C) VM. The
relative numbers of Lactobacillus in the AM group and VM
group were higher than those in the SM group. VM, vaginal bacteria;
SM, sperm bacteria; AM, vaginal and sperm bacteria combined; ND,
not detectable. |
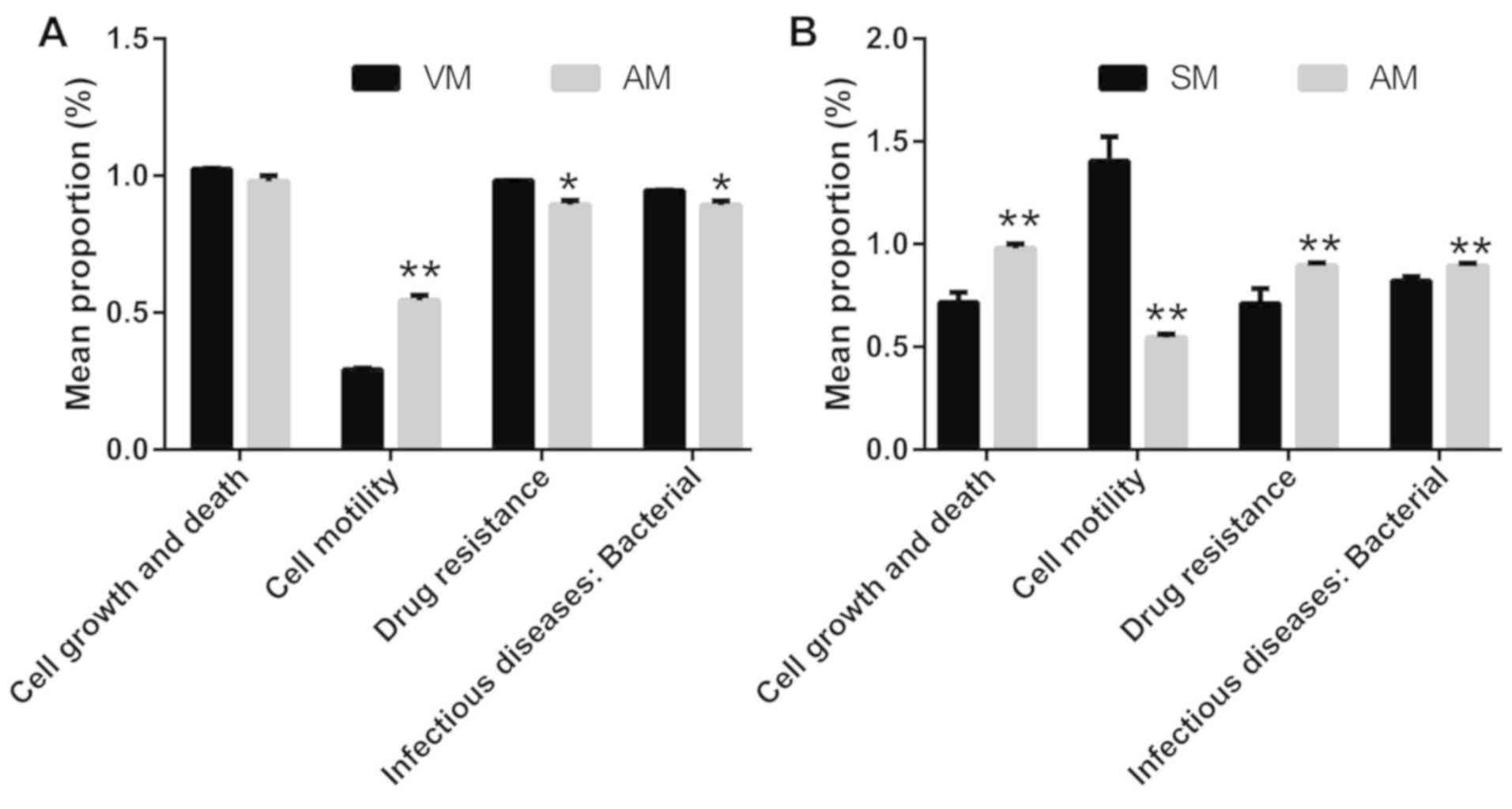
A bioinformatics analysis based on high-throughput
sequencing was then performed and the KEGG metabolic pathway
variance analysis revealed that adhered vaginal bacteria were
significantly associated with the terms promotion of sperm motility
and reduced drug resistance and rates of infection, as compared
with non-adherent bacteria (P<0.05; Fig. 4A). Conversely, the bacteria in the AM
group were associated significantly with the terms increased drug
resistance, infection, cell growth and death, as well as reduced
motility, as compared with those in the SM group (P<0.05;
Fig. 4B).
