Introduction
Acute myocardial infarction (AMI) is an ischemic
heart disease, which is characterized by myocardial necrosis due to
persistent and severe myocardial ischemia (1). AMI is the cause of more than a third of
deaths in developed nations annually, and is responsible for a
large number of annual deaths in the USA and more than 4 million
annual deaths in Europe and Northern Asia (2,3). In
recent decades the incidence of AMI has begun to decline due to the
development of new therapies and changing lifestyles. However,
approximately 7 million individuals still suffer from AMI worldwide
each year (4).
If reperfusion is not performed shortly after
ischemic injury following AMI, it results in irreversible necrosis
of cardiomyocytes, which leads to myocardial fibrosis and scar
tissue formation. Progression of this fibrosis affects contraction
of the ventricle, leading to ventricular remodeling, and ultimately
contributes to the occurrence of heart failure (5).
Metastasis associated lung adenocarcinoma transcript
1 (MALAT1) is a long non-coding RNA (lncRNA) also known as
nuclear-enriched abundant transcript 2, and was originally
identified as a prognostic marker for lung cancer metastasis
(6). Two modes of action have been
proposed for MALAT1: i) Alternative splicing of a subset of
pre-micro RNAs is regulated by MALAT1's recruitment of
phosphorylated SR splicing factors (7); and ii) MALAT1 is associated with
transcriptional control of the gene expression of cell cycle genes
(8). Though the regulatory
mechanisms of MALAT1 action in other conditions remains to be
clarified, it has been reported that MALAT1 is expressed at high
levels in tumor tissue and its expression has been linked to tumor
cell proliferation, apoptosis, migration, invasion and metastatic
spread (9). A study showed that the
development of bladder cancer is controlled by MALAT1 through
regulation of NAD-dependent protease sirtuin-7 and microRNA
(miR)-125b (10). Additional
research indicated that MALAT1 also promotes the proliferation,
migration and invasion of esophageal squamous cell carcinoma
(11). Studies have also shown that
MALAT1 contributes to endothelial cell function, vessel growth and
inflammation (12,13). In 2017, Zhao et al (14) found that MALAT1 functions as a
mediator of the cardioprotective effect of fentanyl during
myocardial ischemia-reperfusion injury. However, the detailed
molecular mechanisms involved in MALAT1-mediated modulation of AMI
need further elucidation.
Nucleotide binding and oligomerization domain-like
receptors (NLRs) are a group of pattern-recognition receptors,
which have various functions in innate immunity (15). Nucleotide binding and oligomerization
domain-like receptor C5 (NLRC5), the largest member of the NLR
protein family, has been well characterized as a regulator of
antigen presentation via modulation of major histocompatibility
complex class I gene expression (16). NLRC5 has also been linked to
suppression of type I interferon production through the IκB kinase
complex or retinoic-acid inducible protein
I/melanoma-differentiation-associated gene 5 (17). It has been confirmed that NLRC5
participates in the transformation and invasion of malignant cancer
cells (18,19) and an increasing number of studies
have found that NLRC5 is associated with non-immunological diseases
(20). MiR-125b-5p is a known
regulator of apoptosis in several types of cells (21–23).
Recently, NLRC5 was reported to have a protective effect against
heart disease (24). This led to the
hypothesis that the effect of MALAT1 on AMI-induced heart damage
may be associated with NLRC5 and miR-125b-5p.
In the present study, downregulation of MALAT1 was
found to prevent AMI-induced tissue damage in rats. In
vitro, expression of NLRC5 and miR-125b-5p were both influenced
by MALAT1. TargetScan and dual-luciferase reporter assays provided
further confirmation that NLRC5 is the direct target gene of
miR-125b-5p. Furthermore, NLRC5 overexpression inhibited
miR-125b-5p-associated cytoprotection in
ischemia/reperfusion-treated HL-1 cells. These results suggest that
MALAT1 abrogates the protective effect of miR-125b-5p on
cardiomyocytes through upregulation of NLCR5 expression. These
findings suggest an emerging role for MALAT1 in the pathogenesis of
AMI and will guide future development of genetic therapeutic
strategies to treat AMI.
Materials and methods
Animals
A total of 32 adult male Sprague-Dawley (SD) rats
(14 weeks) were purchased from the experimental animal center of
the Dezhou People's Hospital. All procedures were approved by the
Dezhou People's Hospital Ethics Review Committee for Animal
Experimentation (experiment number 2017-TX-007). Rats were divided
into four groups (sham NC-siRNA, sham MALAT1-siRNA, AMI NC-siRNA
and AMI MALAT-siRNA, 8 rats per each group) and housed at room
temperature (22–26°C), humidity 50–70% on a 12-h light/dark cycle.
Food and water were provided ad libitum. The body weight of
each rat was recorded weekly. The SD rats (220–250 g) were
anesthetized using 10% chloral hydrate (350 mg/kg; Sinopharm
Chemical Reagent Co., Ltd.) intraperitoneally and were mechanically
ventilated by a TOPO Dual Mode Ventilator (Kent Scientific
Corporation). Peritonitis was not observed following the
administration of 10% chloral hydrate.
A rat model of AMI was developed by ligation of the
left anterior descending coronary artery. In brief, the
anesthetized rats were placed in a supine position and the trachea
carefully exposed. An endotracheal tube was introduced into the
trachea (polyethylene size 90). Mechanical ventilation was achieved
by connecting the endotracheal tube to a TOPO Dual Mode Ventilator
(Kent Scientific Corporation) cycling at 80 breaths per minute with
a tidal volume of 1.2 ml per 100 g body weight. Once steady
breathing was established, the left ventricle was exposed by
opening the chest at the 4th left intercostal space. Ischemia was
induced by extruding the heart and quickly and accurately ligating
the left anterior descending coronary artery with a 6-0 suture.
Reperfusion was established by cutting the knot in the ligature, 30
min after ligation. Before closing the chest, rats were monitored
for 5 min to ensure there was no bleeding due to the surgery. The
chest was closed using 6-0 prolene sutures with a continuous suture
pattern. Post-surgery, the vital signs of rats were measured until
sample collection. In sham control rats, the procedure was
identical, but there was no transient artery ligation. For the
MALAT1 knockdown experiment, the rats were intravenously injected
through the tail vein with small interfering (si)RNA against MALAT1
and siRNA-non-coding (NC) at the final concentration of 10 mg/kg
for 24 h before surgery. When pulsatile blood was observed in the
needle hub, the siRNA was injected with a continuous and slow
movement. The plasmids of siRNA against MALAT1 and NC were
generated using the pcDNA3.1 vector, and purchased from Shanghai
GenePharma Co., Ltd. siMALAT1, 5′-AAGAAAAAUAAAAGCUUUCCU-3′ and
siNC, 5′-ACGUCACACGUUCGGAGAATT-3′.
Determination of myocardial
infarction
The left ventricle was cut into 2-mm-thick sections
perpendicular to the long axis of the heart. A section from each
heart was wrapped with gauze, followed by incubation of the heart
in 1% 2,3,5-triphenyl-2H-tetrazolium chloride (TTC; Beijing
Solarbio Science & Technology Co., Ltd.) for 60 min at 37°C. An
additional section was incubated with phosphate-buffered saline
(PBS) as a negative control for the same length of time. To avoid
sample oxidation, each section was sealed with the PBS in a closed
freezer bag and gently shaken in a hot bath to obtain uniform
staining throughout the incubation at 37°C for 2 h. The sizes of
the TTC-stained area (red staining, ischemic but viable tissue) and
unstained area [infarct myocardium (INF)] were evaluated using
Image Pro Plus 6.0 (Media Cybernetics Inc.). The myocardial INF
size was expressed as a percentage of INF over total area at
risk.
Analysis of cardiac function
Cardiac function was evaluated by motion-mode
echocardiography using the VEVO 770 high-resolution in vivo
imaging system (FUJIFILM VisualSonics Inc.). Left ventricular
fractional shortening (LVFS) was calculated by computerized
algorithms. In brief, the left ventricular end-systolic diameter
(LVESD) and the left ventricular end-diastolic diameter (LVEDD)
were measured by M-mode of echocardiography. These parameters refer
to the size of the ventricle at the end of systole and diastole.
Based on the formula: (LVEDD-LVESD/LVEDD) ×100, the percentage of
size differences of the left ventricle as a parameter of how well
the left ventricle is contracting itself (25). All measurements represented the mean
of 5 consecutive cardiac cycles. Left ventricular internal
dimension (LVIDs) were measured at end-systole (s) through
four-chamber view. By using this, the endocardial margin of the
lateral wall and the septum were analyzed through Vevo 770 (V2.2.0;
VisualSonics, Toronto, Canada).
Cell culture
The mouse cardiomyocyte cell line HL-1 (cat. no.
SCC065) was purchased from Sigma-Aldrich (Merck KGaA). HL-1 cells
were cultured in Claycomb medium (Sigma-Aldrich; Merck KGaA)
supplemented with 10% heat-inactivated fetal bovine serum (MP
Biomedicals LLC.), 0.1 mM norepinephrine and 1%
penicillin-streptomycin (Nacalai Tesque Inc.) in a 5%
CO2 humidified incubator at 37°C. To establish a
hypoxia/reoxygenation (H/R) model the cells were cultured in a
Napco 8000WJ hypoxia (1% O2−5% CO2−94% N2)
incubator (Thermo Fisher Scientific, Inc.). After 10 h of hypoxia,
the HL-1 cells were plated at 5×105 cells/ml and
incubated under normoxic conditions in a CO2 incubator
for an additional 4 h. The HL-1 cells passage time was less than
20.
Plasmid construction
The MALAT1 siRNA were designed and obtained from
Invitrogen (4455877; Thermo Fisher Scientific, Inc.). The sequence
of MALAT1 siRNA was 5′-GCAGAGGCAUUUCAUCCUU-3′. The sequence of the
negative control siRNA was 5′-ACGUCACACGUUCGGAGAATT-3′ and it was
provided by Thermo Fisher Scientific, Inc. The human NLRC5 (NCBI
reference sequence, NP_115582) was obtained by nested PCR from a
human cDNA library (Marathon-ready cDNA; Clontech Laboratories,
Inc.) using the following primers: Forward,
5′-CGTGGGGACCCTAGAGCACCTATCA-3′ and reverse,
5′-GCATCACTTGGCTGGATTCCAAAGG-3′. The PCR products were cloned using
a Takara one step PCR kit (Takara Biotechnology Co., Ltd.) and
initial denaturation at 98°C for 30 sec, 30 cycles of denaturation
at 98°C for 5 sec, annealing at 60°C for 10 sec, and extension at
72°C for 10 sec, finally, extension at 72°C for 2 min. For MALAT1
overexpression, the human MALAT1-expressing vector (ORF023250,
NR_002819.3) was obtained from Applied Biological Materials Inc
(Richmond, BC, Canada).
Transfection
The function of MALTA1 was inhibited using siRNA.
HL-1 cells were plated on 24-well plates at a density of
1×105 cell per well and cultured overnight. The
following day, the cells were transfected with MALAT1 siRNA (500
ng/well) using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) and Opti-MEM (Gibco; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
Overexpression of NLRC5 or empty vector in HL-1 cells was also
carried out using the Lipofectamine 2000 system (500 ng/well of
plasmid). For the overexpression of miR-125b-5p, cells were
transfected with miR-125b-5p mimic or negative control mimic (NC
mimic) using a NanoFectin transfection reagent (Shanghai Excell
Biology, Inc.) according to the manufacturer's protocol. The
miR-125b-5p mimic (5′-AUUUGCGGCCGCUCAGCUGAGAUGUUCUGAGGU-3′) or
NC-mimic (5′-UUCUCCGAACGUGUCACGUTT-3′) was used at a concentration
of 100 nM (Shanghai GenePharma Co., Ltd.).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells or tissue using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. RNase-free DNase (Takara
Biotechnology Co., Ltd.) was then added to the RNA following a
manufacturer's protocol. A total of 2 µl of RNA was
reverse-transcribed to cDNA using the SuperScript IV First-Strand
Synthesis System (Invitrogen; Thermo Fisher Scientific, Inc.)
following the instructions provided by the manufacturer. The qPCR
amplification conditions consisted of pre-denaturation for 3 min at
94°C, followed by a total of 30 cycles of denaturation for 30 sec
at 94°C, annealing at 58°C for 30 sec and extension for 60 sec at
72°C. The expression of genes was measured by SYBR-green master mix
(Applied Biosystems; Thermo Fisher Scientific, Inc.) with the
primer sets described below using an ABIViiA7 Real-Time PCR System.
U6 was used as an internal control for miR-125b-5p. Expression of
β-actin was used as internal control for the analysis of other
genes while the 2−ΔΔCq method was used for data analysis
(26). The primer sequences were
(5′→3′) as follows: Malat1 forward, TCTTAGAGGGTGGGCTTTTGTT
and reverse, CTGCATCTAGGCCATCATACTG; miR-125b-5p forward,
TGCGCTCCCTGAGACCCTAAC and reverse, CCAGTGCAGGGTCCGAGGTATT;
nlrc5 forward, CTCAGCCAGGAGCACGTAG and reverse,
CCAGCAGGTTTGCTGAGAGA; β-actin forward, CCTCTATGCCAACACAGTGC
and reverse, CATCGTACTCCTGCTTGCTG; and U6 forward,
CTCGCTTCGGCAGCACA and reverse, AACGCTTCACGAATTTGCG.
Western blotting
The samples were lysed and prepared using Laemmli
sample buffer (cat. no. 161-0747, Bio-Rad Laboratories, Inc.). The
concentration of protein in each sample was determined using a BCA
Protein Assay kit (Thermo Fisher Scientific, Inc.). A total of 15
µg of protein were subjected to 10% SDS-PAGE and separated proteins
were transferred to polyvinylidene difluoride membrane
(Immobilon-P; EMD Millipore). The membrane was blocked with 5%
skimmed milk in TBS-T for 1 h at room temperature and then
incubated with the primary antibody against NLRC5 (1:1,000, cat.
no. sc-515668; Santa Cruz Biotechnology, Inc.) overnight at 4°C.