Adhesion of selected bacteria to
sperm
Isolation of bacteria from vaginal fluid yielded the
following species: Staphylococcus epidermidis, L. crispatus, L.
acidophilus, L. salivarius, L. helveticus, Pseudomonas stutzeri,
Staphylococcus capitis, L. gasseri and Enterococcus
faecalis. Of note, all of the tested bacteria, whether
probiotic or pathogenic, adhered to sperm in large numbers. The
number of adhered L. crispatus, L. gasseri and E. faecalis
cells was 565, 806 and 881 per 100 sperm, respectively (Fig. 5).
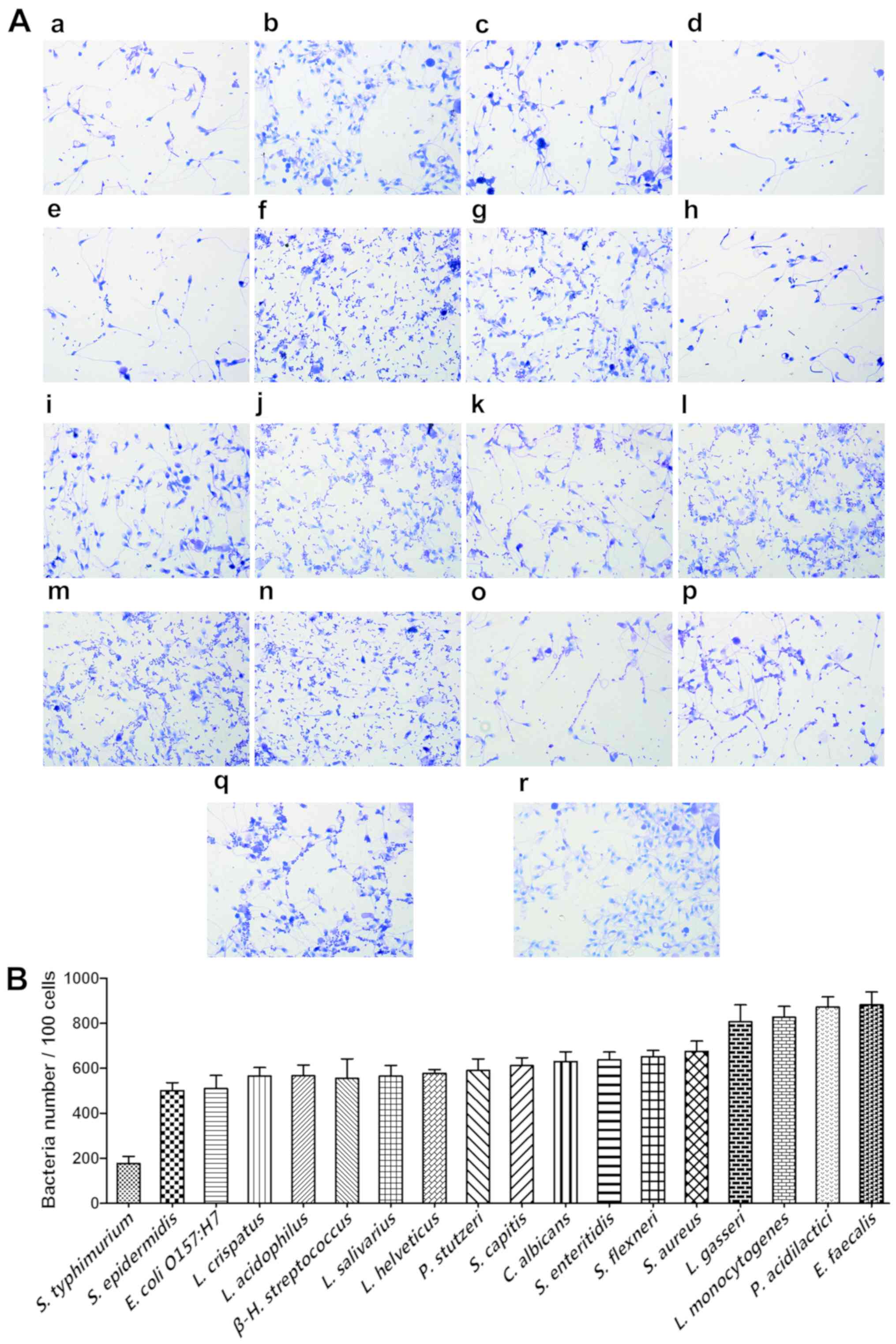 | Figure 5.Adhesion of the selected bacteria to
sperm cells. (A) The gram stain results (magnification, ×100) and
(B) adhesion numbers of S. typhimurium, S. epidermidis, E.
coli O157:H7, L. crispatus, L. acidophilus, β-H.
streptococcus, L. salivarius, L. helveticus, P. stutzeri, S.
capitis, C. Albicans, S. enteritidis, S. flexneri, S. aureus, L.
gasseri, L. monocytogenes, P. acidilactici and E.