After washing three times in TBS-T, the membrane was incubated with
secondary antibody (1:5,000, anti-mouse; AP31535BT-N, OriGene
Technologies Inc.) for 1 h at room temperature. The expression of
β-actin was measured by using an anti-β-actin antibody for 1 h at
room temperature (1:1,000; cat. no. 3700, CST Biological Reagents
Co., Ltd.), before incubation with the same secondary antibody
following the aforementioned conditions. Protein expression was
detected using Luminata Forte Western HRP Substrate (EMD Millipore)
with a Bio-Rad ChemiDoc XRS + imaging system (Bio-Rad Laboratories,
Inc.).
Luciferase activity
The TargetScan 7.2 (targetscan.org) was used for prediction of the direct
target gene of miR-125b-5p, and the confirmation was further
performed by dual-luciferase reporter assay. To construct a
luciferase reporter containing the wild type NLRC5 3′untranslated
region (3′UTR) with a predicted miR-125b-5p binding site and mutant
NLRC5 3′UTR, the region of NLRC5 and its mutant were amplified
using genomic DNA as a template. PCR products were inserted into
the pMIR-report plasmid (Ambion; Thermo Fisher Scientific, Inc.).
In brief, HL-1 cells were cultured in 24-well plates at the density
of 2.5×104 cells/well and transfected with 0.3 µg of
firefly luciferase reporter plasmid (Ambion, Thermo Fisher
Scientific, Inc.) using Lipofectamine® 2000. An equal
amount of β-galactosidase expression vector (Ambion; Thermo Fisher
Scientific, Inc.) was transfected as a control. After a 48-h
transfection, the relative activity of luciferase was determined
using the Dual-Luciferase Reporter Assay kit (Promega Corporation)
according to the manufacturer's instructions. The miR-125b-5p mimic
or negative control mimic (Guangzhou RiboBio Co., Ltd.) was
co-transfected with the wild-type or mutant reporter vectors. The
luciferase activities were then determined using Beckman Coulter
DTX880 Multimode Detector (Beckman Coulter, Inc.) at 560 nm and the
data were normalized against Renilla luciferase
activity.
Measurement of L-lactate dehydrogenase
(LDH) and malondialdehyde (MDA)
The LDH level in the supernatant was quantified
using an LDH assay kit (ab102526, Abcam) according to the
manufacturer's instructions. In brief, the HL-1 cell culture
supernatants were incubated for 1 h at room temperature in 96-well
plates coated with an anti-LDH antibody. After washing, a
biotin-conjugated capture antibody was added and incubated for 1 h
at room temperature, followed by addition of an avidin-horseradish
peroxidase conjugate to bind to any captured antigen. Absorbance
was measured at 450 nm. The values were expressed as the percentage
of LDH released relative to the value obtained following
permeabilization of the cells with digitonin. The MDA level was
analyzed using a Cellular Malondialdehyde Test kit (A003-2, Nanjing
Jiancheng Bioengineering Institute). The standards, of known
concentrations, were incubated with N-methyl-2-phenylindole for 1 h
at 45°C The resulting chromophore was analyzed with a
spectrophotometer at 586 nm. The MDA concentration in the samples
was interpolated from the standard curve.
Fluorescence activated cell sorting
(FACs)
After 48 h of transfection, the HL-1 cells were
recovered, washed with PBS and resuspended. The culture
supernatants were discarded by centrifugation at 100 × g at 4°C for
10 min. Cells were then resuspended in 150 µl of 10 mM HEPES (pH
7.4), 140 mM NaCl and 2.5 mM CaCl2, and placed on ice
for 5 min. A further 50 µl of buffer containing 5 µl propidium
iodide and 10 µl Annexin V-FITC (FITC Annexin V Apoptosis Detection
Kit I; cat. no. 556547; BD Biosciences) was added. After mixing,
the cells were incubated at room temperature in the dark for 15
min. A FACS Calibur cell analyzer (BD Biosciences) and FlowJo
10.6.1 (FlowJo LLC) were used to analyze cell apoptosis and
calculate the apoptosis rate. The apoptotic cells including
apoptotic cells in early stage (Annexin
V+/PI− cells) and late stage (Annexin
V+/PI+ cells), which were used to calculation
of apoptotic cells/viable cells.
Statistical analysis
All data are presented as the mean ± standard
deviation. To determine statistical significance, GraphPad Prism 7
(GraphPad Software, Inc.) was used. One-way ANOVA with Tukey's post
hoc test was used for multiple comparisons. Student's t-test was
used when comparing between 2 conditions. P<0.05 indicated
statistical significance.
Results
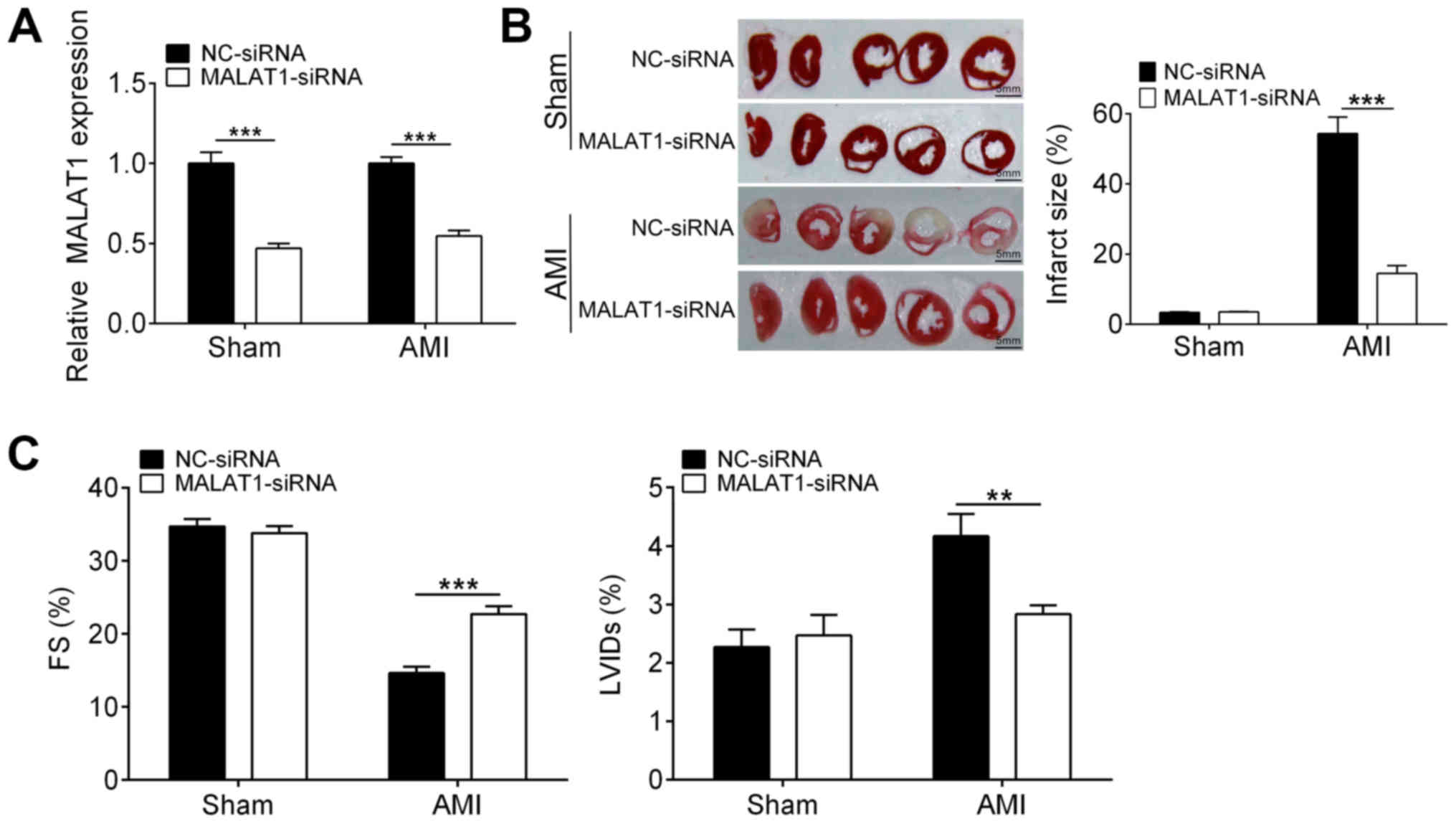
Downregulation of MALAT1 attenuates
heart damage in a rat model of AMI
To investigate the exact role of MALAT1 in
myocardial infarction, a rat model of AMI was established and
MALAT1 was knocked down by siRNA transfection. As shown in Fig. 1A, the mRNA levels of MALAT1 in both
sham and AMI rats were significantly downregulated through siRNA.