faecalis. The initial sperm/bacteria ratio was 1:10 and S.
epidermidis, L. crispatus, L. acidophilus, L. salivarius, L.
helveticus, P. stutzeri, S. capitis, L. gasseri and E.
Faecalis were isolated from vaginal secretions of healthy
females, while the others were available in-house. Statistical
analysis was not performed due to the lack of an obvious control
strain. (B) Adherence index is defined as the number of adherent
bacteria per 100 sperm and was determined from 18 random
microscopic fields. Each adherence assay was performed in
triplicate. All of the tested bacteria, whether probiotic or
pathogenic, adhered to sperms in large numbers. |
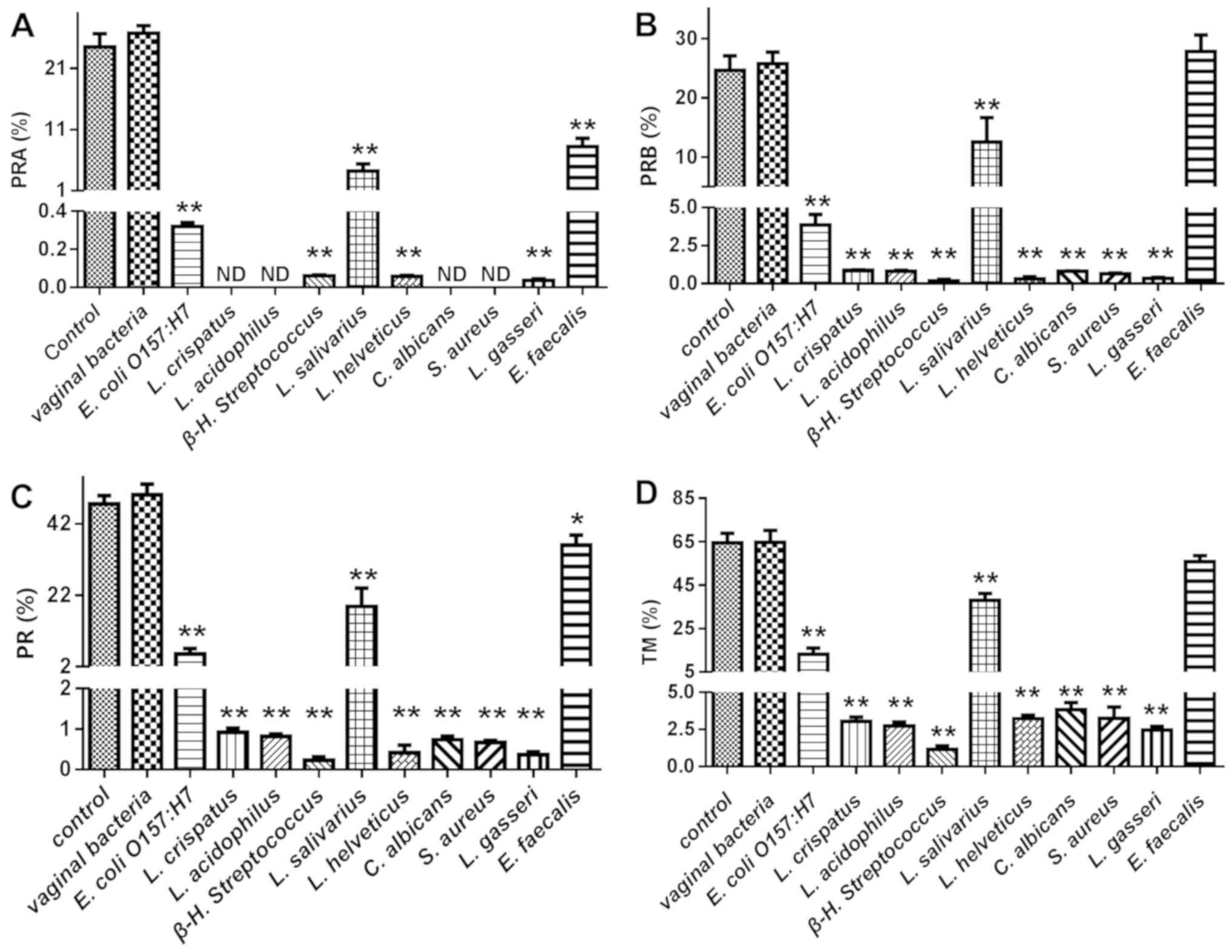
Effect of adhered bacteria on human
sperm motility
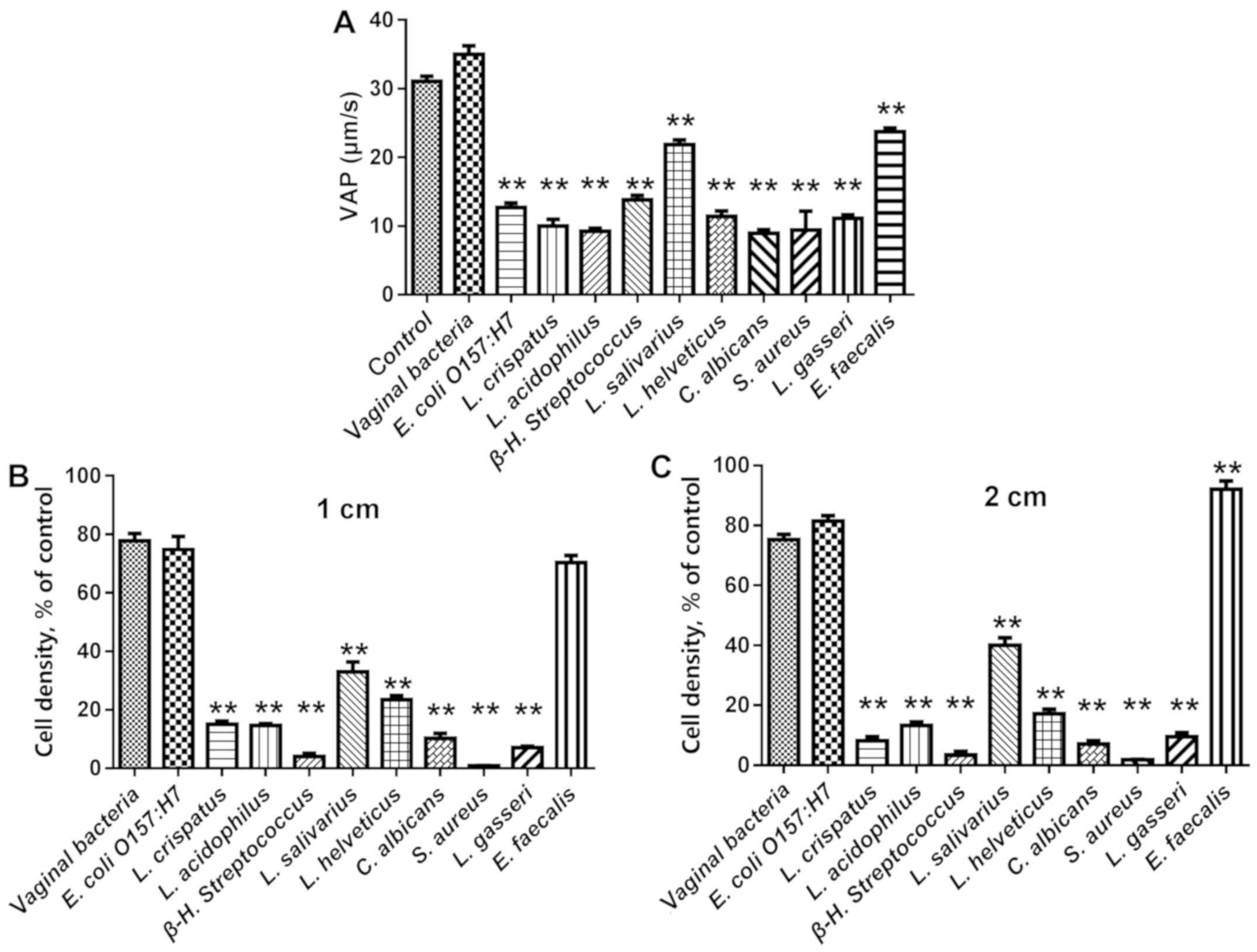
The vaginal bacteria E. coli O157:H7, L.
crispatus, L. acidophilus, β-H. streptococcus, L.
salivarius, L. helveticus, C. Albicans, S. aureus, L. gasseri
and E. faecalis were selected for evaluation of their
effects on human sperm motility. As presented in Figs. 6 and 7, nearly all tested bacteria caused a
statistically significant reduction in all sperm motility
parameters, including PRA, PRB, RP, TM and VAP, while E.
faecalis had a reduced affect on these parameters, though it
adhered to sperm in high numbers. The effects of the adhered
bacteria on sperm motility in a viscous medium were a better
indicator of sperm motility in the female reproductive system.
Comparison of the sperm numbers at 1 and 2 cm from the base of the
tube revealed that L. crispatus, L. acidophilus, β-H.
streptococcus, L. salivarius, L. helveticus, C. Albicans, S.
aureus and L. gasseri significantly reduced sperm
motility, while E. coli O157:H7 and E. faecalis did
not (Fig. 7B and C). Of note, the
probiotic bacteria L. crispatus, L. acidophilus, L.
helveticus and L. gasseri caused a marked reduction in
all of the motility parameters of human sperm, while the effects of
L. salivarius were less pronounced compared with those of
the other Lactobacillus.
Discussion
It has been indicated that changes in the vaginal
microbiome may affect the risk of gynaecological cancers (19). A healthy vaginal microbiome is
dominated by species of Lactobacillus that have a protective
effect and may have therapeutic potential (4,5). As with
the intestinal microbiome, disruption of the vaginal microbiota may
affect immunity or lead to increased growth of pathogens,
potentially resulting in the occurrence of diseases (20). It has been suggested that vaginal
microbiota with abundant Lactobacillus may also reduce the
risk of HIV transmission, particularly female-to-male transmission,
and reduce the rate of preterm birth (4,5). The
ability of the human vagina and its microflora to affect vaginal
discharge, menses, neonates and the overall health of individuals
is currently not sufficiently recognized (133).