Infarction size of heart from sham rats were not affected by
downregulation of MALAT1. Interestingly, knockdown of MALAT1
strongly decreased the myocardial infarction area in AMI rats as
compared with the NC-siRNA group (Fig.
1B). Although AMI caused a reduction in the LVFS and increased
the LVIDs (Fig. 1C), downregulation
of MALAT1 increased the LVFS and reduced LVIDs in AMI rats when
compared with the control (Fig. 1C).
These findings suggested that MALAT1 knockdown ameliorated
AMI-induced heart damage.
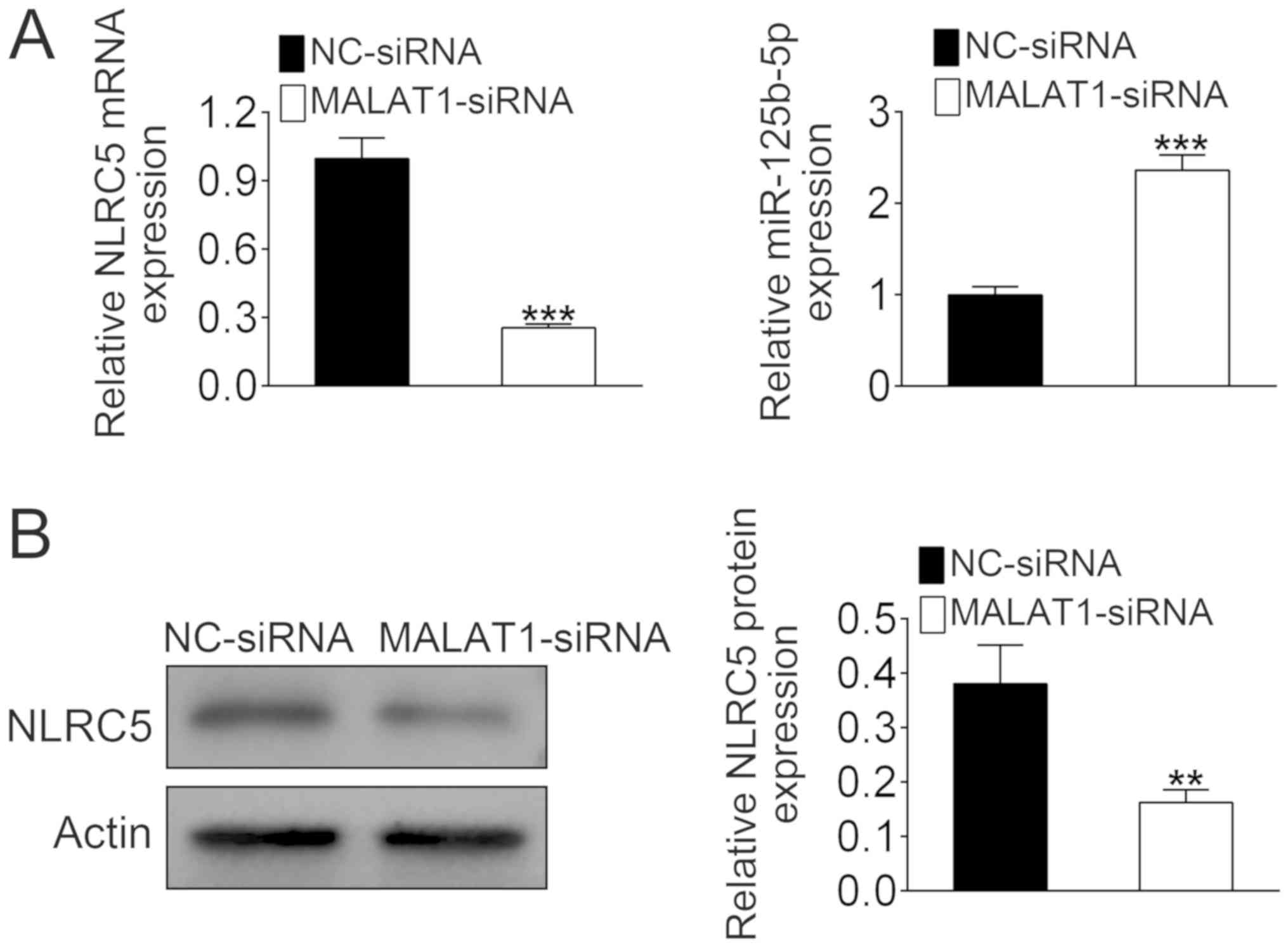
MALAT1 negatively regulates
miR-125b-5p expression and positively regulates the expression of
NLRC5
MALAT1 siRNA was transfected into the mouse
cardiomyocyte cell line HL-1. The mRNA level of NLRC5 was reduced
by more than 60% with MALAT1 knockdown compared with the control
group (Fig. 2A). By contrast, MALAT1
siRNA treatment caused a significant increase in the expression of
miR-125b-5p (Fig. 2A). Western
blotting confirmed that downregulation of MALAT1 significantly
reduced the expression of NLRC5 (Fig.
2B). These data indicated that MALAT1 could regulate the
expression levels of NLRC5 and miR-125b-5p.