Motility is the key factor of sperm function and is
predictive of its fertilisation potential in vitro. In
previous studies, the adhesion of E. coli and P.
aeruginosa to sperms has been extensively studied (4,5), while
the effects of Lactobacillus adhesion on sperm function have
remained to be evaluated, to the best of our knowledge. In
preliminary study by our group, vaginal bacteria were isolated and
it was indicated that most isolates were able to adhere to HeLa
cells (data not shown); therefore, the present study was performed
to assess whether these isolates are able to adhere to sperms
also.
Considering the challenges of assessing the effects
of microorganisms on sperm motility in vivo, an in
vitro model was used in the present study. First of all,
microbial diversity was compared between vaginal secretions and
semen, and a lower bacterial number was identified in the vagina,
possibly due to the harsh environment (low pH of 3.8–4.4) in the
vagina. In total, only 17 common OTUs were identified from the
groups AM, SM and VM, and Lactobacillus, Enterococcus and
Prevotella were identified from these OTUs.
The top 10 genus populations were then further
analysed using the UPGMA method, revealing that
Lactobacillus accounted for only 2.3% of the total OTUs in
the SM group, while it was as high as 49% in the AM group,
indicating Lactobacillus was able to effectively adhere to
sperms. The real-time PCR analysis further confirmed that the
number of Lactobacillus in the AM group was 332-fold higher
than that of Enterococcus (set as the control strain due to
its constant number in all groups), while the number of
Lactobacillus in the SM group was 2.6-fold lower than that
of Enterococcus.
Subsequently, KEGG metabolic pathway variance
analyses were performed and the results indicated that adhered
microbes markedly reduced sperm cell motility, and enhanced
bacterial cell growth and death, drug resistance and bacterial
infection compared to seminal bacteria (P<0.05). Therefore, the
effect of the vaginal isolates and certain in-house strains on
human sperm motility was evaluated. The results revealed that all
tested bacteria were able to adhere to sperms and nearly all of
them caused significant decreases in sperm motility parameters and
decreased sperm motility in a viscous medium, indicating their
potential negative effect on sperm function.
It is known that the predominance of vaginal
Lactobacillus has important health-promoting effects, which
may maintain the reproductive fitness of females through direct and
indirect anti-pathogenic mechanisms (21). Certain bacteria, fungi, viruses and
parasites are known to interfere with reproductive functions in
either sex and infections of the genitourinary tract account for
15% of male infertility cases (3,4). The
results of the present study revealed that all of the
Lactobacillus assessed (L. crispatus, L. acidophilus, L.
salivarius, L. helveticus and L. gasseri) were able to
effectively adhere to sperm and markedly reduce sperm motility. As
one of the most important Lactobacillus species, L.
crispatus appears to dominate the vaginal microbiota of most
healthy females (4). This species is
positively associated with anti-inflammatory cytokines [e.g.
interleukin (IL-10)], potentially decreasing the production of
pro-inflammatory cytokines, includings IL-6, IL-8 and tumour
necrosis factor-α (22). However,
the high adhesion of Lactobacillus species to sperms
significantly reduced the sperm functions, showcasing the dual
character of Lactobacillus in vaginal health and
reproduction.
Of note, the present results indicated that E.
faecalis had relatively minor negative effects on the PR, TM or
VAP of sperm or the ability of sperm to penetrate a viscous medium,
although this strain was able to adhere well to sperm cells. E.
faecalis are Gram-positive cocci that may survive harsh
conditions and certain strains are the cause of serious human and
animal infections (23–25). It is therefore worthwhile studying
the role of E. faecalis in the human vagina.