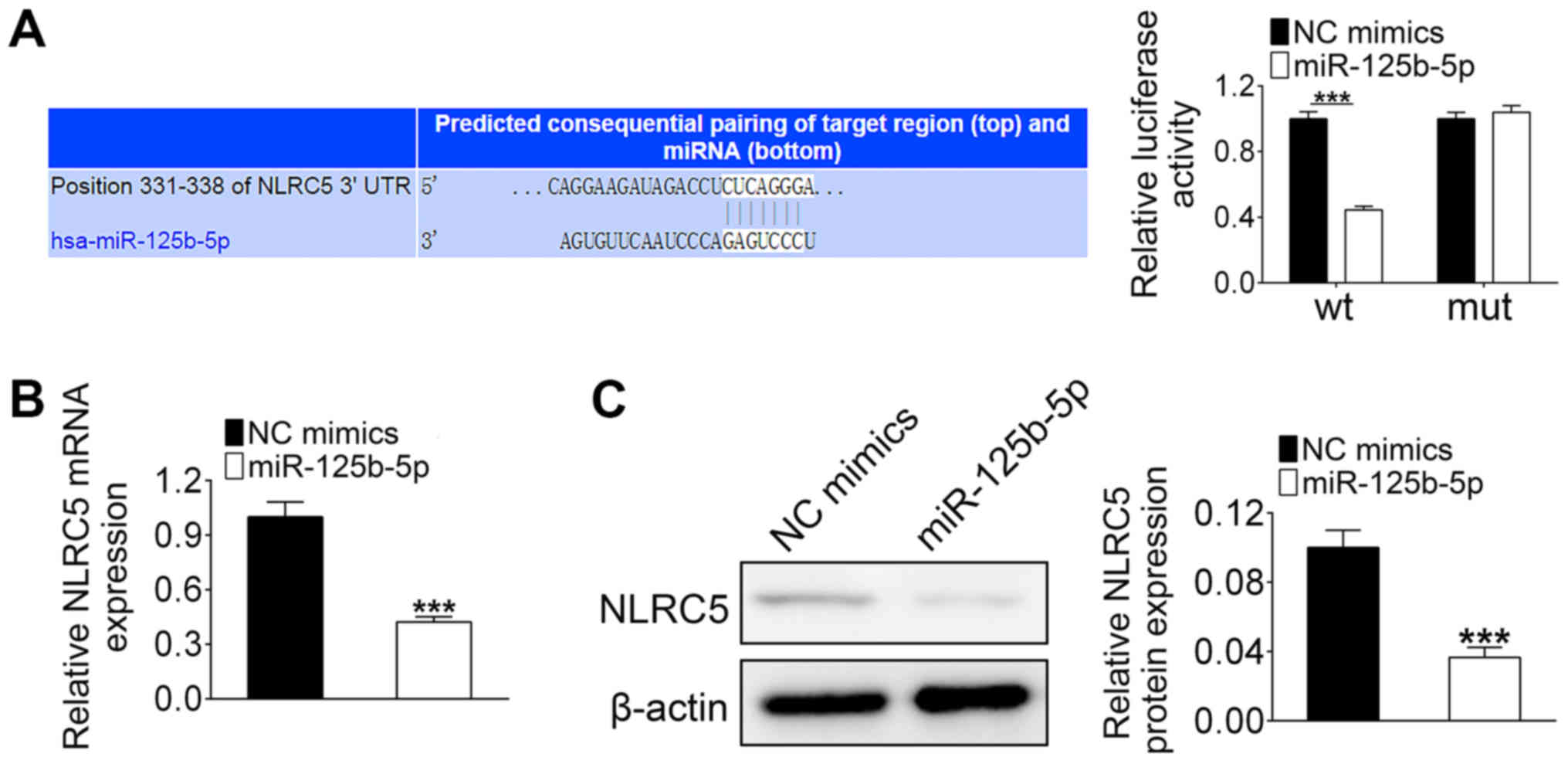
NLRC5 is a target of miR-125b-5p
Since MALAT1 directly regulates the expression of
miR-125b-5p, it was hypothesized that MALAT1 could regulate NLRC5
expression via miR-125b-5p. The binding sites between miR-125b-5p
and NLRC5 are shown in Fig. 3A.
Luciferase activity was significantly decreased in HL-1 cells that
were co-transfected with miR-125b-5p mimics and wild-type 3′UTR of
NLRC5 compared with that co-transfected with the miR-125b-5p mimics
and the mutant 3′UTR of NLRC5 (Fig.
3A). Additionally, both mRNA and protein levels of NLRC5 in
HL-1 cells were significantly decreased by miR-125b-5p
overexpression (Fig. 3B and C).
These results indicated that NLRC5 is a target of miR-125b-5p.
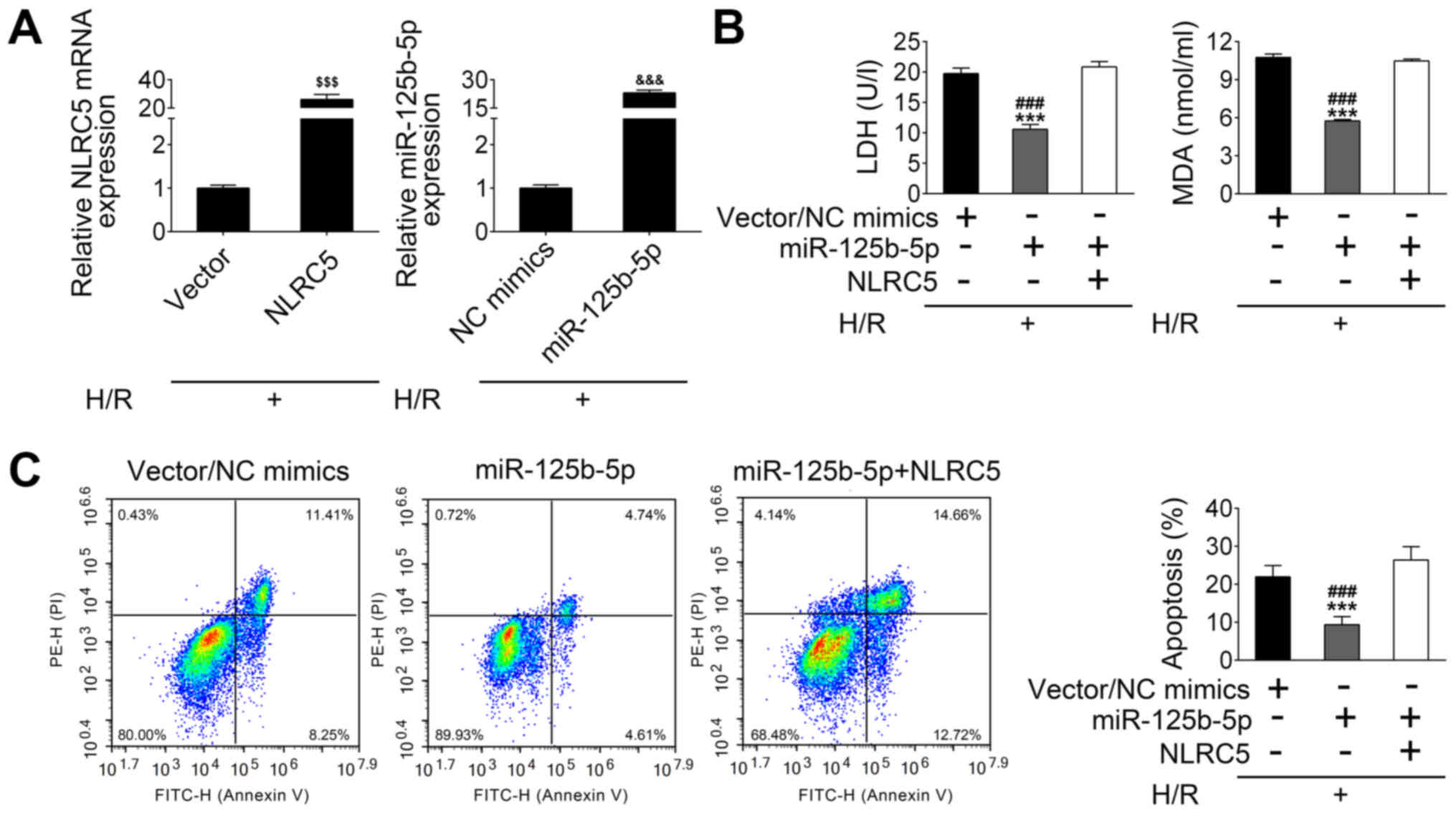
MiR-125b-5p exerts an inhibitory
effect on H/R-induced apoptosis of a cardiomyocyte cell line by
suppressing NLRC5 expression
HL-1 cells were treated with H/R to establish an
in vitro ischemia/reperfusion I/R model. By using these
cells, NLRC5 and miR-125b-5p mRNA expression was increased
following the transfection of H/R-treated HL-1 cells with the NLRC5
vector or the miR-125b-5p mimics, respectively (Fig. 4A). As shown in Fig. 4B, the concentration of lactate
dehydrogenase (LDH), a marker of cell pyroptosis, was decreased by
a miR-125b-5p mimic in H/R-treated HL-1 cells compared with control
vector/mimic transfected cells. However, NLRC5 overexpression
significantly reversed these results (Fig. 4B). The level of MDA, an index of
reactive nitrogen species formation, was also decreased in
miR-125b-5p-overexpressing HL-1 cells under H/R conditions.
Similarly, the decreased MDA level in miR-125b-5p-overexpressing
H/R-treated HL-1 cells was blocked by co-transfection with
miR-125b-5p mimics and NLRC5 overexpression plasmid (Fig. 4B). FACs analysis showed that H/R
treatment causes HL-1 cells apoptosis including 8.25% at early
stage and 11.41% at late stage, and reduction of cell death (~10%
reduction), including 4.61% at early stage and 4.74% at late stage,
was observed when the miR-125b-5p expression was raised in HL-1
cells, as compared with control. Meanwhile, overexpression of NLRC5
in raised miR-125b-5p expressing HL-1 cells suppressed the
cytoprotective effect of miR-125b-5p, as observed by the increased
apoptotic cell number (Fig. 4C).
These data suggest that miR-125b-5p overexpression may exert an
inhibitory effect on H/R-induced apoptosis through negatively
regulating NLRC5.
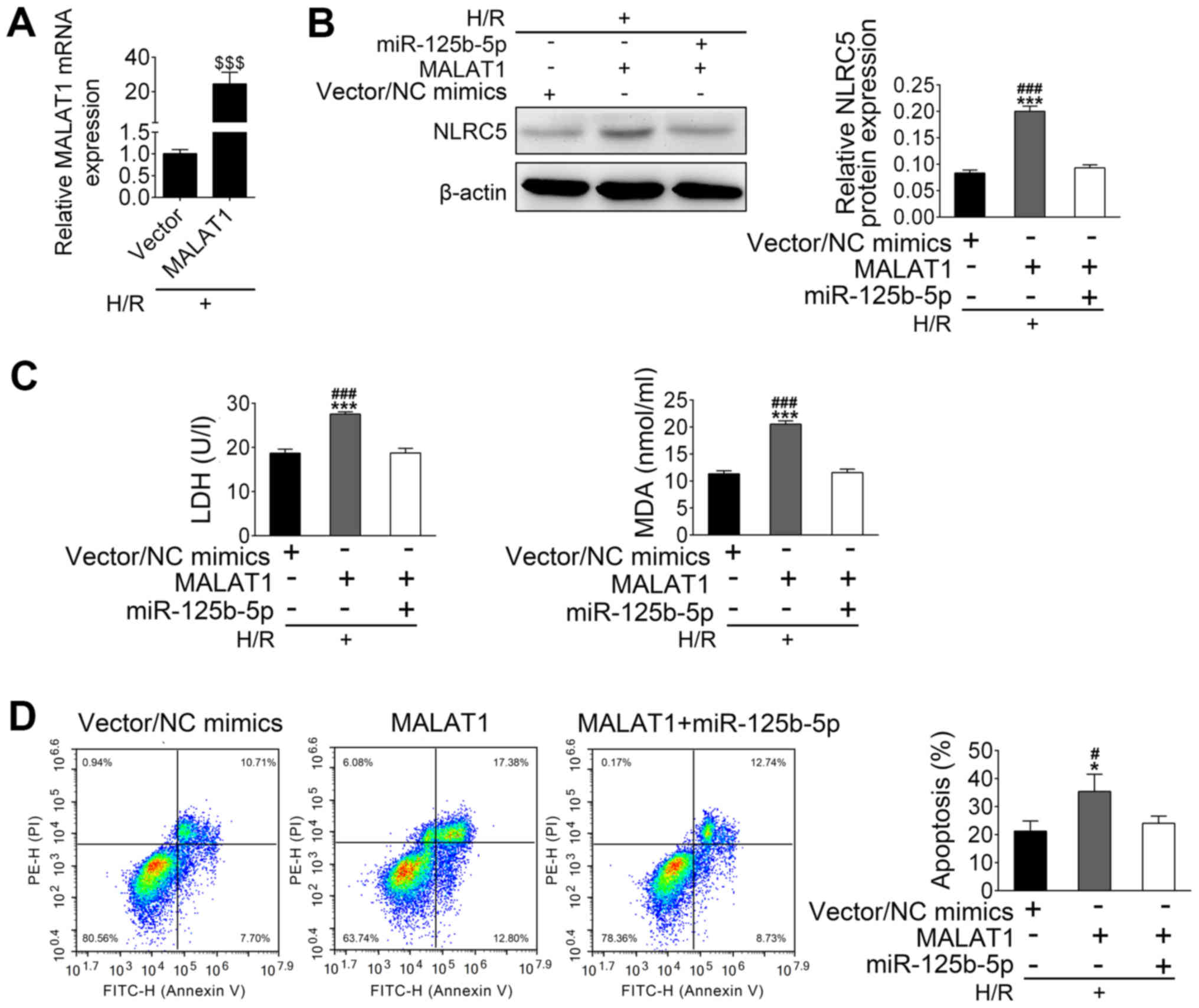
MALAT1 alleviates the protective
effect of miR-125b-5p on H/R-induced apoptosis by upregulation of
NLRC5
After establishing that transfected HL-1 cells
overexpressed MALAT-1 (Fig. 5A),
further experiments aimed to investigate whether MALAT1 could
inhibit the protective effect of miR-125b-5p on H/R-induced
apoptosis. H/R treated HL-1 cells were co-transfected with MALAT1
vector and miR-125b-5p. In H/R-treated HL-1 cells, the expression
of NLRC5 was increased by MALAT1 overexpression and suppressed by
transfection of miR-125b-5p and MALAT1 (Fig. 5B). Higher levels of LDH and MDA were
seen in the cultured supernatants of MALAT1-overexpressing
H/R-treated HL-1 cells compared with the control group. This
increased LDH release and MDA production induced by MALAT1
overexpression was reduced after overexpression of miR-125b-5p in
H/R-treated HL-1 cells (Fig. 5C).
MALAT1 overexpression also induced apoptosis in H/R-treated HL-1
cells, which was inhibited by upregulation of miR-125b-5p (Fig. 5D). Taken together, these data suggest
that MALAT1 may induce HL-1 apoptosis through downregulation of
miR-125b-5p, which leads to upregulation of NLRC5.