In conclusion, the present study evaluated the
effect of bacterial adhesion on sperm motility. All bacteria that
were tested adhered effectively to sperm and reduced their
motility, which was particularly obvious for the probiotic strains
of Lactobacillus. It may be postulated that the reduction in
motility caused by adhesion of Lactobacillus may be
beneficial for healthy couples attempting to conceive, as it may
reduce the chance of poor-quality sperm combining with eggs.
However, this effect may be detrimental for males with severe
asthenospermia, oligospermia or aspermia, as the bacteria may
eliminate the potential of the sperm combining with the egg.
Therefore, the high adhesion capability of vaginal bacteria should
be considered in the development of therapeutics, and the use of
probiotics in the treatment of gynaecological diseases should be
investigated. However, considering the small number of samples used
and the challenges of accurately controlling and quantifying
bacterial adherence in vitro, animal experiments, which may
provide a better model for the interaction of bacteria and sperms,
may be required in future work to obtain more robust
conclusions.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National
Natural Science Foundation of China (grant nos. 91639106, 81873659,
31560264 and 81503364), the Excellent Youth Foundation of Jiangxi
Scientific Committee (grant no. 20171BCB23028), the Jiangxi
Provincial Collaborative Innovation Center of Biopharmaceutics and
Biotechnology (grant no. 2015202004), the Key R&D Plan of
Jiangxi Science and Technology Agency (grant no. 20181BBG70028),
Jiangxi Provincial Science and Technology Innovation Team (grant
no. 20181BCB24003) and the Science and Technology Plan of Jianxi
Health Planning Committee (grant no. 20175526).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
TC and HX designed the experiment. HW, YC, TL, HC
and BT performed the experiments. TC analysed the data and wrote
the manuscript. All authors discussed the results and commented on
the manuscript.
Ethics approval and consent to
participate
This study was approved by the institutional review
boards of Jiangxi Maternal and Child Health Hospital (Nanchang,
China). Patients signed a written informed consent form for the use
of their samples for scientific research and for the publication of
their data. All participants in the study were adults.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dohle GR: Inflammatory-associated
obstructions of the male reproductive tract. Andrologia.
35:321–324. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jenmalm MC: The mother-offspring dyad:
Microbial transmission, immune interactions and allergy
development. J Intern Med. 282:484–495. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Petrova MI, Lievens E, Malik S, Imholz N
and Lebeer S: Lactobacillus species as biomarkers and agents that
can promote various aspects of vaginal health. Front Physiol.
6:812015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ravel J, Gajer P, Abdo Z, Schneider GM,
Koenig SS, McCulle SL, Karlebach S, Gorle R, Russell J, Tacket CO,
et al: Vaginal microbiome of reproductive-age women. Proc Natl Acad
Sci USA. 108 (Suppl 1):S4680–S4687. 2011. View Article : Google Scholar
|
|
5
|
van de Wijgert JH, Borgdorff H, Verhelst
R, Crucitti T, Francis S, Verstraelen H and Jespers V: The vaginal
microbiota: What have we learned after a decade of molecular
characterization? PLoS One. 9:e1059982014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hou D, Zhou X, Zhong X, Settles ML,
Herring J, Wang L, Abdo Z, Forney LJ and Xu C: Microbiota of the
seminal fluid from healthy and infertile men. Fertil Steril.
100:1261–1269. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jones C: Intersex, infertility and the
future: Early diagnoses and the imagined life course. Sociol Health
Illn. 2019. View Article : Google Scholar
|
|
8
|
Jarvi K, Lacroix JM, Jain A, Dumitru I,
Heritz D and Mittelman MW: Polymerase chain reaction-based
detection of bacteria in semen. Fertil Steril. 66:463–467. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mändar R, Punab M, Borovkova N, Lapp E,
Kiiker R, Korrovits P, Metspalu A, Krjutškov K, Nõlvak H, Preem JK,
et al: Complementary seminovaginal microbiome in couples. Res
Microbiol. 166:440–447. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mändar R, Punab M, Korrovits P, Türk S,
Ausmees K, Lapp E, Preem JK, Oopkaup K, Salumets A and Truu J:
Seminal microbiome in men with and without prostatitis. Int J Urol.
24:211–216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen H, Luo T, Chen T and Wang G: Seminal
bacterial composition in patients with obstructive and
non-obstructive azoospermia. Exp Ther Med. 15:2884–2890.
2018.PubMed/NCBI
|
|
12
|
Armanini D, Giorgino FL and Fiore C:
Editorial comment to receptor dependent immobilization of
spermatozoa by sperm immobilization factor isolated from
Escherichia coli: Proof of evidence. Int J Urol. 18:603–604. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Champer M, Wong AM, Champer J, Brito IL,
Messer PW, Hou JY and Wright JD: The role of the vaginal microbiome
in gynaecological cancer: A review. BJOG-Int J Obstet Gy.
125:309–315. 2018. View Article : Google Scholar
|
|
14
|
Meng F, Chen T, Wang X, Wang X, Wei H,
Tian P, Wang H, Zhao X, Shen L and Xin H: Evaluation of the
accuracy and sensitivity of high-throughput sequencing technology
using known microbiota. Mol Med Rep. 17:408–413. 2018.PubMed/NCBI
|
|
15
|
Zhang F, Zhang M, Wang Y, Li C and Chen T:
Comparison of the common bacteria in human and mouse tumours using
high-throughput sequencing. Mol Med Rep. 17:6717–6722.
2018.PubMed/NCBI
|
|
16
|
Drummond A and Rodrigo AG: Reconstructing
genealogies of serial samples under the assumption of a molecular
clock using serial-sample UPGMA. Mol Biol Evol. 17:1807–1815. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rao X, Huang X, Zhou Z and Lin X: An
improvement of the 2ˆ(-delta delta CT) method for quantitative
real-time polymerase chain reaction data analysis. Biostat
Bioinforma Biomath. 3:71–85. 2013.PubMed/NCBI
|
|
18
|
Luo T, Li N, He YQ, Weng SQ, Wang T, Zou
QX and Zeng XH: Emodin inhibits human sperm functions by reducing
sperm [Ca(2+)]i and tyrosine phosphorylation. Reprod Toxicol.
51:14–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ramchander NC and Crosbie EJ: The vaginal
microbiome and gynaecological cancer: Exercise caution when
considering causation. BJOG. 125:2017.PubMed/NCBI
|
|
20
|
Macklaim JM, Cohen CR, Donders G, Gloor
GB, Hill JE, Parham GP, Ravel J, Spear G, van de Wijgert J and Reid
G: Exploring a road map to counter misconceptions about the
cervicovaginal microbiome and disease. Reprod Sci. 19:1154–1162.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Verdenelli MC, Coman MM, Cecchini C, Silvi
S, Orpianesi C and Cresci A: Evaluation of antipathogenic activity
and adherence properties of human Lactobacillus strains for vaginal
formulations. J Appl Microbiol. 116:1297–1307. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kyongo JK, Jespers V, Goovaerts O,
Michiels J, Menten J, Fichorova RN, Crucitti T, Vanham G and Ariën
KK: Searching for lower female genital tract soluble and cellular
biomarkers: Defining levels and predictors in a cohort of healthy
Caucasian women. PLoS One. 7:e439512012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harwood VJ, Delahoya NC, Ulrich RM, Kramer
MF, Whitlock JE, Garey JR and Lim DV: Molecular confirmation of
Enterococcus faecalis and E. faecium from clinical, faecal and
environmental sources. Lett Appl Microbiol. 38:476–482. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Eirich J, Orth R and Sieber SA: Unraveling
the protein targets of vancomycin in living S. aureus and E.
faecalis cells. J Am Chem Soc. 133:12144–12153. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Creti R, Imperi M, Bertuccini L, Fabretti
F, Orefici G, Di Rosa R and Baldassarri L: A survey for virulence
determinants among E. faecalis isolated from different sources. J
Med Microbiol. 53:13–20. 2004. View Article : Google Scholar : PubMed/NCBI
|