Discussion
Although lncRNA MALAT1 has been investigated in
multiple human cancers (6,26), little is known about its involvement
and regu3lation in the pathogenesis of heart disease. A recent
study revealed that MALAT1 acts as an inducer of myocardial injury
through negatively regulating miR-204 expression to increase
cardiomyocyte autophagy and I/R-induced myocardial injury (27). Huang et al (28) showed that MALAT1 mediates cardiac
fibrosis and reduces cardiac function in a MI mouse model.
Furthermore, MALAT1 has been linked with inflammation in myocardial
I/R injury (29). It has also been
shown to block the cardioprotective effects of fentanyl by
negatively regulating the miR-145/Bcl2 interacting protein 3
pathway (14). By contrast, Zhao
et al (30) reported that
MAPK1 upregulated the expression of MALAT1 to promote the
proliferation of cardiomyocytes. MALAT1 has also been found to
promote cardiac stem cell proliferation and migration (29). In the present study, downregulation
of MALAT1 significantly reduced infarction area and LVIDs, and
enhanced LVFS. However, the reason for the differential expression
of MALAT1 in the myocardium is unclear and requires further
investigation in the future.
The regulatory mechanisms underlying MALAT1 activity
in cardiovascular biology and disease are complex (31). It has been reported that lncRNA can
modify mRNA expression by acting as an miRNA sponge and blunting
the regulatory effects of relevant miRNAs (32). Oral squamous cell carcinoma
development is known to be promoted by MALAT1 via the
miR-125b/STAT3 axis (33). Han et
al (10) reported that miR-125b
suppressed bladder cancer development via inhibition of MALAT1.
Both of these publications suggest that miR-125b is a direct target
of MALAT1. In the present study the expression of miR-125b-5p was
negatively regulated by MALAT1 in cardiomyocyte cell line HL-1. In
H/R-treated HL-1 cells, cell apoptosis was significantly reduced by
miR-125b-5p overexpression, as were LDH release and MDA production
compared with controls. However, these protective effects of
miR-125b-5p on H/R-treated HL-1 cells were reversed by MALAT1
overexpression. These data suggested that miR-125b-5p is a
potential apoptosis regulator, which was downregulated by
MALAT1.
NLR proteins act as pattern-recognition receptors
and are critical components of the innate immune response (34). NLRC5, the largest member of the NLR
protein family, is widely expressed in numerous tissues (35). NLRC5 has been linked with major
histocompatibility complex class I gene expression (36). The involvement of NLRC5 in major
histocompatibility complex class I mediated CD8+ T cell
activation, proliferation and cytotoxicity is critical for host
defense against intracellular bacterial infections (37,38). The
biological function of NLRC5 in MI remains poorly understood. In
2017, Zhou et al (24)
reported that silencing of NLRC5 ameliorates cardiac fibrosis.
Consistent with these data, the present study showed that
downregulation of NLRC5 by miR-125b-5p mimic transfection decreased
apoptosis and intracellular oxidative stress, evidenced by
measurement of LDH and MDA levels as well as FACs analysis.
The expression of NLRC5 was downregulated by
treatment of HL-1 cells with MALAT1 siRNA and it was also observed
that its expression was upregulated by MALAT1 overexpression.
Together, these results indicated that MALAT1 modulated the
expression of NLRC5 in AMI. Previous studies demonstrated that
MALAT1 directly regulates the expression of miR-125b-5p (10,34).
Bioinformatics analysis as well as miR-125b-5p overexpression
experiments showed that NLRC5 is the target of miR-125b-5p. Indeed,
MALAT1 abrogated the miR-125b-5p-mediated cytoprotection in
H/R-induced apoptosis through regulation of NLRC5 expression
levels. Ma et al (39) found
that knockout of NLRC5 led to myocardial damage in mice on a high
fat diet. These different results may be due to differences in
experimental model, aim or conditions.
Taken together, to the best of our knowledge, the
results of the present study provide the first evidence for a
crosstalk between MALAT1, miR-125b and NLRC5 in AMI. It was
demonstrated that MALAT1 may function as a negative regulator of
miR-125b-5p to abrogate the protective effects of miR-125b-5p on
myocardial cells in AMI through modulating NLRC5 expression. The
MALAT1/NCLR5/miR-125b-5p axis could provide an effective
therapeutic target in AMI treatment.
Acknowledgements
Not applicable.
Funding
This work was supported by the Medical and Health
Science and Technology development project in Shandong province
(grant no. 2017WS766).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL and JL performed the experiments and wrote the
manuscript. YW, JX and ZW collected and analyzed the experimental
data. PW and HS analyzed the experimental data and revised the
manuscript. ZS and QL designed the experiments and approved the
final version manuscript.
Ethics approval and consent to
participate
The Dezhou People's Hospital Ethics Review Committee
for Animal Experimentation (experiment number 2017-TX-007) approved
this experiment.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AMI
|
acute myocardial infarction
|
|
MALAT1
|
metastasis associated lung
adenocarcinoma transcript 1
|
|
TTC
|
2,3,5-triphenyl-2H-tetrazolium
chloride
|
|
MDA
|
malondialdehyde
|
|
SD
|
Sprague Dawley
|
References
|
1
|
Thygesen K, Alpert JS, Jaffe AS, Chaitman
BR, Bax JJ, Morrow DA, et al: Fourth universal definition of
myocardial infarction (2018). Eur Heart J. 2018. View Article : Google Scholar
|
|
2
|
Nichols M, Townsend N, Scarborough P and
Rayner M: Cardiovascular disease in Europe 2014: Epidemiological
update. Eur Heart J. 35:2950–2959. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yeh RW, Sidney S, Chandra M, Sorel M,
Selby JV and Go AS: Population trends in the incidence and outcomes
of acute myocardial infarction. N Engl J Med. 362:2155–2165. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Writing Group Members, ; Mozaffarian D,
Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, Das SR, de
Ferranti S, Després JP, et al: Heart disease and stroke
statistics-2016 update: A report from the American Heart
Association. Circulation. 133:e38–e360. 2016.PubMed/NCBI
|
|
5
|
Talman V and Ruskoaho H: Cardiac fibrosis
in myocardial infarction-from repair and remodeling to
regeneration. Cell Tissue Res. 365:563–581. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ji P, Diederichs S, Wang W, Boing S,
Metzger R, Schneider PM, Tidow N, Brandt B, Buerger H, Bulk E, et
al: MALAT-1, a novel noncoding RNA, and thymosin beta4 predict
metastasis and survival in early-stage non-small cell lung cancer.
Oncogene. 22:8031–8041. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tripathi V, Ellis JD, Shen Z, Song DY, Pan
Q, Watt AT, Freier SM, Bennett CF, Sharma A, Bubulya PA, et al: The
nuclear-retained noncoding RNA MALAT1 regulates alternative
splicing by modulating SR splicing factor phosphorylation. Mol
Cell. 39:925–938. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang L, Lin C, Liu W, Zhang J, Ohgi KA,
Grinstein JD, Dorrestein PC and Rosenfeld MG: ncRNA- and Pc2
methylation-dependent gene relocation between nuclear structures
mediates gene activation programs. Cell. 147:773–788. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gutschner T, Hammerle M and Diederichs S:
MALAT1-a paradigm for long noncoding RNA function in cancer. J Mol
Med (Berl). 91:791–801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han Y, Liu Y, Zhang H, Wang T, Diao R,
Jiang Z, Gui Y and Cai Z: Hsa-miR-125b suppresses bladder cancer
development by down-regulating oncogene SIRT7 and oncogenic long
non-coding RNA MALAT1. FEBS Lett. 587:3875–3882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang X, Li M, Wang Z, Han S, Tang X, Ge Y,
Zhou L, Zhou C, Yuan Q and Yang M: Silencing of long noncoding RNA
MALAT1 by miR-101 and miR-217 inhibits proliferation, migration,
and invasion of esophageal squamous cell carcinoma cells. J Biol
Chem. 290:3925–3935. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Michalik KM, You X, Manavski Y,
Doddaballapur A, Zornig M, Braun T, John D, Ponomareva Y, Chen W,
Uchida S, et al: Long noncoding RNA MALAT1 regulates endothelial
cell function and vessel growth. Circ Res. 114:1389–1397. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Puthanveetil P, Chen S, Feng B, Gautam A
and Chakrabarti S: Long non-coding RNA MALAT1 regulates
hyperglycaemia induced inflammatory process in the endothelial
cells. J Cell Mol Med. 19:1418–1425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao ZH, Hao W, Meng QT, Du XB, Lei SQ and
Xia ZY: Long non-coding RNA MALAT1 functions as a mediator in
cardioprotective effects of fentanyl in myocardial
ischemia-reperfusion injury. Cell Biol Int. 41:62–70. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lupfer C and Kanneganti TD: Unsolved
mysteries in NLR biology. Front Immunol. 4:2852013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Downs I, Vijayan S, Sidiq T and Kobayashi
KS: CITA/NLRC5: A critical transcriptional regulator of MHC class I
gene expression. Biofactors. 42:349–357. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kuenzel S, Till A, Winkler M, Hasler R,
Lipinski S, Jung S, Grötzinger J, Fickenscher H, Schreiber S and
Rosenstiel P: The nucleotide-binding oligomerization domain-like
receptor NLRC5 is involved in IFN-dependent antiviral immune
responses. J Immunol. 184:1990–2000. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Neerincx A, Jakobshagen K, Utermöhlen O,
Buning H, Steimle V and Kufer TA: The N-terminal domain of NLRC5
confers transcriptional activity for MHC class I and II gene
expression. J Immunol. 193:3090–3100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yoshihama S, Roszik J, Downs I, Meissner
TB, Vijayan S, Chapuy B, Sidiq T, Shipp MA, Lizee GA and Kobayashi
KS: NLRC5/MHC class I transactivator is a target for immune evasion
in cancer. Proc Natl Acad Sci USA. 113:5999–6004. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chelbi ST and Guarda G: NLRC5, a promising
new entry in tumor immunology. J Immunother Cancer. 4:392016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bolha L, Ravnik-Glavač M and Glavač D:
Long noncoding RNAs as biomarkers in cancer. Dis Markers.
2017:72439682017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsang FH, Au V, Lu WJ, Shek FH, Liu AM,
Luk JM, Fan ST, Poon RT and Lee NP: Prognostic marker microRNA-125b
inhibits tumorigenic properties of hepatocellular carcinoma cells
via suppressing tumorigenic molecule eIF5A2. Dig Dis Sci.
59:2477–2487. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bayoumi AS, Park K-m, Wang Y, Teoh JP,
Aonuma T, Tang Y, Su H, Weintraub NL and Kim IM: A
carvedilol-responsive microRNA, miR-125b-5p protects the heart from
acute myocardial infarction by repressing pro-apoptotic bak1 and
klf13 in cardiomyocytes. J Mol Cell Cardiol. 114:72–82. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhou H, Yu X and Zhou G: NLRC5 silencing
ameliorates cardiac fibrosis by inhibiting the TGFbeta1/Smad3
signaling pathway. Mol Med Rep. 16:3551–3556. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pasha Z, Haider HKh and Ashraf M:
Efficient non-viral reprogramming of myoblasts to stemness with a
single small molecule to generate cardiac progenitor cells. PLoS
One. 6:e236672011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gutschner T, Hammerle M, Eissmann M, Hsu
J, Kim Y, Hung G, Revenko A, Arun G, Stentrup M, Gross M, et al:
The noncoding RNA MALAT1 is a critical regulator of the metastasis
phenotype of lung cancer cells. Cancer Res. 73:1180–1189. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang S, Yu W, Luo X, Chen J and Deng F:
MALAT1/miR-204/LC3-II: A potential regulated axis of autophagy in
myocardial ischemia-reperfusion injury. Int J Cardiol. 277:2222019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang S, Zhang L, Song J, Wang Z, Huang X,
Guo Z, Chen F and Zhao X: Long noncoding RNA MALAT1 mediates
cardiac fibrosis in experimental postinfarct myocardium mice model.
J Cell Physiol. 234:2997–3006. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang Q, Lu G and Chen Z: MALAT1 promoted
cell proliferation and migration via MALAT1/miR-155/MEF2A pathway
in hypoxia of cardiac stem cells. J Cell Biochem. 120:6384–6394.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao J, Li L and Peng L: MAPK1
up-regulates the expression of MALAT1 to promote the proliferation
of cardiomyocytes through PI3K/AKT signaling pathway. Int J Clin
Exp Pathol. 8:15947–15953. 2015.PubMed/NCBI
|
|
31
|
Hou J, Zhou C, Long H, Zheng S, Guo T, Wu
Q, Wu H, Zhong T and Wang T: Long noncoding RNAs: Novel molecules
in cardiovascular biology, disease and regeneration. Exp Mol
Pathol. 100:493–501. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Paraskevopoulou MD and Hatzigeorgiou AG:
Analyzing MiRNA-LncRNA Interactions. Methods Mol Biol.
1402:271–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chang SM and Hu WW: Long non-coding RNA
MALAT1 promotes oral squamous cell carcinoma development via
microRNA-125b/STAT3 axis. J Cell Physiol. 233:3384–3396. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Motta V, Soares F, Sun T and Philpott DJ:
NOD-like receptors: Versatile cytosolic sentinels. Physiol Rev.
95:149–178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Benko S, Magalhaes JG, Philpott DJ and
Girardin SE: NLRC5 limits the activation of inflammatory pathways.
J Immunol. 185:1681–1691. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kobayashi KS and van den Elsen PJ: NLRC5:
A key regulator of MHC class I-dependent immune responses. Nat Rev
Immunol. 12:813–820. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Biswas A, Meissner TB, Kawai T and
Kobayashi KS: Cutting edge: Impaired MHC class I expression in mice
deficient for Nlrc5/class I transactivator. J Immunol. 189:516–520.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yao Y, Wang Y, Chen F, Huang Y, Zhu S,
Leng Q, Wang H, Shi Y and Qian Y: NLRC5 regulates MHC class I
antigen presentation in host defense against intracellular
pathogens. Cell Res. 22:836–847. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ma SR and Xie XW: NLRC5 deficiency
promotes myocardial damage induced by high fat diet in mice through
activating TLR4/NF-κB. Biomed Pharmacother. 91:755–766. 2017.
View Article : Google Scholar : PubMed/NCBI
|